Abstract
Recent investigations in the upper Lower–Middle Pleistocene deposits of the Megalopolis Basin (Greece) led to the discovery of several sites/findspots with abundant faunal material. Here, we provide an updated overview including new results on the micro- and macro-mammal fauna. Important new discoveries comprise partial hippopotamus skeletons from Marathousa 1 and the new Lower Pleistocene site Choremi 6, as well as a second partial elephant skeleton from Marathousa 1, including a complete tusk and the rarely found stylohyoideum. Based on the first results from the newly collected micromammals, we discuss age constraints of the sites, and we provide biostratigraphic/biochronologic remarks on key mammal taxa for the Middle Pleistocene of Greece and southeastern Europe. The presence of mammals highly dependent on freshwater for their survival, together with temperate-adapted ones in several stratigraphic layers of the basin, including those correlated with glacial stages, when conditions were colder and/or drier, indicate the capacity of the basin to retain perennial freshwater bodies under milder climatic conditions, even during the harsher glacial periods of the European Middle Pleistocene, and further support its refugial status. Yet, the smaller dimensions of the Megalopolis hippopotamuses may represent a response to the changing environmental conditions of the epoch, not optimal for hippopotamuses. Overall, the Megalopolis Basin comprises a unique fossil record for southeastern Europe and provides valuable insights into the Middle Pleistocene terrestrial ecosystems of Europe, and hominin adaptations in particular.
Keywords:
Marathousa; Choremi; Kyparissia; Palaeoloxodon; Hippopotamus; Mimomys; Arvicola; Pleistocene; Galerian; Greece 1. Introduction
The Megalopolis Basin (Arcadia, Peloponnesus, Greece; Figure 1a) is a tectonic half-graben that was filled from the Neogene onwards by continental deposits of fluviolacustrine origin. During the late Early and the Middle Pleistocene (ca. 900 to 150 ka), the basin hosted a large, shallow lake, which resulted in a stratigraphic sequence composed mainly of lacustrine sediments with lignite seams (Figure 1c) [1,2,3]. Since 1969, the mining of the lignite resulted in the exposure of long and high fossiliferous sections that offer the unique opportunity to study the stratigraphy and the paleoenvironment of the Pleistocene paleolake and its environs.
The existence of fossil remains in the Megalopolis Basin, especially of “giant” bones that we now know belong to elephants and hippopotamuses, is known since antiquity, when these were attributed to mythical beings (see, e.g., [4,5]). The first systematic excavations by T. Skouphos (University of Athens) in 1902 unearthed an important number of fossils and highlighted the wealth of the basin in fossil vertebrates and its potential for further paleontological research [6,7]. A part of this first collected material was studied in detail in a series of articles by I. Melentis during the 1960s [4,8,9,10,11,12,13,14,15]. Yet, as is common for historical collections, those specimens lack precise stratigraphic information, and the assemblage does not represent a single, stratified accumulation. Since then, several field surveys and excavations of Pleistocene fossils took place in the basin sporadically throughout the years, and numerous vertebrates’ remains have been collected and studied (e.g., [2,16,17,18,19,20]). Among other vertebrate remains collected was a surface-collected hominin molar ([17], see the Appendix by Marinos), which was recently attributed to the Neanderthal lineage [21].
Recent investigations in the basin were conducted by a joint team of the Ephorate of Paleoanthropology–Speleology, the University of Tübingen, and the American School of Classical Studies at Athens, in three phases: the field survey of 2012–2013, the systematic excavation of the site Marathousa 1 in 2013–2019, and the field survey of 2018–2022 (for further details, see, e.g., [22,23,24,25]). Aiming primarily to locate sites with paleoanthropological/archaeological interest from securely stratified contexts and to perform detailed paleoenvironmental analyses with chronological control, these investigations targeted the Choremi Formation (Fm) and, in particular, its lower part, the Marathousa Member (Mb). The Marathousa Mb, dated to late Early Pleistocene–Middle Pleistocene, is formed by cyclic sedimentation of lignite and clastic (detrital intervals) sediments, mostly of lacustrine origin, with the lignites thought to correspond to interglacial (warm) stages and the detrital intervals to glacial (cold) stages (Figure 1c) [2,3,26,27]. Three main lignite seams are identified (lignite seams I, II, and III, abbreviated as LI, LII, and LIII, respectively) and subdivided into individual seams within them (LIa–c, LIIa–b, and LIIIa–c), of which LIb encompasses the Matuyama/Brunhes geomagnetic polarity reversal, at 0.78 Ma, and is correlated with the warm Marine Isotope Stage (MIS) 19 (Figure 1c) [3,28].
The aims of the present study are the following: (1) to present an overview of the micro- and macro-mammalian fauna, including updates and new results; (2) to provide age constraints based on the newly collected micromammal fauna; (3) to provide biostratigraphic/biochronological remarks on the first and last occurrences of Middle Pleistocene mammals in Greece, as well as in the wider region of southeastern Europe; (4) to discuss the potential link between the harsher environmental conditions during the glacial stages of the Middle Pleistocene and the smaller size of the Megalopolis hippopotamuses; (5) to highlight the importance of the Megalopolis Basin as a glacial refugial area for several mammals, including elephants, hippopotamuses, and macaques.

Figure 1.
(a) Geographic position of the Megalopolis Basin (maps taken from Copernicus Land Monitoring Service: https://land.copernicus.eu/ and https://maps-for-free.com/ (accessed on 30 April 2024)). (b) Geographic position of the investigated sites within the lignite mines (numbered 1–4) of the Megalopolis Basin (satellite image from Google Earth). (c) Panoramic view of the eastern quarry of the Marathousa mine, indicating the Marathousa and Megalopolis Members of the Choremi Formation, and the lignite seams. (d) Simplified stratigraphic column of the Marathousa Member showing the lignite seams and the intercalating detrital intervals, their correlation to the Marine Isotope Stages (MIS) following the age model proposed by Tourloukis et al. [3], and the stratigraphic position of the sites and survey units following Karkanas et al. [25,28].
2. Materials and Methods
The material was recovered during the field investigations (field surveys and excavation) that took place in the Megalopolis Basin during 2012–2022. It is stored at the Ephorate of Paleoanthropology–Speleology in Athens. Systematic excavation was conducted at the site of Marathousa 1 from 2014 to 2019. The rest of the finds (stratified or surface finds, as noted accordingly in the article) were collected from sites (with a sufficient concentration of fossils; Figure 1b) or from findspots during the targeted survey units that systematically prospected the natural or artificial profiles of the mines. A survey unit (SU; CHO, KYP, and MAR in the Choremi, Kyparissia, and Marathousa mines, respectively; Figure 1b) is an arbitrary unit of observation delimited by natural, stratigraphic, or artificial boundaries, e.g., the start/end of a tier inside the mine [22,25]. All this material was collected from exposed artificial profiles, without the means of systematic excavation. The geodetic coordinates of survey units, collected or not collected specimens, and clear stratigraphic contacts were recorded by means of a Differential GPS in RTK mode, using the Hellenic Geodetic Reference System 1987 (HGRS87) and the local datum provided by the Megalopolis DEH.
In the following section, the fossiliferous sites and survey units are presented in chronological order (besides those that correspond roughly to the same stratigraphic position). Their stratigraphic position within the basin and their correlation to MIS are shown in Figure 1d. For the stratigraphic placement of the new find-bearing horizons, the first stratigraphic data from Karkanas et al. [25] were used, while for their age correlation, we employed the age model proposed by Tourloukis et al. [3]. This age model matches the second-option model proposed by Okuda et al. [27], and it was essentially (but preliminary) confirmed by both individual dating assays from the newly identified sites throughout the basin sequence (e.g., [25,29]), as well as by the correlation of the sequence with the Oxygen Isotope Curve (ongoing study).
Sediment samples (ca. 485 kg, dried weight) for small vertebrates were collected from the sites Kyparissia 4 (209 kg, including the test samples studied by Kolfschoten et al. [30]), Kyparissia 3 (ca. 91 kg), and Choremi 7 (ca. 162 kg), as well as from KYP SU 6 (23 kg) during the fieldwork seasons of 2019–2022. Samples were air-dried, soaked in a weak water solution of H2O2 (about 1% H2O2), wet sieved using a 0.5 mm mesh, and then left to air-dry. Sorting for vertebrate remains was conducted under a stereoscope. Here, we investigated only Rodentia, with a special focus on arvicolines. The study of the entire small mammal fauna, including Eulipotyphla, is pending. We focused on the m1, which is the most diagnostic and informative dental element in arvicolines. Measurements and terminology of arvicoline molars follow van der Meulen [31]. The La/Li index was calculated based on Luzi and Lopez Garcia [32]. Photographs and measurements (in mm, 0.01 accuracy) were taken with a Leica MZ 16A.
Upper premolars/molars are indicated with the upper-case letters P and M, respectively, and the corresponding lower ones with p and m. Dental and postcranial measurements of the large mammal specimens, as well as the updated faunal lists for the sites and SUs, are provided in the Supplementary Materials (Tables S1 and S2).
3. Fossiliferous Sites—Selected Survey Units and Findspots
3.1. Choremi 6 (CHO-6)
CHO-6 (Figure 1b and Figure 2) was discovered during the field survey of 2021, when several Hippopotamus bones were found stratified in grey clay, some of them partially exposed on the dried surface due the low water level of that year (Figure 2). CHO-6 is located just below LIa (Figure 1d); therefore, it predates the Matuyama/Bruhnes boundary and has an age older than MIS 19 [3,25]. As such, it is dated toward the end of the Lower Pleistocene and represents the oldest known fossiliferous site of the Choremi Fm.

Figure 2.
(a) Panoramic view of the site Choremi 6. (b–e) Fossils belonging to the Hippopotamus antiquus skeleton in situ at Choremi 6. (f) Distribution map showing the position of the hippopotamus remains (recorded with the use of a Differential GPS—WGS84 datum).
The proximate spatial distribution of the hippopotamus skeletal elements, the lack of duplication, and the consistency in size and ontogenetic age indicate the presence of a single, adult individual (all epiphyses of the long limb bones, the heads of the ribs, and the plates of the vertebrae are fused). Despite the absence of anatomical connection between the bones, the skeleton is not dissociated, and several originally articulated bones were found in very close spatial association (Figure 2). Moreover, most of the available bones belong to the posterior part of the skeleton (femur, patella, tibia, tarsals, metatarsals, pelvis, and lumbar vertebrae), and only few (scapula and mandible) are from the anterior part, most elements of which had already been removed before the discovery of the site (Figure 2 and Figure 3).
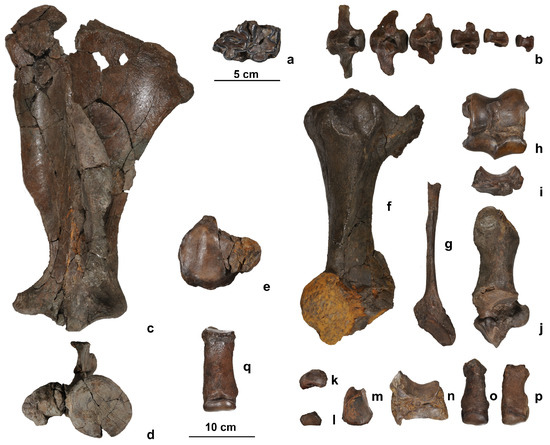
Figure 3.
Hippopotamus antiquus remains from Choremi 6. (a) Lower molar (m1?) in occlusal view; (b) series of caudal vertebrae (CHO-6-36, 37, 38, 39, 40, and 41); (c,d) left scapula (CHO-6-49) in lateral (c) and distal (d) view; (e) right patella (CHO-6-10) in caudal view; (f) left tibia (CHO-6-42) in dorsal view (note the presence of a limonite concretion on the distal end); (g) left fibula (CHO-6-43) in medial view; (h) left astragalus (CHO-6-7) in dorsal view; (i) left navicular (central tarsal, CHO-6-15) in lateral view; (j) left calcaneus (CHO-6-8) in medial view; (k) left ectocuneiform (tarsal I, CHO-6-17) in proximal view; (l) left mesocuneiform (tarsal II, CHO-6-25) in proximal view; (m) left entocuneiform (tarsal III, CHO-6-11) in distal view; (n) left cuboid (tarsal IV, CHO-6-13) in medial view; (o) left Mt II (CHO-6-20) in dorsal view; (p) left MT V (CHO-6-24) in dorsal view; (q) right MT III (CHO-6-21) in dorsal view.
In the biplots comparing variables of postcranial elements, the CHO-6 specimens are distinct from Hippopotamus amphibius, and due to their large size, they are plotted well within the sample of Hippopotamus antiquus, in most cases toward its lower range (Figure 4; Table S1). Compared to the other known hippopotamuses from the Megalopolis Basin (Kyparissia 1, Kyparissia 4, Kyparissia T, and Marathousa 1), the CHO-6 hippopotamus is the largest one, which is also reflected in the estimated body size of ca. 2400 kg (Table 1).
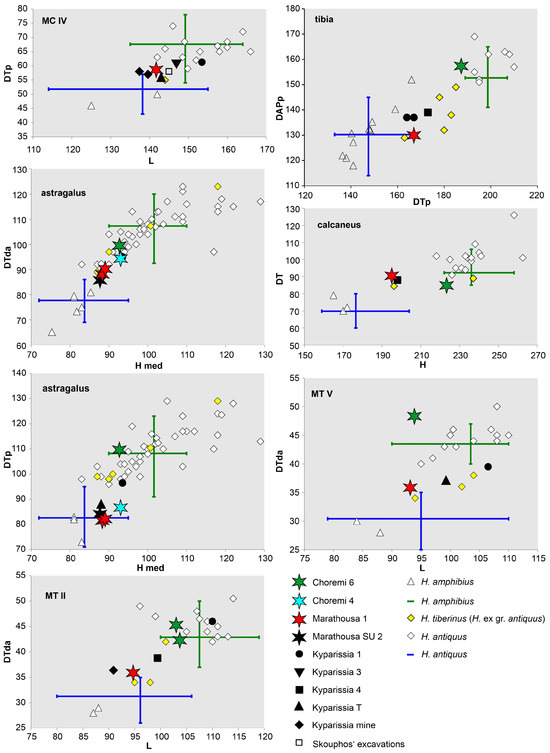
Figure 4.
