
Soil Solution Properties of Tropical Soils and Brachiaria Growth as Affected by Humic Acid Concentration


 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Soils Characterization
2.2. Humic Acid Characterization
2.3. Experimental Design and Treatments
2.4. Soil Solution Sampling and Analysis
2.5. Plant Analysis
2.6. Statistical Analysis
3. Results
3.1. Soil Solution
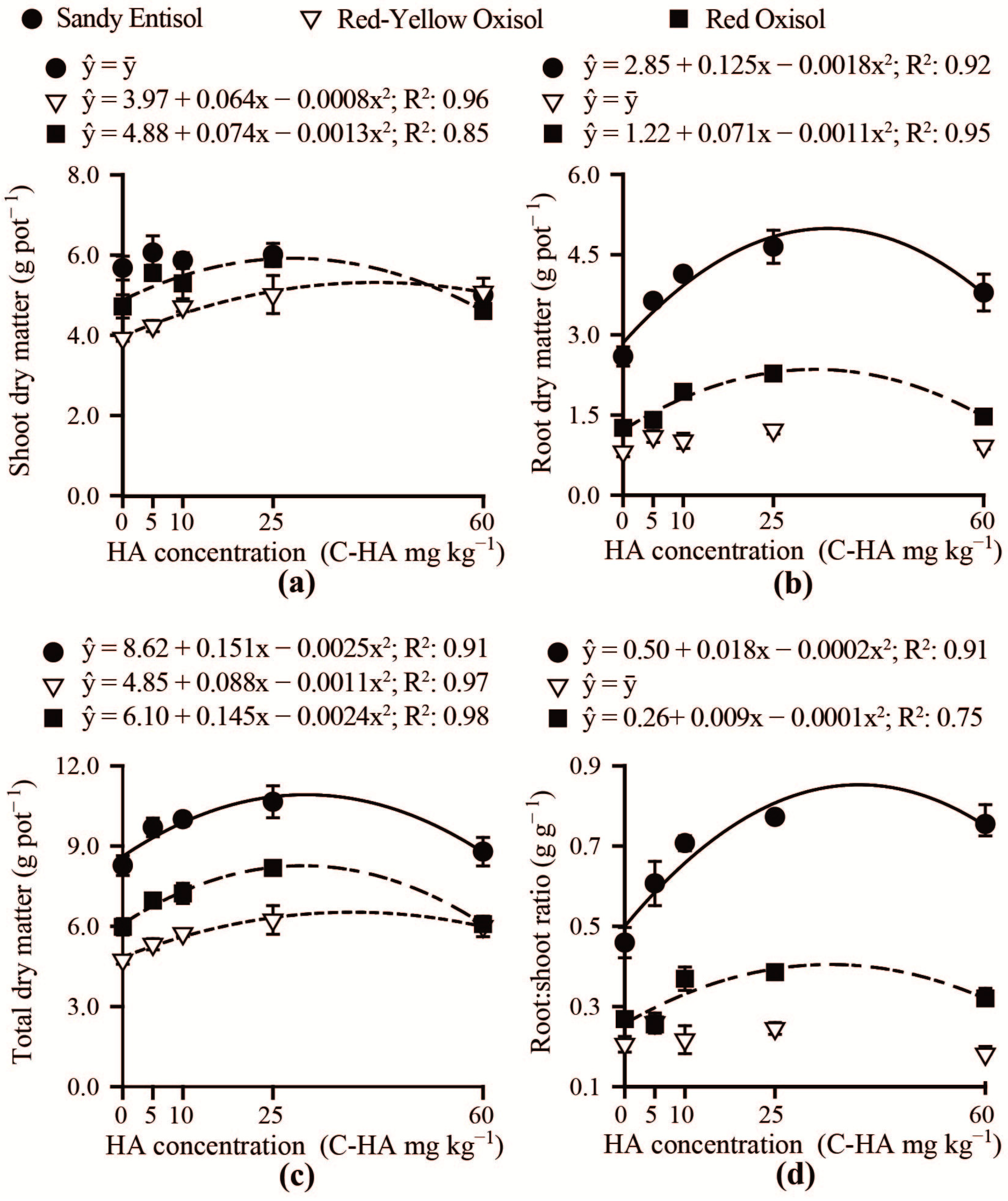
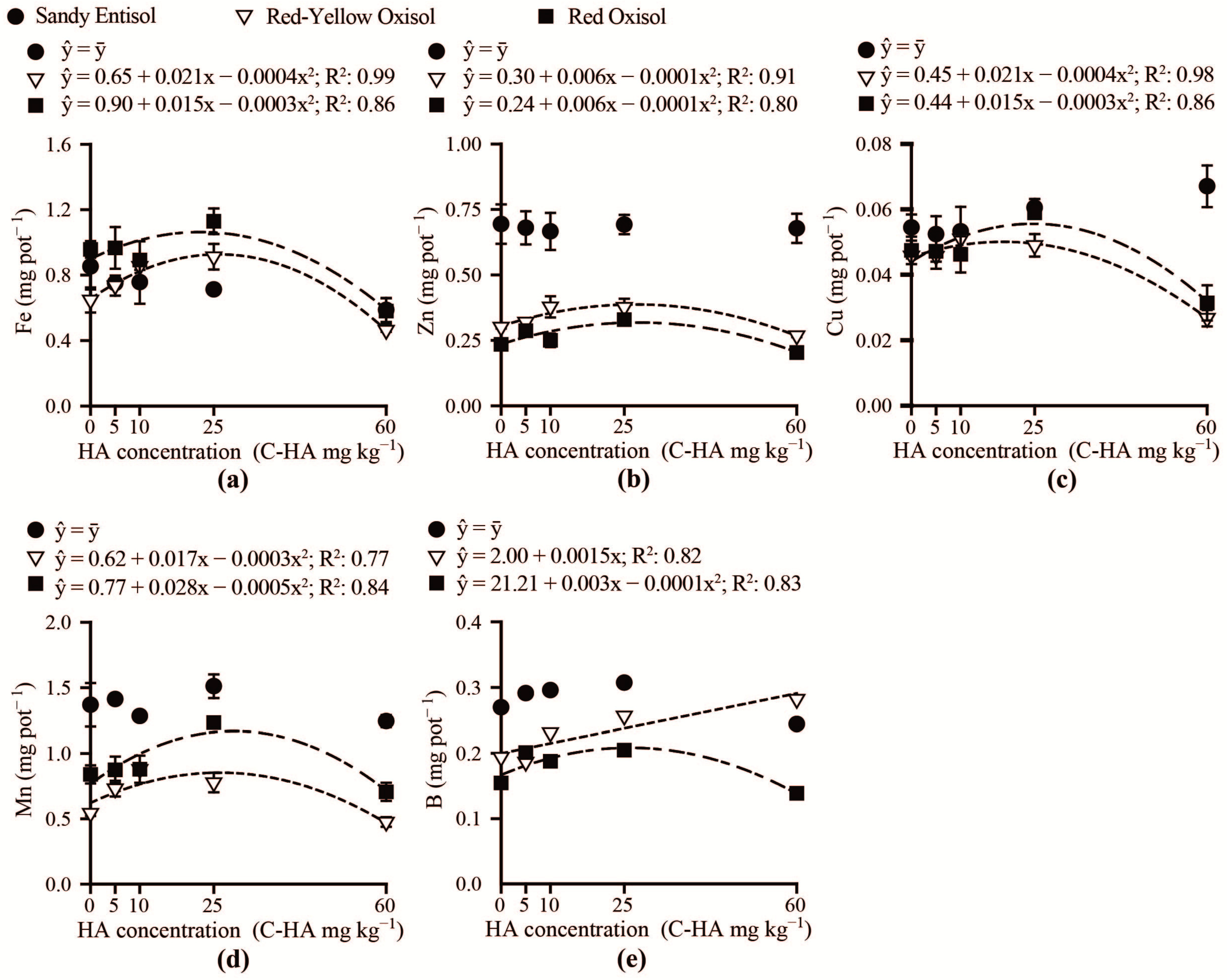
3.2. Brachiaria Nutrition and Growth
