
Analysis of Molecular Structure Changes in Humic Acids from Manure-Amended Soils over 17 Years Using Elemental Analysis and Solid-State 13C Nuclear Magnetic Resonance Spectroscopy

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Treatments
2.2. Soil Sampling and Analysis
2.3. Extraction and Purification of Humic Acid
2.4. Elemental Analysis
2.5. NMR Experiments
2.6. Data Processing and Analysis
3. Results
3.1. Elemental Composition of HA
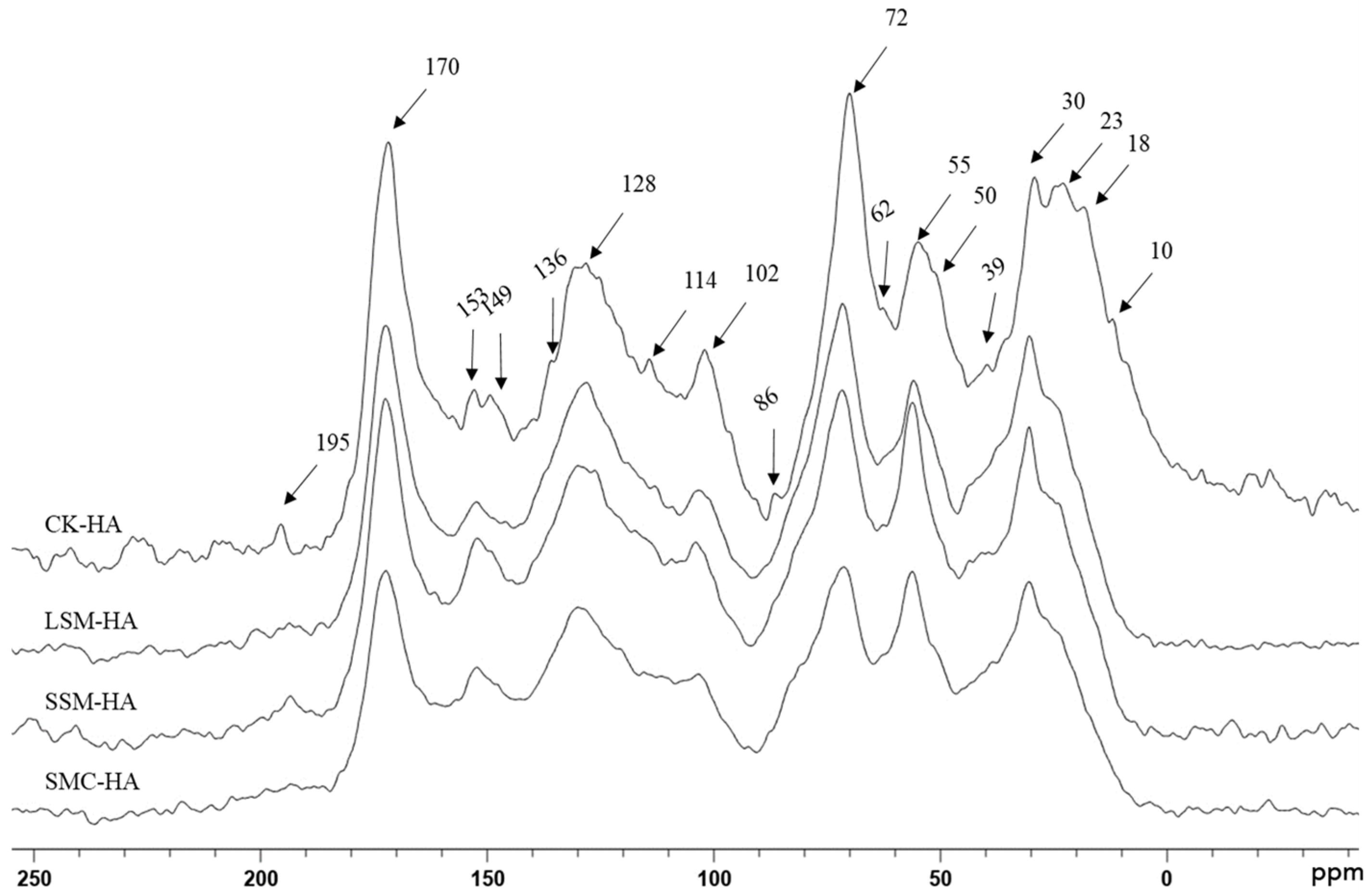
3.2. Solid-State 13C NMR of HA
4. Discussion
4.1. Formation and Composition of HA
4.2. Solid-State 13C NMR of HA
4.3. Concentration of Carboxylic Functional Group in hAs
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Soman, C.; Li, D.; Wander, M.M.; Kent, A.D. Long-term fertilizer and crop-rotation treatments differentially affect soil bacterial community structure. Plant Soil 2017, 413, 145–159. [Google Scholar] [CrossRef]
- Audette, Y.; Congreves, K.A.; Schneider, K.; Zaro, G.C.; Nunes, A.L.P.; Zhang, H.; Voroney, R.P. The effect of agroecosystem management on the distribution of C functional groups in soil organic matter: A review. Biol. Fertil. Soils 2022, 57, 881–894. [Google Scholar] [CrossRef] [PubMed]
- Gerke, J. Carbon Accumulation in Arable Soils: Mechanisms and the Effect of Cultivation Practices and Organic Fertilizers. Agronomy 2021, 11, 1079. [Google Scholar] [CrossRef]
- Essington, M.E. Soil and Water Chemistry: An Integrative Approach, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Scotti, R.; Bonanomi, G.; Scelza, R.; Zoina, A.; Rao, M. Organic amendments as sustainable tool to recovery fertility in intensive agricultural systems. J. Soil Sci. Plant Nutr. 2015, 15, 333–352. [Google Scholar] [CrossRef]
- Mitchell, P.J.; Simpson, A.J.; Soong, R.; Schurman, J.S.; Thomas, S.C.; Simpson, M.J. Biochar amendment and phosphorus fertilization altered forest soil microbial community and native soil organic matter molecular composition. Biogeochemistry 2016, 130, 227–245. [Google Scholar] [CrossRef]
- DiDonato, N.; Chen, H.; Waggoner, D.; Hatcher, P.G. Potential origin and formation for molecular components of humic acids in soils. Geochim. Cosmochim. Acta 2016, 178, 210–222. [Google Scholar] [CrossRef]
- Guppy, C.N.; Menzies, N.W.; Moody, P.W.; Blamey, F.P.C. Competitive sorption reactions between phosphorus and organic matter in soil: A review. Soil Res. 2004, 43, 189–202. [Google Scholar] [CrossRef]
- Kalbitz, K.; Schmerwitz, J.; Schwesig, D.; Matzner, E. Biodegradation of soil-derived dissolved organic matter as related to its properties. Geoderma 2003, 113, 273–291. [Google Scholar] [CrossRef]