Metrical comparison of Hippopotamus postcranial specimens from Megalopolis Basin with Hippopotamus antiquus, Hippopotamus tiberinus (H. ex gr. antiquus), and Hippopotamus amphibius. Data from [12,20,33,34,35,36,37]. The blue and green lines correspond to the ranges of H. amphibius and H. antiquus given by [38]. Abbreviations: a, articular facet; d, distal; DAP, anteroposterior diameter; DT, transverse diameter; H, height; L, length; med, medial; p, proximal.
Besides this individual, a large-sized cervid is represented by a right and left hemimandible and a maxilla fragment. All bear the deciduous dentition and the erupting first molar and belong to the same individual. The dimensions of the m1 are similar to those of the giant deer Praemegaceros verticornis from Kyparissia 4, yet in the absence of comparative material and of more diagnostic specimens, the CHO-6 specimen is attributed to Cervidae indet. (large-sized). Additionally, an avian large-sized distal phalanx and a turtle shell fragment were found.

Table 1.
Estimated body masses of Hippopotamus from several sites of the Megalopolis Basin (based on equations for humerus (H), femur (F), and tibia (T) in [39]) compared to Hippopotamus amphibius and Hippopotamus antiquus (data from [40]). Measurements for Kyparissia 1 and Kyparissia 4 were taken from [20]. Number in parentheses denotes the number of specimens measured.
Table 1.
Estimated body masses of Hippopotamus from several sites of the Megalopolis Basin (based on equations for humerus (H), femur (F), and tibia (T) in [39]) compared to Hippopotamus amphibius and Hippopotamus antiquus (data from [40]). Measurements for Kyparissia 1 and Kyparissia 4 were taken from [20]. Number in parentheses denotes the number of specimens measured.
| Site/Species | Measurement | Body Mass (kg) |
|---|---|---|
| Choremi 6 | T2 (1) | 2366 |
| Kyparissia 4 * | F5, T2, T4 (2) | 2111 |
| Kyparissia 1 ** | H5, T2, T4 (5) | 1893 |
| Marathousa 1 *** | H5, T2 (2) | 1738 |
| H. amphibius (range/mean) | 1048–2090/1476 | |
| H. antiquus (range/mean) | 1602–4694/3197 |
* Unknown whether femur and tibia belong to the same individual. ** Minimum of two individuals. *** Skeletally immature individual.
3.2. Kyparissia 4 (KYP-4)
KYP-4 (Figure 1b) was discovered in 2007 and since then has provided a rich and diverse assemblage of large mammals and other vertebrates [19,20]. The site lies above the limestone basement and is placed stratigraphically within clastic sediments close to the top of LI (Figure 1d) [25]. The site was revisited during the field survey of 2019 and subsequently in 2020–2022, further enriching the faunal assemblage in both the number of specimens and species. Of importance are the abundance of cranial and postcranial remains of the giant deer Praemegaceros verticornis (including three partial skeletons), a partial skeleton of Hippopotamus antiquus, and the first evidence for the presence of macaques at the site. Furthermore, the discovery of lithic artefacts provides evidence for the presence of humans.
The updated large mammal fauna includes the following taxa ([19,20,41,42] and ongoing study): Castor fiber (beaver), Macaca sylvanus (macaque), Vulpes sp. (fox), Felis sp. (wild cat), Palaeoloxodon antiquus (elephant), Stephanorhinus cf. hemitoechus (rhinoceros), Equus spp. (horse), Hippopotamus antiquus (hippopotamus), Sus scrofa (wild boar), Bison sp. (bison), Dama sp. (fallow deer), Cervus elaphus (red deer), and Praemegaceros verticornis (giant deer). A detailed study of the newly collected materials is in progress.
In addition to the large mammals and aiming to provide a secure biochronological framework of the site, sampling for micromammals was conducted in two phases. The first phase (test sampling) took place in 2019–2020 and showed the potential of the site for the preservation and diversity of micromammals [30], while during the second phase in 2021–2022, the systematic sampling significantly increased the available material. The KYP-4 rodent assemblage is dominated by the genus Mimomys. There is a clear size difference among the Mimomys molars, allowing the distinction of two taxa, the larger Mimomys, attributed to Mimomys cf. savini, and the smaller one attributed to Mimomys sp. (Figure 5a–d). Some rooted molars without crown cementum but with “Mimomys-enamel differentiation” are ascribed to the genus Pliomys (Figure 5e,f), yet their specific attribution is pending. Moreover, some unrooted molars show morphological features (e.g., in the m1, five more or less closed triangles and an arvalis-type anterior cap) that indicate the presence of the subgenus Microtus (Microtus). Additionally, two murid taxa are recognized, one with smaller dimensions and morphology that are compatible with Mus spretus, preliminary assigned here to Mus cf. spretus, and the other one with larger dimensions, attributed to Apodemus sp., which could possibly belong to Apodemus sylvaticus. Finally, the presence of the shrew Sorex sp. is also recognized.

Figure 5.
Small mammals from several sites and findspots of the Megalopolis Basin. (a,b) Right M2 (KYP-4-MM008) of Mimomys sp. (smaller size) from Kyparissia 4 in occlusal (a) and lingual (b) view; (c,d) left m1 (KYP-4-MM017) of Mimomys cf. savini from Kyparissia 4 in occlusal (c) and lingual (d) view; (e,f) left M3 (KYP-4-MM015) of Pliomys sp. from Kyparissia 4 in occlusal (e) and lingual (f) view; (g,h) left m2 (KYP-3-MM001) of Mimomys sp. (smaller size) from Kyparissia 3 in occlusal (g) and lingual (h) view; (i) right m1 (KYP SU 6-MM002) of Pliomys cf. episcopalis from Kyparrisia SU 6 in occlusal view; (j,k) left m1 (CHO-7-MM135) of Arvicola mosbachensis from Choremi 7 in occlusal (j) and lingual (k) view; (l) left m1 (CHO-7-MM66) of Microtus sp. (cf. Microtus arvalis) from Choremi 7 in occlusal view; (m) right m1 (CHO-7-MM214) of Microtus (Terricola) subterraneus from Choremi 7 in occlusal view.
3.3. Choremi 4 (CHO-4)
CHO-4 (Figure 1b and Figure 6) was discovered during the field survey of 2018 in Choremi Survey Unit 6 (CHO SU 6). Mammal fossils at CHO-4 were found within a mollusk-rich layer stratigraphically positioned between LIc and LIIa (Figure 1d and Figure 6a–c); therefore, a correlation to MIS 16 is possible (the detrital interval 3, in [3]). As such, CHO-4 stands stratigraphically higher and is younger than CHO-6 and KYP-4. The identified fossils include remains of turtle, beaver (cheek teeth, tibia fragment, and astragalus), deer, and hippopotamus. The latter is represented by an atlas, an astragalus, an ectocuneiform, and a proximal phalanx, all of which were found in very close spatial proximity (Figure 6c–h). The astragalus shows different proportions than the CHO-6 one, especially in terms of the DT proximal, and is similar to the astragali from KYP-T and MAR-1 (Figure 4). Even though most of the variables fall in the lower range of H. antiquus, it is closer to the mean values of H. tiberinus (H. ex gr. H. antiquus, according to [43]), except for the DT proximal, which is lower than in the latter species (Figure 4). In addition, a large bovid astragalus (Figure 6i–l) found on the surface close to the CHO-4 section is morphologically compatible with an attribution to Bison, following the criteria in [44].

Figure 6.
(a) Panoramic view of the Choremi 4 section showing the fossiliferous layer. (b) A tooth of Castor fiber in situ within a shell-rich layer. (c) In situ concentration of hippopotamus bones. (d,e) Right astragalus of Hippopotamus antiquus in dorsal (d) and plantar (e) view. (f) Right entocuneiform (tarsal III) in proximal view. (g,h) Atlas of H. antiquus in cranial (g) and caudal (h) view. (i–l) Right astragalus of Bison sp. found at the surface near the Choremi 4 section in dorsal (i), plantar (j), medial (k), and lateral (l) view.
3.4. Kyparissia 3 (KYP-3) and Kyparissia Survey Unit 6 (KYP SU 6)
KYP-3 (Figure 1b) was discovered in 2006 and studied in [19,20]. KYP-3 is located stratigraphically above KYP-4 and is placed in the upper part of the clastic unit underlying LII (Figure 1d) [19,25]. Similar to CHO-4, a correlation to MIS 16 is possible [3]. The site was revisited during the field survey of 2022 and resulted in the discovery of a partial skeleton of the elephant Palaeoloxodon antiquus (a third upper molar and several bones, mainly ribs), as well as artiodactyls (Hippopotamidae, Cervidae, and Suidae), turtles, and birds, in direct spatial and stratigraphic association with lithic artefacts and, thus, human presence [25]. The large mammal fauna includes the following taxa ([19,20]; ongoing study): Hyaenidae indet., Felis sp., Panthera sp. (Panthera ?pardus), Palaeoloxodon antiquus, Sus scrofa, Hippopotamus antiquus, Dama sp., Cervus elaphus, and Praemegaceros verticornis. A detailed study of the new materials is in progress.
The samples for micromammals collected from Kyparissia 3 were very poor and yielded only one arvicoline molar, a rooted m2 (Figure 5g,h), which, although not diagnostic, its dimensions denote the presence of a Mimomys species, smaller than M. savini. On the other hand, test sampling from an adjacent section to KYP-3 (named as KYP SU 6), of comparable stratigraphic position, proved to be much richer and promising for future investigations. From KYP SU 6, rooted arvicoline molars preserving crown cementum belong to Mimomys. The size difference among the molars indicates the presence of the larger M. cf. savini and of a smaller sized one, attributed to Mimomys sp. A third arvicoline is identified by a m1 (Figure 5i), which preserves no cement in the synclines, thick enamel with positive differentiation, a small T6, very shallow BRA4, and a rounded AC (anteroconid). These traits are similar to those of Pliomys episcopalis, yet due to the single status of the specimen and its larger size, we prefer to ascribe it to Pliomys cf. episcopalis. Larger-sized Pliomys cf. episcopalis were also found in Choremiou Section 3, a feature that was interpreted as an endemic feature [2].
3.5. Marathousa Survey Unit 7 (MAR SU 7)
MAR SU 7 was investigated in the field seasons of 2018 and 2019 and comprises a long section along the terrace at the northern part of the Marathousa mine, just below LIIa (Figure 1d and Figure 7a,b), thus possibly corresponding to MIS 16 [3]. Several fossils were discovered within two stratigraphically close clastic layers rich in mollusks and organic material. Of interest are fossils of a small-sized cervid and of a mustelid. To the former belong a radius and a metatarsal (Figure 7i–l; Table S1), whose dimensions are smaller than corresponding specimens from Kyparissia 4, Marathousa 1, and Marathousa 2, referred to Dama sp. [24,42,45], as well as from “Cervus” peloponnesiacus [17], and indicate a Capreolus-sized cervid (e.g., [46]), which is a rare faunal element in the Megalopolis Basin [19]. The mustelid is represented by an almost complete humerus characterized by strong dorsoventral curvature of the shaft, a laterally expanding supracondylar crest, large deltoid tuberosity, strong lateral curvature of the deltoid ridge, and broad distal epiphysis (Figure 7e–h). These traits, combined with its dimensions (Table S1), are similar to the humerus of the otter Lutra simplicidens from Voigtstedt (Germany [47]), and it is, therefore, attributed to this species. This is the second finding of L. simplicidens in the Megalopolis Basin, previously known only from MAR-1 [45]. From these mollusk-rich layers, fossils of Castor fiber (calcaneus, distal radius; Figure 7c,d), Cervus elaphus (metacarpal, radius), turtle shells, and bird bones were also found.

Figure 7.
(a) Panoramic view of the northern profile in the Marathousa mine, indicating the Marathousa and Megalopolis Members, the lignite seams, and the location of the MAR SU 7. (b) Details of the profile showing the MAR SU 7 section. (c,d) Left calcaneus of Castor fiber in medial (c) and plantar (d) view. (e–h) Left humerus of Lutra simplicidens in cranial (e), caudal (f), medial (g), and lateral (h) view. (i,j) Left radius of a Capreolus-sized cervid in dorsal (i) and volar (j) view. (k,l) Metacarpal of a Capreolus-sized cervid in dorsal (k) and volar (l) view.
3.6. Kyparissia T (KYP-T)
KYP-T (Figure 1b) was discovered during the field survey of 2012 close to the western margin of the Megalopolis Basin and was systematically excavated in the same year [20]. Stratigraphically, KYP-T was preliminarily placed on the top of LII (Figure 1d) [20], based on the presence of a thin calcareous layer that was identified as the guide horizon for LII, according to [48]. Recent stratigraphic studies at Kyparissia position KYP-3 and KYP-4 lower in the stratigraphic sequence of the Megalopolis Basin, namely, in the upper part of the clastic unit underlying LII, and close to the top of LI, respectively [25]. Thus, KYP-T may actually be older, as it lies at the same altitude as KYP-4. However, the current conditions at KYP-T (collapsed section and growth of dense vegetation) do not allow for new, detailed stratigraphic observations.
The faunal content of KYP-T consists mainly of several Hippopotamus antiquus specimens and scarce cervid, avian, and chelonian finds [20]. Those of Hippopotamus are rather small sized and may belong to a single individual.
3.7. Marathousa 1 (MAR-1)
MAR-1 (Figure 1b) was discovered during the field survey of 2013 and was systematically excavated until 2019 [22,23,49,50]. The locality is situated between LIIb and LIIIa (Figure 1d), is radiometrically, magnetostratigraphically, and biochronologically dated to ca. 450 ka, and is correlated to the glacial Marine Isotope Stage (MIS) 12 [3,28,45,51,52,53]. MAR-1 has yielded a stratified and exceptionally well-preserved archaeological and paleontological assemblage, including lithic and bone artefacts, micro- and macro-fauna (insects, ostracods, mollusks, fishes, amphibians, reptiles, birds, and mammals), and micro- and macro-flora ([45,52,54,55,56,57,58,59,60] and ongoing studies). Additionally, it preserves evidence of hominin butchering of elephant and other large mammal carcasses, in accordance with the traits of the associated lithic artefacts and the presence of use-wear traces involving butchering activities [45,59,61]. As such, it provides important insights into hominin subsistence strategies and, in particular, megafauna exploitation.
