
The Role of Large Mammals as Vitamin C Sources for MIS 3 Hominins


Abstract
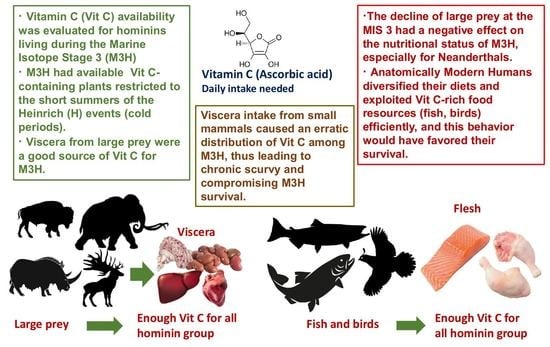
1. Introduction
1.1. Controversy in the Diet of M3H
1.2. Prevalence of Scurvy in M3H Populations
1.3. Vit C Status of Arctic Populations: Implications for MIS 3 Hominins
1.4. The Bottleneck of Vitamin C for the Survival of M3H
2. Material and Methods

{kind=link}
{kind=link}
| Species | mg/100 g Fresh wt | Method of Vitamin C Determination | Reference |
|---|---|---|---|
| Bone marrow | |||
| Caribou/Reindeer (Rangifer tarandus L.) | 0.00 | Microfluorometric | [86] |
| Brain | |||
| Musk ox (Ovibos moschatus Zimmermann) | 12.80 | Tritation with 2,6-dichlorophenolindophenol | [87] |
| Depot fat | |||
| Caribou/Reindeer (R. tarandus L.) | 1.80 | Spectrophotometric with 2,4-dinitrophenylhydrazine | [59] |
| Caribou/Reindeer (R. tarandus L.) | 0.00 | Microfluorometric | [86] |
| Hearth | |||
| Caribou/Reindeer (R. tarandus L.) | 2.60 | HPLC-ED | [88] |
| Muskox (O. moschatus Zimmermann) | 1.50 | Tritation with 2,6-dichlorophenolindophenol | [87] |
| Kidney | |||
| Caribou/Reindeer (R. tarandus L.) | 8.88 | HPLC-ED | [88] |
| Muskox (O. moschatus Zimmermann) | 5.90 | Tritation with 2,6-dichlorophenolindophenol | [87] |
| Liver | |||
| Caribou/Reindeer (R. tarandus L.) | 23.76 | HPLC-ED | [88] |
| Caribou/Reindeer (R. tarandus L.) | 11.88 | Microfluorometric | [86] |
| Mean for caribou/reindeer liver | 17.82 ± 8.40 | ||
| Muskox (O. moschatus Zimmermann) | 10.40 | Tritation with 2,6-dichlorophenolindophenol | [87] |
| Meat | |||
| Bear, polar (Ursus maritimus Phipps) | 1.00 | Spectrophotometric with 2,4-dinitrophenylhydrazine | [59] |
| Bear, polar (U. maritimus Phipps) | 2.00 | - | [89] FDC ID: 169794 |
| Mean for polar bear meat | 1.5 ± 0.7 | ||
| Bear, brown (Ursus arctos L.) | 0.00 | - | [89] FDC ID: 173845 |
| Beaver (Castor spp. L.) | 2.00 | - | [89] FDC ID: 175294 |
| Bison (Bison bison L.) | 0.00 | HPLC-FD | [90] |
| Bison (B. bison L.) | 0.00 | Tritation with 2,6-dichlorophenolindophenol | [91] |
| Bison (B. bison L.) | 0.00 | - | [89] FDC ID: 175293 |
| Mean for bison meat | 0.00 | ||
| Caribou/Reindeer (R. tarandus L.) | 0.86 | HPLC-ED | [88] |
| Caribou/Reindeer (R. tarandus L.) | 1.40 | Spectrophotometric with 2,4-dinitrophenylhydrazine | [59] |
| Caribou/Reindeer (R. tarandus L.) | 0.00 | Microfluorometric | [86] |
| Caribou/Reindeer (R. tarandus L.) | 0.00 | - | [89] FDC ID: 173853 |
| Mean for caribou meat | 0.56 ± 0.69 | ||
| Horse (Equus ferus caballus L.) | 0.00 | Tritation with 2,6-dichlorophenolindophenol | [92] |
| Horse (E. ferus caballus L.) | 1.00 | - | [89] FDC ID: 175086 |
| Mean for horse meat | 0.50 ± 0.71 | ||
| Moose (Alces alces L.) | 4.00 | - | [89] FDC ID: 174344 |
| Muskox (O. moschatus Zimmermann) | 1.50 | Spectrophotometric with 2,4-dinitrophenylhydrazine | [59] |
| Muskox (O. moschatus Zimmermann) | 0.80 | Tritation with 2,6-dichlorophenolindophenol | [87] |
| Mean for muskox meat | 1.15 ± 0.49 | ||
| Red deer (Cervus elaphus L.) | 0.00 | - | [89] FDC ID: 173855 |
| Wild boar (Sus scrofa L.) | 0.00 | - | [89] FDC ID: 175297 |
| Stomach | |||
| Caribou/Reindeer (Rangifer tarandus L.) | 0.45 | HPLC-ED | [88] |
| Organ/Animal | mg/100 g | Reference |
|---|---|---|
| Red bone marrow | ||
| Beef | 1.0–3.7 | [93] |
| Brain | ||
| Beef | 19.20 | [94] |
| Lamb/sheep/mutton | 19.40 | [94] |
| Pork | 13.50 | [94] |
| Mean for brain | 17.37 ± 3.35 | |
| Heart | ||
| Beef | 7.00 | [94] |
| Lamb/sheep/mutton | 7.30 | [94] |
| Pork | 0.60 | [95] |
| Pork | 2.54 | [96] |
| Pork | 5.00 | [94] |
| Mean for hearth | 4.49 ± 2.89 | |
| Kidney | ||
| Beef | 10.40 | [94] |
| Lamb/sheep/mutton | 12.90 | [94] |
| Pork | 2.52 | [95] |
| Pork | 14.20 | [94] |
| Mean for kidney | 10.01 ± 5.23 | |
| Liver | ||
| Beef | 22.40 | [94] |
| Beef | 21.80 | [97] |
| Beef | 25.00 | [98] |
| Beef | 32.00 | [99] |
| Lamb/sheep/mutton | 25.00 | [94] |
| Lamb/sheep/mutton | 46.00 | [99] |
| Pork | 8.35 | [95] |
| Pork | 14.66 | [96] |
| Pork | 21.60 | [94] |
| Pork | 12.00 | [99] |
| Mean for liver | 22.88 ± 10.68 | |
| Lung | ||
| Beef | 38.50 | [94] |
| Lamb/sheep/mutton | 31.40 | [94] |
| Lamb/sheep/mutton | 24.50 | [100] |
| Pork | 13.10 | [94] |
| Pork | 27.00 | [100] |
| Rabbit | 29.00 | [100] |
| Mean for lung | 27.25 ± 8.42 | |
| Meat | ||
| Beef | 0.00 | [101] |
| Beef | 2.53 | [102] |
| Beef | 0.00 | [103] |
| Beef | 1.60 | [98] |
| Buffalo | 0.00 | [104] |
| Camel | 0.00 | [103] |
| Lamb/sheep/mutton | 0.60 | [105] |
| Mutton | 0.00 | [103] |
| Pork | 0.27 | [95] |
| Pork | 0.90 | [96] |
| Mean for meat | 0.66 ± 0.89 | |
| Pancreas | ||
| Lamb/sheep/mutton | 17.50 | [94] |
| Pork | 15.30 | [94] |
| Mean for pancreas | 16.40 ± 1.56 | |
| Spleen | ||
| Beef | 45.50 | [94] |
| Lamb/sheep/mutton | 23.20 | [94] |
| Pork | 19.51 | [95] |
| Pork | 27.22 | [96] |
| Pork | 30.00 | [94] |
| Mean for spleen | 29.09 ± 10.00 | |
| Tonge | ||
| Beef | 3.30 | [94] |
| Lamb/sheep/mutton | 6.80 | [94] |
| Pork | 4.40 | [94] |
| Mean for tonge | 4.83 ± 1.79 | |
| Tripe/intestines | ||
| Beef | 3.40 | [94] |
| Lamb/sheep/mutton | 6.60 | [94] |
| Mean for tripe/intestines | 5.00 ± 2.26 | |
| Subcutaneous fat | ||
| Beef raw | 0.00 | [89] FDC ID: 173091 |
| Lamb raw | 0.00 | [89] FDC ID: 174435 |
| Pork | 0.00 | [95] |
| Pork | 0.00 | [89] FDC ID: 167813 |
| Mean for subcutaneous fat | 0.00 ± 0.00 | |
| Thymus | ||
| Beef | 34.00 | [94] |