- Swift, R.S. Macromolecular properties of soil humic substances: Fact, fiction, and opinion. Soil Sci. 1999, 164, 790–802. [Google Scholar] [CrossRef]
- Piccolo, A. The Supramolecular Structure of Humic Substances. Soil Sci. 2001, 166, 810–832. [Google Scholar] [CrossRef]
- Saurette, D.; Warren, J.; Heck, R. Soils of Ontario. In Digging into Canadian Soils: An Introduction to Soil Science; Krzic, M., Walley, F.L., Diochon, A., Paré, M.C., Farrell, R.E., Eds.; CSSS: Pinawa, MB, Canada, 2021. [Google Scholar]
- Leytem, A.B.; Mikkelsen, R.L. The nature of phosphorus in calcareous soils. Better Crop. 2005, 89, 11–13. [Google Scholar]
- von Wandruszka, R. Phosphorus retention in calcareous soils and the effect of organic matter on its mobility. Geochem. Trans. 2006, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- Zanin, L.; Tomasi, N.; Cesco, S.; Varanini, Z.; Pinton, R. Humic Substances Contribute to Plant Iron Nutrition Acting as Chelators and Biostimulants. Front. Plant Sci. 2019, 10, 675. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.J.; Zhang, T.Q.; Tan, C.S.; Welacky, T.; Wang, Y.T.; Lawrence, D.; Hong, J.P. Crop yield and phosphorus uptake as affected by phosphorus-based swine manure application under long-term corn-soybean rotation. Nutr. Cycl. Agroecosystems 2015, 103, 217–228. [Google Scholar] [CrossRef]
- Lavallee, J.M.; Soong, J.L.; Cotrufo, M.F. Conceptualizing soil organic matter into particulate and mineral-associated forms to address global change in the 21st century. Glob. Chang. Biol. 2020, 26, 261–273. [Google Scholar] [CrossRef]
- Rodrigues, L.A.T.; Dieckow, J.; Giacomini, S.; Ottonelli, A.S.; Zorzo, G.P.P.; Bayer, C. Carbon sequestration capacity in no-till soil decreases in the long-term due to saturation of fine silt plus clay-size fraction. Geoderma 2022, 412, 115711. [Google Scholar] [CrossRef]
- Maillard, É.; Angers, D.A.; Chantigny, M.; Bittman, S.; Rochette, P.; Lévesque, G.; Hunt, D.; Parent, L.-É. Carbon accumulates in organo-mineral complexes after long-term liquid dairy manure application. Agric. Ecosyst. Environ. 2015, 202, 108–119. [Google Scholar] [CrossRef]
- Olk, D.C.; Bloom, P.R.; De Nobili, M.; Chen, Y.; McKnight, D.M.; Wells, M.J.M.; Weber, J. Using Humic Fractions to Understand Natural Organic Matter Processes in Soil and Water: Selected Studies and Applications. J. Environ. Qual. 2019, 48, 1633–1643. [Google Scholar] [CrossRef]
- Kleber, M.; Lehmann, J. Humic Substances Extracted by Alkali Are Invalid Proxies for the Dynamics and Functions of Organic Matter in Terrestrial and Aquatic Ecosystems. J. Environ. Qual. 2019, 48, 207–216. [Google Scholar] [CrossRef]