The micro- and macro-mammal fauna of MAR-1 was studied in [52] and [45], respectively. The latter study included the mammal remains from the 2013–2016 collection. The subsequent excavation seasons of 2017–2019 significantly enriched the collection from both areas. In Area A, several more bones of the Palaeoloxodon skeleton were unearthed, and these will be treated in detail separately. In Area B, the mammal assemblage increased significantly in the number of specimens, including a complete mandible of the macaque Macaca sylvanus cf. pliocena, which represents a new entry in the faunal list [57], a partial skeleton of Hippopotamus, several elephant remains belonging to a partial skeleton, as well as several remains of beavers, carnivorans, deer, and bovids.
Hippopotamus skeleton from Area B. The skeleton of Hippopotamus was found during the mechanical expansion in the southern part of Area B in 2019, within the sedimentary unit UB2b (dark grey, laminated, organic-rich silty sand, and sand rich in shell fragments, interpreted to represent relatively high-energy fluviatile flows entering the margins of the lake within the zone of lake-surface fluctuation [28]) and was subsequently systematically excavated (Figure 8). Several hippopotamus teeth and postcranial remains collected in this southern part of the trench during the initial opening of the site in 2013 belong most possibly to this skeleton, as they fully match in the status of bone preservation, dimensions, and ontogenetic age, while there is no duplication of skeletal elements (Figure 8). The spatial distribution of the recorded skeletal remains shows that the skeleton, although dissociated, retained in proximity anatomically connected bones, indicating minimal disturbance (Figure 8a). Moreover, most of the recorded bones belong to the posterior part of the skeleton (sacrum, lumbar vertebra, femur, tibia, and tarsals). Considering all available specimens, the skeleton is represented by dental and all types of axial and appendicular (both front- and hind-limb) elements, indicating that there was no selective bone survival.

Figure 8.
Hippopotamus antiquus skeleton from Marathousa 1 (Area B; UB2b). (a) Distribution map showing the position (recorded with the use of a total station) of the hippopotamus remains; (b) hippopotamus fossils in situ; (c) sacrum (MAR-1B-926/587-17) in ventral view; (d) right radius (MAR-1B-1) in dorsal view; (e) right patella (MAR-1B-925/587-2) in caudal view; (f) right femur (MAR-1B-925/587-1) in cranial view; (g) right tibia (MAR-1B-926/587-10) in dorsal view; (h) right astragalus (MAR-1B-926/587-4) in dorsal view; (i) right calcaneus (MAR-1B-2) in medial view; (j) right navicular (central tarsal, MAR-1B-924/587-3) in lateral view; (k) right MT II (MAR-1B-4) in dorsal view; (l) right MT V (MAR-1B-926/586-1) in dorsal view; (m) lumbar vertebra (MAR-1B-926/587-9) in cranial view.
The hippopotamus remains belonged to a skeletally immature individual with unfused heads of ribs and plates of vertebrae, unfused distal epiphysis of radius, and partially fused ilium, head, and distal epiphysis of femur, and proximal epiphysis of tibia (Figure 8). This stage of epiphyseal fusion is compatible with the slightly worn m2 and the minimally worn M3 (Figure 9a,b,d,e), which is equivalent to a mean age of 17 years (with a lifespan mean 43) of H. amphibius (group X [62]), indicating an early prime individual.
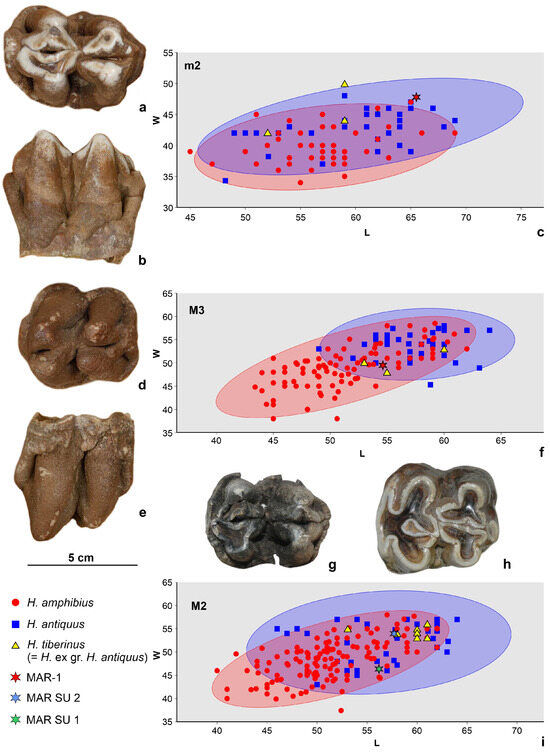
Figure 9.
(a,b) Left m2 (MAR-1B-8) of Hippopotamus antiquus from Marathousa 1 (Area B), most possibly belonging to the hippopotamus skeleton, in occlusal (a) and buccal (b) view. (c) Biplot comparing Hippopotamus m2 from various localities. (d,e) Left M3 (MAR-1B-7) of H. antiquus from Marathousa 1 (Area B), most possibly belonging to the hippopotamus skeleton, in occlusal (d) and buccal (e) view. (f) Biplot comparing Hippopotamus M3 from various localities. (g) Left M2 of H. antiquus from MAR SU 1 in occlusal view. (h) Left M2 of H. antiquus from MAR SU 2 in occlusal view. (i) Biplot comparing Hippopotamus M2 from various localities. Biplots with 95% confidence ellipses for Hippopotamus amphibius and H. antiquus, performed with the software package PAST v. 4.16 [63]. Data are from [33,64,65,66,67,68].
For the taxonomic attribution, in the absence of a skull, we focus our comparisons on the dimensions and proportions of the molars and of the more informative postcranial bones (Table S1). In the biplots for the m2 and M3 (Figure 9c,f), H. amphibius and H. antiquus are largely overlapping, and only in the case of M3 H. antiquus presents generally larger dimensions. The MAR-1 molars plot toward the spectrum of the larger dimensions. The m2 stands at the upper ranges of both species but within the 95% confidence ellipses of H. antiquus, whereas the M3 at the lower values of H. antiquus, and close to H. tiberinus. The postcranial bones are smaller than those of CHO-6 and H. antiquus in general, and for most skeletal elements, they are close to specimens from Kyparissia (KYP-1, KYP-4, and KYP-T; Figure 4). Despite being largely within the variation of H. amphibius, their best fit is H. tiberinus; yet, it should be noted again that the MAR-1 bones belonged to a skeletally immature individual and, therefore, in the case of long limb bones where the epiphyses were not completely fused (radius, femur, and tibia), the measurements are a slight underestimate of the expected ones if the epiphyses were fully fused.
Palaeoloxodon skeleton from Area B. Besides the elephant skeleton of Area A, a second partial skeleton was unearthed in the southern part of Area B in 2019, within the sedimentary unit UB4c (dark grey, massive organic, and intra-clast-rich silty sand, interpreted to represent subaerial-originated dilute mudflows and hyper-concentrated flows plunged into the lake margin [28]). The discovered skeletal elements include a tusk, a stylohyoid bone, three cervical vertebrae (including the axis), two ribs, and a distal fragment of the humerus (Figure 10). All these remains were found in close spatial proximity (Figure 10a,b) and possibly belonged to the same individual. The left tusk (MAR-1B-928/587-48) is completely preserved, as the pulp cavity is open and filled with sediment. It is long (length = 3.4 m), robust toward its proximal end (circumference = 560 mm, maximum diameters 176 × 171 mm; 56 cm from the base), and shows the typical slight curvature and weak torsion of Palaeoloxodon antiquus (Figure 10a; Table S1). The large dimensions of the tusk indicate most possibly a male individual, which agrees with the large size of the axis (height = 320 mm; comparable to the height of 333 mm of the male individual from Area A [45]).

Figure 10.
Palaeoloxodon antiquus skeleton from Marathousa 1 (Area B; UB4c). (a) Tusk and cervical vertebrae (including the axis) in situ. (b) Distribution map of the southwestern part of the trench at Area B showing the position (recorded with the use of a total station) of the elephant remains, stratigraphically and spatially associated with lithic artefacts and other faunal remains. (c,d) Axis (MAR-1B-928/588-39) in cranial (c) and right lateral (d) view; (e) cervical vertebra (MAR-1B-928/586-16) in cranial view; (f–h) left stylohyoid (MAR-1B-928/587-32) in medial (f), lateral (g), and anterior (h) view.
The reversed-Y-shaped left stylohyoid (MAR-1B- 928/587-32) is almost complete (Figure 10f–h; Table S1), missing only the ventral part of the inferior ramus. The distal ends of the superior and posterior rami are completely preserved, showing the typically rugged edges [69]. The superior ramus is elongate and slender, the posterior ramus is rather short and moderately flattened at its proximal part, and the inferior ramus is slender and shows a weak deflection (z = 8°). The angle between the inferior and posterior rami is narrow (α = 49° and β = 39°; measurements according to [70]). In these traits, the MAR-1 stylohyoid is different from those of the mammoth Mammuthus trogontherii (including that from Loussika [71]) and matches better with those of P. antiquus [70]. Within P. antiquus, the MAR-1 specimen fits best with the proportions shown in the stylohyoid from La Polledrara (MIS 9; Italy), rather than those of Neumark Nord 1 (MIS 5e; Germany), which generally have rather short and stout superior ramus, relatively longer posterior ramus, and a differently angled inferior ramus in regard to posterior ones [70]. To investigate whether these traits have evolutionary significance (and in turn, biochronological value) or represent intraspecific variability requires the discovery of additional specimens. However, the presence of this rare and fragile bone at MAR-1 not only highlights the exceptional preservation of the faunal assemblage at this site, but also enriches the available sample of this important bone for phylogenetic and taxonomic studies on proboscideans [72].
Whether the elephant remains that were found in previous years (proximal tibia, partial ribs and vertebrae, and several other bone fragments) belong to this individual is possible but not certain, especially because a second individual is present in Area B (and thus a third one for MAR-1, collectively), identified by a fragment of an axis (MAR-1B-923/586-1).
Other new faunal remains. Two hemimandibles, five diaphyseal/distal fragments of tibiae (Figure 11a–c; Table S1), and several other postcranial elements show morphology (e.g., lower incisors with smooth enamel on the outer side and triangular cross-section, and high-crowned cheek teeth with deep striids) and dimensions similar to the beaver Castor fiber [73,74], a species already reported from the locality [45]. A carnivoran fourth metatarsal from Area A (Figure 11s; Table S1) resembles in morphology and dimensions, those of a wolf-sized canid and is attributed to Canis sp., a taxon previously known only from Area B [45].

Figure 11.
Mammal remains from Marathousa 1. (a,b) Left hemimandible with the incisor and p4–m3 of Castor fiber (surface find at Area B) in medial (a) and dorsal (b) view. (c) Left hemimandible (MAR-1A-941/677-41) of Castor fiber in situ at Area A. (d,e) Second phalanx of the front limb (MAR-1B-930/593-46) of Bison sp. in dorsal (d) and lateral (e) view. (f,g) Third phalanx of the front limb (MAR-1B-931/594-42) of Bison sp. in lateral (f) and medial (g) view. (h) Refitting mandibular fragments (MAR-1B-934/594-71, 932/598-51, 933/595-60, 931/593-38, and 931/596-39) of the same individual of Dama sp., above the right hemimandible in medial view, and below the left hemimandible in lateral view. (i,j) Left scapula fragment (MAR-1B-934/597-35) of Dama sp. in lateral (i) and distal (j) view. (k) Left calcaneus (MAR-1B-928/588-21) of Dama sp. in plantar view. (l,m) Left scapula fragment (MAR-1A-935/672-20) of Cervus elaphus in lateral (l) and distal (m) view. (n) Distal fragment of left humerus (MAR-1B-931/564-58) of C. elaphus in cranial view. (o) Proximal fragment of left radius (MAR-1B-926-587-22) in dorsal view. (p) Distal fragment of left tibia (MAR-1B-925/586-7) in dorsal view. (q) Right astragalus (MAR-1B-926/586-4) of C. elaphus in dorsal view. (r) Left calcaneus (MAR-1-939/635-1) of C. elaphus in medial view. (s) Left MT IV (MAR-1A-941/672-44) of Canis sp. in dorsal view. Silhouette images from PhyloPic, phylopic.org.
In the cervid assemblage, two size groups are identified, the medium-sized one belonging to a fallow deer (Dama sp.) and the larger one to the red deer (Cervus elaphus). Additional material further enriched the specimens of both taxa. In Konidaris et al.’s study [45], a right hemimandible fragment of Dama sp. was described, which was found in two refitting pieces (MAR-1B-934/594-71 and 932/598-51). During the subsequent excavations, the left hemimandible bearing the p3–m3 of the same individual was also found (the wear of the teeth and the preservation are identical) in three pieces that again refit (MAR-1B-933/595-60, 931/593-38, and 931/596-39; Figure 11h). Moreover, to the previous published fallow deer material, we add here further dental specimens (P2, m2, and m3), a posterior part of a cranium preserving the occipital condyles, a sacrum, a distal fragment of a scapula, two calcanei, and a proximal phalanx (Figure 11i–k; Table S1). Additional cervid material that shows morphological and metrical traits compatible with an attribution to Cervus elaphus [46,75,76] includes isolated cheek teeth (dp3, p3, p4, and m1), a scapula, a distal humerus, a proximal radius, a distal tibia, two astragali, a calcaneus, and a second phalanx (Figure 11l–r; Table S1).
One second and one third phalanx (both of the front limb; Figure 11d–g; Table S1) belong to a bovine [77]. Their morphology and proportions allow for an attribution to Bison [44]. A specific attribution is not possible based on the available material, yet similar to the already known second phalanx of the hindlimb [45], an attribution to Bison schoetensacki is possible. Finally, a fragment of a fairly worn cheek tooth shows a bunodont pattern with plicated enamel, consistent with an attribution to Sus, a genus reported for the first time from the locality.