| Body% | g | Energy kcal/g | Total kcal | Vit C mg/g | Total Vit C in Organ, mg | Reference for Vit C | Reference for Organ wt.% | Reference for Energy | |
|---|---|---|---|---|---|---|---|---|---|
| Viscera | |||||||||
| Adrenal | 0.0209 | 59.5 | 0.680 | 40 | 0.392 | 23.3 | [87] b | [106] c | Estimated as testes |
| Bile | 0.0522 | 148.7 | 0.045 | 7 | 0.01 | 1.5 | [87] b | [107] c | Derived from [108] d |
| Blood | 3.4052 | 9704.7 | 0.700 | 6793 | 0.0007 | 6.8 | [109] e | [110] e | [111] e |
| Brain | 0.2210 | 629.8 | 1.234 | 777 | 0.128 | 80.6 | [87] b | [106] f | [73] g |
| Digestive tract | 5.3000 | 15,105.0 | 0.960 | 14,501 | 0.074 | 1117.8 | [86] b | [106] e | [112] f |
| Epididymis | 0.1098 | 312.9 | 0.680 | 213 | 0.042 | 13.1 | [87] b | [113] i | Estimated as testes |
| Eyes | 0.0393 | 112.0 | 3.260 | 365 | 0.007 | 0.8 | [87] b | [106] c | [89] FDC ID: 169801f |
| Heart | 0.4400 | 1254.0 | 1.120 | 1404 | 0.015 | 18.8 | [87] b | [114] b | [112] f, [115] f |
| Kidney | 0.1855 | 528.7 | 0.917 | 485 | 0.059 | 31.2 | [87] b | [114] b | [73] g |
| Liver | 1.0382 | 2958.9 | 1.240 | 3669 | 0.104 | 307.7 | [87] b | [114] b | [115] f |
| Lung | 1.3528 | 3855.5 | 1.090 | 4202 | 0.081 | 312.3 | [87] b | [106] c | [112] f |
| Pancreas | 0.1405 | 400.4 | 2.350 | 941 | 0.039 | 15.6 | [87] b | [116] e | [89] FDC ID: 169452 h |
| Spleen | 0.2284 | 650.9 | 0.900 | 586 | 0.06 | 39.1 | [87] b | [106] c | [73] g |
| Testes | 0.5160 | 1470.6 | 0.680 | 1000 | 0.182 | 267.6 | [87] b | [107] c | [89] FDC ID: 172619 c |
| Tongue | 0.1891 | 538.9 | 2.60 | 1401 | 0.01 | 5.4 | [87] b | [107] c | [89] FDC ID: 168985 f |
| Thyroid | 0.0196 | 55.9 | 0.680 | 38 | 0.038 | 2.1 | [87] b | [107] c | Estimated as testes |
| Total viscera | 13.2584 | 37,786.4 | - | 36,423 | - | 2243.8 | - | - | - |
| Bones | 14.2748 | 40,683.2 | 0.000 | - | - | - | - | [114] b | - |
| Bone marrow | 2.5000 | 7125.0 | 4.837 | 34,464 | 0.000 | 0.000 | [86] f | [117] f | [73] g |
| Dissectible fat | 17.7099 | 50,473.2 | 7.894 | 398,436 | 0.000 | 0.000 | [87] b | [114] b | [73] b |
| Total fat | 57,715.5 | - | 432,899 | - | - | - | - | - | |
| Muscles | 41.9084 | 119,438.9 | 1.132 | 135,205 | 0.0115 | 1373.5 | [87] b; [59] b | [114] b | [73] b |
| Skin | 9.0043 | 25,662.3 | 5.440 | 139,603 | 0.0050 | 128.3 | [89] FDC ID: 234130 j | [110] e | [89] FDC ID: 2341308 j |
| Rest k | 3.6675 | 10,452.4 | 1.000 | 10,452 | 0.0010 | 10.5 | Estimated | Estimated | Estimated |
| Scientific Name | Organ | mg/100 g Fresh wt | Reference | |
|---|---|---|---|---|
| Birds | ||||
| Duck | Anas platyrhynchos | Flesh | 6.2 | [89] FDC ID: 174469 |
| Goose | Anser anser | Flesh | 7.2 | [89] FDC ID: 172413 |
| Pigeon | Columba livia | Flesh | 7.2 | [89] FDC ID: 171079 |
| Quail | Perdix perdix | Flesh | 7.2 | [89] FDC ID: 172419 |
| Seaweeds | ||||
| Dulse | Palmaria palmata | Thallus | 0.61 | [119] |
| Irishmoss | Chondrus crispus | Thallus | 3 | [89] FDC ID: 168456 |
| Kelp | Laminariales | Thallus | 3 | [89] FDC ID: 168457 |
| Kombu | Laminaria sp. | Thallus | 1.34 | [119] |
| Laver | Porphyra umbilicalis | Thallus | 39 | [89] FDC ID: 168458 |
| Laver | P. umbilicalis | Thallus | 33.29 | [119] |
| Sea spaghetti | Himanthalia elongata | Thallus | 46.66 | [119] |
| Wakame | Undaria pinnatifida | Thallus | 3 | [89] FDC ID: 170496 |
| Invertebrates | ||||
| Abalone | Haliotidae | Edible portion | 2 | [89] FDC ID: 174212 |
| Crab | Clibanarius vittatus | Whole body | 1.8 | [120] |
| Eastern Oysters | Crassostrea virginica | Edible portion | 3 | [121] |
| Eastern Oysters | Crassostrea virginica | Edible portion | 4.7 | [89] FDC ID: 175172 |
| Mussels | Mytilus edulis | Edible portion | 8 | [89] FDC ID: 174216 |
| Octopus, common | Octopus vulgaris | Edible portion | 5 | [89] FDC ID: 174218 |
| Polychaete | Neanthes virens | Whole body | 1.56 | [122] |
| Sea urchin | Paracentrotus lividus | Gonad | 26.57 | [123] |
| Squid, mixed species | Cephalopods | Edible portion | 4.7 | [89] FDC ID: 174223 |
| Shrimp | Palaemonetes pugio | Edible portion | 1.64 | [122] |
| Snail | Polinices duplicatus | Kidney | 2.34 | [120] |
| Whelk, unspecified | Mollusk | Edible portion | 4 | [89] FDC ID: 171983 |
| Freshwater fish | ||||
| Arctic char | Salvelinys alpinus | Whole fish | 5.8 | [59] |
| Arctic char | Salvelinys alpinus | Flesh | 1.8 | [59] |
| Arctic char | Salvelinusalpinus | Flesh | 1.3 | [88] |
| Bass, fresh water, mixed species | Perciformes | Flesh | 2 | [89] FDC ID: 174184 |
| Broad whitefish | Coregonos nasus | Flesh | 2.83 | [88] |
| Carp | Cyprinidae | Flesh | 1.6 | [89] FDC ID: 171952 |
| Carp | Cyprinus carpio | Flesh | 1.14 | [124] |
| Cisco | Coregonos nasus | Eggs | 49.6 | [87] |
| European catfish | Silurus glanis | Flesh | 2.15 | [124] |
| Pike, northern, raw | Esox lucius | Flesh | 3.8 | [89] FDC ID: 173680 |
| Pike perch | Sander lucioperca | Flesh | 1.91 | [124] |
| Whitefish | Coregonus clupeaformis | Eggs | 12 | [89] FDC ID: 167643 |
| Salmon, chinook | Oncorhynchus tshawytscha | Flesh | 4.0 | [89] FDC ID: 173688 |
| Sculpin | Myoxocephalus spp. | Flesh | 1.05 | [88] |
| Trout, rainbow | Oncorhynchus mykiss | Flesh | 2.4 | [89] FDC ID: 175154 |
| Trout, rainbow | Oncorhynchus mykiss | Flesh | 1.8 | [121] |
| Marine fish | ||||
| Mullet | Fam. Mugilidae | Flesh | 1.2 | [89] FDC ID: 175123 |
| Needlefish | Fam. Belonidae | Brain | 29.5 | [122] |
| Gill | 3.88 | [122] | ||
| Kidney | 2.93 | [122] | ||
| Flesh | 1.41 | [122] | ||
