
A Glacial–Interglacial Malacofauna Record from the Titel Loess Plateau, Serbia, between ~350 and 250 ka
 , , , ,
, , , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
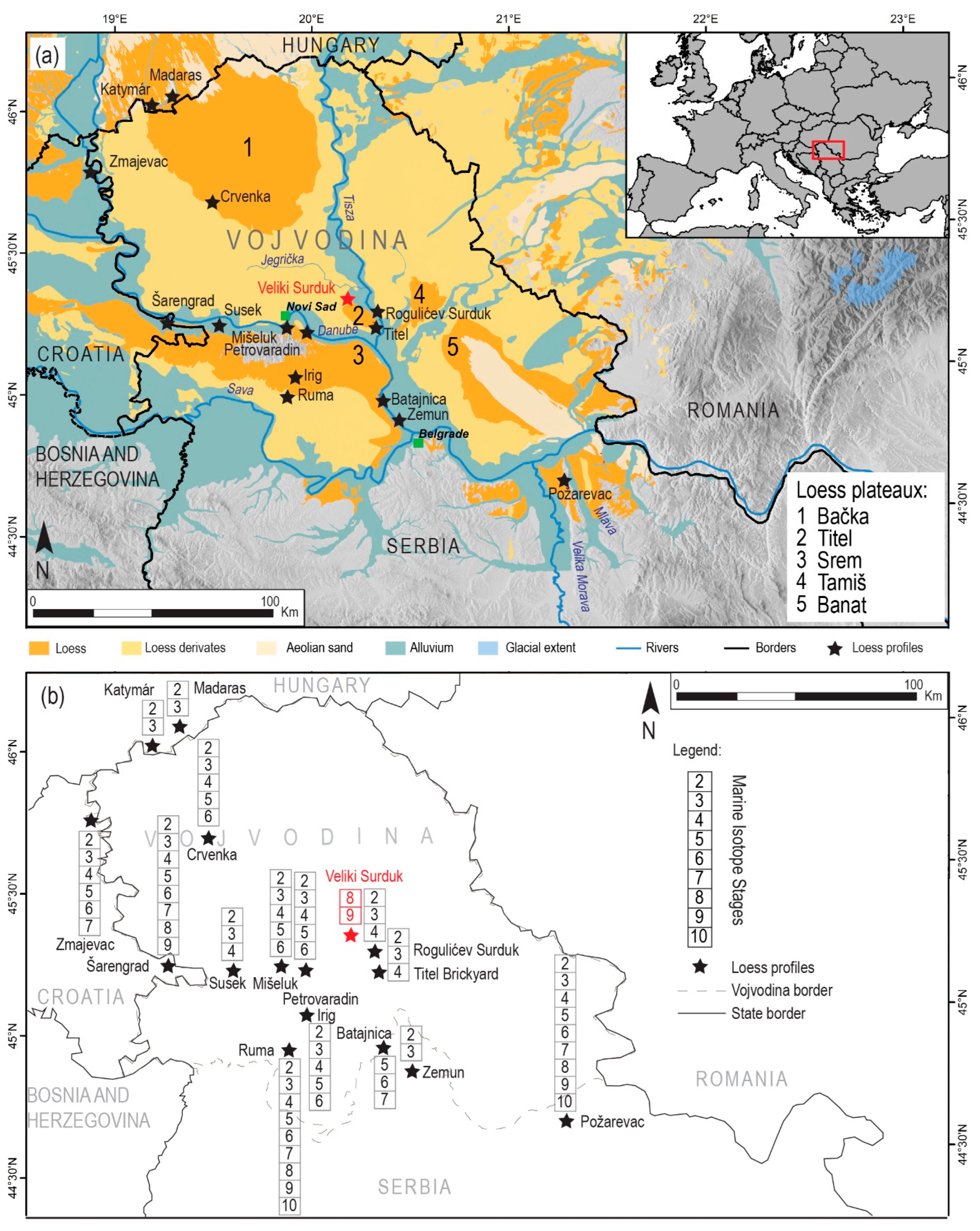
2.1. Description of the Study Site
2.2. Sampling Strategy
2.3. Malacothermometer July Paleotemperatures
2.4. Time-Scale Construction and Grain Size Measurement
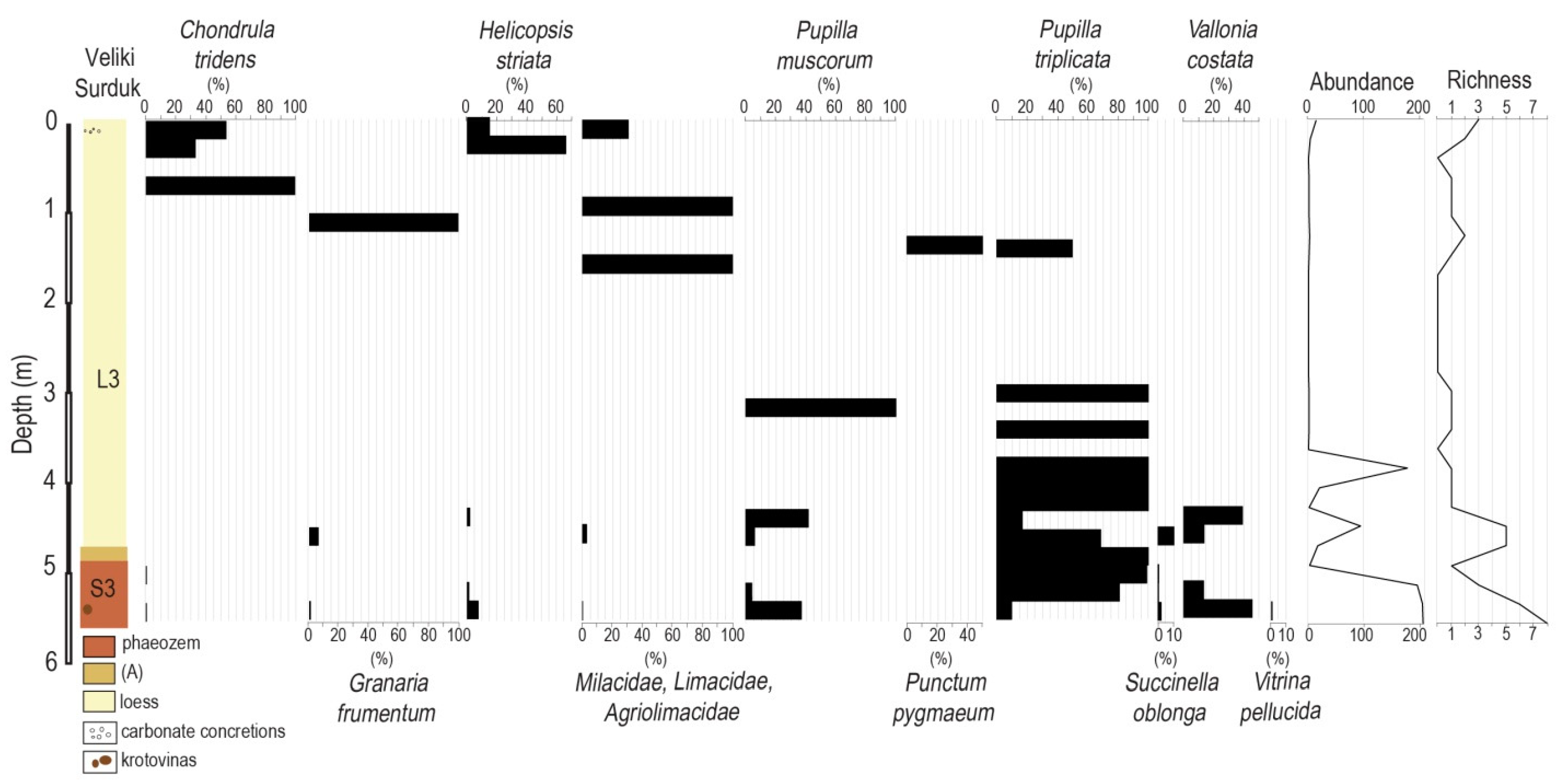
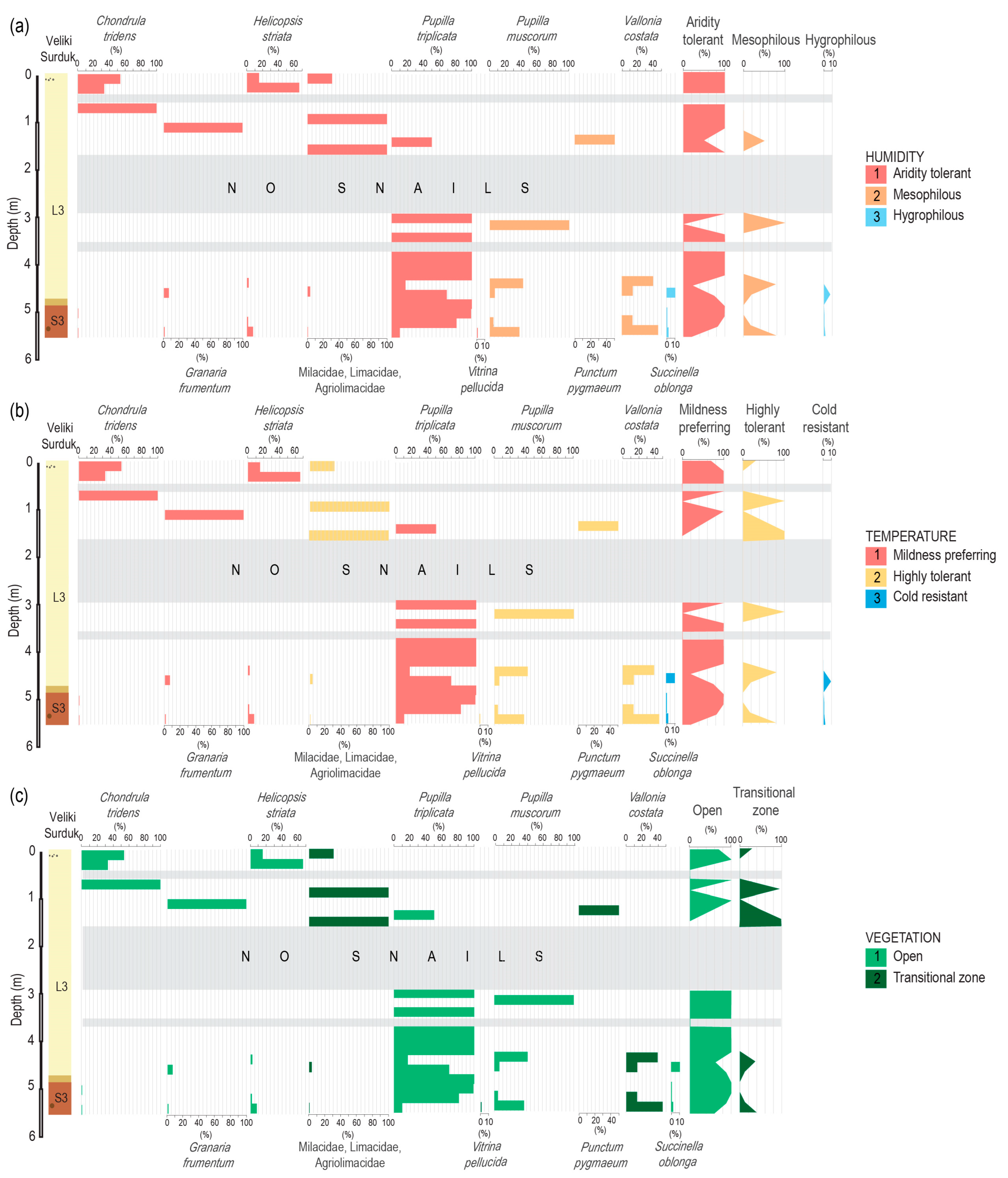
3. Results
Malacology of the Loess Section
4. Discussion
4.1. Chronostratigraphic Context
4.2. Comparison to Other Malacological Records
4.3. Orbital Forcing
4.4. Connection to Regional Records
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ložek, V. Quartärmollusken der Tschechoslowakei; Herausgegeben von der Geologischen Zentralanstaltim Verlag der Tschechoslowakischen Akademie der Wissenschaften: Praha, Czech Republic, 1964; Volume 31. (In German) [Google Scholar]
- Ložek, V. Molluscs in loess, their paleoecological significance and role in geochronology—Principles and methods. Quatern. Int. 1990, 7, 71–79. [Google Scholar] [CrossRef]
- Ložek, V. Molluscan fauna from the loess series of Bohemia and Moravia. Quat. Int. 2001, 76–77, 141–156. [Google Scholar] [CrossRef]
- Sümegi, P.; Gulyás, S.; Csökmei, B.; Molnár, D.; Hambach, U.; Stevens, T.; Markovic, S.B.; Almond, P.C. Climatic fluctuations inferred for the Middle and Late Pleniglacial (MIS 2) based on high-resolution (~ca. 20 y) preliminary environmental magnetic investigation of the loess section of the Madaras brickyard (Hungary). Central Eur. Geol. 2012, 55, 329–345. [Google Scholar] [CrossRef]
- Újvári, G.; Molnár, M.; Novothny, Á.; Páll-Gergely, B.; Kovács, J.; Várhegyi, A. AMS 14C and OSL/IRSL dating of the Dunaszekcső loess sequence (Hungary): Chronology for 20 to 150 ka and implications for establishing reliable age–depth models for the last 40 ka. Quat. Sci. Rev. 2014, 106, 140–154. [Google Scholar] [CrossRef]