3.8. Marathousa 2 (MAR-2)
MAR-2 was identified during the targeted field survey of 2018, ca. 1.5 km east of MAR-1 and at a similar stratigraphic position (between LII and LIII; Figure 1d); therefore, a comparable age and a correlation to a glacial stage, ca. 450 ka (MIS 12), was suggested [24]. Fossils were collected from exposed sections in two defined areas, Area A and Area B, both yielding mainly hippopotamus bones. In particular, at Area A, the spatial association of the bones (vertebrae and ribs) and the consistency in size and ontogenetic age indicate the presence of a single, skeletally immature, hippopotamus individual (perhaps of close ontogenetic age with the MAR-1 hippopotamus that also has unfused vertebral plates). Additionally, a partial hemimandible of a fallow deer and a femur diaphysis of a beaver were also found. From Area B, isolated hippopotamus ribs and teeth were recovered, of which a large-sized canine permits the attribution to H. antiquus. The taphonomic study revealed the presence of cut marks in the spine of a thoracic vertebra of the hippopotamus from Area A, indicating butchering activities, further supported by the discovery of a spatially and stratigraphically associated lithic artefact. This represents one of the very few examples of hippopotami carcass processing during the Early–Middle Pleistocene of western Eurasia [24].
3.9. Tripotamos 4 (TRP-4)
TRP-4 (Figure 1b) was discovered during the field survey of 2020 in the southwestern margin of the mine, when lithic and faunal remains were found within a light-colored silty-sand layer positioned possibly above LIIIa, thus overlying MAR-1 (Figure 1d) [25]. The study of the large mammals is pending. The material consisted mostly of bone fragments and isolated teeth. Artiodactyls are represented by Dama sp. (hemimandible and other dental and postcranial remains), Cervus elaphus (a proximal part of an antler preserving two basal tines), and a bovine (several isolated, hypsodont, molars), while a canine fragment of small dimensions belongs to Hippopotamus (Figure 12a–g). The presence of an elephantid is confirmed by several lamellar fragments (Figure 12h).

Figure 12.
Mammal remains from Tripotamos 4 (TRP-4) and Choremi 7 (CHO-7). (a) Antler fragment (TRP-4-F2) of Cervus elaphus in lateral view; (b) distal metacarpal (TRP-4-F57) of Dama sp. in dorsal view; (c) distal metacarpal (TRP-4-F47) of Dama sp. in dorsal view; (d–f) lower molar (TRP-4-F2, surface) of Bovini in lingual (d), occlusal (e), and buccal (f) view; (g) fragment of upper canine (TRP-4-F70) of Hippopotamus sp.; (h) lamellar fragment (TRP-4-F36, F44, F58, and F73) of Elephantidae indet.; (i–k) right hemimandible fragment with m1–m3 (CHO-7-F125) of Cervus elaphus in lateral (i), lingual (j), and occlusal (k) view; (l–n) right m3 (CHO-7-F147) of Bos sp. in occlusal (l), lingual (m), and buccal (n); (o–q) upper molar (CHO-7-F68) of Bos sp. in occlusal (o), buccal (p), and lingual (q) view.
3.10. Choremi 7 (CHO-7)
CHO-7 was discovered during the field survey of 2021 in the southeastern margin of the mine (Figure 1b) and yielded both faunal and cultural material. CHO-7 stands at the highest stratigraphic position of all investigated sites, located above the entire lignite sequence of the Marathousa Mb (above the LIIIc; Figure 1d), just below the overlying Megalopolis Mb; therefore, it comprises the youngest of the investigated sites [25].
The systematic sampling for micromammals from the different layers of the site provided abundant specimens, and the first results of their study are presented herein. The assemblage is dominated by the genus Microtus. The occlusal surface of the m1 is characterized by five closed triangles and an arvalis-type mesial cap (Figure 5l). The morphology is different than Microtus agrestis and indicates the presence of Microtus arvalis. In the M2 of the former species, there is an additional closed loop at the distal end of the tooth [78], and this loop is not observed in any of the studied specimens. Additionally, based on the La/Li index, i.e., T4 width/T5 width [32], the two species (M. arvalis/M. agrestis) can be distinguished, except for an overlap in the larger values of M. agrestis and the lower ones of M. arvalis. In the CHO-7 specimens, the T4/T5 index (62.3–101.4, mean = 81.9; n = 17) falls within the range of M. arvalis or higher. Because the dentition of M. arvalis and Microtus rossiameridionalis is extremely similar [79], making their distinction impossible, we assign the CHO-7 material to Microtus sp. (cf. M. arvalis). Moreover, some Microtus molars show confluent T4 and T5 triangles (Figure 5m), and their dimensions are within the range of Microtus (Terricola) subterraneus; as such, they are ascribed to this species. Furthermore, the CHO-7 rodent assemblage includes three complete and five incomplete rootless m1s with three closed triangles (Figure 5j,k). The enamel of the mesial and distal sides of each triangle differs. The length of the complete specimens (mean = 2.71 mm; n = 33), the SDQ (Schmelzband–Differenzierung Quotient [80]) values (114.5–133.3, mean = 120.0; n = 7), and the shape of their occlusal surface permit their attribution to Arvicola mosbachensis. Finally, some murid molars that are similar to Apodemus, and some soricid ones, have not been studied yet.
The study of macro-mammals is pending. The material consists predominantly of bone fragments, which show clear evidence of rounding of their broken edges, and polishing and abrasive marks of their cortical surfaces, indicating substantial transport by water. Three cervid taxa are present at CHO-7 (following morphological and metrical data in [46,75], and comparison with corresponding specimens from other Megalopolis sites): a Capreolus-sized (molars, phalanges, and distal tibia), Dama sp. (upper molar, phalanges, and distal humerus), and Cervus elaphus (hemimandible, molars, and tibia; Figure 12i–k; Table S1). Several isolated, hypsodont molars are morphologically compatible (following [44]) with an attribution to Bos (Figure 12l–q).
3.11. Selected Isolated Finds
Two proboscidean large tusk fragments were discovered, one from the Marathousa mine (above LII) and the other from the Choremi mine (below LIII; Figure 13a,b). Both show obtuse Schreger angles in their cross-sections and, therefore, can be ascribed to Palaeoloxodon antiquus. Several other small tusk fragments collected from various findspots in the Megalopolis Basin also show the same Schreger pattern, also permitting the attribution to this species.

Figure 13.
(a,b) Tusk fragment of Palaeoloxodon antiquus from CHO SU 11 in cross-sectional (a) and lateral (b) view. (c–e) Left upper molar of cf. Bos from Choremi 5 in occlusal (c), lingual (d), and buccal (e) view. (f) Distal metacarpal of Bos sp. from CHO SU 11 in dorsal view. (g,h) Right upper premolar (P3/P4) of Equus sp. from CHO SU 10, close to the site Choremi 6, in lingual (g) and occlusal (h) view. (i) Left astragalus of Equus sp. from MAR SU North in dorsal view. (j) Left astragalus of Equus sp. from CHO SU 7 in dorsal view. (k) Left radius and ulna of Hippopotamus antiquus from the Marathousa mine in lateral view.
Five isolated horse (Equus) specimens were discovered during the field surveys and are considered important, because horses constitute a rare faunal element in the Megalopolis Basin ([9,19,42] and a study in progress). An upper premolar (P3/P4) was identified close to CHO-6, at the same stratigraphic level as the site, thus preceding the Matuyama/Brunhes boundary (Figure 13g,h). Two more specimens were found in the southern–southwestern part of the Choremi mine within clastic sediments below LIIa, an astragalus, collected at CHO SU 7 (Figure 13j), and a proximal phalanx, collected during the sampling for magnetostratigraphy at the level of sample K7 [3] (Figure 4). Two more astragali were found within MAR SU 8 (below LII) and MAR SU North (surface; Figure 13i). Interestingly, all the newly collected horse material (including the few specimens from KYP-4) originated from the lower parts of the stratigraphic sequence.
Some more hippopotamus remains were collected during the field survey of 2013. These include a stratified M2 (MAR SU 1; Figure 9g) and some surface-collected finds (MAR SU 2) that comprise an M2 (Figure 9h), a navicular, and an astragalus. Both M2 specimens fall in the overlap region of H. amphibius and H. antiquus in their crown dimensions (Figure 9i). The dimensions of both the navicular and the astragalus are smaller than that of CHO-6 and very close to the MAR-1B hippopotamus skeleton (Figure 4). An additional surface-collected find is a complete radio-ulna (Figure 13k) from the lower parts of the sequence in the Marathousa mine, possibly originating from clastic sediments below LII.
Finally, some more Bovini remains were found in the Choremi mine. The oldest ones originated from the site Choremi 5 (CHO-5, Figure 1b), discovered during the field survey of 2018. The find horizon is located between two lignite seams, possibly LIIa and LIIb, and thus might correspond to MIS 14 (Figure 1d) [3]. From this layer, the collected Bovini remains include a lower premolar, an upper molar, a proximal phalanx, and a sesamoid. The morphology of the upper molar (Figure 13c–e; following [44]) fit better with an attribution to an aurochs (Bos primigenius) rather than to a bison. However, the material is very limited for a definite assignment and, therefore, the molar is attributed to cf. Bos. Additionally, a distal metacarpal fragment of a Bovini (Figure 13f) was collected within CHO SU 11 toward the upper the part of the Marathousa Mb sequence of the Choremi mine. The absence of marked supra-articular tubercles (so that the greatest distal width is measured across the trochleae [44]) allow for an attribution to Bos sp.
4. Discussion
4.1. Biostratigraphy–Biochronology
The updated faunal lists of the Megalopolis Basin sites and survey units are presented in Table S2. Most of the large mammals present in the Megalopolis Basin have a wide biostratigraphic/biochronological range within the Galerian and Aurelian European Land Mammal Ages, if not appearing even earlier, during the late Villafranchian–Epivillafranchian. As such, it is the study of the micromammals that offers the most important biochronological information that contributes to the age constraints of the sites and allows for the identification and dating of the currently known First and Last Greek Occurrence (FGO and LGO), as well as for the First and Last Megalopolis Occurrence (FMO and LMO) of large mammals.
The data from the level of CHO-6 are limited, yet based on the local stratigraphy, its dating at the end of the Lower Pleistocene renders CHO-6 as the oldest large mammal site of the Choremi Fm and marks the FMO of Hippopotamus antiquus and Equus (Figure 14).
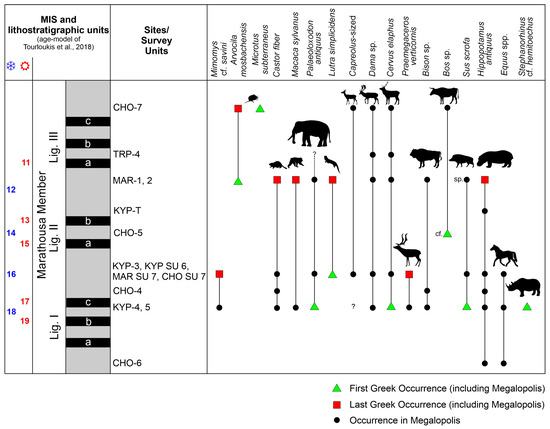
Figure 14.
Simplified stratigraphic column of the Marathousa Member showing the lignite seams and the intercalating detrital intervals, their correlation to the Marine Isotope Stages (MIS) following the age model proposed by Tourloukis et al. [3], the stratigraphic position of the sites and survey units following Karkanas et al. [25,28], and the biostratigraphic range of selected micro- and macro-mammals from the Megalopolis Basin, indicating their First and Last Occurrences in the Greek fossil record. Silhouette images from PhyloPic, phylopic.org.
The available data for KYP-4 are more informative. The presence of Microtus (Microtus) indicates that the fauna post-dates the Allophaiomys/Microtus transition that took place close to the Early/Middle Pleistocene transition (780 ka). The occurrence of Mimomys savini is also biochronologically important. The Mimomys/Arvicola transition was roughly synchronous throughout Europe and took place around MIS 15 [81,82,83]. Transitional populations showing primitive Arvicola traits were identified at Isernia (Italy, ca. MIS15/MIS14 [84]) and Mauer (Germany, MIS 15, ca. 600 ka [85]). Therefore, the presence of Mimomys at KYP-4 indicates an age older than 600–650 ka. Perhaps the most informative taxon at KYP-4 is the smaller-sized Mimomys, a taxon which is found only in a limited number of western Eurasian sites (identified as Mimomys pusillus or Mimomys sp.) dated to or predating MIS 16. These include the Choremiou Section of the Megalopolis Basin [2], Kärlich in Germany [86], Pakefield in the UK [87], and Chiu-Atasova in Russia (Southern Urals [88]). As a whole, the micromammal assemblage of KYP-4 indicates an early Middle Pleistocene age, after 780 ka and before MIS 16, in agreement with the stratigraphic placement of the site inside the lowermost lignite (LI).
This age for KYP-4 marks the FMO and FGO for several of the large mammals (Figure 14). These include Homo (based on the presence of lithic artefacts), Palaeoloxodon antiquus, Cervus elaphus, Sus scrofa, and perhaps, Stephanorhinus hemitoechus, marking their currently known FGO in the early Middle Pleistocene, while depending on the followed taxonomy, this period may also mark the FGO of Praemegaceros verticornis.
The earliest palaeoloxodont elephants outside of Africa are documented at Gesher Benot Ya’aqov at ca. 780 ka (MIS 19, Israel) [89,90], while their arrival in Europe is documented with P. antiquus at Slivia (Slivia faunal unit) in Italy, estimated to be slightly older than the Early/Middle Pleistocene boundary based on micromammals [91]. Palaeoloxodon is present in Croatia in sediments dated between 940 and 470 ka [91,92]; therefore, the KYP-4 record is the secure first known occurrence of palaeoloxodont elephants is SE Europe.
The red deer remains are scarce and fragmentary at KYP-4, yet from the roughly same level of KYP-5 (slightly to the north of KYP-4; Figure 1b), the presence of Cervus elaphus is secure based on a large-sized antler, showing the characteristic adjacent disposition of brow and bez tines [42]. Cervus elaphus is already known from Epivillafranchian localities of Europe, dated between 1.0 and 0.8 Ma, but in Greece, the KYP-4, 5 red deer record is the earliest one identified so far.
The replacement of Sus strozzii by Sus scrofa took place at or soon after the Early/Middle Pleistocene transition [93], and the KYP-4 record is one of the earliest known occurrences of the species in Europe, biochronologically close perhaps to the record from Petralona Cave [94]. Accordingly, if Stephanorhinus hemitoechus is indeed present at KYP-4, it marks its FGO, again perhaps close to the Petralona occurrence [94].