| Liver | 2.13 | [122] | ||
| Snapper, mixed species | - | Flesh | 1.6 | [89] FDC ID: 173698 |
| Marine mammals | ||||
| Beluga | Delphinapterus leucas | Mattak | 36.02 | [88] |
| Narwhal | Monodon monoceros | Mattak | 31.51 | [88] |
| Ringed seal | Phoca hispida | Brain | 14.86 | [88] |
| Liver | 23.8 | [88] | ||
| Liver | 35 | [59] | ||
| Meat | 1.55 | [88] | ||
| Meat | 3 | [59] |
3. Results and Discussion
3.1. Sources of Vitamin C for M3H
3.2. M3H Viscera, a Valuable Food Resource Hard to Equally Distribute among M3H
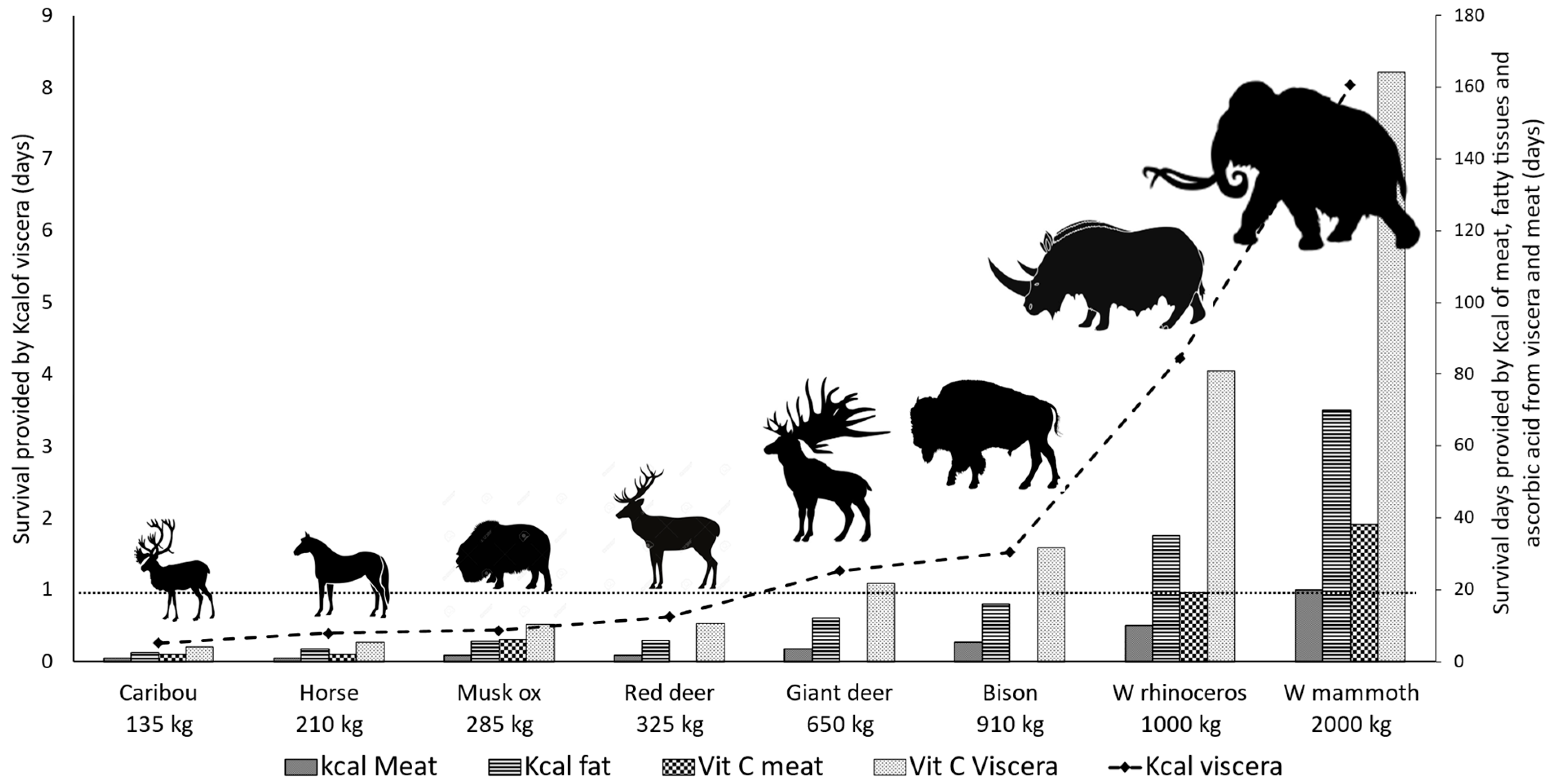
3.3. Paleolithic Mammals as Vitamin C Suppliers
3.4. The Importance of Large Prey for Paleolithic Hominins
3.5. The Decline of in Mammals during the Marine Isotope Stage 3: Differential Consequences for the Vit C Status of M3H
3.6. Vitamin C-Rich Food Resources Available to M3H
4. Conclusions
Supplementary Materials
Funding
Data Availability Statement
Conflicts of Interest
References
- Arsuaga, J.L.; Martínez, I.; Arnold, L.J.; Aranburu, A.; Gracia-Téllez, A.; Sharp, W.D.; Quam, R.M.; Falguères, C.; Pantoja-Pérez, A.; Bischoff, J.; et al. Neandertal roots: Cranial and chronological evidence from Sima de los Huesos. Science 2014, 344, 1358–1363. [Google Scholar] [CrossRef] [PubMed]
- Devièse, T.; Abrams, G.; Hajdinjak, M.; Pirson, S.; De Groote, I.; Di Modica, K.; Toussaint, M.; Fischer, V.; Comeskey, D.; Spindler, L.; et al. Reevaluating the timing of Neanderthal disappearance in Northwest Europe. Proc. Natl. Acad. Sci. USA 2021, 118, e2022466118. [Google Scholar] [CrossRef] [PubMed]
- Guil-Guerrero, J.L.; Manzano-Agugliaro, F. Worldwide research trends on Neanderthals. J. Quat. Sci. 2022, 38, 208–220. [Google Scholar] [CrossRef]
- Finlayson, C.; Carrión, J.S. Rapid ecological turnover and its impact on Neanderthal and other human populations. Trends Ecol. Evol. 2007, 22, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Staubwasser, M.; Drăgușin, V.; Onac, B.P.; Assonov, S.; Ersek, V.; Hoffmann, D.L.; Veres, D. Impact of climate change on the transition of Neanderthals to modern humans in Europe. Proc. Natl. Acad. Sci. USA 2018, 115, 9116–9121. [Google Scholar] [CrossRef]
- Banks, W.E.; d’Errico, F.; Peterson, A.T.; Kageyama, M.; Sima, A.; Sánchez-Goñi, M.F. Neanderthal extinction by competitive exclusion. PLoS ONE 2008, 3, e3972. [Google Scholar] [CrossRef]
- Houldcroft, C.; Underdown, S.J. Neanderthal genomics suggests a pleistocene time frame for the first epidemiologic transition. Am. J. Phys. Anthropol. 2016, 160, 379–388. [Google Scholar] [CrossRef]
- Smith, F.H.; Janković, I.; Karavanić, I. The assimilation model, modern human origins in Europe, and the extinction of Neandertals. Quat. Int. 2005, 137, 7–19. [Google Scholar] [CrossRef]
- Lalueza-Fox, C. Neanderthal assimilation? Nat. Ecol. Evol. 2021, 5, 711–712. [Google Scholar] [CrossRef]
- Degioanni, A.; Bonenfant, C.; Cabut, S.; Condemi, S. Living on the edge: Was demographic weakness the cause of Neanderthal demise? PLoS ONE 2019, 14, e0216742. [Google Scholar] [CrossRef]
- Fitzsimmons, K.E.; Hambach, U.; Veres, D.; Iovita, R. The Campanian Ignimbrite eruption: New data on volcanic ash dispersal and its potential impact on human evolution. PLoS ONE 2013, 8, e65839. [Google Scholar] [CrossRef] [PubMed]
- Timmermann, A. Quantifying the potential causes of Neanderthal extinction: Abrupt climate change versus competition and interbreeding. Quat. Sci. Rev. 2020, 238, 106331. [Google Scholar] [CrossRef]
- Fabre, V.; Condemi, S.; Degioanni, A. Genetic evidence of geographical groups among Neanderthals. PLoS ONE 2009, 4, e5151. [Google Scholar] [CrossRef] [PubMed]
- Haws, J.A.; Benedetti, M.M.; Talamo, S.; Bicho, N.; Cascalheira, J.; Ellis, M.G.; Carvalho, M.M.; Friedl, L.; Pereira, T.; Zinsious, B.K. The early Aurignacian dispersal of modern humans into westernmost Eurasia. Proc. Natl. Acad. Sci. USA 2020, 117, 25414–25422. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.R. Neanderthal extinction as part of the faunal change in Europe during Marine Isotope Stage 3. Acta Zool. Crac.-Ser. A Vertebr. 2007, 50, 93–124. [Google Scholar] [CrossRef]
- Bunn, H.T., III. Meat-Eating and Human Evolution: Studies on the Diet and Subsistence Patterns of Plio-Pleistocene Hominids in East Africa; University of California: Berkeley, CA, USA, 1982. [Google Scholar]
- Kolobova, K.A.; Roberts, R.G.; Chabai, V.P.; Jacobs, Z.; Krajcarz, M.T.; Shalagina, A.V.; Krivoshapkin, A.I.; Li, B.; Uthmeier, T.; Markin, S.V.; et al. Archaeological evidence for two separate dispersals of Neanderthals into southern Siberia. Proc. Natl. Acad. Sci. USA 2020, 117, 2879–2885. [Google Scholar] [CrossRef]
- Stuart, A.J. Late Quaternary megafaunal extinctions on the continents: A short review. Geol. J. 2015, 50, 338–363. [Google Scholar] [CrossRef]
- Richards, M.P.; Trinkaus, E. Out of Africa: Modern human origins special feature: Isotopic evidence for the diets of European Neanderthals and early modern humans. Proc. Natl. Acad. Sci. USA 2009, 106, 16034-9. [Google Scholar] [CrossRef]
- Wißing, C.; Rougier, H.; Crevecoeur, I.; Germonpré, M.; Naito, Y.I.; Semal, P.; Bocherens, H. Isotopic evidence for dietary ecology of late Neandertals in North-Western Europe. Quat. Int. 2016, 411, 327–345. [Google Scholar] [CrossRef]