- Savarese, C.; Drosos, M.; Spaccini, R.; Cozzolino, V.; Piccolo, A. Molecular characterization of soil organic matter and its extractable humic fraction from long-term field experiments under different cropping systems. Geoderma 2021, 383, 114700. [Google Scholar] [CrossRef]
- Zou, J.; Zhang, H.; Yue, D.; Huang, J. Is the traditional alkali extraction method valid in isolating chemically distinct humic acid? Chem. Eng. J. Adv. 2021, 6, 100077. [Google Scholar] [CrossRef]
- Jiménez-González, M.A.; Almendros, G.; Waggoner, D.C.; Álvarez, A.M.; Hatcher, P.G. Assessment of the molecular composition of humic acid as an indicator of soil carbon levels by ultra-high-resolution mass spectrometric analysis. Org. Geochem. 2020, 143, 104012. [Google Scholar] [CrossRef]
- Jakab, G.; Vancsik, A.; Filep, T.; Madarász, B.; Zacháry, D.; Ringer, M.; Ujházy, N.; Szalai, Z. Soil organic matter characterisation using alkali and water extraction, and its relation to soil properties. Geoderma Reg. 2022, 28, e00469. [Google Scholar] [CrossRef]
- Audette, Y.; Longstaffe, J.G.; Gillespie, A.W.; Smith, D.S.; Voroney, R.P. Validation and comparisons of NaOH and Na 4 P 2 O 7 extraction methods for the characterization of organic amendments. Soil Sci. Soc. Am. J. 2021, 85, 273–285. [Google Scholar] [CrossRef]
- Kim, S.; Kramer, R.W.; Hatcher, P.G. Graphical Method for Analysis of Ultrahigh-Resolution Broadband Mass Spectra of Natural Organic Matter, the Van Krevelen Diagram. Anal. Chem. 2003, 75, 5336–5344. [Google Scholar] [CrossRef]
- Ussiri, D.A.; Johnson, C.E. Characterization of organic matter in a northern hardwood forest soil by 13C NMR spectroscopy and chemical methods. Geoderma 2003, 111, 123–149. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS/STAT® User’s Guide; SAS Institute Inc.: Cary, NC, USA, 2021. [Google Scholar]
- Sposito, G. The Chemistry of Soils, 2nd ed.; Oxford University Press: New York, NY, USA, 2016. [Google Scholar]
- Dodla, S.K.; Wang, J.J.; Cook, R.L. Molecular Composition of Humic Acids from Coastal Wetland Soils along a Salinity Gradient. Soil Sci. Soc. Am. J. 2012, 76, 1592–1605. [Google Scholar] [CrossRef]
- Wong, S.; Hanna, J.V.; King, S.; Carroll, T.J.; Eldridge, R.J.; Dixon, D.R.; Bolto, B.A.; Hesse, S.; Abbt-Braun, G.; Frimmel, F.H. Fractionation of Natural Organic Matter in Drinking Water and Characterization by 13C Cross-Polarization Magic-Angle Spinning NMR Spectroscopy and Size Exclusion Chromatography. Environ. Sci. Technol. 2002, 36, 3497–3503. [Google Scholar] [CrossRef]