- Sümegi, P.; Marković, S.B.; Molnár, D.; Sávai, S.; Náfrádi, K.; Szelepcsényi, Z.; Novák, Z. Črvenka loess-paleosol sequence revisited: Local and regional Quaternary biogeographical inferences of the southern Carpathian Basin. Open Geosci. 2016, 8, 390–404. [Google Scholar] [CrossRef]
- Zhai, J.; Wang, X.; Qin, B.; Cui, L.; Zhang, S.; Ding, Z. Clumped isotopes in land snail shells over China: Towards establishing a biogenic carbonate paleothermometer. Geochim. Cosmochim. Acta 2019, 257, 68–79. [Google Scholar] [CrossRef]
- Kukla, J.; Ložek, V.; Záruba, Q. Zur Stratigraphie der Lösse in der Tschechoslowakei. Quartär-Int. Jahrbuchzur Erforsch. Des Eiszeitalt. Und Der Steinzeit 1961, 13, 1–29. [Google Scholar]
- Alexandrowicz, S.W. Malacological analyses in Quaternary research. Kwart. AGH Geol. 1987, 13, 1–240. [Google Scholar]
- Alexandrowicz, S.W. Late holocene molluscan assemblages from Czorsztyn (Pieniny Klippen Belt, southern Poland). Folia Malacol. 2016, 5, 15–24. [Google Scholar] [CrossRef]
- Puisségur, J.J. Mollusques Continentaux Quaternaires de Bourgogne: Significations Stratigraphiques et Climatiques, Rapports avec d’autres Faunes Boréales de France; Mémoires géologiques de l’Université de Dijon: Dijon, France, 1976; Volume 3, pp. 1–241. [Google Scholar]
- Magnin, F. Mollusques Continentaux et Histoire Quaternaire des Milieux Méditerranéens (Sud-Est de la France, Catalogne). Ph.D. Dissertation, Aix-Marseille Université, Marseille, France, 1991. [Google Scholar]
- Kerney, M.P. British Quaternary non-marine Mollusca: A brief review. In British Quaternary Studies—Recent Advances; Shotton, F.W., Ed.; Clarendon Press: Oxford, UK, 1978; pp. 31–42. [Google Scholar]
- Preece, R.C. The molluscan fauna of the middle pleistocene interglacial deposits at Little Oakley, Essex, and its environmental and stratigraphical implications. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1990, 328, 387–407. [Google Scholar] [CrossRef]
- Soós, L. The past of the Hungarian mollusk fauna. Ann. Museihistorico-Nat. Hung. 1926, 24, 392–421. [Google Scholar]