- Ben-Dor, M.; Barkai, R. Supersize Does Matter: The Importance of Large Prey in Paleolithic Subsistence and a Method for Measurement of Its Significance in Zooarchaeological Assemblages, in Human–Elephant Interactions: From Past to Present; Konidaris, G., Barkai, R., Tourloukis, V., Eds.; Tübingen University Press: Tübingen, Germany, 2021. [Google Scholar]
- Ben-Dor, M.; Barkai, R. Prey Size Decline as a Unifying Ecological Selecting Agent in Pleistocene Human Evolution. Quaternary 2021, 4, 7. [Google Scholar] [CrossRef]
- Dodat, P.J.; Tacail, T.; Albalat, E.; Gómez-Olivencia, A.; Couture-Veschambre, C.; Holliday, T.; Madelaine, S.; Martin, J.E.; Rmoutilova, R.; Maureille, B.; et al. Isotopic calcium biogeochemistry of MIS 5 fossil vertebrate bones: Application to the study of the dietary reconstruction of Regourdou 1 Neandertal fossil. J. Hum. Evol. 2021, 151, 102925. [Google Scholar] [CrossRef] [PubMed]
- Balter, V.; Person, A.; Labourdette, N.; Drucker, D.; Renard, M.; Vandermeersch, B. Les Néandertaliens étaient-ils essentiellement carnivores? Résultats préliminaires sur les teneurs en Sr et en Ba de la paléobiocénose mammalienne de Saint-Césaire. Comptes Rendus L’acad. Sci.-Ser. IIA-Earth Planet. Sci. 2001, 332, 59–65. [Google Scholar] [CrossRef]
- Kuzmin, Y.V.; Bondarev, A.A.; Kosintsev, P.A.; Zazovskaya, E.P. The Paleolithic diet of Siberia and Eastern Europe: Evidence based on stable isotopes (δ13C and δ15N) in hominin and animal bone collagen. Archaeol. Anthropol. Sci. 2021, 13, 179. [Google Scholar] [CrossRef]
- Drucker, D.G.; Naito, Y.I.; Péan, S.; Prat, S.; Crépin, L.; Chikaraishi, Y.; Ohkouchi, N.; Puaud, S.; Lázničková-Galetová, M.; Patou-Mathis, M.; et al. Isotopic analyses suggest mammoth and plant in the diet of the oldest anatomically modern humans from far southeast Europe. Sci. Rep. 2017, 7, 6833. [Google Scholar] [CrossRef]
- Wißing, C.; Rougier, H.; Baumann, C.; Comeyne, A.; Crevecoeur, I.; Drucker, D.G.; Gaudzinski-Windheuser, S.; Germonpré, M.; Gómez-Olivencia, A.; Krause, J.; et al. Stable isotopes reveal patterns of diet and mobility in the last Neandertals and first modern humans in Europe. Sci. Rep. 2019, 9, 4433. [Google Scholar] [CrossRef]
- Miras, Y.; Barbier-Pain, D.; Ejarque, A.; Allain, E.; Allué, E.; Marín, J.; Vettese, D.; Hardy, B.; Puaud, S.; Llach, J.M.; et al. Neanderthal plant use and stone tool function investigated through non-pollen palynomorphs analyses and pollen washes in the Abri du Maras, South-East France. J. Archaeol. Sci. Rep. 2020, 33, 102569. [Google Scholar] [CrossRef]
- Hardy, K.; Bocherens, H.; Miller, J.B.; Copeland, L. Reconstructing Neanderthal diet: The case for carbohydrates. J. Hum. Evol. 2022, 162, 103105. [Google Scholar] [CrossRef]
- Henry, A.G.; Brooks, A.S.; Piperno, D.R. Plant foods and the dietary ecology of Neanderthals and early modern humans. J. Hum. Evol. 2014, 69, 44–54. [Google Scholar] [CrossRef]
- Churchill, S.E. Thin on the Ground: Neandertal Biology, Archeology, and Ecology; John Wiley & Sons: Hoboken, NJ, USA, 2014. [Google Scholar]
- Buck, L.T.; Stringer, C.B. Having the stomach for it: A contribution to Neanderthal diets? Quat. Sci. Rev. 2014, 96, 161–167. [Google Scholar] [CrossRef]
- Fiorenza, L.; Benazzi, S.; Tausch, J.; Kullmer, O.; Bromage, T.G.; Schrenk, F. Molar macrowear reveals Neanderthal eco-geographic dietary variation. PLoS ONE 2011, 6, e14769. [Google Scholar] [CrossRef]
- Droke, J.L.; Schmidt, C.W.; Williams, F.L.; Karriger, W.M.; Smith, F.H.; Becam, G.; de Lumley, M.-A. Regional variability in diet between Northern European and Mediterranean Neandertals: Evidence from dental microwear texture analysis. In Dental Wear in Evolutionary and Biocultural Contexts; Academic Press: Cambridge, MA, USA, 2020; pp. 225–241. [Google Scholar] [CrossRef]
- Ben-Dor, M.; Gopher, A.; Barkai, R. Neandertals’ large lower thorax may represent adaptation to high protein diet. Am. J. Phys. Anthropol. 2016, 160, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Hardy, B.L.; Moncel, M.-H. Neanderthal use of fish, mammals, birds, starchy plants and wood 125–250,000 years ago. PLoS ONE 2011, 6, e23768. [Google Scholar] [CrossRef] [PubMed]
- Fiore, I.; Gala, M.; Romandini, M.; Cocca, E.; Tagliacozzo, A.; Peresani, M. From feathers to food: Reconstructing the complete exploitation of avifaunal resources by Neanderthals at Fumane cave, unit A9. Quat. Int. 2016, 421, 134–153. [Google Scholar] [CrossRef]
- Finlayson, S.; Finlayson, G.; Guzman, F.J.G.; Finlayson, C. Neanderthals and the cult of the Sun Bird. Quat. Sci. Rev. 2019, 217, 217–224. [Google Scholar] [CrossRef]
- Morin, E.; Meier, J.; El Guennouni, K.; Moigne, A.-M.; Lebreton, L.; Rusch, L.; Valensi, P.; Conolly, J.; Cochard, D. New evidence of broader diets for archaic Homo populations in the northwestern Mediterranean. Sci. Adv. 2019, 5, eaav9106. [Google Scholar] [CrossRef]
- Guillaud, E.; Béarez, P.; Daujeard, C.; Defleur, A.R.; Desclaux, E.; Roselló-Izquierdo, E.; Morales-Muñiz, A.; Moncel, M.-H. Neanderthal foraging in freshwater ecosystems: A reappraisal of the Middle Paleolithic archaeological fish record from continental Western Europe. Quat. Sci. Rev. 2021, 252, 106731. [Google Scholar] [CrossRef]
- Tanner, A. An embarrassment of riches: The ontological aspect of meat and fat harvesting among subarctic hunters. In Human–Elephant Interactions: From Past to Present; Konidaris, G., Barkai, R., Tourloukis, V., Eds.; Tübingen University Press: Tübingen, Germany, 2021; Available online: https://publikationen.uni-tuebingen.de/xmlui/handle/10900/114229 (accessed on 19 November 2022).