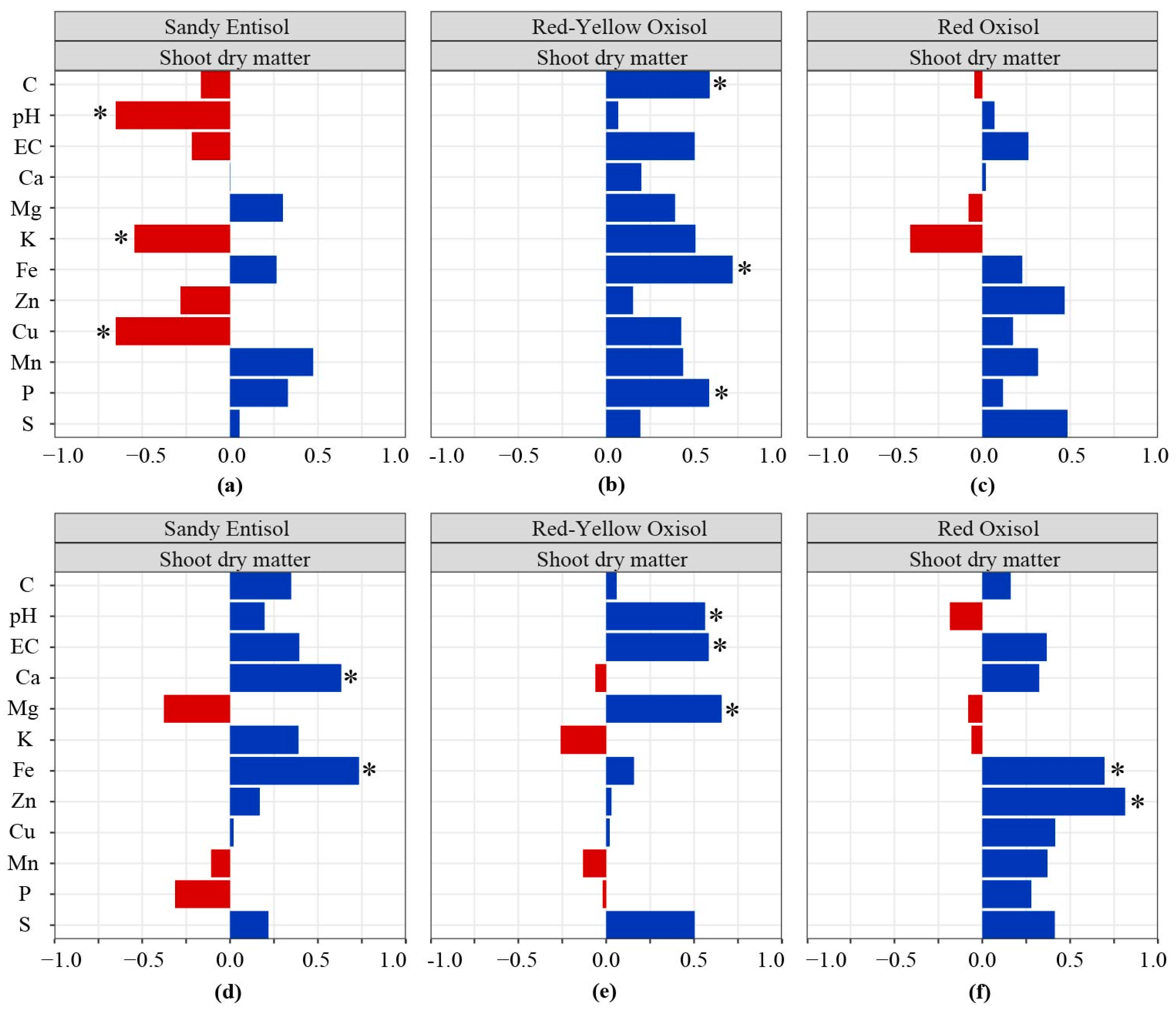
3.3. Pearson’s Correlation Matrix
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- ABIEC Beef Report: Perfil da Pecuária No Brasil. Available online: http://abiec.com.br/publicacoes/beef-report-2021/ (accessed on 10 April 2024).