- Mao, J.-D.; Tremblay, L.; Gagné, J.-P.; Kohl, S.; Rice, J.; Schmidt-Rohr, K. Humic acids from particulate organic matter in the Saguenay Fjord and the St. Lawrence Estuary investigated by advanced solid-state NMR. Geochim. Cosmochim. Acta 2007, 71, 5483–5499. [Google Scholar] [CrossRef]
- SWIFT, R.S. Organic matter characterization. In Methods of Soil Analysis. Part 3. Chemical Methods; SSSA: Madison, WI, USA, 1996; Volume 5, pp. 1001–1069. [Google Scholar]
- Pu, Y.; Hallac, B.; Ragauskas, A.J. Plant Biomass Characterization: Application of Solution- and Solid-State NMR Spectroscopy. In Aqueous Pretreatment of Plant Biomass for Biological and Chemical Conversion to Fuels and Chemicals; John Wiley & Sons: Hoboken, NJ, USA, 2013; pp. 369–390. [Google Scholar] [CrossRef]
- Hatcher, P.G.; Rowan, R.; Mattingly, M.A. 1H and 13C NMR of marine humic acids. Org. Geochem. 1980, 2, 77–85. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Hattenschwiler, S.; Olander, L.; Allison, S. Nitrogen and nature. AMBIO A J. Hum. Environ. 2002, 31, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Bernal, M.; Sánchez-Monedero, M.; Paredes, C.; Roig, A. Carbon mineralization from organic wastes at different composting stages during their incubation with soil. Agric. Ecosyst. Environ. 1998, 69, 175–189. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Wallenstein, M.D.; Boot, C.M.; Denef, K.; Paul, E. The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant inputs form stable soil organic matter? Glob. Chang. Biol. 2013, 19, 988–995. [Google Scholar] [CrossRef] [PubMed]
- Kallenbach, C.M.; Frey, S.D.; Grandy, A.S. Direct evidence for microbial-derived soil organic matter formation and its ecophysiological controls. Nat. Commun. 2016, 7, 13630. [Google Scholar] [CrossRef]
- Dijkstra, P.; Thomas, S.C.; Heinrich, P.L.; Koch, G.W.; Schwartz, E.; Hungate, B.A. Effect of temperature on metabolic activity of intact microbial communities: Evidence for altered metabolic pathway activity but not for increased maintenance respiration and reduced carbon use efficiency. Soil Biol. Biochem. 2011, 43, 2023–2031. [Google Scholar] [CrossRef]
- Bahri, H.; Rasse, D.P.; Rumpel, C.; Dignac, M.-F.; Bardoux, G.; Mariotti, A. Lignin degradation during a laboratory incubation followed by 13C isotope analysis. Soil Biol. Biochem. 2008, 40, 1916–1922. [Google Scholar] [CrossRef]
- Lisý, A.; Ház, A.; Nadányi, R.; Jablonský, M.; Šurina, I. About Hydrophobicity of Lignin: A Review of Selected Chemical Methods for Lignin Valorisation in Biopolymer Production. Energies 2022, 15, 6213. [Google Scholar] [CrossRef]