- Ben-Dor, M.; Gopher, A.; Hershkovitz, I.; Barkai, R. Man the fat hunter: The demise of Homo erectus and the emergence of a new hominin lineage in the Middle Pleistocene (ca. 400 kyr) Levant. PLoS ONE 2011, 6, e28689. [Google Scholar] [CrossRef]
- Navazo Ruiz, M.; Benito-Calvo, A.; Alonso-Alcalde, R.; Alonso, P.; de la Fuente, H.; Santamaría, M.; Santamaría, C.; Álvarez-Vena, A.; Arnold, L.J.; Iriarte-Chiapusso, M.J.; et al. Late Neanderthal subsistence strategies and cultural traditions in the northern Iberia Peninsula: Insights from Prado Vargas, Burgos, Spain. Quat. Sci. Rev. 2021, 254, 106795. [Google Scholar] [CrossRef]
- McLeod, H. Plenty of Fish in the Sea? A Comparison of Marine Resource Use in Early Hominins. Ethnograph 2018, 4, 36–55. [Google Scholar]
- Bertacchi, A.; Starkovich, B.M.; Conard, N.J. The Zooarchaeology of Sirgenstein Cave: A Middle and Upper Paleolithic site in the Swabian Jura, SW Germany. J. Paleolit. Archaeol. 2021, 4, 7. [Google Scholar] [CrossRef]
- Englard, S.; Seifter, S. The biochemical functions of ascorbic acid. Annu. Rev. Nutr. 1986, 6, 365–406. [Google Scholar] [CrossRef]
- Padh, H. Cellular functions of ascorbic acid. Biochem. Cell Biol. 1990, 68, 1166–1173. [Google Scholar] [CrossRef]
- Baker, E.M.; Hodges, R.E.; Hood, J.; Sauberlich, H.E.; March, S.C.; Canham, J.E. Metabolism of 14C and 3H-labeled L-ascorbic acid in human scurvy. Am. J. Clin. Nutr. 1971, 24, 444–454. [Google Scholar] [CrossRef]
- Burri, B.J.; Jacob, R.A. Human metabolism and the requirement for vitamin C. In Vitamin C in Health and Disease; Packer, L., Fuchs, J., Eds.; Marcel Dekker Inc.: New York, NY, USA, 1997; pp. 341–366. [Google Scholar]
- Olmedo, J.M.; Yiannias, J.A.; Windgassen, E.B.; Gornet, M.K. Scurvy: A disease almost forgotten. Int. J. Dermatol. 2006, 45, 909–913. [Google Scholar] [CrossRef] [PubMed]
- Huss-Ashmore, R.; Goodman, A.H.; Armelagos, G.J. Nutritional inference from paleopathology. In Advances in Archaeological Method and Theory; Academic Press: Cambridge, MA, USA, 1982; pp. 395–474. Available online: https://www.jstor.org/stable/20210060 (accessed on 12 January 2023).
- Ortner, D.J.; Ericksen, M.F. Bone changes in the human skull probably resulting from scurvy in infancy and childhood. Int. J. Osteoarchaeol. 1997, 7, 212–220. [Google Scholar] [CrossRef]
- Eddie, D.M. Examination of Trauma in a Neandertal Ulna. Master’s Thesis, University of Kansas, Lawrence, KS, USA, 2013. Available online: https://kuscholarworks.ku.edu/handle/1808/12956 (accessed on 27 October 2022).
- Mednikova, M.B. Bioarcheology of Neanderthal burials from the territory of France and Crimea. Brief Commun. Inst. Archeol. 2015, 238, 243–261. [Google Scholar]
- Fennell, K.J.; Trinkaus, E. Bilateral femoral and tibial periostitis in the La Ferrassie 1 Neanderthal. J. Archaeol. Sci. 1997, 24, 985–995. [Google Scholar] [CrossRef]
- Gardner, J.C.; Smith, F.H. The paleopathology of the Krapina Neandertals. Period. Biol. 2006, 108, 471. [Google Scholar]
- Teschler-Nicola, M.; Czerny, C.; Oliva, M.; Schamall, D.; Schultz, M. Pathological alterations and traumas in the human skeletal remains from Mladeč. In Early Modern Humans at the Moravian Gate; Springer: Vienna, Austria, 2006; pp. 473–489. [Google Scholar]
- Garralda, M.D.; Vandermeersch, B. ¿Escorbuto en los neandertales? Posibles casos en Combe-Grenal (Domme, Francia). Veleia 2008, 24–25, 385–395. [Google Scholar]
- Geraci, J.R.; Smith, T.G. Ascorbic acid in the diet of Inuit hunters from Holman, Northwest Territories. Arctic 1979, 32, 135–139. [Google Scholar] [CrossRef]
- Rodahl, K. Nutritional requirements in cold climates. J. Nutr. 1954, 53, 575–588. [Google Scholar] [CrossRef] [PubMed]
- Fediuk, K. Vitamin C in the Inuit Diet: Past and Present. School of Dietetics & Human Nutrition. Master’s Thesis, McGill University, Montreal, QC, Canada, 2000. [Google Scholar]
- Delanghe, J.R.; Langlois, M.R.; De Buyzere, M.L.; Na, N.; Ouyang, J.; Speeckaert, M.M.; Torck, M.A. Vitamin C deficiency: More than just a nutritional disorder. Genes Nutr. 2011, 6, 341–346. [Google Scholar] [CrossRef]
- Clemens, Z.; Tóth, C. Vitamin C and disease: Insights from the evolutionary perspective. J. Evol. Health Jt. Publ. Ancestral Health Soc. Soc. Evol. Med. Health 2013, 1, 13. [Google Scholar] [CrossRef]
- Høygaard, A.; Rasmussen, H.W. Vitamin C sources in eskimo food. Nature 1939, 3631, 943. [Google Scholar] [CrossRef]
- Mullie, P.; Deliens, T.; Clarys, P. Vitamin C in East-Greenland traditional nutrition: A reanalysis of the Høygaard nutritional data (1936–1937). Int. J. Circumpolar Health 2021, 80, 1951471. [Google Scholar] [CrossRef]
- Levine, V.E. The value of meat as an antiscorbutic. Am. J. Dig. Dis. 1941, 8, 454–463. [Google Scholar] [CrossRef]
- Stefansson, V. The Dilemma in Vitamins. Science 1939, 89, 484. [Google Scholar] [CrossRef]
- Pierson, E.M. The Antiscorbutic Properties of Some Common Food Materials. Ph.D. Thesis, The University of Minnesota, Minneapolis, MN, USA, 1922. [Google Scholar]
- Stringer, C.B.; Finlayson, J.C.; Barton, R.N.E.; Fernández-Jalvo, Y.; Cáceres, I.; Sabin, R.C.; Rhodes, E.J.; Currant, A.P.; Rodríguez-Vidal, J.; Giles-Pacheco, F.; et al. Neanderthal exploitation of marine mammals in Gibraltar. Proc. Natl. Acad. Sci. USA 2008, 105, 14319–14324. [Google Scholar] [CrossRef]
- Dembitzer, J.; Barkai, R.; Ben-Dor, M.; Meiri, S. Levantine overkill: 1.5 million years of hunting down the body size distribution. Quat. Sci. Rev. 2022, 276, 107316. [Google Scholar] [CrossRef]
- Ben-Dor, M.; Barkai, R. The importance of large prey animals during the Pleistocene and the implications of their extinction on the use of dietary ethnographic analogies. J. Anthropol. Archaeol. 2020, 59, 101192. [Google Scholar] [CrossRef]
- Ben-Dor, M.; Sirtoli, R.; Barkai, R. The evolution of the human trophic level during the Pleistocene. Am. J. Phys. Anthropol. 2021, 175, 27–56. [Google Scholar] [CrossRef] [PubMed]
- Guil-Guerrero, J.L. Evidence for chronic omega-3 fatty acids and ascorbic acid deficiency in Palaeolithic hominins in Europe at the emergence of cannibalism. Quat. Sci. Rev. 2017, 157, 176–187. [Google Scholar] [CrossRef]
- Goldfield, A.E.; Booton, R.; Marston, J.M. Modeling the role of fire and cooking in the competitive exclusion of Neanderthals. J. Hum. Evol. 2018, 124, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Venner, S.J. A New Estimate for Neanderthal Energy Expenditure; CUNY Hunter College: New York, NY, USA, 2018. [Google Scholar]
- Paddon-Jones, D.; Rasmussen, B. Dietary protein recommendations and the prevention of sarcopenia: Protein, amino acid metabolism and therapy. Curr. Opin. Clin. Nutr. Metab. Care 2009, 12, 86–90. [Google Scholar] [CrossRef]
- Bilsborough, S.; Mann, N. A review of issues of dietary protein intake in humans. Int. J. Sport. Nutr. Exerc. Metab. 2006, 16, 129. [Google Scholar] [CrossRef]
- Shaffer, P.A. Antiketogenesis. II. The ketogenic antiketogenic balance in man. J. Biol. Chem. 1921, 47, 463–473. [Google Scholar]
- Hassan, F.A. Demographic archaeology. In Advances in Archaeological Method and Theory; Academic Press: Cambridge, MA, USA, 1981; pp. 225–279. [Google Scholar]
- Kelly, R.L. The Lifeways of Hunter-Gatherers: The Foraging Spectrum, 2nd ed.; Cambridge University Press: Cambridge, CA, USA, 2013. [Google Scholar]
- Goren-Inbar, N.; Belfer-Cohen, A. Reappraisal of hominin group size in the Lower Paleolithic: An introduction to the special issue. J. Hum. Evol. 2020, 144, 102821. [Google Scholar] [CrossRef]
- Smith, E.A. The application of optimal foraging theory to the analysis of hunter-gatherer group size. Hunt.-Gatherer Strateg. 1981, 137, 36–65. [Google Scholar]
- Trinkaus, E. Neanderthal mortality patterns. J. Archaeol. Sci. 1995, 22, 121–142. [Google Scholar] [CrossRef]
- Trinkaus, E. Late Pleistocene adult mortality patterns and modern human establishment. Proc. Natl. Acad. Sci. USA 2011, 108, 1267–1271. [Google Scholar] [CrossRef]
- Dietary Guidelines for Americans. United States Department of Agriculture and United States Department of Health and Human Services, 7th ed.; U.S. Government Printing Office: Washington, DC, USA, 2010. Available online: http://www.dietaryguidelines.gov (accessed on 25 December 2022).