- Jank, L.; Barrios, S.C.; do Valle, C.B.; Simeão, R.M.; Alves, G.F. The Value of Improved Pastures to Brazilian Beef Production. Crop Pasture Sci. 2014, 65, 1132. [Google Scholar] [CrossRef]
- Pereira, L.E.T.; Herling, V.R.; Tech, A.R.B. Current Scenario and Perspectives for Nitrogen Fertilization Strategies on Tropical Perennial Grass Pastures: A Review. Agronomy 2022, 12, 2079. [Google Scholar] [CrossRef]
- Pinheiro, P.L.; Passos, R.R.; Peçanha, A.L.; Canellas, L.P.; Olivares, F.L.; de Sá Mendonça, E. Promoting the Growth of Brachiaria Decumbens by Humic Acids (HAs). Aust. J. Crop Sci. 2018, 12, 1114–1121. [Google Scholar] [CrossRef]
- Verlinden, G.; Coussens, T.; De Vliegher, A.; Baert, G.; Haesaert, G. Effect of Humic Substances on Nutrient Uptake by Herbage and on Production and Nutritive Value of Herbage from Sown Grass Pastures. Grass Forage Sci. 2010, 65, 133–144. [Google Scholar] [CrossRef]
- Daur, I. Feed value of blue panic (Panicum antidotale Retz.) grass at different growth stages and under varying levels of humic acid in saline conditions. Turk. J. Field Crops 2016, 21, 210. [Google Scholar] [CrossRef]
- Neves, R.G.; Freitas, G.S.; Deminicis, B.B.; de Sá Mendonça, E.; Peçanha, A.L.; Dobbss, L.B.; Chambela Neto, A.; da Sá Deminicis, R.G. Dry Matter Yield, Growth Index, Chemical Composition and Digestibility of Marandu Grass under Nitrogen and Organic Fertilization. Semin. Ciências Agrárias 2019, 40, 1901. [Google Scholar] [CrossRef]
- Olaetxea, M.; De Hita, D.; Garcia, C.A.; Fuentes, M.; Baigorri, R.; Mora, V.; Garnica, M.; Urrutia, O.; Erro, J.; Zamarreño, A.M.; et al. Hypothetical Framework Integrating the Main Mechanisms Involved in the Promoting Action of Rhizospheric Humic Substances on Plant Root- and Shoot- Growth. Appl. Soil Ecol. 2018, 123, 521–537. [Google Scholar] [CrossRef]
- Shah, Z.H.; Rehman, H.M.; Akhtar, T.; Alsamadany, H.; Hamooh, B.T.; Mujtaba, T.; Daur, I.; Al Zahrani, Y.; Alzahrani, H.A.S.; Ali, S.; et al. Humic Substances: Determining Potential Molecular Regulatory Processes in Plants. Front. Plant Sci. 2018, 9, 263. [Google Scholar] [CrossRef] [PubMed]
- de Morais, E.G.; Silva, C.A.; Maluf, H.J.G.M. Soaking of Seedlings Roots in Humic Acid as an Effective Practice to Improve Eucalyptus Nutrition and Growth. Commun. Soil Sci. Plant Anal. 2021, 52, 1399–1415. [Google Scholar] [CrossRef]
- Nardi, S.; Schiavon, M.; Francioso, O. Chemical Structure and Biological Activity of Humic Substances Define Their Role as Plant Growth Promoters. Molecules 2021, 26, 2256. [Google Scholar] [CrossRef]
- Trevisan, S.; Francioso, O.; Quaggiotti, S.; Nardi, S. Humic Substances Biological Activity at the Plant-Soil Interface. Plant Signal. Behav. 2010, 5, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Nardi, S.; Pizzeghello, D.; Schiavon, M.; Ertani, A. Plant Biostimulants: Physiological Responses Induced by Protein Hydrolyzed-Based Products and Humic Substances in Plant Metabolism. Sci. Agric. 2016, 73, 18–23. [Google Scholar] [CrossRef]