- Krolopp, E. Middle Pleistocene mollusc fauna from the Vértesszőlős campsite of prehistoric man. Földrajzi Közlemények 1978, 35, 188–211. [Google Scholar]
- Oches, E.A.; McCoy, W.D. Historical developments and recent advances in amino acid geochronology applied to loess research: Examples from North America, Europe, and China. Earth-Sci. Rev. 2001, 54, 173–192. [Google Scholar] [CrossRef]
- Rousseau, D.-D. Loess biostratigraphy: New advances and approaches in mollusk studies. Earth-Sci. Rev. 2001, 54, 157–171. [Google Scholar] [CrossRef]
- Moine, O.; Rousseau, D.D.; Antoine, P. Terrestrial molluscan records of Weichselian Lower to Middle Pleniglacial climatic changes from the Nussloch loess series (Rhine Valley, Germany): The impact of local factors. Boreas 2005, 34, 363–380. [Google Scholar] [CrossRef]
- Moine, O.; Rousseau, D.-D.; Antoine, P. The impact of Dansgaard–Oeschger cycles on the loessic environment and malacofauna of Nussloch (Germany) during the Upper Weichselian. Quat. Res. 2008, 70, 91–104. [Google Scholar] [CrossRef]
- Sümegi, P.; Krolopp, E. Quatermalacological analyses for modeling of the Upper Weichselianpalaeoenvironmental changes in the Carpathian Basin. Quatern. Int. 2002, 91, 53–63. [Google Scholar] [CrossRef]
- Sümegi, P.; Molnár, D.; Náfrádi, K.; Makó, L.; Cseh, P.; Törőcsik, T.; Molnár, M.; Zhou, L. Vegetation and land snail-based reconstruction of the palaeocological changes in the forest steppe eco-region of the Carpathian Basin during last glacial warming. Glob. Ecol. Conserv. 2022, 33, e01976. [Google Scholar] [CrossRef]
- Czébely, A.; Túri, M.; Kiss, D.; Újvári, G.; Kertész, T.; Rinyu, L. Clumped Isotope Temperatures from Secondary Carbonates in Loess: Comparability of Different Preparation Methods of Snail Shells and Earthworm Biospheroids (No. EGU24-16279). Copernicus Meetings. 2024. Available online: https://meetingorganizer.copernicus.org/EGU24/EGU24-16279.html (accessed on 25 April 2024).