The uncertain taxonomic status of the various Praemegaceros species/subspecies and the validity of several distinctive morphological traits makes the earliest occurrence of Praemegaceros verticornis in Europe problematic. Nonetheless, the rich KYP-4 material, including antlers with distal palmation, is more advanced than the Early Pleistocene Praemegaceros material from Greece, commonly attributed to Praemegaceros pliotarantoindes [19,42,95]. Praemegaceros verticornis is thought to have existed in Europe until ca. 400 ka [96], yet it is not present at MAR-1 (dated at ca. 450 ka). Its absence at MAR-1 and MAR-2, if not accidental, may be due to paleoecological reasons, or to earlier local extinction [19].
Additionally, Castor fiber, Macaca sylvanus, Dama sp., and Bison sp. document their FMO (Figure 14).
As in the case of the micromammals at KYP-4, the presence at the level of KYP-3/KYP SU 6 of both a larger Mimomys (M. cf. savini) and a smaller one (Mimomys sp.) indicates a correlation to or before MIS 16, and certainly older than the Arvicola-bearing MAR-1. The stratigraphic data suggest that KYP-3 overlies KYP-4, and following the age model of Tourloukis et al. [3], a correlation to MIS 16 is possible. This marks the LGO of Mimomys (Figure 14). As a whole, the level of KYP-3/KYP SU 6/MAR SU 7/CHO SU 7 marks the so far known LGO of Praemegaceros. The genus is present at Petralona [97], yet of unknown age. In addition, Sus scrofa and Equus record their LMO, at least a dated one, since both taxa were recorded by Melentis [9,10] and Sickenberg [17]. On the other hand, this level marks the FGO of Lutra simplicidens and of a Capreolus-sized cervid (Figure 14), the latter documented from unknown horizons [11,17,19].
At CHO-5 (likely MIS 14, based on the basin stratigraphy) the FGO of Bos is possibly recorded (Figure 14), a genus that is otherwise present at higher stratigraphic positions in the basin (e.g., CHO-7 and CHO SU 11) and was also documented by Melentis [13] but from unknown horizons.
The biostratigraphy/biochronology of the mammal fauna from MAR-1 and MAR-2 has been already discussed [24,45,52]. Of particular interest is Arvicola mosbachensis from MAR-1, whose molars’ SDQ values and size suggest a correlation to MIS 12/11 [52]. This is also the so far known FGO of Arvicola, although its slightly advanced stage might indicate that an earlier occurrence of more primitive Arvicola in lower stratigraphic layers of the basin is possible. In terms of large mammals, the levels of MAR-1 and 2 mark the LGO of Castor fiber, Macaca sylvanus, Lutra simplicidens, and Hippopotamus antiquus (Figure 14). Particularly, the MAR-1 and 2 hippopotamus record corresponds to one of the last occurrences of H. antiquus in Europe at ca. 450 ka (see also [98]). The last occurrences of L. simplicidens in Europe are reported at Hundsheim (Austria) and Mosbach-2 (Germany), both correlated to MIS 15 or MIS 13 (ca. 600–500 ka), while the modern Lutra lutra is reported from Hoxne, England, at MIS 11 (ca. 400 ka) [99,100] (note, however, that the Hoxne lutrine is regarded as Lutra sp. in [101]). As such, the MAR-1 record during the MIS 12 may document one of the last occurrences of L. simplicidens in Europe.
The rich rodent assemblage from CHO-7 is important in terms of biochronology. The earliest occurrence of Microtus (Terricola) subterraneus in Central Europe is recorded at Maastricht-Belvédère 4 (MIS 7, The Netherlands) [102,103], although a slightly older age (MIS 8) for the earliest occurrence of the species is also possible [104] (Figure 2). This indicates that CHO-7 is significantly younger than MAR-1. The presence of Microtus sp. (cf. M. arvalis) and Arvicola mosbachensis is common between CHO-7 and MAR-1. The mean SDQ values of the Arvicola molars are also comparable. Yet, the available molars of Arvicola from CHO-7 are limited and, as such, the mean SDQ value should be considered only as indicative.
In a recent study, Konidaris and Kostopoulos [105] introduced the Late Pliocene to Middle Pleistocene large mammal faunal units (FUs) of Greece, which are defined by a set of First and Last Occurrences. In this biochronological unit scheme, they recognized the distinct Marathousa FU, dated between <800 and ca. 450 ka, which encompasses the Galerian faunal assemblages from the Megalopolis Basin and the Petralona Cave (the older faunal assemblage) in Chalkidiki Peninsula (e.g., [94,97]). Marathousa FU records the FGO of Homo, Palaeoloxodon antiquus, Lutra simplicidens, Ursus deningeri, Panthera spelaea, Crocuta crocuta, “Hyaena” prisca”, Stephanorhinus hemitoechus, Bison, Sus scrofa, the Cervus elaphus lineage, and possibly Bos, while additional large mammals may also include Panthera pardus (if present at KYP-3) and Mammuthus trogontherii from Loussika (Peloponnese) and other localities [71]. On the other hand, this FU marks the LGO of Macaca sylvanus, Equus altidens, Hippopotamus antiquus, Praemegaceros, and Lutra simplicidens [105]. In a wider European context, the Marathousa FU represents a unit within Isernia and Fontana Ranuccio FUs of Italy, and corresponds to Gran Dolina TD8, Sima de los Huesos (Spain), and L’Escale and Arago CM (France) faunal assemblages (e.g., [91], and references therein). Marathousa FU is part of the major large mammal reorganization that took place close to the Early/Middle Pleistocene transition and is associated with the “Mid-Pleistocene Revolution” that affected the terrestrial ecosystems (e.g., [91]). The faunal turnover documents the disappearance of several Villafranchian–Epivillafranchian taxa (whose last occurrences in Greece are recorded in the Apollonia FU) and the arrival of several Galerian immigrants (Marathousa FU). The Marathousa FU is followed by the Apidima FU, which is characterized by the establishment of modern large mammals from the end of the Middle Pleistocene onwards [105,106].
4.2. Hippopotamuses in the Megalopolis Basin
In the last few years, several studies dealing with the status of the Middle Pleistocene hippopotamuses from Europe were published (e.g., [24,67,107,108,109,110]). Most of these studies underline that in the absence of the diagnostic cranium and mandible from securely dated contexts, the taxonomy (largely focused on postcranial dimensions) and the time of replacement of the two or three commonly recognized species remain vague. Despite this, and some different opinions among the researchers, there is a broad consensus that H. antiquus or closely related forms classified under H. ex gr. antiquus were replaced by H. amphibius broadly between 500 and 400 ka [111].
The hippopotamus material from the Megalopolis Basin does not include adequately preserved cranial material, yet the quite abundant dental and postcranial specimens (in some cases belonging to partial skeletons), with secure chronological placement and from different stratigraphic levels covering the late Early–Middle Pleistocene, are important and contribute to the current investigations and discussions.
The study of Athanassiou et al. [20] on the hippopotamuses from the early Middle Pleistocene sites of the Kyparissia mine (KYP-1, KYP-3, KYP-4, and KYP-T) showed their intermediate position in terms of bone dimensions between H. amphibius and H. antiquus, and a better match with specimens classified as H. tiberinus. The present study confirms that the hippopotamuses from other Middle Pleistocene deposits of the basin (CHO-4, MAR SU 2, and MAR-1; a larger-sized third metacarpal from MAR-1 Area A [45] is of comparable size to a recently discovered third metacarpal from KYP-4) are also consistently placed at the lower range of H. antiquus, reflected also in the estimated body masses (Figure 4; Table 1 and Table S1; note that the MAR-1 individual is skeletally immature). The skeletal elements (and the estimated body mass) of the CHO-6 hippopotamus indicate a large-sized individual, larger than the other hippopotamuses from Megalopolis and as large as those of “typical” H. antiquus [40], in agreement with the older (Lower Pleistocene) age of the find-bearing sediment.
Therefore, in terms of taxonomy, either the middle-sized H. tiberinus is a distinct species (according to [112] after 1.4–1.2 Ma; pending confirmation from cranial remains), implying a second dispersal of African hippopotamuses to Europe (related perhaps to Hippopotamus gorgops [33,112]), or the smaller dimensions of the Middle Pleistocene hippopotamuses correspond to a body size decrease of the single Early Pleistocene arrival of hippopotamuses in Europe (between ca. 2.1 and 1.6 Ma; H. antiquus) as a response to climatic conditions [43]. Considering that, at present, there exists no clear-cut morphological distinction between H. tiberinus and H. antiquus (e.g., [67,113]) that would support the first option, and the fact that the studied Hippopotamus remains from Megalopolis Basin originated from glacial (or stadial) sediments with supposedly harsher paleoenvironmental conditions (see also below), the second option is preferred for the moment (besides hippopotamuses, the potential size reduction of other large mammals in the Megalopolis Basin requires further investigation). This is also in agreement with the study on dental enamel hypoplasia (related to physiological stress) in hippopotamus incisors and canines from Early Pleistocene sites of Europe [114]. In this study, a high incidence of hypoplasia was observed at sites dated to ca. 900–860 ka when a long and severe glacial phase was recorded.
Therefore, the Middle Pleistocene hippopotamuses from the Megalopolis Basin are attributed to the smaller-sized representatives of H. antiquus (the H. ex gr. H. antiquus of [43]). The even smaller-sized canine from TRP-4 (younger than MAR-1 and 2) might hint at an attribution to H. amphibius, but the evidence is too fragmentary and, therefore, is ascribed to Hippopotamus sp.
4.3. Paleoenrvironmental Remarks
Palaeoloxodon antiquus had wide ecological adaptations, inhabiting a broad spectrum of environments and having flexible dietary preferences and foraging behavior (e.g., [115,116,117]). Despite this, in central and northern Europe, the species existed predominantly during the warmer interglacial stages and was generally absent from the intervening cold stages of open conditions, when its geographic range was restricted in refugia of peri-Mediterranean Europe (e.g., [118,119]). The isotopic study on the elephant individual from the Area A of MAR-1 (i.e., during the glacial MIS 12) corroborated the refugial status of Greece and the Megalopolis Basin in particular, by finding a restricted geographical range within a C3-dominated open woodland environment under relatively stable climatic conditions [120]. This conclusion is further supported by the continuous presence of straight-tusked elephants in several stratigraphic levels of the Megalopolis Basin, corresponding to both interglacial and glacial, which further highlights the adaptability of the species to survive under diverse climatic conditions and to inhabit a variety of environments. The refugial status of the region is additionally attested from the also temperate-adapted fallow deer (present in several sites/findspots) and macaques (the latter present at both KYP-4 and MAR-1) that prefer settings with milder conditions (e.g., [121,122]), and especially the former is found in several stratigraphic layers of the Megalopolis Basin.
Although not exclusively linked with a warm/temperate environment, the presence during glacial stages of beavers, hippopotamuses, and otters, which are highly dependent on permanent freshwater bodies for their survival (e.g., [43]), indicates the capacity of the basin to retain freshwater bodies throughout the annual cycle, even under the harsher climatic conditions [54]. Such an environment is also supported from the presence of two freshwater turtles, the European pond turtle Emys orbicularis and the Balkan terrapin Mauremys rivulata, which further require high enough average summer temperatures that allow for sexual differentiation ([60]; see also [123] regarding the refugial status of Greece for E. orbicularis).
The presence of warmer-adapted floral and faunal taxa, for instance, Palmae and the darter (Anhinga; present at both MAR-1 and KYP-T), a bird that is not found today in Greece and is associated with warm/tropical regions, indicate, together with the other botanical material (aquatic, waterside, and damp ground vegetation, combined with willow, alder, elm, deciduous oak, and maple) and avian remains (ducks, geese, and swans), a warm-temperate environment [20,56,58]. On the other hand, cooler conditions than today’s Greece are inferred for MAR-1 by the multiproxy paleoenvironmental study including ostracods that indicate mean summer temperatures of 10–15 °C [54], as well as for MAR-2 by the mollusk study [24]; yet, in both cases, temperatures allowed the existence of water bodies under non-freezing conditions throughout the year.
For KYP-4, most paleoenvironmental analyses are currently in progress. The large mammal fauna generally shows similarities with MAR-1 in the occurrence of elephants, hippopotamuses, macaques, deer, and beavers. However, the presence at KYP-4 of the giant deer Praemegaceros verticornis (abundant in the faunal assemblage), rhinoceroses, and horses [19,42] might indicate slightly more open conditions compared to MAR-1. This stands in agreement with the preliminary results from the mollusk fauna from KYP-4, which show a slight increase in the terrestrial/semi-terrestrial taxa [124] as well as with the presence of the (terrestrial) tortoise Testudo marginata in the wider Kyparissia region (e.g., at KYP-3), a species absent in the particularly abundant (freshwater) turtle assemblage of MAR-1 [20,60]. The ostracod analysis for KYP-4 indicates the presence of rather cold (constantly lower than 15 °C) carbonate springs and a relatively stable temperature throughout the year [125].
Overall, despite the evidence of colder conditions than present-day Greece, the presence/abundance of taxa adapted to milder climatic conditions (elephants, hippopotamuses, macaques, and fallow deer), and some even to warmer ones (Palmae and the darter), indicate that the prevailing climate at MAR-1, and possibly at the other Megalopolis sites (at least at MAR-2, KYP-3, and KYP-4), was temperate, even during cold—oxygen isotopic—stages, while both flora and fauna support the existence of a landscape with substantial woodland components and more open areas, close to permanent and large freshwater bodies. The presence of taxa generally indicative of milder conditions in different stratigraphic layers of the basin, including detrital deposits (corresponding to the cold, glacial/stadial stages), suggests their continuous presence and a refugium region for their populations.
5. Conclusions
The late Early to Middle Pleistocene localities of the Megalopolis Basin permit the identification and tracking of faunal and evolutionary changes within the stratigraphic succession of the same sedimentary basin, thus allowing the study of the taxonomy and evolution of several vertebrate taxa and mammal turnovers between ca. 900 and 300 ka. Additionally, they provide paleoenvironmental (e.g., the refugial status of the basin for fauna and flora during glacial stages, dietary preferences of mammals, and seasonal climatic variations) and taphonomic interpretations (e.g., carnivore and anthropogenic modifications of mammal bones and particularly megafauna, site formation processes) that offer valuable insights into the Middle Pleistocene terrestrial ecosystems of Europe and hominin adaptations in particular, thus comprising a unique record for SE Europe. Particularly for Greece, the Megalopolis Basin localities, together with the Lower Pleistocene ones, e.g., Dafnero and Mygdonia Basin, and the Middle–Upper Pleistocene caves Petralona and Apidima, all of which are characterized by long stratigraphic sequences, altogether represent a rather continuous faunal sequence that captures the entire Pleistocene and allows for the detailed study of the faunal changes and the paleoenvironmental record of Greece.