- Xu, Z.; Peng, B.; Kitata, R.B.; Nicora, C.D.; Weitz, K.K.; Pu, Y.; Shi, T.; Cort, J.R.; Ragauskas, A.J.; Yang, B. Understanding of bacterial lignin extracellular degradation mechanisms by Pseudomonas putida KT2440 via secretomic analysis. Biotechnol. Biofuels Bioprod. 2022, 15, 117. [Google Scholar] [CrossRef]
- Zhao, C.; Xie, S.; Pu, Y.; Zhang, R.; Huang, F.; Ragauskas, A.J.; Yuan, J.S. Synergistic enzymatic and microbial lignin conversion. Green Chem. 2016, 18, 1306–1312. [Google Scholar] [CrossRef]
- Lam, B.; Baer, A.; Alaee, M.; Lefebvre, B.; Moser, A.; Williams, A.; Simpson, A.J. Major Structural Components in Freshwater Dissolved Organic Matter. Environ. Sci. Technol. 2007, 41, 8240–8247. [Google Scholar] [CrossRef]
- Woods, G.C.; Simpson, M.J.; Simpson, A.J. Oxidized sterols as a significant component of dissolved organic matter: Evidence from 2D HPLC in combination with 2D and 3D NMR spectroscopy. Water Res. 2012, 46, 3398–3408. [Google Scholar] [CrossRef] [PubMed]
- Christiernin, M. Lignin composition in cambial tissues of poplar. Plant Physiol. Biochem. 2006, 44, 700–706. [Google Scholar] [CrossRef] [PubMed]
- Wikberg, H.; Liisamaunu, S. Characterisation of thermally modified hard- and softwoods by 13C CPMAS NMR. Carbohydr. Polym. 2004, 58, 461–466. [Google Scholar] [CrossRef]
- Wang, Y.; Griffin, P.; Jin, K.; Fogel, M.L.; Steele, A.; Cody, G.D. Tracing H isotope effects in the dynamic metabolic network using multi-nuclear (1H, 2H and 13C) solid state NMR and GC–MS. Org. Geochem. 2013, 57, 84–94. [Google Scholar] [CrossRef]
- Higuchi, T. Microbial degradation of lignin: Role of lignin peroxidase, manganese peroxidase, and laccase. Proc. Jpn. Acad. B Phys. Biol. Sci. 2004, 80, 204–214. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Treatment Plots | pH | P (mg kg−1) | K (mg kg−1) | Mg (mg kg−1) | Ca (mg kg−1) | Base Saturation (K%) | Base Saturation (Mg%) | Base Saturation (Ca%) | SOM (%) | Amendment |
|---|---|---|---|---|---|---|---|---|---|---|
| CK | 6.3 | 6.3 | 129.3 | 405.7 | 2646.7 | 1.9 | 18.6 | 72.9 | 2.6 | No Manure (control) |
| LSM | 6.5 | 40.0 | 236.3 | 309.7 | 2056.7 | 4.2 | 17.4 | 70.0 | 3.2 | Liquid Swine Manure |
| SSM | 7.3 | 65.3 | 306.0 | 389.7 | 2803.3 | 4.1 | 16.8 | 72.7 | 4.4 | Solid Swine Manure |
| SMC | 7.1 | 51.3 | 257.0 | 433.3 | 3226.7 | 3.1 | 16.7 | 74.7 | 4.4 | Swine Manure Compost |
| Sample | Carbon (%) | Hydrogen (%) | Nitrogen (%) | Sulfur (%) | Oxygen (%) | O/C | H/C | C/N | HA Extracted (% SOM) |
|---|---|---|---|---|---|---|---|---|---|
| CK-HA | 44.01 c | 3.82 d | 3.50 c | 0.52 c | 32.82 b | 0.56 | 1.04 | 14.70 | 1.10c |
| LSM-HA | 49.73 a | 4.65 a | 4.61 a | 0.73 b | 31.21 c | 0.47 | 1.12 | 12.61 | 32.02a |
| SSM-HA | 45.55 b | 4.49 b | 4.41 b | 0.81 a | 36.00 a | 0.59 | 1.18 | 12.06 | 0.91c |