- Rousseau, D.-D. A tribute to Ložek (1965): The problem of loess formation and the loess molluscs. E&G Quat. Sci. J. 2021, 70, 229–233. [Google Scholar] [CrossRef]
- Marković, S.B.; Bokhorst, M.P.; Vandenberghe, J.; McCoy, W.D.; Oches, E.A.; Hambach, U.; Gaudenyi, T.; Jovanović, M.; Zöller, L.; Stevens, T. Late Pleistocene loess-palaeosol sequences in the Vojvodina region, north Serbia. J. Quat. Sci. 2008, 23, 73–84. [Google Scholar] [CrossRef]
- Marković, S.B.; Stevens, T.; Kukla, G.J.; Hambach, U.; Fitzsimmons, K.E.; Gibbard, P.; Buggle, B.; Zech, M.; Guo, Z.; Hao, Q.; et al. Danube loess stratigraphy—Towards a pan-European loess stratigraphic model. Earth-Sci. Rev. 2015, 148, 228–258. [Google Scholar] [CrossRef]
- Fitzsimmons, K.E.; Marković, S.B.; Hambach, U. Pleistocene environmental dynamics recorded in the loess of the middle and lower Danube basin. Quat. Sci. Rev. 2012, 41, 104–118. [Google Scholar] [CrossRef]
- Perić, Z.; Adolphi, E.L.; Stevens, T.; Újvári, G.; Zeeden, C.; Buylaert, J.-P.; Marković, S.B.; Hambach, U.; Fischer, P.; Schmidt, C.; et al. Quartz OSL dating of late quaternary Chinese and Serbian loess: A cross Eurasian comparison of dust mass accumulation rates. Quat. Int. 2019, 502, 30–44. [Google Scholar] [CrossRef]
- Fenn, K.; Millar, I.L.; Durcan, J.A.; Thomas, D.S.; Banak, A.; Marković, S.B.; Veres, D.; Stevens, T. The provenance of Danubian loess. Earth-Sci. Rev. 2022, 226, 103920. [Google Scholar] [CrossRef]
- Lukić, T.; Radaković, M.; Marković, R.; Thompson, W.; Ponjiger, T.M.; Basarin, B.; Tomić, N.; Tomović, V.; Raljić, J.P.; Gavrilov, M.; et al. Initial results of the colorimetric indices of the oldest exposed pedocomplex (Titel loess plateau, Serbia). Geol. Croat. 2023, 76, 73–85. [Google Scholar] [CrossRef]
- Marković, S.B.; Vandenberghe, J.; Perić, Z.M.; Filyó, D.; Bartyik, T.; Radaković, M.G.; Hao, Q.; Marković, R.S.; Lukić, T.; Tomić, N.; et al. Local Differentiation in the Loess Deposition as a Function of Dust Source: Key Study Novo Orahovo Loess Paleosol Sequence (Vojvodina, Serbia). Quaternary 2023, 6, 23. [Google Scholar] [CrossRef]
- Song, Y.; Guo, Z.; Marković, S.; Hambach, U.; Deng, C.; Chang, L.; Wu, J.; Hao, Q. Magnetic stratigraphy of the Danube loess: A composite Titel-Stari Slankamen loess section over the last one million years in Vojvodina, Serbia. J. Asian Earth Sci. 2018, 155, 68–80. [Google Scholar] [CrossRef]
- Marković, S.B.; Hambach, U.; Stevens, T.; Basarin, B.; O’Hara-Dhand, K.; Gavrilov, M.M.; Gavrilov, M.B.; Smalley, I.; Teofanov, N. Relating the astronomical timescale to the loess–paleosol sequences in Vojvodina, Northern Serbia. In Climate Change; Springer: Vienna, Austria, 2012; pp. 65–78. [Google Scholar]
- Basarin, B.; Buggle, B.; Hambach, U.; Marković, S.B.; Dhand, K.O.; Kovačević, A.; Stevens, T.; Guo, Z.; Lukić, T. Time-scale and astronomical forcing of Serbian loess–paleosol sequences. Glob. Planet. Chang. 2014, 122, 89–106. [Google Scholar] [CrossRef]
- Zeeden, C.; Hark, M.; Hambach, U.; Markovic, S.; Zöller, L. Depressions on the Titel loess plateau: Form, pattern, genesis. Geogr. Pannonica 2007, 11, 4–8. [Google Scholar] [CrossRef]
- Sümegi, P.; Molnár, D.; Gulyás, S.; Náfrádi, K.; Sümegi, B.P.; Törőcsik, T.; Persaits, G.; Molnár, M.; Vandenberghe, J.; Zhou, L. High-resolution proxy record of the environmental response to climatic variations during transition MIS3/MIS2 and MIS2 in Central Europe: The loess-paleosol sequence of Katymár brickyard (Hungary). Quatern. Int. 2019, 504, 40–55. [Google Scholar] [CrossRef]
- Sümegi, P.; Gulyás, S.; Molnár, D.; Bozsó, G.; Fekete, I.; Makó, L.; Cseh, P.; Molnár, M.; Sümegi, B.P.; Almond, P.; et al. New chronology and extended palaeoenvironmental data to the 1975 loess profile of Madaras brickyard, South Hungary. J. Quat. Sci. 2021, 36, 1364–1381. [Google Scholar] [CrossRef]
- Osipova, E.; Danukalova, G.; Marković, S.B. Malacological characteristics of the Middle to Upper Pleistocene transitional interval (MIS 7–5) observed in the Batajnica locality (Serbia). Quat. Int. 2013, 292, 86–100. [Google Scholar] [CrossRef]
- Molnár, D.; Makó, L.; Cseh, P.; Sümegi, P.; Fekete, I.; Galović, L. Middle and Late Pleistocene loess-palaeosol archives in East Croatia: Multi-proxy palaeoecological studies on Zmajevac and Šarengrad II sequences. Stud. Quat. 2021, 38, 3–17. [Google Scholar] [CrossRef]
- Hupuczi, J.; Molnár, D.; Galović, L.; Sümegi, P. Preliminary malacological investigation of the loess profile at Šarengrad, Croatia. Open Geosci. 2010, 2, 57–63. [Google Scholar] [CrossRef]
- Molnár, D.; Hupuczi, J.; Galović, L.; Sümegi, P. Preliminary malacological investigation on the loess profile at Zmajevac, Croatia. Open Geosci. 2010, 2, 52–56. [Google Scholar] [CrossRef]
- Marković, S.B.; Oches, E.A.; Perić, Z.M.; Gaudenyi, T.; Jovanović, M.; Sipos, G.; Thiel, C.; Buylaert, J.; Savić, S.; McCoy, W.D.; et al. The Požarevac loess–paleosol sequence: A record of increased aridity in the south-eastern margin of the Carpathian Basin during the last 350 ka. J. Quat. Sci. 2021, 36, 1436–1447. [Google Scholar] [CrossRef]