- Hassan, A.A.; Sandanger, T.M.; Brustad, M. Level of selected nutrients in meat, liver, tallow and bone marrow from semi-domesticated reindeer (Rangifer t. tarandus L.). Int. J. Circumpolar Health 2012, 71, 17997. [Google Scholar] [CrossRef] [PubMed]
- Rodahl, K. Vitamin sources in Arctic regions. Nor. Polarinst. Skr. 1949, 91, 1–47. [Google Scholar]
- Fediuk, K.; Hidiroglou, N.; Madère, R.; Kuhnlein, H.V. Ascorbic acid in Inuit traditional food and women’s diets. J. Food Compos. Anal. 2002, 15, 221–235. [Google Scholar] [CrossRef]
- USDA National Nutrient Database for Standard Reference Legacy Release. Available online: https://ndb.nal.usda.gov (accessed on 25 June 2022).
- Driskell, J.A.; Marchello, M.J.; Giraud, D.W.; Sulaeman, A. Vitamin and selenium content of ribeye cuts from grass-and grain-finished bison of the same herd. J. Food Qual. 2004, 27, 388–398. [Google Scholar] [CrossRef]
- Marchello, M.J.; Driskell, J.A. Nutrient composition of grass-and grain-finished bison. Great Plains Res. 2001, 11, 65–82. [Google Scholar] [CrossRef]
- Lee, C.-E.; Seong, P.-N.; Oh, W.-Y.; Ko, M.-S.; Kim, K.-I.; Jeong, J.-H. Nutritional characteristics of horsemeat in comparison with those of beef and pork. Nutr. Res. Pract. 2007, 1, 70–73. [Google Scholar] [CrossRef] [PubMed]
- Serum, D.K.; Field, R.A.; Miller, G.J. Contribution of bone marrow to the vitamin content of mechanically separated meat. Meat Sci. 1986, 17, 73–77. [Google Scholar] [CrossRef]
- Kizlaitis, L.; Steinfeld, M.I.; Siedler, A.J. Nutrient Content of Variety Meats. I. Vitamin A, Vitamin C, Iron, and Proximate Composition a, b. J. Food Sci. 1962, 27, 459–462. [Google Scholar] [CrossRef]
- Eichenberger, B.; Pfirter, H.P.; Wenk, C.; Gebert, S. Influence of dietary Vitamin E and C supplementation on Vitamin E and C content and thiobarbituric acid reactive substances (TBARS) in different tissues of growing pigs. Arch. Anim. Nutr. 2004, 58, 195–208. [Google Scholar] [CrossRef]
- Gebert, S.; Eichenberger, B.; Pfirter, H.P.; Wenk, C. Influence of different dietary vitamin C levels on vitamin E and C content and oxidative stability in various tissues and stored m. longissimus dorsi of growing pigs. Meat Sci. 2006, 73, 362–367. [Google Scholar] [CrossRef]
- Mori, K.; Kidawara, M.; Iseki, M.; Umegaki, C.; Kishi, T. A simple fluorometric determination of vitamin C. Chem. Pharm. Bull. 1998, 46, 1474–1476. [Google Scholar] [CrossRef] [PubMed]
- Phillips, P.H.; Stare, U.F. The distribution of a reducing substance (vitamin C) in the tissues of fluorine-fed cows. J. Biol. Chem. 1934, 104, 351–358. [Google Scholar] [CrossRef]
- Svirbely, J.L.; Szent-Györgyi, A. The chemical nature of Vitamin C. Biochem. J. 1933, 27, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Slade, R.; Stead, A.G.; Graham, J.A.; Hatch, G.E. Comparison of lung antioxidant levels in humans and laboratory animals. Am. Rev. Respir. Dis. 1985, 131, 742–746. [Google Scholar]
- Chant, J.L.; Day, L.; Field, R.A.; Kruggel, W.G.; Chang, Y. Composition and palatability of mechanically deboned meat and mechanically separated tissue. J. Food Sci. 1977, 42, 306–309. [Google Scholar] [CrossRef]
- Descalzo, A.M.; Rossetti, L.; Grigioni, G.; Irurueta, M.; Sancho, A.M.; Carrete, J.; Pensel, N.A. Antioxidant status and odour profile in fresh beef from pasture or grain-fed cattle. Meat Sci. 2007, 75, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, H.H.H.; Jin, G.; Ma, M.; Khalifa, I.; Shukat, R.; Elkhedir, A.E.; Zeng, Q.; Noman, A.E. Comparative characterization of proximate nutritional compositions, microbial quality and safety of camel meat in relation to mutton, beef, and chicken. LWT 2020, 118, 108714. [Google Scholar] [CrossRef]
- Tamburrano, A.; Tavazzi, B.; Callà, C.A.M.; Amorini, A.M.; Lazzarino, G.; Vincenti, S.; Zottola, T.; Campagna, M.C.; Moscato, U.; Laurenti, P. Biochemical and nutritional characteristics of buffalo meat and potential implications on human health for a personalized nutrition. Ital. J. Food Saf. 2019, 8, 8317. [Google Scholar] [CrossRef]
- Young, O.A.; Lane, G.A.; Podmore, C.; Fraser, K.; Agnew, M.; Cummings, T.L.; Cox, N.R. Changes in composition and quality characteristics of ovine meat and fat from castrates and rams aged to 2 years. N. Z. J. Agric. Res. 2006, 49, 419–430. [Google Scholar] [CrossRef]
- Crile, G.; Quiring, D.P. A record of the body weight and certain organ and gland weights of 3690 animals. Ohio J. Sci. 1940, 15, 219–259. [Google Scholar]
- Kirton, A.H.; Fourie, P.D.; Jury, K.E. Growth and development of sheep: III. Growth of the carcass and non-carcass components of the Southdown and Romney and their cross and some relationships with composition. N. Z. J. Agric. Res. 1972, 15, 214–227. [Google Scholar] [CrossRef]
- Barrett, K.E. Ganong’s Review of Medical Physiology, 24th ed.; McGraw-Hill Medical: New York, NY, USA, 2012. [Google Scholar]
- Richmond, M.S.; Satterfield, G.H.; Grinnells, C.D.; Dann, W.J. Ascorbic acid Content of Goat’s Milk and Blood: Influence of Ascorbic Acid Injection and Diet: Three Figures. J. Nutr. 1940, 20, 99–108. [Google Scholar] [CrossRef]
- Awuk, A.; Tamir, B. Feed Intake, Weight Gain and Carcass Yield Characteristics of Intact Hararghe Highland Male Goats Fed on Different Hay to Concentrate Ratios. East Afr. J. Sci. 2007, 1, 45–54. [Google Scholar] [CrossRef]
- Nollet, L.M.; Toldrá, F. (Eds.) Handbook of Analysis of Edible Animal by-Products; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Kuhnleini, H.V.; Soueida, R. Use and nutrient composition of traditional Baffin Inuit foods. J. Food Compos. Anal. 1992, 5, 112–126. [Google Scholar] [CrossRef]
- Bercovitch, F.B.; Rodriguez, J.F. Testis size, epididymis weight, and sperm competition in rhesus macaques. Am. J. Primatol. 1993, 30, 163–168. [Google Scholar] [CrossRef]
- Adamczewski, J.Z.; Flood, P.F.; Gunn, A. Seasonal Patterns in Body Composition and Reproduction of Female Muskoxen (Ovibos moschatus). J. Zool. 1997, 241, 245–269. [Google Scholar] [CrossRef]
- Kuhnlein, H.; Chan, H.; Leggee, D.; Barthet, V. Macronutrient, mineral and fatty acid composition of Canadian Arctic traditional food. J. Food Compos. Anal. 2002, 15, 545–566. [Google Scholar] [CrossRef]
- Sebsibe, A.; Casey, N.; Van Niekerk, W.; Tegegne, A.; Coertze, R. Growth performance and carcass characteristics of three Ethiopian goat breeds fed grainless diets varying in concentrate to roughage ratios. S. Afr. J. Anim. Sci. 2007, 37, 221–232. [Google Scholar] [CrossRef]
- Nieminen, M.; Laitinen, M. Bone marrow and kidney fat as indicators of condition in reindeer. Rangifer 1986, 6, 219–226. [Google Scholar] [CrossRef]
- Knott, K.K.; Barboza, P.S.; Bowyer, R.T. Growth in arctic ungulates: Postnatal development and organ maturation in Rangifer tarandus and Ovibos moschatus. J. Mammal. 2005, 86, 121–130. [Google Scholar] [CrossRef]