- Tavares, O.C.H.; Santos, L.A.; Ferreira, L.M.; Sperandio, M.V.L.; da Rocha, J.G.; García, A.C.; Dobbss, L.B.; Berbara, R.L.L.; de Souza, S.R.; Fernandes, M.S. Humic Acid Differentially Improves Nitrate Kinetics under Low- and High-Affinity Systems and Alters the Expression of Plasma Membrane H+-ATPases and Nitrate Transporters in Rice. Ann. Appl. Biol. 2017, 170, 89–103. [Google Scholar] [CrossRef]
- Rose, M.T.; Patti, A.F.; Little, K.R.; Brown, A.L.; Jackson, W.R.; Cavagnaro, T.R. A Meta-Analysis and Review of Plant-Growth Response to Humic Substances. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 2014; pp. 37–89. [Google Scholar]
- Lyons, G.; Genc, Y. Commercial Humates in Agriculture: Real Substance or Smoke and Mirrors? Agronomy 2016, 6, 50. [Google Scholar] [CrossRef]
- Maluf, H.J.G.M.; Silva, C.A.; de Morais, E.G.; Paula, L.H.D. de Is Composting a Route to Solubilize Low-Grade Phosphate Rocks and Improve MAP-Based Composts? Rev. Bras. Ciência Solo 2018, 42, e0170079. [Google Scholar] [CrossRef]
- Rosa, S.D.; Silva, C.A.; Maluf, H.J.G.M. Phosphorus Availability and Soybean Growth in Contrasting Oxisols in Response to Humic Acid Concentrations Combined with Phosphate Sources. Arch. Agron. Soil Sci. 2020, 66, 220–235. [Google Scholar] [CrossRef]
- Tiwari, J.; Ramanathan, A.; Bauddh, K.; Korstad, J. Humic Substances: Structure, Function and Benefits for Agroecosystems—A Review. Pedosphere 2023, 33, 237–249. [Google Scholar] [CrossRef]
- Ampong, K.; Thilakaranthna, M.S.; Gorim, L.Y. Understanding the Role of Humic Acids on Crop Performance and Soil Health. Front. Agron. 2022, 4, 848621. [Google Scholar] [CrossRef]
- Rosolem, C.A.; Nascimento, C.A.C.; Bertolino, K.M.; Picoli, L.B. Humic Acid Enhances Phosphorus Transport in Soil and Uptake by Maize. J. Plant Nutr. Soil Sci. 2024, 187, 401–414. [Google Scholar] [CrossRef]
- Zavaschi, E.; de Abreu Faria, L.; Ferraz-Almeida, R.; do Nascimento, C.A.C.; Pavinato, P.S.; Otto, R.; Vitti, A.C.; Vitti, G.C. Dynamic of P Flux in Tropical Acid Soils Fertilized with Humic Acid–Complexed Phosphate. J. Soil Sci. Plant Nutr. 2020, 20, 1937–1948. [Google Scholar] [CrossRef]
- Gerke, J. Review Article: The Effect of Humic Substances on Phosphate and Iron Acquisition by Higher Plants: Qualitative and Quantitative Aspects. J. Plant Nutr. Soil Sci. 2021, 184, 329–338. [Google Scholar] [CrossRef]
- Macdonald, L.M.; Farrell, M.; Van Zwieten, L.; Krull, E.S. Plant Growth Responses to Biochar Addition: An Australian Soils Perspective. Biol. Fertil. Soils 2014, 50, 1035–1045. [Google Scholar] [CrossRef]
- Do Carmo, D.L.; Silva, C.A.; de Lima, J.M.; Pinheiro, G.L. Electrical Conductivity and Chemical Composition of Soil Solution: Comparison of Solution Samplers in Tropical Soils. Rev. Bras. Ciência Solo 2016, 40, e0140795. [Google Scholar] [CrossRef]