- Marković, S.B.; Oches, E.; Sümegi, P.; Jovanović, M.; Gaudenyi, T. An introduction to the Middle and Upper Pleistocene loess–paleosol sequence at Ruma brickyard, Vojvodina, Serbia. Quatern. Int. 2006, 149, 80–86. [Google Scholar] [CrossRef]
- Markovic, S.B.; Oches, E.; Gaudenyi, T.; Jovanovic, M.; Hambach, U.; Zöller, L.; Sümegi, P. Palaeoclimate record in the Late Pleistocene loess-paleosol sequence at Miseluk (Vojvodina, Serbia) [Le cycle climatique du Pléistocènesupérieur dans la séquenceloessique de Miseluk (Vojvodina, Serbie)]. Quaternaire 2004, 15, 361–368. [Google Scholar] [CrossRef]
- Marković, S.; Gaudenyi, T.; Jovanović, M.; Stevens, T.; Walther, R.; Ivanišević, P.; Galić, Z. Paleoclimate record in the Upper Pleistocene loess-paleosol sequence at Petrovaradin brickyard (Vojvodina, Serbia). Geol. Carpath. 2005, 56, 545–552. [Google Scholar]
- Markovic, S.; Hambach, U.; Gaudenyi, T.; Jovanovic, M.; Zöller, L.; Machalett, B.; Savic, S.; Romelic, J.; Plavsa, J.; Mesaros, M. An introduction to the late Pleistocene loess-paleosol sequence at Susek, Vojvodina, Serbia. Geogr. Pannonica 2006, 10, 4–8. [Google Scholar] [CrossRef]
- Marković, S.B.; Oches, E.A.; McCoy, W.D.; Frechen, M.; Gaudenyi, T. Malacological and sedimentological evidence for “warm” glacial climate from the Irig loess sequence, Vojvodina, Serbia. Geochem. Geophy. Geosy. 2007, 8, 3309. [Google Scholar] [CrossRef]
- Gaudenyi, T. Paleoclimatical and Paleoecological Reconstruction of the Last Glacial on the Titel Loess Plateau. Ph.D. Thesis, University of Novi Sad, Novi Sad, Serbia, 2009. in press. [Google Scholar]
- Gavrilović, B.; Sümegi, P.; Ćirić, M.; Radaković, M.G.; Gavrilov, M.B.; Mlađan, D.; Marković, S.B. The Middle and Late Pleniglacial (Weichselian) malacofauna of the Zemun loess-paleosol sequence, Serbia. PalZ 2019, 94, 519–531. [Google Scholar] [CrossRef]
- Constantin, D.; Veres, D.; Panaiotu, C.; Anechitei-Deacu, V.; Groza, S.; Begy, R.; Kelemen, S.; Buylaert, J.-P.; Hambach, U.; Marković, S.; et al. Luminescence age constraints on the Pleistocene-Holocene transition recorded in loess sequences across SE Europe. Quat. Geochronol. 2018, 49, 71–77. [Google Scholar] [CrossRef]
- Constantin, D.; Mason, J.; Veres, D.; Hambach, U.; Panaiotu, C.; Zeeden, C.; Zhou, L.; Marković, S.; Gerasimenko, N.; Avram, A.; et al. OSL-dating of the Pleistocene-Holocene climatic transition in loess from China, Europe and North America, and evidence for accretionary pedogenesis. Earth-Sci. Rev. 2021, 221, 103769. [Google Scholar] [CrossRef]
- Lehmkuhl, F.; Bösken, J.; Hošek, J.; Sprafke, T.; Marković, S.B.; Obreht, I.; Hambach, U.; Sümegi, P.; Thiemann, A.; Steffens, S.; et al. Loess distribution and related Quaternary sediments in the Carpathian Basin. J. Maps 2018, 14, 661–670. [Google Scholar] [CrossRef]
- Kerney, M.P.; Jungbluth, J.H. Die Landschneckennord-und Mitteleuropas; Paul Parey: Berlin, Germany, 1983. [Google Scholar]
- Pfleger, V. A Field Guide in Colour to Molluscs; Silverdale Books: Prague, Czech Republic, 2000. [Google Scholar]
- Fehér, Z.; Deli, T.; Solymos, P. Revision of Granaria frumentum (Draparnaud 1801) (Mollusca, Gastropoda, Chondrinidae) subspecies occurring in the eastern part of the species’ range. J. Conchol. 2010, 40, 201. [Google Scholar]
- Welter-Schultes, F.W. European Non-Marine Molluscs, a Guide for Species Identification: Bestimmungsbuch für Europäische Land-und Süsswassermollusken; Planet Poster Editions: Goettingen, Germany, 2012. [Google Scholar]
- Nekola, J.C.; Coles, B.F.; Horsák, M. Species assignment in Pupilla (Gastropoda: Pulmonata: Pupillidae): Integration of DNA-sequence data and conchology. J. Molluscan Stud. 2015, 81, 196–216. [Google Scholar] [CrossRef]
- Sysoev, A.; Shileyko, A. Land Snails and Slugs of Russian and Adjacent Countries Sofia-Moscow; Pensoft Publ.: Sofia, Bulgaria, 2009; p. 312. [Google Scholar]
- Juřičková, L.; Ložek, V.; Horáčková, J.; Tlachač, P.; Horáček, I. Holocene succession and biogeographical importance of mollusc fauna in the Western Sudetes (Czech Republic). Quat. Int. 2014, 353, 210–224. [Google Scholar] [CrossRef]
- Hertelendi, E.; Sümegi, P.; Szöör, G. Geochronologic and Paleoclimatic Characterization of Quaternary Sediments in the Great Hungarian Plain. Radiocarbon 1992, 34, 833–839. [Google Scholar] [CrossRef]
- Sümegi, P. Upper Pleistocene Evolution History of the Hajdusag (Hungary) Region on the Basis of Stratigraphical Investigations; University of Debrecen: Debrecen, Hungary, 1989; p. 93. [Google Scholar]
- Sólymos, P.; Sümegi, P. The Shell Morpho-Thermometer Method and Its Application in Palaeoclimatic Reconstruction. Ann. Univ. Sci. Budapestiensis Sect. Geol. 1999, 32, 137–148. [Google Scholar]
- Sümegi, P. Refuting ideas based on small batch of data: Malacothermometry aid in the reconstruction of mean july paleo-temperatures in the Carpathian basin for the last glacial of the pleistocene [Több természettudományi adatot és kevesebb mítoszt-malakohőmérő módszerrel rekonstruált egykori júliusi hőmérsékleti adatok a jégkor utolsó löszképződési periódusában kifejlődött valódi szárazföldi környezetre vonatkozóan a Kárpát-medencében]. Archeometriai Műhely 2019, 16, 143–166. [Google Scholar]