Ongoing dating (e.g., electron spin resonance (ESR) and post-infrared infrared stimulated luminescence (pIRIR)), paleontological, paleoenvironmental (e.g., pollen, ostracods, mollusks, and stable isotope biogeochemistry, and dental micro- and meso-wear analyses in the mammal remains), and archaeological research in the Megalopolis Basin will refine the age constraints of the localities and establish a secure paleoecological and cultural framework, leading to more accurate interpretations for the first occurrence of hominins and other taxa in the southern-most tip of the Balkans.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/quat7040041/s1. Table S1: Dental and postcranial measurements (in mm) of new large mammal fossils from the Megalopolis Basin. Table S2: Updated faunal lists of the Megalopolis Basin sites and survey units. Data from [19,20,24,41,42,45,52,57] and the present study.
Author Contributions
Conceptualization, G.E.K.; methodology, G.E.K., A.A. and T.v.K.; validation, G.E.K., A.A., V.T., K.C., T.v.K., D.G., N.T., G.T., E.R., E.P., P.K. and K.H.; formal analysis, G.E.K., A.A., K.C. and T.v.K.; data curation, G.E.K. and A.A.; writing—original draft preparation, G.E.K. and K.C.; writing—review and editing, G.E.K., A.A., V.T., K.C., T.v.K., D.G., N.T., G.T., E.R., E.P., P.K. and K.H.; visualization, G.E.K. and K.C.; project administration, K.H., T.v.K. and E.P.; funding acquisition, K.H. All authors have read and agreed to the published version of the manuscript.
Funding
Excavation at Marathousa 1 was supported by the ERC-StG-283503 (“PaGE”) and the ERC-CoG-724703 (“CROSSROADS”), both awarded to K.H. The 2018–2022 field survey in the Megalopolis Basin was supported by the ERC-CoG-724703 (“CROSSROADS”). G.E.K., V.T., and K.H. were supported by the Deutsche Forschungsgemeinschaft (DFG Project No. 463225251; “MEGALOPOLIS”). K.C., E.R., and K.H. were supported by the ERC-AdG-101019659 (“FIRSTSTEPS”).
Data Availability Statement
Data are contained within the article and the Supplementary Materials.
Acknowledgments
Excavation at Marathousa 1 was conducted under a permit granted to the Ephorate of Paleoanthropology–Speleology, Hellenic Ministry of Culture. The 2018–2022 field survey in the Megalopolis Basin was conducted in the framework of the Megalopolis Paleoenvironmental Project (MegaPal) under a permit granted to the Ephorate of Paleoanthropology–Speleology, Hellenic Ministry of Culture, and the American School of Classical Studies at Athens. Many thanks to all field team members for their contribution in the fieldwork, the preparation of material, and the sieving and sorting of micromammals, particularly, A. Varis, C. Siali, M. Psyrri, G. Boni, N. Nikolakopoulou, and K. Kouveli. We greatly thank D. Kostopoulos for his valuable help with the bovine material from Megalopolis. Many thanks to the academic editors and the four reviewers for providing constructive comments and suggestions that improved the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Vinken, R. Stratigraphie und Tektonik des Beckens von Megalopolis (Peloponnes, Griechenland). Geol. Jahrb. 1965, 83, 97–148. [Google Scholar]
- van Vugt, N. Orbital Forcing in Late Neogene Lacustrine Basins from the Mediterranean. A Magnetostratigraphic and Cyclostratigraphic Study. Ph.D. Thesis, Utrecht University, Utrecht, The Netherland, 2000. [Google Scholar]
- Tourloukis, V.; Muttoni, G.; Karkanas, P.; Monesi, E.; Scardia, G.; Panagopoulou, E.; Harvati, K. Magnetostratigraphic and chronostratigraphic constraints on the Marathousa 1 Lower Palaeolithic site and the Middle Pleistocene deposits of the Megalopolis basin, Greece. Quat. Int. 2018, 497, 154–169. [Google Scholar] [CrossRef]
- Melentis, J.K. Die Dentition der pleistozänen Proboscidier des Beckens von Megalopolis im Peloponnes (Griechenland). Ann. Géologiques Pays Helléniques 1961, 12, 153–262. [Google Scholar]
- Darlas, A. Palaeolithic finds from the Megalopolis Basin: Their relation to the fossils of the same area. In Argonautis: Honorary Issue to C.G. Doumas; Vlachopoulos, A., Birtaha, K., Eds.; Antiquity: Kathimerini, Athens, 2003; pp. 27–37, (In Greek with English Summary). [Google Scholar]
- Bürchner, L. Wichtige Funde fossiler Knochen in Arkadien. Berichte Naturwissenschaftlichen Vereines Regensbg. 1903, 9, 119–123. [Google Scholar]
- Skouphos, T. Über die paläontologischen Ausgrabungen in Griechenland in Beziehung auf das Vorhandensein des Menschen. In Proceedings of the Comptes Rendus du Congrès International d’Archéologie, 1re Session, Athens, Greece, 7–13 April 1905; pp. 231–236. [Google Scholar]
- Melentis, J.K. Die Osteologie der pleistozänen Proboscidier des Beckens von Megalopolis im Peloponnes (Griechenland). Ann. Géologiques Pays Helléniques 1963, 14, 1–107. [Google Scholar]
- Melentis, J.K. Über Equus abeli aus dem Mittelpleistozän des Beckens von Megalopolis im Peloponnes (Griechenland). Ann. Géologiques Pays Helléniques 1963, 16, 507–519. [Google Scholar]
- Melentis, J.K. Sus scrofa L. aus dem Jungpleistozän des Beckens von Megalopolis im Peloponnes (Griechenland). Ann. Géologiques Pays Helléniques 1965, 16, 436–445. [Google Scholar]
- Melentis, J.K. Die pleistozänen Cerviden des Beckens von Megalopolis im Peloponnes (Griechenland). Ann. Géologiques Pays Helléniques 1965, 16, 1–92. [Google Scholar]
- Melentis, J.K. Über Hippopotamus antiquus Desmarest aus dem Mittelpleistozän des Beckens von Megalopolis im Peloponnes (Griechenland). Ann. Géologiques Pays Helléniques 1965, 16, 403–435. [Google Scholar]
- Melentis, J.K. Die Boviden des Jungpleistozäns des Beckens von Megalopolis im Peloponnes (Griechenland). Ann. Géologiques Pays Helléniques 1965, 16, 446–472. [Google Scholar]
- Melentis, J.K. Die pleistozänen Nashörner des Beckens von Megalopolis im Peloponnes (Griechenland). Ann. Géologiques Pays Helléniques 1965, 16, 363–402. [Google Scholar]
- Melentis, J.K. Clemmys caspica aus dem Pleistozän des Beckens von Megalopolis im Peleponnes (Griechenland). Ann. Géologiques Pays Helléniques 1966, 17, 169–181. [Google Scholar]
- Melentis, J.K. Erster Fund von Palaeoloxodon antiquus germanicus in den jungpleistozänen Ablagerungen des Beckens von Megalopolis (Peloponnes). Prakt. Akad. Athin. 1965, 40, 197–207. [Google Scholar]
- Sickenberg, O. Eine Säugetierfauna des tieferen Bihariums aus dem Becken von Megalopolis (Peloponnes, Griechenland). Ann. Géologiques Pays Helléniques 1976, 27, 25–73. [Google Scholar]
- Theodorou, G. Megalopolis—112 years after the first excavation by National and Kapodistrian University of Athens (NKUA)—And the Post Lignite era. In Proceedings of the VIth International Conference on Mammoths and their Relatives, Grevena-Siatista, Greece, 5–12 May 2014; pp. 195–196. [Google Scholar]
- Athanassiou, A. Pleistocene vertebrates from the Kyparíssia lignite mine, Megalopolis Basin, S. Greece: Rodentia, Carnivora, Proboscidea, Perissodactyla, Ruminantia. Quat. Int. 2018, 497, 198–221. [Google Scholar] [CrossRef]
- Athanassiou, A.; Michailidis, D.; Vlachos, E.; Tourloukis, V.; Thompson, N.; Harvati, K. Pleistocene vertebrates from the Kyparíssia lignite mine, Megalopolis Basin, S. Greece: Testudines, Aves, Suiformes. Quat. Int. 2018, 497, 178–197. [Google Scholar] [CrossRef]
- Röding, C.; Zastrow, J.; Scherf, H.; Doukas, C.; Harvati, K. Crown outline analysis of the hominin upper third molar from the Megalopolis Basin, Peloponnese, Greece. In Ancient Connections in Eurasia; Reyes-Centeno, H., Harvati, K., Eds.; Kerns Verlag: Tübingen, Germany, 2021; pp. 13–36. [Google Scholar]
- Thompson, N.; Tourloukis, V.; Panagopoulou, E.; Harvati, K. In search of Pleistocene remains at the Gates of Europe: Directed surface survey of the Megalopolis Basin (Greece). Quat. Int. 2018, 497, 22–32. [Google Scholar] [CrossRef]
- Panagopoulou, E.; Tourloukis, V.; Thompson, N.; Konidaris, G.; Athanassiou, A.; Giusti, D.; Tsartsidou, G.; Karkanas, P.; Harvati, K. The Lower Palaeolithic site of Marathousa 1, Megalopolis, Greece: Overview of the evidence. Quat. Int. 2018, 497, 33–46. [Google Scholar] [CrossRef]
- Konidaris, G.E.; Tourloukis, V.; Boni, G.; Athanassiou, A.; Giusti, D.; Thompson, N.; Syrides, G.; Panagopoulou, E.; Karkanas, T.; Harvati, K. Marathousa 2: A new Middle Pleistocene locality in the Megalopolis Basin (Greece) with evidence of hominin exploitation of megafauna (Hippopotamus). PaleoAnthropology 2023, 2023, 34–55. [Google Scholar]
- Karkanas, P.; Tourloukis, V.; Thompson, N.; Giusti, D.; Tsartsidou, G.; Athanassiou, A.; Konidaris, G.; Roditi, E.; Panagopoulou, E.; Harvati, K. The Megalopolis Paleoenvironmental Project (MegaPal). In Human Evolution at the CROSSROADS: Research in Greece and Beyond; Harvati, Κ., Ioannidou, Μ., Eds.; Tübingen University Press: Tübingen, Germany, 2024. [Google Scholar]
- Nickel, B.; Riegel, W.; Schönherr, T.; Velitzelos, E. Environments of coal formation in the Pleistocene lignite at Megalopolis, Peloponnesus (Greece)—Reconstructions from palynological and petrological investigations. Neues Jahrb. Geol. Paläontologie—Abh. 1996, 200, 201–220. [Google Scholar] [CrossRef]
- Okuda, M.; van Vugt, N.; Nakagawa, T.; Ikeya, M.; Hayashida, A.; Yasuda, Y.; Setoguchi, T. Palynological evidence for the astronomical origin of lignite–detritus sequence in the Middle Pleistocene Marathousa Member, Megalopolis, SW Greece. Earth Planet. Sci. Lett. 2002, 201, 143–157. [Google Scholar] [CrossRef]
- Karkanas, P.; Tourloukis, V.; Thompson, N.; Giusti, D.; Panagopoulou, E.; Harvati, K. Sedimentology and micromorphology of the Lower Palaeolithic lakeshore site Marathousa 1, Megalopolis basin, Greece. Quat. Int. 2018, 497, 123–136. [Google Scholar] [CrossRef]
- Jacobs, Z.; Li, B.; O’Gorman, K.; Tourloukis, V.; Panagopoulou, E.; Karkanas, P.; Harvati, K. Single-grain optical dating of sediment samples from Marathousa 1, southern Greece. In Human Evolution at the CROSSROADS: Research in Greece and Beyond; Harvati, K., Ioannidou, M., Eds.; Tübingen University Press: Tübingen, Germany, 2024. [Google Scholar]
- van Kolfschoten, T.; Konidaris, G.E.; Doukas, C.; Athanassiou, A.; Tourloukis, V.; Panagopoulou, E.; Karkanas, P.; Harvati, K. Voles (Rodentia, Mammalia) as a proxy to date the site Kyparissia 4 (Megalopolis Basin, Greece). In Human Evolution at the CROSSROADS: Research in Greece and Beyond; Harvati, K., Ioannidou, M., Eds.; Tübingen University Press: Tübingen, Germany, 2024. [Google Scholar]
- van der Meulen, A.J. Middle Pleistocene smaller mammals from the Monte Peglia (Orvieto, Italy) with special reference to the phylogeny of Microtus (Arvicolidae, Rodentia). Quaternaria 1973, 17, 1–144. [Google Scholar]
- Luzi, E.; López-García, J.M. Patterns of variation in Microtus arvalis and Microtus agrestis populations from Middle to Late Pleistocene in southwestern Europe. Hist. Biol. 2019, 31, 535–543. [Google Scholar] [CrossRef]
- Mazza, P. New evidence on the Pleistocene hippopotamuses of Western Europe. Geol. Romana 1995, 31, 61–241. [Google Scholar]
- Kahlke, R.D. Die Hippopotamus-Reste aus dem Unterpleistozän von Untermaßfeld. In Das Pleistozän von Untermaßfeld bei Meiningen (Thüringen), Teil 1; Kahlke, R.D., Ed.; Monographien des Römisch-Germanischen Zentralmuseums Mainz, Dr. Rudolf Habelt: Bonn, Germany, 1997; Volume 40, pp. 277–374. [Google Scholar]