- Ferraces-Casais, P.; Lage-Yusty, M.A.; De Quirós, A.R.-B.; López-Hernández, J. Evaluation of bioactive compounds in fresh edible seaweeds. Food Anal. Methods 2012, 5, 828–834. [Google Scholar] [CrossRef]
- Carr, R.S.; Neff, J.M. Determination of ascorbic acid in tissues of marine animals by liquid chromatography with electrochemical detection. Anal. Chem. 1980, 52, 2428–2430. [Google Scholar] [CrossRef] [PubMed]
- Nettleton, J.A.; Exler, J. Nutrients in wild and farmed fish and shellfish. J. Food Sci. 1992, 57, 257–260. [Google Scholar] [CrossRef]
- Carr, R.S.; Bally, M.B.; Thomas, P.; Neff, J.M. Comparison of methods for determination of ascorbic acid in animal tissues. Anal. Chem. 1983, 55, 1229–1232. [Google Scholar] [CrossRef]
- de Quirós, A.R.-B.; Lopez-Hernandez, J.; Simal-Lozano, J. Determination of ascorbic acid in sea urchin: Comparison of two HPLC methods. Chromatographia 2001, 53, S246–S249. [Google Scholar] [CrossRef]
- Özyurt, G.; Polat, A.; Loker, G.B. Vitamin and mineral content of pike perch (Sander lucioperca), common carp (Cyprinus carpio), and European catfish (Silurus glanis). Turk. J. Vet. Anim. Sci. 2009, 33, 351–356. [Google Scholar] [CrossRef]
- Daujeard, C.; Abrams, G.; Germonpré, M.; Le Pape, J.-M.; Wampach, A.; Di Modica, K.; Moncel, M.-H. Neanderthal and animal karstic occupations from southern Belgium and south-eastern France: Regional or common features? Quat. Int. 2016, 411, 179–197. [Google Scholar] [CrossRef]
- Lebreton, L.; Moigne, A.-M.; Filoux, A.; Perrenoud, C. A specific small game exploitation for Lower Paleolithic: The beaver (Castor fiber) exploitation at the Caune de l’Arago (Pyrénées-Orientales, France). J. Archaeol. Sci. Rep. 2017, 11, 53–58. [Google Scholar] [CrossRef]
- Collie, J.T.; Greaves, R.F.; Jones, O.A.; Eastwood, G.; Bellomo, R. Ascorbic acid measurement in critical illness: Challenges, methodologies and quality improvements. Clin. Chem. Lab. Med. (CCLM) 2020, 58, 460–470. [Google Scholar] [CrossRef]
- Carr, A.C.; Lykkesfeldt, J. Discrepancies in global vitamin C recommendations: A review of RDA criteria and underlying health perspectives. Crit. Rev. Food Sci. Nutr. 2021, 61, 742–755. [Google Scholar] [CrossRef]
- Hodges, R.E.; Hood, J.; Canham, J.E.; Sauberlich, H.E.; Baker, E.M. Clinical manifestations of ascorbic acid deficiency in man. Am. J. Clin. Nutr. 1971, 24, 432–443. [Google Scholar] [CrossRef] [PubMed]
- Masci, D.; Rubino, C.; Basile, M.; Indolfi, G.; Trapani, S. When the limp has a dietary cause: A retrospective study on scurvy in a tertiary Italian pediatric hospital. Front. Pediatr. 2022, 10, 1581. [Google Scholar] [CrossRef] [PubMed]
- Henry, A.G.; van Amerongen, Y. Tubers in Winter. Seventh Annual Meeting of the European Society for the Study of Human Evolution. Evol. Anthropol. Issues News Rev. 2017, 27, 87. [Google Scholar]
- Schwarcz, H.P. Winter sources of ascorbic acid for Pleistocene hominins in northern Eurasia. Res. Sq. 2023, preprint. [Google Scholar] [CrossRef]
- Ng, W.Y.; Hung, L.Y.; Lam, Y.H.; Chan, S.S.; Pang, K.S.; Chong, Y.K.; Ching, C.K.; Mak, T.W. Poisoning by toxic plants in Hong Kong: A 15-year review. Hong Kong Med. J. 2019, 25, 102. [Google Scholar] [CrossRef]
- Applequist, W.L.; Bridges, M.C.; Moerman, D.E. North American Fertility–Regulating Botanicals: A Review. Econ. Bot. 2022, 76, 84–113. [Google Scholar] [CrossRef]
- Ali, M.; Hossain, M.; Akhter, S.; Islam; Hashem, M. Effect of age on slaughterhouse by-products of indigenous cattle of Bangladesh. Bangladesh J. Anim. Sci. 2013, 42, 62–66. [Google Scholar] [CrossRef]
- Bunn, H.T.; Ezzo, J.A. Hunting and scavenging by Plio-Pleistocene hominids: Nutritional constraints, archaeological patterns, and behavioural implications. J. Archaeol. Sci. 1993, 20, 365–398. [Google Scholar] [CrossRef]
- Devatkal, S.; Mendiratta, S. Evaluation of shelf life and spoilage pattern of aerobically stored buffalo liver. J. Muscle Foods 2007, 18, 276–284. [Google Scholar] [CrossRef]
- Tennie, C.; O’Malley, R.C.; Gilby, I.C. Why do chimpanzees hunt? Considering the benefits and costs of acquiring and consuming vertebrate versus invertebrate prey. J. Hum. Evol. 2014, 71, 38–45. [Google Scholar] [CrossRef]
- McHugh, T. The Time of the Buffalo; University of Nebraska Press: Lincoln, NE, USA, 1979; p. 85. [Google Scholar]
- Saario, D.J. Human Ecological Investigations at Kivalina, Alaska. Final Report. No. PNE-425; University of Alaska and Oak Ridge: Fairbanks, AK, USA, 1962. [Google Scholar]
- Blumenschine, R.J.; Caro, T.M. Unit flesh weights of some East African bovids. Afr. J. Ecol. 1986, 24, 273–286. [Google Scholar] [CrossRef]
- Nilssen, P.J. An actualistic Butchery Study in South Africa and Its Implications for Reconstructing Hominid Strategies of Carcass Acquisition and Butchery in the Upper Pleistocene and Plio-Pleistocene. Ph.D. Thesis, University of Cape Town, Cape Town, South Africa, 2000. [Google Scholar]
- Blumenschine, R.J. Carcass consumption sequences and the archaeological distinction of scavenging and hunting. J. Hum. Evol. 1986, 15, 639–659. [Google Scholar] [CrossRef]
- Morin, E.; Ready, E. Foraging Goals and Transport Decisions in Western Europe During the Paleolithic and Early Holocene. In Zooarchaeology and Modern Human Origins: Human Hunting Behavior During the Later Pleistocene; Clark, J., Speth, J., Eds.; Springer: New York, NY, USA, 2013; pp. 227–269. [Google Scholar]
- Blasco, R.; Rosell, J.; Arilla, M.; Margalida, A.; Villalba, D.; Gopher, A.; Barkai, R. Bone marrow storage and delayed consumption at Middle Pleistocene Qesem Cave, Israel (420 to 200 ka). Sci. Adv. 2019, 5, eaav9822. [Google Scholar] [CrossRef]
- Wu, G. Dietary protein intake and human health. Food Funct. 2016, 7, 1251–1265. [Google Scholar] [CrossRef]
- Rendu, W.; Costamagno, S.; Meignen, L.; Soulier, M.-C. Monospecific faunal spectra in Mousterian contexts: Implications for social behavior. Quat. Int. 2012, 247, 50–58. [Google Scholar] [CrossRef]
- Driver, J.C.; Maxwell, D. Bison death assemblages and the interpretation of human hunting behaviour. Quat. Int. 2013, 297, 100–109. [Google Scholar] [CrossRef]
- Dusseldorp, G.L. Studying prehistoric hunting proficiency: Applying optimal foraging theory to the Middle Palaeolithic and Middle Stone Age. Quat. Int. 2012, 252, 3–15. [Google Scholar] [CrossRef]
- Rodríguez-Hidalgo, A.; Saladié, P.; Ollé, A.; Arsuaga, J.L.; de Castro, J.M.B.; Carbonell, E. Human predatory behavior and the social implications of communal hunting based on evidence from the TD10. 2 bison bone bed at Gran Dolina (Atapuerca, Spain). J. Hum. Evol. 2017, 105, 89–122. [Google Scholar] [CrossRef] [PubMed]