- Kordos, L. Change in the Holocene climate of Hungary reflected by the “vole thermometer” method. Földrajzi Közlemények 1957, 25, 222–229. [Google Scholar]
- Coope, G.; Morgan, A.; Osborne, P. Fossil coleoptera as indicators of climatic fluctuations during the last glaciation in Britain. Palaeogeogr. Palaeoclim. Palaeoecol. 1971, 10, 87–101. [Google Scholar] [CrossRef]
- Heusser, C.J. Climatic records of the Last Ice Age for Pacific mid-latitudes in the Americas. In Proceedings of the 9th INQUA Congress, Christchurch, New Zealand, 2–10 December 1973; 150p. [Google Scholar]
- Ludwig, P.; Gavrilov, M.B.; Radaković, M.G.; Marković, S.B. Malaco temperature reconstructions and numerical simulation of environmental conditions in the southeastern Carpathian Basin during the Last Glacial Maximum. J. Quat. Sci. 2021, 36, 1426–1435. [Google Scholar] [CrossRef]
- Hao, Q.; Wang, L.; Oldfield, F.; Peng, S.; Qin, L.; Song, Y.; Xu, B.; Qiao, Y.; Bloemendal, J.; Guo, Z. Delayed build-up of Arctic ice sheets during 400,000-year minima in insolation variability. Nature 2012, 490, 393–396. [Google Scholar] [CrossRef] [PubMed]
- Lisiecki, L.E.; Raymo, M.E. A Pliocene-Pleistocene stack of 57 globally distributed benthic δ 18O records. Paleoceanography 2005, 20, PA1003. [Google Scholar] [CrossRef]
- Porter, S.C.; Zhisheng, A. Correlation between climate events in the North Atlantic and China during the last glaciation. Nature 1995, 375, 305–308. [Google Scholar] [CrossRef]
- Vandenberghe, J.; Mucher, H.J.; Roebroeks, W.; Gemke, D. Lithostratigraphy and paleoenvironment of the Pleistocene deposits at Maaschricht-Belvédère, Southern Limburg, the Netherlands. Meded. Rijks Geol. Dienst 1985, 39, 7–29. [Google Scholar]
- Vandenberghe, J.; Zhisheng, A.; Nugteren, G.; Huayu, L.; Van Huissteden, K. New absolute time scale for the Quaternary climate in the Chinese Loess region by grain-size analysis. Geology 1997, 25, 35–38. [Google Scholar] [CrossRef]
- Sümegi, P.; Krolopp, E. Palaeoecological reconstruction of the Hungarian Weichselian loess deveopment. Földtani Közlöny 1995, 124, 125–148, (In Hungarian with English Abstract). [Google Scholar]
- Schmidt, E.; Machalett, B.; Marković, S.; Tsukamoto, S.; Frechen, M. Luminescence chronology of the upper part of the Stari Slankamen loess sequence (Vojvodina, Serbia). Quat. Geochronol. 2010, 5, 137–142. [Google Scholar] [CrossRef]
- Murray, A.S.; Schmidt, E.D.; Stevens, T.; Buylaert, J.P.; Marković, S.B.; Tsukamoto, S.; Frechen, M. Dating Middle Pleistocene loess from Stari Slankamen (Vojvodina, Serbia)—Limitations imposed by the saturation behaviour of an elevated temperature IRSL signal. Catena 2014, 117, 34–42. [Google Scholar] [CrossRef]
- Avram, A.; Constantin, D.; Veres, D.; Kelemen, S.; Obreht, I.; Hambach, U.; Slobodan, B.; Marković, A.B.; Timar-Gabor, A. Testing polymineral post-IR IRSL and quartz SAR-OSL protocols on Middle to Late Pleistocene loess at Batajnica, Serbia. Boreas 2020, 49, 615–633. [Google Scholar] [CrossRef]
- Namier, N.; Hao, Q.; Gao, X.; Fu, Y.; Marković, S.B.; Hambach, U.; Veres, D.; Mason, J.A.; Song, Y.; Deng, C.; et al. Comprehensive magnetic analysis of the tephras in Middle-Late Pleistocene loess records of Serbia, and implications for tephra identification, correlation and loess chronology. Quat. Sci. Rev. 2023, 313, 108202. [Google Scholar] [CrossRef]
- Laag, C.; Hambach, U.; Zeeden, C.; Lagroix, F.; Guyodo, Y.; Veres, D.; Jovanović, M.; Marković, S.B. A detailed paleoclimate proxy record for the middle danube basin over the last 430 kyr: A rock magnetic and colorimetric study of the Zemun loess-paleosol sequence. Front. Earth Sci. 2021, 9, 600086. [Google Scholar] [CrossRef]
- Marković, S.B.; Hambach, U.; Catto, N.; Jovanović, M.; Buggle, B.; Machalett, B.; Zöller, L.; Glaser, B.; Frechen, M. Middle and Late Pleistocene loess sequences at Batajnica, Vojvodina, Serbia. Quat. Int. 2009, 198, 255–266. [Google Scholar] [CrossRef]
- Marković, S.B.; Hughes, P.D.; Schaetzl, R.; Gibbard, P.L.; Hao, Q.; Radaković, M.G.; Vandenberghe, J.; Obreht, I.; Sipos, G.; Laag, C.; et al. The relationship between the loess stratigraphy in the Vojvodina region of northern Serbia and the Saalian and Rissian Stage glaciations—A review. Boreas, 2024; online version of record before inclusion in an issue. [Google Scholar]
- Kukla, G.J. Loess Stratigraphy of Central Europe. In After the Australopithecines; Butzer, K.W., Isaac, G.L., Eds.; Mouton: The Hague, The Netherlands, 1975; pp. 99–187. [Google Scholar]
- Marković-Marjanović, J. Data concerning the stratigraphy and the fauna of the lower and middle pleistocene of Yugoslavia. Palaeogeogr. Palaeoclim. Palaeoecol. 1970, 8, 153–163. [Google Scholar] [CrossRef]
- Marković-Marjanović, J. Rasprostranjenje i stratigrafija lesa u Juguslaviji. Glas. Prir. Muzeja Ser. A 1972, 27, 93–107. [Google Scholar]