- De Morais, E.G.; Silva, C.A. Novel Slow-Release NPK Biochar-Based Fertilizers with Acidulated Apatite: Evaluation of the Fertilization Value in a Short-Term Experiment. J. Soil Sci. Plant Nutr. 2023, 23, 4937–4954. [Google Scholar] [CrossRef]
- Maluf, H.J.G.M.; Silva, C.A.; Curi, N.; Norton, L.D.; Rosa, S.D. Adsorption and Availability of Phosphorus in Response to Humic Acid Rates in Soils Limed with CaCO3 or MgCO3. Ciência Agrotecnologia 2018, 42, 7–20. [Google Scholar] [CrossRef]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural Uses of Plant Biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef]
- Halpern, M.; Bar-Tal, A.; Ofek, M.; Minz, D.; Muller, T.; Yermiyahu, U. The Use of Biostimulants for Enhancing Nutrient Uptake. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 2015; pp. 141–174. [Google Scholar]
- Chotzen, R.A.; Polubesova, T.; Chefetz, B.; Mishael, Y.G. Adsorption of Soil-Derived Humic Acid by Seven Clay Minerals: A Systematic Study. Clays Clay Miner. 2016, 64, 628–638. [Google Scholar] [CrossRef]
- De Morais, E.G.; Silva, C.A.; Rosa, S.D. Nutrient Acquisition and Eucalyptus Growth Affected by Humic Acid Sources and Concentrations. Semin. Ciências Agrárias 2018, 39, 1417. [Google Scholar] [CrossRef]
- Berbara, R.L.L.; García, A.C. Humic Substances and Plant Defense Metabolism. In Physiological Mechanisms and Adaptation Strategies in Plants under Changing Environment; Springer: New York, NY, USA, 2014; pp. 297–319. [Google Scholar]
- Canellas, L.P.; Olivares, F.L. Production of Border Cells and Colonization of Maize Root Tips by Herbaspirillum Seropedicae Are Modulated by Humic Acid. Plant Soil 2017, 417, 403–413. [Google Scholar] [CrossRef]
- Kjeldahl, J. Neue Methode Zur Bestimmung Des Stickstoffs in Organischen Körpern. Fresenius Z. Anal. Chem. 1883, 22, 366–382. [Google Scholar] [CrossRef]
- Silva, S.R.; de Barros, N.F.; de Novais, R.F.; Comerford, N.B. Eucalyptus Growth and Phosphorus Nutritional Efficiency as Affected by Soil Compaction and Phosphorus Fertilization. Commun. Soil Sci. Plant Anal. 2018, 49, 2700–2714. [Google Scholar] [CrossRef]
- Kalra, Y. Handbook of Reference Methods for Plant Analysis; Kalra, Y., Ed.; CRC Press: Boca Raton, FL, USA, 1997. [Google Scholar]
- Baligar, V.C.; Fageria, N.K.; He, Z.L. Nutrient Use Efficiency in Plants. Commun. Soil Sci. Plant Anal. 2001, 32, 921–950. [Google Scholar] [CrossRef]
- Ferreira, E.B.; Cavalcanti, P.P.; Nogueira, D.A. ExpDes.Pt: Pacote Experimental Designs (Portugues); R Core Team: Vienna, Austria, 2022. [Google Scholar]
- Ward, J.H. Hierarchical Grouping to Optimize an Objective Function. J. Am. Stat. Assoc. 1963, 58, 236–244. [Google Scholar] [CrossRef]
- Suzuki, R.; Terada, Y.; Shimodaira, H. Pvclust: Hierarchical Clustering with P-Values via Multiscale Bootstrap Resampling; R Core Team: Vienna, Austria, 2019. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2022. [Google Scholar]