- Galobart, À.; Ros, X.; Maroro, J.; Vila, B. Descripción del material de hipopótamo (Hippopotamus antiquus Desmarest, 1822) de los yacimientos del Pleistoceno inferior de Incarcal (Girona, NE de la Península Ibérica). Paleontol. Evol. 2003, 34, 153–173. [Google Scholar]
- Tsoukala, E.; Chatzopoulou, K. A new Early Pleistocene (latest Villafranchian) site with mammals in Kalamotó (Mygdonia Basin, Macedonia, Greece)—Preliminary report. Mitteilungen Komm. Quartärforschung Osterr. Akad. Wiss. 2005, 14, 213–233. [Google Scholar]
- Georgitsis, M.K.; Liakopoulou, D.Ε.; Theodorou, G.E.; Tsiolakis, E. Functional morphology of the hindlimb of fossilized pygmy hippopotamus from Ayia Napa (Cyprus). J. Morphol. 2022, 283, 1048–1079. [Google Scholar] [CrossRef]
- Faure, M. Les hippopotames quaternaires non-insulaires d’Europe occidentale. Nouv. Arch. Muséum D’histoire Nat. Lyon 1985, 23, 13–79. [Google Scholar] [CrossRef]
- Scott, K.M. Postcranial dimensions of ungulates as predictors of body mass. In Body Size in Mammalian Paleobiology. Estimation and Biological Implications; Damuth, J., MacFadden, B.J., Eds.; Cambridge University Press: Cambridge, UK, 1990; pp. 301–335. [Google Scholar]
- Palmqvist, P.; Rodríguez-Gómez, G.; Figueirido, B.; García-Aguilar, J.M.; Pérez-Claros, J.A. On the ecological scenario of the first hominin dispersal out of Africa. L’Anthropologie 2022, 126, 102998. [Google Scholar] [CrossRef]
- Konidaris, G.E.; Athanassiou, A.; Panagopoulou, E.; Karkanas, P.; Harvati, K. Fossil macaques (Cercopithecidae, Primates) from the Middle Pleistocene of the Megalopolis Basin (Greece) with description of a new specimen from Kyparissia 4. In Human Evolution at the CROSSROADS: Research in Greece and Beyond; Harvati, K., Ioannidou, M., Eds.; Tübingen University Press: Tübingen, Germany, 2024. [Google Scholar]
- Athanassiou, A.; Konidaris, G.E.; Tourloukis, V.; Thompson, N.; Giusti, D.; Panagopoulou, E.; Karkanas, P.; Harvati, K. The Middle Pleistocene large mammal fauna from Kyparissia (Peloponnese, S. Greece): New collected material. In Human Evolution at the CROSSROADS: Research in Greece and Beyond; Harvati, K., Ioannidou, M., Eds.; Tübingen University Press: Tübingen, Germany, 2024. [Google Scholar]
- Mazza, P.P.A.; Bertini, A. Were Pleistocene hippopotamuses exposed to climate-driven body size changes? Boreas 2013, 42, 194–209. [Google Scholar] [CrossRef]
- Sala, B. Bison schoetensacki Freud. from Isernia la Pineta (early Mid-Pleistocene-Italy) and revision of the European species of bison. Palaeontogr. Ital. 1986, 74, 113–170. [Google Scholar]
- Konidaris, G.E.; Athanassiou, A.; Tourloukis, V.; Thompson, N.; Giusti, D.; Panagopoulou, E.; Harvati, K. The skeleton of a straight-tusked elephant (Palaeoloxodon antiquus) and other large mammals from the Middle Pleistocene butchering locality Marathousa 1 (Megalopolis Basin, Greece): Preliminary results. Quat. Int. 2018, 497, 65–84. [Google Scholar] [CrossRef]
- Stefaniak, K. Neogene and Quaternary Cervidae from Poland; Institute of Systematics and Evolution of Animals of the Polish Academy of Sciences: Kraków, Poland, 2015; p. 260. [Google Scholar]
- Cherin, M. New material of Lutra simplicidens (Carnivora, Mustelidae, Lutrinae), a key taxon for understanding the evolution of European otters. Riv. Ital. Paleontol. Stratigr. 2017, 123, 433–441. [Google Scholar]
- Löhnert, E.; Nowak, H. Die Braunkohlenlagerstätte von Khoremi im Becken von Megalopolis/Peloponnes. Geol. Jahrb. 1965, 82, 847–868. [Google Scholar]
- Panagopoulou, E.; Tourloukis, V.; Thompson, N.; Athanassiou, A.; Tsartsidou, G.; Konidaris, G.E.; Giusti, D.; Karkanas, P.; Harvati, K. Marathousa 1: A new Middle Pleistocene archaeological site from Greece. Antiquity 2015, 343, 1–8. Available online: https://antiquity.ac.uk/projgall/panagopoulou343 (accessed on 28 May 2024).
- Harvati, K.; Konidaris, G.; Tourloukis, V. Paleoanthropology at the Gates of Europe: Recent research in Greece in the frame of the PaGE ERC Starting Grant project. Quat. Int. 2018, 497, 1–3. [Google Scholar] [CrossRef]
- Blackwell, B.A.B.; Sakhrani, N.; Singh, I.K.; Gopalkrishna, K.K.; Tourloukis, V.; Panagopoulou, E.; Karkanas, P.; Blickstein, J.I.B.; Skinner, A.R.; Florentin, J.A.; et al. ESR dating ungulate teeth and molluscs from the Paleolithic site Marathousa 1, Megalopolis Basin, Greece. Quaternary 2018, 1, 22. [Google Scholar] [CrossRef]
- Doukas, C.; van Kolfschoten, T.; Papayianni, K.; Panagopoulou, E.; Harvati, K. The small mammal fauna from the palaeolithic site Marathousa 1 (Greece). Quat. Int. 2018, 497, 95–107. [Google Scholar] [CrossRef]
- Jacobs, Z.; Li, B.; Karkanas, P.; Tourloukis, V.; Thompson, N.; Panagopoulou, E.; Harvati, K. Optical dating of K-feldspar grains from Middle Pleistocene lacustrine sediment at Marathousa 1 (Greece). Quat. Int. 2018, 497, 170–177. [Google Scholar] [CrossRef]
- Bludau, I.J.E.; Papadopoulou, P.; Iliopoulos, G.; Weiss, M.; Schnabel, E.; Thompson, N.; Tourloukis, V.; Zachow, C.; Kyrikou, S.; Konidaris, G.E.; et al. Lake-level changes and their paleoclimatic implications at the MIS12 Lower Paleolithic (Middle Pleistocene) site Marathousa 1, Greece. Front. Earth Sci. 2021, 9, 668445. [Google Scholar] [CrossRef]
- Boni, G. Paleoenvironmental reconstruction based on mollusc macrofaunal analysis: A case study from the Middle Pleistocene of Megalopolis basin (Peloponnesus, Greece). Master’s Thesis, Aristotle University of Thessaloniki, Thessaloniki, Greece, 2022. [Google Scholar]
- Field, M.H.; Ntinou, M.; Tsartsidou, G.; van Berge Henegouwen, D.; Risberg, J.; Tourloukis, V.; Thompson, N.; Karkanas, P.; Panagopoulou, E.; Harvati, K. A palaeoenvironmental reconstruction (based on palaeobotanical data and diatoms) of the Middle Pleistocene elephant (Palaeoloxodon antiquus) butchery site at Marathousa, Megalopolis, Greece. Quat. Int. 2018, 497, 108–122. [Google Scholar] [CrossRef]
- Konidaris, G.E.; Athanassiou, A.; Panagopoulou, E.; Harvati, K. First record of Macaca (Cercopithecidae, Primates) in the Middle Pleistocene of Greece. J. Hum. Evol. 2022, 162, 103104. [Google Scholar] [CrossRef] [PubMed]
- Michailidis, D.; Konidaris, G.E.; Athanassiou, A.; Panagopoulou, E.; Harvati, K. The ornithological remains from Marathousa 1 (Middle Pleistocene; Megalopolis Basin, Greece). Quat. Int. 2018, 497, 85–94. [Google Scholar] [CrossRef]
- Tourloukis, V.; Thompson, N.; Panagopoulou, E.; Giusti, D.; Konidaris, G.E.; Karkanas, P.; Harvati, K. Lithic artifacts and bone tools from the Lower Palaeolithic site Marathousa 1, Megalopolis, Greece: Preliminary results. Quat. Int. 2018, 497, 47–64. [Google Scholar] [CrossRef]
- Vlachos, E.; Georgalis, G.L.; Konidaris, G.E.; Athanassiou, A.; Tourloukis, V.; Thompson, N.; Panagopoulou, E.; Harvati, K. Preliminary results on the reptiles from the Middle Pleistocene of Marathousa 1, Megalopolis Basin (Greece). In Human Evolution at the CROSSROADS: Research in Greece and Beyond; Harvati, K., Ioannidou, M., Eds.; Tübingen University Press: Tübingen, Germany, 2024. [Google Scholar]
- Guibert-Cardin, J.; Tourloukis, V.; Thompson, N.; Panagopoulou, E.; Harvati, K.; Nicoud, E.; Beyries, S. The function of small tools in Europe during the Middle Pleistocene: The case of Marathousa 1 (Megalopolis, Greece). J. Lithic Stud. 2022, 9, 1–29. [Google Scholar] [CrossRef]
- Laws, R.M. Dentition and ageing of the hippopotamus. East Afr. Wildl. J. 1968, 6, 19–52. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Hooijer, D.A. The fossil Hippopotamidae of Asia, with notes on the recent species. Zool. Verh. 1950, 8, 1–123. [Google Scholar]
- Alberdi, M.T.; Ruiz Bustos, A. Descripción y significado biostratigráfico y climático del Equus e Hippopotamus, en el yacimiento de Venta Micena (Granada). Estud. Geológicos 1985, 41, 251–261. [Google Scholar] [CrossRef]
- Kahlke, R.D. Schädelreste von Hippopotamus aus dem Unterpleistozän von Untermaßfeld. In Das Pleistozän von Untermaßfeld bei Meiningen (Thüringen), Teil 2; Kahlke, R.D., Ed.; Monographien des Römisch-Germanischen Zentralmuseums Mainz, Dr. Rudolf Habelt: Bonn, Germany, 2001; Volume 40, pp. 483–500. [Google Scholar]
- Fidalgo, D.; Rosas, A.; Madurell-Malapeira, J.; Pineda, A.; Huguet, R.; García-Tabernero, A.; Cáceres, I.; Ollé, A.; Vallverdú, J.; Saladie, P. A review on the Pleistocene occurrences and palaeobiology of Hippopotamus antiquus based on the record from the Barranc de la Boella Section (Francolí Basin, NE Iberia). Quat. Sci. Rev. 2023, 307, 108034. [Google Scholar] [CrossRef]
- Martínez-Navarro, B.; Palmqvist, P.; Madurell-Malapeira, J.; Ros-Montoya, S.; Espigares, M.P.; Torregrosa, V.; Pérez-Claros, J.A. La fauna de grandes mamíferos de Fuente Nueva-3 y Barranco León-5: Estado de la cuestión. In Ocupaciones Humanas en el Pleistoceno Inferior y Medio de la Cuenca de Guadix-Baza; Toro, I., Martínez-Navarro, B., Agustí, J., Eds.; Consejería de Cultura, Junta de Andalucía, Arqueología Monografías: Sevilla, Spain, 2010; pp. 197–236. [Google Scholar]
- Inuzuka, N.; Hasegawa, Y.; Nogariya, H.; Kamei, T. On the stylohyoid bone of Naumann’s elephant (Elephas naumanni Makiyama) from Lake Nojiri. Mem. Fac. Sci. Kyoto Univ. Ser. Geol. Mineral. 1975, 41, 49–65. [Google Scholar]
- Palombo, M.R. Intra-specific variation of stylohyoid bones in Palaeoloxodon: A case study of Neumark-Nord 1 (Geiseltal, Sachsen-Anhalt, Germany) straight-tusked elephants. Quat. Int. 2012, 276–277, 77–92. [Google Scholar] [CrossRef]
- Athanassiou, A. A skeleton of Mammuthus trogontherii (Proboscidea, Elephantidae) from NW Peloponnese, Greece. Quat. Int. 2012, 255, 9–28. [Google Scholar] [CrossRef]
- Shoshani, J.; Ferretti, M.P.; Lister, A.M.; Agenbroad, L.D.; Saegusa, H.; Mol, D.; Takahashi, K. Relationships within the Elephantinae using hyoid characters. Quat. Int. 2007, 169–170, 174–185. [Google Scholar] [CrossRef]
- Guenther, E.W. Die Biber (Trogontherium cuvieri Fisch. und Castor fiber L.) der altpleistozänen Fundstelle von Voigtstedt in Thüringen. Paläontologische Abh. A 1965, 2, 566–582. [Google Scholar]
- Fischer, K. Postkraniale Skelettreste von Bibern (Castor L., Trogontherium Fischer, Castoridae, Rodentia, Mamm.) aus dem Mittelpleistozän von Bilzingsleben. In Bilzingsleben IV. Homo erectus—Seine Kultur und Seine Umwelt; Fischer, K., Guenther, E.W., Heinrich, W.D., Mania, D., Musil, R., Nötzold, T., Eds.; Veröffentlichungen des Landesmuseums für Vorgeschichte in Halle: Berlin, Germany, 1991; pp. 63–70. [Google Scholar]
- Lister, A.M. The morphological distinction between bones and teeth of fallow deer (Dama dama) and red deer (Cervus elaphus). Int. J. Osteoarchaeol. 1996, 6, 119–143. [Google Scholar] [CrossRef]
- Pfeiffer, T. Die Stellung von Dama (Cervidae, Mammalia) im System plesiometacarpaler Hirsche des Pleistozäns. Cour. Forschungsinstitut Senckenberg 1999, 211, 1–218. [Google Scholar]
- Heintz, E. Les cervidés Villafranchiens de France et d’Espagne. Volume II: Figures et tableaux. Mem. Mus. Natl. D’histoire Nat. Série C 1970, 22, 1–206. [Google Scholar]
- Zimmerman, K. Zur Evolution der Molarenstruktur der Erdmaus, Microtus agrestis (L.). Zool. Jahrbücher Abt. Syst. Okol. Und Geogr. Tiere 1956, 84, 270–274. [Google Scholar]
- Markova, E.; Malygin, V.; Montuire, S.; Nadachowski, A.; Quéré, J.P.; Ochman, K. Dental variation in sibling species Microtus arvalis and M. rossiaemeridionalis (Arvicolinae, Rodentia): Between-species comparisons and geography of morphotype dental patterns. J. Mamm. Evol. 2010, 17, 121–139. [Google Scholar] [CrossRef]