- Butrym, J. Profil lessowy Stari Slankamen—Čot Yugoslawia. Ann. Univ. Mariae Curie-Sklodowska Sec. B 1974, 26, 113–133. [Google Scholar]
- Bronger, A. Zur Quartören Klima-und Landschaftsentwicklung des Karpatenbeckens auf (Palaeo-) Pedologischer und Bodengeographischer Grundlage. Ph.D. Thesis, Kiel University, Kiel, Germany, 1976. (In German). [Google Scholar]
- Marković, S.B.; Sümegi, P.; Stevens, T.; Schaetzl, R.J.; Obreht, I.; Chu, W.; Buggle, B.; Zech, M.; Zech, R.; Zeeden, C.; et al. The Crvenka loess-paleosol sequence: A record of continuous grassland domination in the southern Carpathian Basin during the Late Pleistocene. Palaeogeogr. Palaeoclim. Palaeoecol. 2018, 509, 33–46. [Google Scholar] [CrossRef]
- Radaković, M.G.; Tošić, I.; Bačević, N.; Mladjan, D.; Gavrilov, M.B.; Marković, S.B. The analysis of aridity in Central Serbia from 1949 to 2015. Theor. Appl. Clim. 2017, 133, 887–898. [Google Scholar] [CrossRef]
- Obreht, I.; Zeeden, C.; Hambach, U.; Veres, D.; Marković, S.B.; Bösken, J.; Svirčev, Z.; Bačević, N.; Gavrilov, M.B.; Lehmkuhl, F. Tracing the influence of Mediterranean climate on Southeastern Europe during the past 350,000 years. Sci. Rep. 2016, 6, 36334. [Google Scholar] [CrossRef] [PubMed]
- Buggle, B.; Hambach, U.; Kehl, M.; Markovic, S.B.; Zoller, L.; Glaser, B. The progressive evolution of a continental climate in southeast-central European lowlands during the Middle Pleistocene recorded in loess paleosol sequences. Geology 2013, 41, 771–774. [Google Scholar] [CrossRef]
- Radaković, M.G.; Oches, E.A.; Hughes, P.D.; Marković, R.S.; Hao, Q.; Perić, Z.M.; Gavrilović, B.; Ludwig, P.; Lukić, T.; Gavrilov, M.B.; et al. Reconstructed Malacothermometer July Paleotemperatures from the Last Nine Glacials over the South-Eastern Carpathian Basin (Serbia). Atmosphere 2023, 14, 791. [Google Scholar] [CrossRef]
- Bennett, K.D. Milankovitch cycles and their effects on species in ecological and evolutionary time. Paleobiology 1990, 16, 11–21. [Google Scholar] [CrossRef]
- Cronin, T.M.; Ikeya, N.; Ross, R.M.; Allmon, W.D. Tectonic events and climatic change: Opportunities for speciation in Cenozoic marine Ostracoda. In Causes of Evolution—A Paleontological Perspective; University of Chicago Press: Chicago, IL, USA; London, UK, 1990; pp. 210–248. [Google Scholar]
- Cronin, T.M. Principles of Paleoclimatology; Columbia University Press: New York, NY, USA, 1999. [Google Scholar]
- Aswan; Ozawa, T. Milankovitch 41,000-year cycles in lithofacies and molluscan content in the tropical Middle Miocene Nyalindung Formation, Jawa, Indonesia. Palaeogeogr. Palaeoclim. Palaeoecol. 2006, 235, 382–405. [Google Scholar] [CrossRef]
- Wu, N.; Rousseau, D.D.; Liu, X. Response of mollusk assemblages from the Luochuan loess section to orbital forcing since the last 250 ka. Chin. Sci. Bull. 2000, 45, 1617–1622. [Google Scholar] [CrossRef]
- Wu, N.; Rousseau, D.D.; Liu, T.; Lu, H.; Zhaoyan, G.; Guo, Z.; Jiang, W. Orbital forcing of terrestrial mollusks and climatic changes from the Loess Plateau of China during the past 350 ka. J. Geophys. Res. Atmos. 2001, 106, 20045–20054. [Google Scholar] [CrossRef]
- Laskar, J.; Robutel, P.; Joutel, F.; Gastineau, M.; Correia, A.C.M.; Levrard, B. A long-term numerical solution for the insolation quantities of the Earth. Astron. Astrophys. 2004, 428, 261–285. [Google Scholar] [CrossRef]
- Drysdale, R.N.; Zanchetta, G.; Hellstrom, J.C.; Fallick, A.E.; Zhao, J.-X.; Isola, I.; Bruschi, G. Palaeoclimatic implications of the growth history and stable isotope (δ18O and δ13C) geochemistry of a Middle to Late Pleistocene stalagmite from central-western Italy. Earth Planet. Sci. Lett. 2004, 227, 215–229. [Google Scholar] [CrossRef]
- Tzedakis, P.C.; Roucoux, K.H.; de Abreu, L.; Shackleton, N.J. The duration of Forest stages in southern europe and interglacial climate variability. Science 2004, 306, 2231–2235. [Google Scholar] [CrossRef] [PubMed]
- Roucoux, K.; Tzedakis, P.; de Abreu, L.; Shackleton, N. Climate and vegetation changes 180,000 to 345,000 years ago recorded in a deep-sea core off Portugal. Earth Planet. Sci. Lett. 2006, 249, 307–325. [Google Scholar] [CrossRef]
- Desprat, S.; Sánchez Goñi, M.F.; McManus, J.F.; Duprat, J.; Cortijo, E. Millennial-scale climatic variability between 340,000 and 270 000 years ago in SW Europe: Evidence from a NW Iberian margin pollen sequence. Clim. Past. 2009, 5, 53–72. [Google Scholar] [CrossRef]
- Railsback, L.B.; Gibbard, P.L.; Head, M.J.; Voarintsoa, N.R.G.; Toucanne, S. An optimized scheme of lettered marine isotope substages for the last 1.0 million years, and the climatostratigraphic nature of isotope stages and substages. Quat. Sci. Rev. 2015, 111, 94–106. [Google Scholar] [CrossRef]
- Cheng, H.; Edwards, R.L.; Broecker, W.S.; Denton, G.H.; Kong, X.; Wang, Y.; Zhang, R.; Wang, X. Ice age terminations. Science 2009, 326, 248–252. [Google Scholar] [CrossRef] [PubMed]
- Hao, Q.; Peng, S.; Gao, X.; Marković, S.B.; Li, S.-H.; Zhang, J.; Li, F.; Han, L.; Fu, Y.; Wu, X.; et al. Unusual weakening trend of the East Asian winter monsoon during MIS 8 revealed by Chinese loess deposits and its implications for ice age dynamics. Glob. Planet. Chang. 2024, 234, 104389. [Google Scholar] [CrossRef]