- Bongiorno, G.; Bünemann, E.K.; Oguejiofor, C.U.; Meier, J.; Gort, G.; Comans, R.; Mäder, P.; Brussaard, L.; de Goede, R. Sensitivity of Labile Carbon Fractions to Tillage and Organic Matter Management and Their Potential as Comprehensive Soil Quality Indicators across Pedoclimatic Conditions in Europe. Ecol. Indic. 2019, 99, 38–50. [Google Scholar] [CrossRef]
- Gmach, M.R.; Cherubin, M.R.; Kaiser, K.; Cerri, C.E.P. Processes That Influence Dissolved Organic Matter in the Soil: A Review. Sci. Agric. 2020, 77, e20180164. [Google Scholar] [CrossRef]
- Al-Mallahi, J.; Tahhan, R.; Khresat, S. Quality of Fresh Plant Residue Affects Sequestration of Residue Derived Organic Material by Humic Acid. Eurasian J. Soil Sci. 2020, 9, 222–230. [Google Scholar] [CrossRef]
- Buragohain, P.; Sreedeep, S.; Lin, P.; Ni, J.; Garg, A. Influence of Soil Variability on Single and Competitive Interaction of Ammonium and Potassium: Experimental Study on Seven Different Soils. J. Soils Sediments 2019, 19, 186–197. [Google Scholar] [CrossRef]
- Basso, C.J.; Ceretta, C.A.; Durigon, R.; Poletto, N.; Girotto, E. Dejeto Líquido de Suínos: II—Perdas de Nitrogênio e Fósforo Por Percolação No Solo Sob Plantio Direto. Ciência Rural. 2005, 35, 1305–1312. [Google Scholar] [CrossRef]
- Urrutia, O.; Erro, J.; Guardado, I.; San Francisco, S.; Mandado, M.; Baigorri, R.; Claude Yvin, J.; Ma Garcia-Mina, J. Physico-chemical Characterization of Humic-metal-phosphate Complexes and Their Potential Application to the Manufacture of New Types of Phosphate-based Fertilizers. J. Plant Nutr. Soil Sci. 2014, 177, 128–136. [Google Scholar] [CrossRef]
- Jindo, K.; Soares, T.S.; Peres, L.E.P.; Azevedo, I.G.; Aguiar, N.O.; Mazzei, P.; Spaccini, R.; Piccolo, A.; Olivares, F.L.; Canellas, L.P. Phosphorus Speciation and High-affinity Transporters Are Influenced by Humic Substances. J. Plant Nutr. Soil Sci. 2016, 179, 206–214. [Google Scholar] [CrossRef]
- Yang, F.; Sui, L.; Tang, C.; Li, J.; Cheng, K.; Xue, Q. Sustainable Advances on Phosphorus Utilization in Soil via Addition of Biochar and Humic Substances. Sci. Total Environ. 2021, 768, 145106. [Google Scholar] [CrossRef] [PubMed]
- López-Velázquez, J.G.; López-López, M.E.; Rubio-Trías, A.; Ayón-Reyna, L.E.; Díaz-Corona, D.A.; Olivas Orozco, G.I.; Molina-Corral, J.; Vega-García, M.O. Cell Wall Stabilization and Calcium Absorption on Mango Fruit Treated with a Quarantine Hot Water Treatment Combined with Calcium Salts and Stored at Chilling Temperature. J. Food Biochem. 2022, 46, e14266. [Google Scholar] [CrossRef] [PubMed]
- Hawkesford, M.J.; Cakmak, I.; Coskun, D.; De Kok, L.J.; Lambers, H.; Schjoerring, J.K.; White, P.J. Functions of Macronutrients. In Marschner’s Mineral Nutrition of Plants; Elsevier: Amsterdam, The Netherlands, 2023; pp. 201–281. [Google Scholar]
- Rengel, Z.; Cakmak, I.; White, P.J. Marschner’s Mineral Nutrition of Plants; Academic Press: Cambridge, MA, USA, 2023. [Google Scholar]