- Heinrich, W.D. Zur Evolution und Biostratigraphie von Arvicola (Rodentia, Mammalia) im Pleistozän Europas. Z. Geol. Wiss. 1982, 10, 683–735. [Google Scholar]
- Mahmoudi, A.; Maul, L.C.; Khoshyar, M.; Darvish, J.; Aliabadian, M.; Kryštufek, B. Evolutionary history of water voles revisited: Confronting a new phylogenetic model from molecular data with the fossil record. Mammalia 2020, 84, 171–184. [Google Scholar] [CrossRef]
- Masini, F.; Maul, L.C.; Abbazzi, L.; Petruso, D.; Savorelli, A. Independent water vole (Mimomys savini, Arvicola: Rodentia, Mammalia) lineages in Italy and Central Europe. Foss. Impr. 2020, 76, 59–83. [Google Scholar] [CrossRef]
- Maul, L.C. Die Kleinsäugerfauna aus der Forschungsbohrung Ummendorf 1/2012. Mitteilungen Geol. Bergwesen Sachs. -Anhalt 2019, 20, 99–108. [Google Scholar]
- Peretto, C.; Arnaud, J.; Moggi-Cecchi, J.; Manzi, G.; Nomade, S.; Pereira, A.; Falguères, C.; Bahain, J.J.; Grimaud-Hervé, D.; Berto, C.; et al. A human deciduous tooth and new 40Ar/39Ar dating results from the Middle Pleistocene archaeological site of Isernia La Pineta, southern Italy. PLoS ONE 2015, 10, e0140091. [Google Scholar] [CrossRef]
- Wagner, G.A.; Maul, L.C.; Löscher, M.; Schreiber, H.D. Mauer—The type site of Homo heidelbergensis: Palaeoenvironment and age. Quat. Sci. Rev. 2011, 30, 1464–1473. [Google Scholar] [CrossRef]
- van Kolfschoten, T.; Turner, E. Early Middle Pleistocene mammalian faunas from Kärlich and Miesenheim I and their biostratigraphical implications. In Proceedings of the Early Middle Pleistocene in Europe, Proceedings of the SEQS Cromer Symposium, Norwich, UK, 3–7 September 1996; pp. 227–253. [Google Scholar]
- Parfitt, S.A.; Barendregt, R.W.; Breda, M.; Candy, I.; Collins, M.J.; Coope, G.R.; Durbidge, P.; Field, M.H.; Lee, J.R.; Lister, A.M.; et al. The earliest record of human activity in northern Europe. Nature 2005, 438, 1008. [Google Scholar] [CrossRef]
- Danukalova, G.; Yakovlev, A.; Osipova, E.; Kurmanov, R.; van Kolfschoten, T. Biostratigraphy of the early Middle Pleistocene of the Southern Fore-Urals. Quat. Int. 2016, 420, 115–135. [Google Scholar] [CrossRef]
- Goren-Inbar, N.; Lister, A.; Werker, E.; Chech, M. A butchered elephant skull and associated artifacts from the Acheulian site of Gesher Benot Ya’aqov, Israel. Paléorient 1994, 20, 99–112. [Google Scholar] [CrossRef]
- Goren-Inbar, N.; Feibel, C.S.; Verosub, K.L.; Melamed, Y.; Kislev, M.E.; Tchernov, E.; Saragusti, I. Pleistocene milestones on the out-of-Africa corridor at Gesher Benot Ya’aqov, Israel. Science 2000, 289, 944–947. [Google Scholar] [CrossRef] [PubMed]
- Palombo, M.R. Deconstructing mammal dispersals and faunal dynamics in SW Europe during the Quaternary. Quat. Sci. Rev. 2014, 96, 50–71. [Google Scholar] [CrossRef]
- Malez, M.; Lenardić-Fabić, J. New subspecies of the southern elephant (Mammuthus meridionalis adriacus n. ssp.) from the bottom of the Adriatic Sea (Croatia, Yugoslavia). Palaeontol. Jugosl. 1988, 37, 1–36. [Google Scholar]
- Cherin, M.; Alba, D.M.; Crotti, M.; Menconero, S.; Moullé, P.É.; Sorbelli, L.; Madurell-Malapeira, J. The post-Jaramillo persistence of Sus strozzii (Suidae, Mammalia) in Europe: New evidence from the Vallparadís Section (NE Iberian Peninsula) and other coeval sites. Quat. Sci. Rev. 2020, 233, 106234. [Google Scholar] [CrossRef]
- Tsoukala, E.; Guérin, C. The Rhinocerotidae and Suidae of the Middle Pleistocene from Petralona Cave (Macedonia, Greece). Acta Zool. Bulg. 2016, 68, 243–264. [Google Scholar]
- Athanassiou, A. The fossil record of continental fossil deer (Mammalia: Artiodactyla: Cervidae) in Greece. In Fossil Vertebrates of Greece Vol. 2: Laurasiatherians, Artiodactyles, Perissodactyles, Carnivorans, and Island Endemics; Vlachos, E., Ed.; Springer: Cham, Switzerland, 2022; pp. 205–247. [Google Scholar]
- Pfeiffer, T. The first complete skeleton of Megaloceros verticornis (Dawkins, 1868) Cervidae, Mammalia, from Bilshausen (Lower Saxony, Germany): Description and phylogenetic implications Mitteilungen Mus. Naturkunde Berl. Geowiss. Reihe 2002, 5, 289–308. [Google Scholar] [CrossRef]
- Tsoukala, E. Contribution to the Study of the Pleistocene Fauna of Large Mammals (Carnivora, Perissodactyla, Artiodactyla) from Petralona Cave, Chalkidiki (N. Greece). Ph.D. Thesis, Aristotle University of Thessaloniki, Thessaloniki, Greece, 1989. [Google Scholar]
- Martino, R.; Marra, F.; Ríos, M.; Pandofli, L. The Middle Pleistocene hippopotamus from Malagrotta (Latium, Italy): New data and future perspectives. Quaternary 2024, 7, 13. [Google Scholar] [CrossRef]
- Stuart, A.J.; Wolff, R.G.; Lister, A.M.; Singer, R.; Egginton, J.M. Fossil vertebrates. In The Lower Paleolithic Site at Hoxne, England; Singer, R., Gladfelter, B.G., Wymer, J.J., Eds.; University of Chicago Press: Chicago, IL, USA, 1993; pp. 163–206. [Google Scholar]
- Kahlke, R.D.; García, N.; Kostopoulos, D.S.; Lacombat, F.; Lister, A.M.; Mazza, P.P.A.; Spassov, N.; Titov, V.V. Western Palaearctic palaeoenvironmental conditions during the Early and early Middle Pleistocene inferred from large mammal communities, and implications for hominin dispersal in Europe. Quat. Sci. Rev. 2011, 30, 1368–1395. [Google Scholar] [CrossRef]
- Willemsen, G.F. A revision of the Pliocene and Quaternary Lutrinae from Europe. Scr. Geol. 1992, 101, 1–115. [Google Scholar]
- van Kolfschoten, T. The evolution of the mammal fauna in the Netherlands and the middle Rhine Area (Western Germany) during the late Middle Pleistocene. Meded. Rijks Geol. Dienst 1990, 43, 1–69. [Google Scholar]
- van Kolfschoten, T. The mammal fauna from the interglacial deposits at Maastricht-Belvédère. Meded. Rijks Geol. Dienst 1993, 47, 51–60. [Google Scholar]
- Maul, L.C.; Markova, A.K. Similarity and regional differences in Quaternary arvicolid evolution in Central and Eastern Europe. Quat. Int. 2007, 160, 81–99. [Google Scholar] [CrossRef]
- Konidaris, G.E.; Kostopoulos, D.S. The Late Pliocene–Middle Pleistocene Large Mammal Faunal Units of Greece. Quaternary 2024, 7, 27. [Google Scholar] [CrossRef]
- Harvati, K.; Röding, C.; Bosman, A.M.; Karakostis, F.A.; Grün, R.; Stringer, C.; Karkanas, P.; Thompson, N.C.; Koutoulidis, V.; Moulopoulos, L.A.; et al. Apidima Cave fossils provide earliest evidence of Homo sapiens in Eurasia. Nature 2019, 571, 500–504. [Google Scholar] [CrossRef]
- Martino, R.; Pandolfi, L. The Quaternary Hippopotamus records from Italy. Hist. Biol. 2022, 34, 1146–1156. [Google Scholar] [CrossRef]
- Martino, R.; Ríos, M.I.; Mateus, O.; Pandolfi, L. Taxonomy, chronology, and dispersal patterns of Western European Quaternary hippopotamuses: New insight from Portuguese fossil material. Quat. Int. 2023, 674–675, 121–137. [Google Scholar] [CrossRef]
- Fidalgo, D.; Madurell-Malapeira, J.; Martino, R.; Pandolfi, L.; Rosas, A. An updated review of the Quaternary hippopotamus fossil records from the Iberian Peninsula. Quaternary 2024, 7, 4. [Google Scholar] [CrossRef]
- Mecozzi, B.; Iannucci, A.; Mancini, M.; Tentori, D.; Cavasinni, C.; Conti, J.; Messina, M.Y.; Sarra, A.; Sardella, R. Reinforcing the idea of an early dispersal of Hippopotamus amphibius in Europe: Restoration and multidisciplinary study of the skull from the Middle Pleistocene of Cava Montanari (Rome, central Italy). PLoS ONE 2023, 18, e0293405. [Google Scholar] [CrossRef]
- Pandolfi, L.; Petronio, C. A brief review of the occurrences of Pleistocene Hippopotamus (Mammalia, Hippopotamidae) in Italy. Geol. Croat. 2015, 68, 313–319. [Google Scholar] [CrossRef]
- Made, J.v.d.; Sahnouni, M.; Boufaghraief, K. Hippopotamus gorgops from El Kherba (Aigeria) and the context of its biogeography. In Proceedings of the II Meeting of African Prehistory, Burgos, Spain, 15–16 April 2017; pp. 135–169. [Google Scholar]
- Petronio, C. Note on the taxonomy of Pleistocene hippopotamuses. Ibex 1995, 3, 53–55. [Google Scholar]
- Fidalgo, D.; Rosas, A.; Bartolini-Lucenti, S.; Boisserie, J.R.; Pandolfi, L.; Martínez-Navarro, B.; Palmqvist, P.; Rook, L.; Madurell-Malapeira, J. Increase on environmental seasonality through the European Early Pleistocene inferred from dental enamel hypoplasia. Sci. Rep. 2023, 13, 16941. [Google Scholar] [CrossRef] [PubMed]
- Palombo, M.R.; Albayrak, E.; Marano, F. The straight-tusked elephants from Neumark-Nord. A glance into a lost world. In Elefantenreich. Eine Fossilwelt in Europa; Meller, H., Ed.; Landesamt für Denkmalpflege und Archäologie Sachsen-Anhalt (Halle): Halle, Germany, 2010; pp. 219–247. [Google Scholar]
- Saarinen, J.; Lister, A.M. Dental mesowear reflects local vegetation and niche separation in Pleistocene proboscideans from Britain. J. Quat. Sci. 2016, 31, 799–808. [Google Scholar] [CrossRef]
- Rivals, F.; Semprebon, G.M.; Lister, A.M. Feeding traits and dietary variation in Pleistocene proboscideans: A tooth microwear review. Quat. Sci. Rev. 2019, 219, 145–153. [Google Scholar] [CrossRef]
- Tsoukala, E.; Lister, A. Remains of straight-tusked elephant, Elephas (Palaeoloxodon) antiquus Falc. & Caut. (1847) ESR-dated to oxygen isotope Stage 6 from Grevena (W. Macedonia, Greece). Boll. Soc. Paleontol. Ital. 1998, 37, 117–139. [Google Scholar]
- Lister, A.M. Ecological interactions of elephantids in Pleistocene Eurasia: Palaeoloxodon and Mammuthus. In Human paleoecology in the Levantine Corridor; Goren-Inbar, N., Speth, J.D., Eds.; Oxbow Books: Oxford, UK, 2004; pp. 53–60. [Google Scholar]
- Roditi, E.; Bocherens, H.; Konidaris, G.E.; Athanassiou, A.; Tourloukis, V.; Karkanas, P.; Panagopoulou, E.; Harvati, K. Life-history of Palaeoloxodon antiquus reveals Middle Pleistocene glacial refugium in the Megalopolis basin, Greece. Sci. Rep. 2024, 14, 1390. [Google Scholar] [CrossRef]
- Lister, A.M. Evolutionary and ecological origins of British deer. Proc. R. Soc. Edinb. 1984, 82B, 205–229. [Google Scholar] [CrossRef]
- Elton, S.; O’Regan, H.J. Macaques at the margins: The biogeography and extinction of Macaca sylvanus in Europe. Quat. Sci. Rev. 2014, 96, 117–130. [Google Scholar] [CrossRef]
- Vlachos, E.; Delfino, M. Food for thought: Sub-fossil and fossil chelonian remains from Franchthi Cave and Megalopolis confirm a glacial refuge for Emys orbicularis in Peloponnesus (S. Greece). Quat. Sci. Rev. 2016, 150, 158–171. [Google Scholar] [CrossRef]
- Boni, G.; Syrides, G.; Konidaris, G.E.; Athanassiou, A.; Tourloukis, V.; Koukousioura, O.; Panagopoulou, E.; Karkanas, P.; Harvati, K. Preliminary results on the taxonomy and palaeoenvironmental analysis of the mollusc fauna from Marathousa 1, Marathousa 2 and Kyparissia 4 (Middle Pleistocene, Megalopolis Basin, Greece). In Human Evolution at the CROSSROADS: Research in Greece and Beyond; Harvati, K., Ioannidou, M., Eds.; Tübingen University Press: Tübingen, Germany, 2024. [Google Scholar]
- Papadopoulou, P.; Tsoni, M.; Konidaris, G.E.; Tourloukis, V.; Panagopoulou, E.; Karkanas, P.; Harvati, K.; Iliopoulos, G. Ostracod contribution to the palaeoenvironmental study of Kyparissia 4 (Megalopolis Basin, Greece). In Human Evolution at the CROSSROADS: Research in Greece and Beyond; Harvati, K., Ioannidou, M., Eds.; Tübingen University Press: Tübingen, Germany, 2024. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).