- Regattieri, E.; Zanchetta, G.; Isola, I.; Bajo, P.; Perchiazzi, N.; Drysdale, R.N.; Boschi, C.; Hellstrom, J.C.; Francke, A.; Wagner, B. A MIS 9/MIS 8 speleothem record of hydrological variability from Macedonia (FYROM). Glob. Planet Change 2018, 162, 39–52. [Google Scholar] [CrossRef]
- Temovski, M.; Wieser, A.; Marchhart, O.; Braun, M.; Madarász, B.; Kiss, G.I.; Palcsu, L.; Ruszkiczay-Rüdiger, Z. Pleistocene valley incision, landscape evolution and inferred tectonic uplift in the central parts of the Balkan Peninsula–Insights from the geochronology of cave deposits in the lower part of Crna Reka basin (N. Macedonia). Geomorphology 2024, 445, 108994. [Google Scholar] [CrossRef]
- Fletcher, W.J.; Müller, U.C.; Koutsodendris, A.; Christanis, K.; Pross, J. A centennial-scale record of vegetation and climate variability from 312 to 240 ka (Marine Isotope Stages 9c–a, 8 and 7e) from Tenaghi Philippon, NE Greece. Quat. Sci. Rev. 2013, 78, 108–125. [Google Scholar] [CrossRef]
- Sadori, L.; Koutsodendris, A.; Panagiotopoulos, K.; Masi, A.; Bertini, A.; Combourieu-Nebout, N.; Francke, A.; Kouli, K.; Joannin, S.; Mercuri, A.M.; et al. Pollen-based paleoenvironmental and paleoclimatic change at Lake Ohrid (south-eastern Europe) during the past 500 ka. Biogeosciences 2016, 13, 1423–1437. [Google Scholar] [CrossRef]
- Wagner, B.; Wilke, T.; Francke, A.; Albrecht, C.; Baumgarten, H.; Bertini, A.; Combourieu-Nebout, N.; Cvetkoska, A.; D’Addabbo, M.; Donders, T.H.; et al. The environmental and evolutionary history of Lake Ohrid (FYROM/Albania): Interim results from the SCOPSCO deep drilling project. Biogeosciences 2017, 14, 2033–2054. [Google Scholar] [CrossRef]
- Wagner, B.; Tauber, P.; Francke, A.; Leicher, N.; Binnie, S.A.; Cvetkoska, A.; Jovanovska, E.; Just, J.; Lacey, J.H.; Levkov, Z.; et al. The geodynamic and limnological evolution of Balkan Lake Ohrid, possibly the oldest extant lake in Europe. Boreas 2022, 52, 1–26. [Google Scholar] [CrossRef]
- Ulfers, A.; Zeeden, C.; Voigt, S.; Abadi, M.S.; Wonik, T. Half-precession signals in Lake Ohrid (Balkan) and their spatio-temporal relations to climate records from the European realm. Quat. Sci. Rev. 2022, 280, 107413. [Google Scholar] [CrossRef]
- Tzedakis, P.; Hooghiemstra, H.; Pälike, H. The last 1.35 million years at Tenaghi Philippon: Revised chronostratigraphy and long-term vegetation trends. Quat. Sci. Rev. 2006, 25, 3416–3430. [Google Scholar] [CrossRef]
- Hughes, P.D.; Gibbard, P.L.; Ehlers, J. The “missing glaciations” of the Middle Pleistocene. Quat. Res. 2020, 96, 161–183. [Google Scholar] [CrossRef]
- Wagner, B.; Vogel, H.; Francke, A.; Friedrich, T.; Donders, T.; Lacey, J.H.; Leng, M.J.; Regattieri, E.; Sadori, L.; Wilke, T.; et al. Mediterranean winter rainfall in phase with African monsoons during the past 1.36 million years. Nature 2019, 573, 256–260. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Unit | Thickness (cm) | Depth (cm) | Description |
|---|---|---|---|
| L3 | 460 | 2370–2830 | Massive, porous pale yellow (5YR 4/3, 5/4) loess with many humus infiltrations and carbonate concretions (ø 1–3 cm), with Fe and Mn nodules in the lowest half meter, intensively bioturbated in the upper part in contact with the base of paleosol S2 above. |
| S3 | 60 | 2830–2840 | Mollic A horizon with brighter color (10YR 6/2–4) and many carbonate pseudomycelia. |
| 2840–2885 | Ah1 lower humic (10YR 6/3, 4/2) horizon with granular structure, carbonate concretions (ø 1–2 cm) and krotovinas. | ||
| 2885–2890 | Upper lighter Ah2 (10YR 6/3-4) horizon with carbonate concretions (ø 1–2); typical granular structure. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radaković, M.G.; Gavrilović, B.; Gavrilov, M.B.; Marković, R.S.; Hao, Q.; Schaetzl, R.J.; Zeeden, C.; Cai, B.; Perić, Z.M.; Antić, A.; et al. A Glacial–Interglacial Malacofauna Record from the Titel Loess Plateau, Serbia, between ~350 and 250 ka. Quaternary 2024, 7, 28. https://doi.org/10.3390/quat7020028
Radaković MG, Gavrilović B, Gavrilov MB, Marković RS, Hao Q, Schaetzl RJ, Zeeden C, Cai B, Perić ZM, Antić A, et al. A Glacial–Interglacial Malacofauna Record from the Titel Loess Plateau, Serbia, between ~350 and 250 ka. Quaternary. 2024; 7(2):28. https://doi.org/10.3390/quat7020028
Chicago/Turabian StyleRadaković, Milica G., Bojan Gavrilović, Milivoj B. Gavrilov, Rastko S. Marković, Qingzhen Hao, Randall J. Schaetzl, Christian Zeeden, Binggui Cai, Zoran M. Perić, Aleksandar Antić, and et al. 2024. "A Glacial–Interglacial Malacofauna Record from the Titel Loess Plateau, Serbia, between ~350 and 250 ka" Quaternary 7, no. 2: 28. https://doi.org/10.3390/quat7020028
APA StyleRadaković, M. G., Gavrilović, B., Gavrilov, M. B., Marković, R. S., Hao, Q., Schaetzl, R. J., Zeeden, C., Cai, B., Perić, Z. M., Antić, A., Lukić, T., & Marković, S. B. (2024). A Glacial–Interglacial Malacofauna Record from the Titel Loess Plateau, Serbia, between ~350 and 250 ka. Quaternary, 7(2), 28. https://doi.org/10.3390/quat7020028