- Soares Filho, C.V.; Cecato, U.; Ribeiro, O.L.; Roma, C.F.D.C.; Jobim, C.C.; Beloni, T.; Perri, S.H.V. Root System and Root and Stem Base Organic Reserves of Pasture Tanzania Grass Fertilizer with Nitrogen under Grazing. Semin. Ciências Agrárias 2013, 34, 2415. [Google Scholar] [CrossRef]
- Ekinci, M.; Esringü, A.; Dursun, A.; Yildirim, E.; Turan, M.; Karaman, M.R.; Arjumend, T. Growth, Yield, and Calcium and Boron Uptake of Tomato (Lycopersicon esculentum L.) and Cucumber (Cucumis sativus L.) Asaffected by Calcium and Boron Humate Application in Greenhouse Conditions. Turk. J. Agric. For. 2015, 39, 613–632. [Google Scholar] [CrossRef]
- Kalbitz, K.; Kaiser, K. Contribution of Dissolved Organic Matter to Carbon Storage in Forest Mineral Soils. J. Plant Nutr. Soil Sci. 2008, 171, 52–60. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil | Sandy Entisol | Red-Yellow Oxisol | Red Oxisol |
|---|---|---|---|
| Clay (g kg−1) | 70 | 230 | 625 |
| Silt (g kg−1) | 10 | 35 | 95 |
| Sand (g kg−1) | 920 | 735 | 280 |
| Texture | Sandy | Medium texture | Clayey |
| pH | 4.9 | 4.8 | 4.1 |
| SOM (g kg−1) | 9 | 16 | 31.7 |
| Al3+ (cmolc kg−1) | 0.7 | 0.5 | 1 |
| H+Al (cmolc kg−1) | 4.3 | 3.9 | 9.7 |
| Ca2+ (cmolc kg−1) | 0.2 | 0.7 | 0.2 |
| Mg2+ (cmolc kg−1) | 0.1 | 0.2 | 0.1 |
| K (mg kg−1) | 47.8 | 1.4 | 1.8 |
| Fe (mg kg−1) | 151 | 59.2 | 89.6 |
| Zn (mg kg−1) | 0.8 | 0.7 | 0.3 |
| Cu (mg kg−1) | 0.5 | 0.3 | 1.7 |
| Mn (mg kg−1) | 7.8 | 10.5 | 8.2 |
| P (mg kg−1) | 19.1 | 65.3 | 30.3 |
| S-SO42− (mg kg−1) | 4.7 | 5.7 | 6.9 |
| B (mg kg−1) | 0.1 | 0.1 | 0.1 |
| P-Rem (mg L−1) | 47 | 39 | 13 |
| pH | EC (dS m−1) | C (g kg−1) | N (g kg−1) | P (g kg−1) | K (g kg−1) | Ca (g kg−1) | Mg (g kg−1) | S (g kg−1) | Fe (mg kg−1) | Cu (mg kg−1) | Mn (mg kg−1) | Zn (mg kg−1) | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Humic acid | 9.7 | 36 | 350 | 5.2 | 0.05 | 42 | 1.4 | 2.6 | 2.8 | 2.7 | 7.6 | 21 | 76 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valenciano, M.N.; de Morais, E.G.; Rosa, S.D.; Silva, C.A. Soil Solution Properties of Tropical Soils and Brachiaria Growth as Affected by Humic Acid Concentration. Soil Syst. 2024, 8, 86. https://doi.org/10.3390/soilsystems8030086
Valenciano MN, de Morais EG, Rosa SD, Silva CA. Soil Solution Properties of Tropical Soils and Brachiaria Growth as Affected by Humic Acid Concentration. Soil Systems. 2024; 8(3):86. https://doi.org/10.3390/soilsystems8030086
Chicago/Turabian StyleValenciano, Murilo Nunes, Everton Geraldo de Morais, Sara Dantas Rosa, and Carlos Alberto Silva. 2024. "Soil Solution Properties of Tropical Soils and Brachiaria Growth as Affected by Humic Acid Concentration" Soil Systems 8, no. 3: 86. https://doi.org/10.3390/soilsystems8030086
APA StyleValenciano, M. N., de Morais, E. G., Rosa, S. D., & Silva, C. A. (2024). Soil Solution Properties of Tropical Soils and Brachiaria Growth as Affected by Humic Acid Concentration. Soil Systems, 8(3), 86. https://doi.org/10.3390/soilsystems8030086