- Saladié, P.; Huguet, R.; Díez, C.; Rodríguez-Hidalgo, A.; Cáceres, I.; Vallverdú, J.; Rosell, J.; de Castro, J.M.B.; Carbonell, E. Carcass transport decisions in Homo antecessor subsistence strategies. J. Hum. Evol. 2011, 61, 425–446. [Google Scholar] [CrossRef]
- O’Connell, J.F.; Hawkes, K.; Jones, N.B. Reanalysis of large mammal body part transport among the Hadza. J. Archaeol. Sci. 1990, 17, 301–316. [Google Scholar] [CrossRef]
- Monchot, H.; Houmard, C.; Dionne, M.M.; Desrosiers, P.M.; Gendron, D. The modus operandi of walrus exploitation during the Palaeoeskimo period at the Tayara site, Arctic Canada. Anthropozoologica 2013, 48, 15–36. [Google Scholar] [CrossRef]
- Rabassa, J.; Ponce, J.F. The Heinrich and Dansgaard-Oeschger climatic events during Marine Isotopic Stage 3: Searching for appropriate times for human colonization of the Americas. Quat. Int. 2013, 299, 94–105. [Google Scholar] [CrossRef]
- Clarke, A.; Gaston, K.J. Climate, energy and diversity. Proc. R. Soc. B Biol. Sci. 2006, 273, 2257–2266. [Google Scholar] [CrossRef] [PubMed]
- Nogués-Bravo, D.; Rodríguez, J.; Hortal, J.; Batra, P.; Araújo, M.B. Climate change, humans, and the extinction of the woolly mammoth. PLoS Biol. 2008, 6, e79. [Google Scholar] [CrossRef]
- D’Errico, F.; Zilhão, J.; Julien, M.; Baffier, D.; Pelegrin, J. Neanderthal acculturation in Western Europe? A critical review of the evidence and its interpretation. Curr. Anthropol. 1998, 39, S1–S44. [Google Scholar] [CrossRef]
- Eeuwes, D.D.M. Megafauna Extinction Caused by Homo Sapiens? Reports Environmental Science No. 589. Master’s Thesis, Radboud University Nijmegen, Nijmegen, The Netherlands, 2018. [Google Scholar]
- Magniez, P.; Boulbes, N. Environment during the Middle to Late Palaeolithic transition in southern France: The archaeological sequence of Tournal Cave (Bize-Minervois, France). Quat. Int. 2014, 337, 43–63. [Google Scholar] [CrossRef]
- Morin, E. Evidence for declines in human population densities during the early Upper Paleolithic in western Europe. Proc. Natl. Acad. Sci. USA 2008, 105, 48–53. [Google Scholar] [CrossRef]
- El Zaatari, S.; Grine, F.E.; Ungar, P.S.; Hublin, J.-J. Neandertal versus modern human dietary responses to climatic fluctuations. PLoS ONE 2016, 11, e0153277. [Google Scholar] [CrossRef]
- Fabre, V.; Condemi, S.; Degioanni, A.; Herrscher, E. Neanderthals versus modern humans: Evidence for resource competition from isotopic modelling. Int. J. Evol. Biol. 2011, 2011, 689315. [Google Scholar] [CrossRef]
- Sepulchre, P.; Ramstein, G.; Kageyama, M.; Vanhaeren, M.; Krinner, G.; Sánchez-Goñi, M.F.; d’Errico, F. H4 abrupt event and late Neanderthal presence in Iberia. Earth Planet. Sci. Lett. 2007, 258, 283–292. [Google Scholar] [CrossRef]
- Defleur, A.R.; Desclaux, E. Impact of the last interglacial climate change on ecosystems and Neanderthals behavior at Baume Moula-Guercy, Ardèche, France. J. Archaeol. Sci. 2019, 104, 114–124. [Google Scholar] [CrossRef]
- Drucker, D.; Bocherens, H. Carbon and nitrogen stable isotopes as tracers of change in diet breadth during Middle and Upper Palaeolithic in Europe. Int. J. Osteoarchaeol. 2004, 14, 162–177. [Google Scholar] [CrossRef]
- Richards, M.P.; Pettitt, P.B.; Stiner, M.C.; Trinkaus, E. Stable isotope evidence for increasing dietary breadth in the European mid-Upper Paleolithic. Proc. Natl. Acad. Sci. USA 2001, 98, 6528–6532. [Google Scholar] [CrossRef] [PubMed]
- Torke, W. Fischreste als Quellen der Okologie und Okonomie in der Steinzeit Siidwest-Deutschlands; Urgeschichtliche Materialhefte Nr. 4; Verlag Archaeologica Venatoria; Institut für Urgeschichte der Universität: Tübingen, Germany, 1981. [Google Scholar]
- Conard, N.J.; Kitagawa, K.; Krönneck, P.; Böhme, M.; Münzel, S.C. The importance of fish, fowl and small mammals in the Paleolithic diet of the Swabian Jura, southwestern Germany. In Zooarchaeology and Modern Human Origins; Clark, J., Speth, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 173–190. [Google Scholar] [CrossRef]
- Van Neer, W.; Wouters, W.; Germonpré, M. Fish remains from three Upper Palaeolithic cave deposits in southern Belgium. Anthropol. Praehist. 2007, 118, 5–22. [Google Scholar]
- Tortosa, J.; Pardo, J.F.J.; Álvarez-Fernández, E.; Ripoll, M.P.; Aristu, B.A.; Morales-Pérez, J.V.; García, M.J.R.; Marlasca, R.; Alcover, J.A.; Jardón, P.; et al. Palaeolithic-epipalaeolithic Sea People of the southern Iberian coast (Spain): An overview. In Archaeology of Maritime Hunter-Gatherers. From Settlement Function to the Organization of the Coastal Zone; Pôle editorial: Paris, France, 2016; pp. 69–92. [Google Scholar]
- Álvarez-Fernández, E. Humans and Marine Resource Interaction Reappraised: Archaeofauna Remains During the Late Pleistocene and Holocene in Cantabrian Spain. J. Anthropol. Archaeol. 2011, 30, 327–343. [Google Scholar] [CrossRef]
- The Royal Canadian Geographical Society. Indigenous Peoples Atlas of Canada; Canadian Geographic: Ottawa, ON, USA, 2018. [Google Scholar]
- Stiner, M.C. Small game use and expanding diet breadth. Petits animaux et sociétés humaines du complément alimentaire aux ressources utilitaires. In Actes des XXIVe Rencontres Internationales D’archéologie et D’histoire d’Antibes; APDCA: Antibes, France, 2004. [Google Scholar]
- Nishiaki, Y.; Jöris, O. Learning Behaviors Among Neanderthals and Palaeolithic Modern Humans: An Introduction. In Learning Among Neanderthals and Palaeolithic Modern Humans; Springer: Singapore, 2019; pp. 1–6. [Google Scholar]
- Zwir, I.; Del-Val, C.; Hintsanen, M.; Cloninger, K.M.; Romero-Zaliz, R.; Mesa, A.; Arnedo, J.; Salas, R.; Poblete, G.F.; Raitoharju, E.; et al. Evolution of genetic networks for human creativity. Mol. Psychiatry 2021, 27, 354–376. [Google Scholar] [CrossRef]
- Germonpré, M.; Fedorov, S.; Danilov, P.; Galeta, P.; Jimenez, E.-L.; Sablin, M.; Losey, R.J. Palaeolithic and prehistoric dogs and Pleistocene wolves from Yakutia: Identification of isolated skulls. J. Archaeol. Sci. 2017, 78, 1–19. [Google Scholar] [CrossRef]
- Lahtinen, M.; Clinnick, D.; Mannermaa, K.; Salonen, J.S.; Viranta, S. Excess protein enabled dog domestication during severe Ice Age winters. Sci. Rep. 2021, 11, 7. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guil-Guerrero, J.L. The Role of Large Mammals as Vitamin C Sources for MIS 3 Hominins. Quaternary 2023, 6, 20. https://doi.org/10.3390/quat6010020
Guil-Guerrero JL. The Role of Large Mammals as Vitamin C Sources for MIS 3 Hominins. Quaternary. 2023; 6(1):20. https://doi.org/10.3390/quat6010020
Chicago/Turabian StyleGuil-Guerrero, José Luis. 2023. "The Role of Large Mammals as Vitamin C Sources for MIS 3 Hominins" Quaternary 6, no. 1: 20. https://doi.org/10.3390/quat6010020
APA StyleGuil-Guerrero, J. L. (2023). The Role of Large Mammals as Vitamin C Sources for MIS 3 Hominins. Quaternary, 6(1), 20. https://doi.org/10.3390/quat6010020