| SMC-HA | 46.59 b | 4.34 c | 4.39 b | 0.80 a | 19.16 d | 0.31 | 1.12 | 12.39 | 10.04b |
| Integral Areas | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Sample | Aliphatic | Methoxy | O-Alkyl | Aromatic | Phenolic | Carboxyl | Ketonic | Aromaticity | AOR |
| 0–50 ppm | 50–60ppm | 60–108 ppm | 108–145 ppm | 145–162 ppm | 162–190 ppm | 190–200 ppm | |||
| CK-HA | 20.4 | 5.1 | 17.8 | 30.6 | 9.2 | 16.6 | 0.2 | 0.48 | 1.15 |
| LSM-HA | 21.4 | 5.2 | 17.8 | 29.7 | 8.3 | 17.5 | 0.1 | 0.46 | 1.20 |
| SSM-HA | 21.1 | 6.5 | 18.5 | 28.8 | 9.4 | 15.7 | <0.1 | 0.45 | 1.14 |
| SMC-HA | 20.6 | 5.8 | 17.0 | 29.1 | 9.9 | 17.2 | 0.4 | 0.47 | 1.21 |
| Integral Areas | |||||
|---|---|---|---|---|---|
| Sample | Aliphatic (25–50 ppm) | Aromatic (120–140 ppm) | Aliphatic Region Normalization Factor * | Normalized Area of CPMAS Aromatic Region | Increase in Area of Aromatic Region from CPMAS |
| CK-HA | 11.71 | 16.88 | 1.33 | 22.41 | 5.53 |
| LSM-HA | 12.83 | 16.97 | 1.41 | 23.95 | 6.98 |
| SSM-HA | 12.5 | 17.04 | 1.31 | 22.29 | 5.25 |
| SMC-HA | 12.33 | 16.77 | 1.4 | 23.44 | 6.67 |
| Treatments | Carboxylic Acidity (cmol/kg) |
|---|---|
| CK-HA | 608.7 c |
| LSM-HA | 724.8 a |
| SSM-HA | 609.7 c |
| SMC-HA | 653.6 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohammed, I.; Kodaolu, B.; Zhang, T.; Wang, Y.; Audette, Y.; Longstaffe, J. Analysis of Molecular Structure Changes in Humic Acids from Manure-Amended Soils over 17 Years Using Elemental Analysis and Solid-State 13C Nuclear Magnetic Resonance Spectroscopy. Soil Syst. 2023, 7, 76. https://doi.org/10.3390/soilsystems7030076
Mohammed I, Kodaolu B, Zhang T, Wang Y, Audette Y, Longstaffe J. Analysis of Molecular Structure Changes in Humic Acids from Manure-Amended Soils over 17 Years Using Elemental Analysis and Solid-State 13C Nuclear Magnetic Resonance Spectroscopy. Soil Systems. 2023; 7(3):76. https://doi.org/10.3390/soilsystems7030076
Chicago/Turabian StyleMohammed, Ibrahim, Busayo Kodaolu, Tiequan Zhang, Yutao Wang, Yuki Audette, and James Longstaffe. 2023. "Analysis of Molecular Structure Changes in Humic Acids from Manure-Amended Soils over 17 Years Using Elemental Analysis and Solid-State 13C Nuclear Magnetic Resonance Spectroscopy" Soil Systems 7, no. 3: 76. https://doi.org/10.3390/soilsystems7030076
APA StyleMohammed, I., Kodaolu, B., Zhang, T., Wang, Y., Audette, Y., & Longstaffe, J. (2023). Analysis of Molecular Structure Changes in Humic Acids from Manure-Amended Soils over 17 Years Using Elemental Analysis and Solid-State 13C Nuclear Magnetic Resonance Spectroscopy. Soil Systems, 7(3), 76. https://doi.org/10.3390/soilsystems7030076