The Reduction of Nitrogen Fertilizer Rate Shifted Soil Bacterial Community Structure in Rice Paddies

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Rice Plot Experimental Design
2.2. Soil Sampling
2.3. Determination of Soil Physiochemical Parameters
2.4. Soil Microbial Community Analyses
3. Results
3.1. Effects of N Application on Soil Physiochemical Parameters
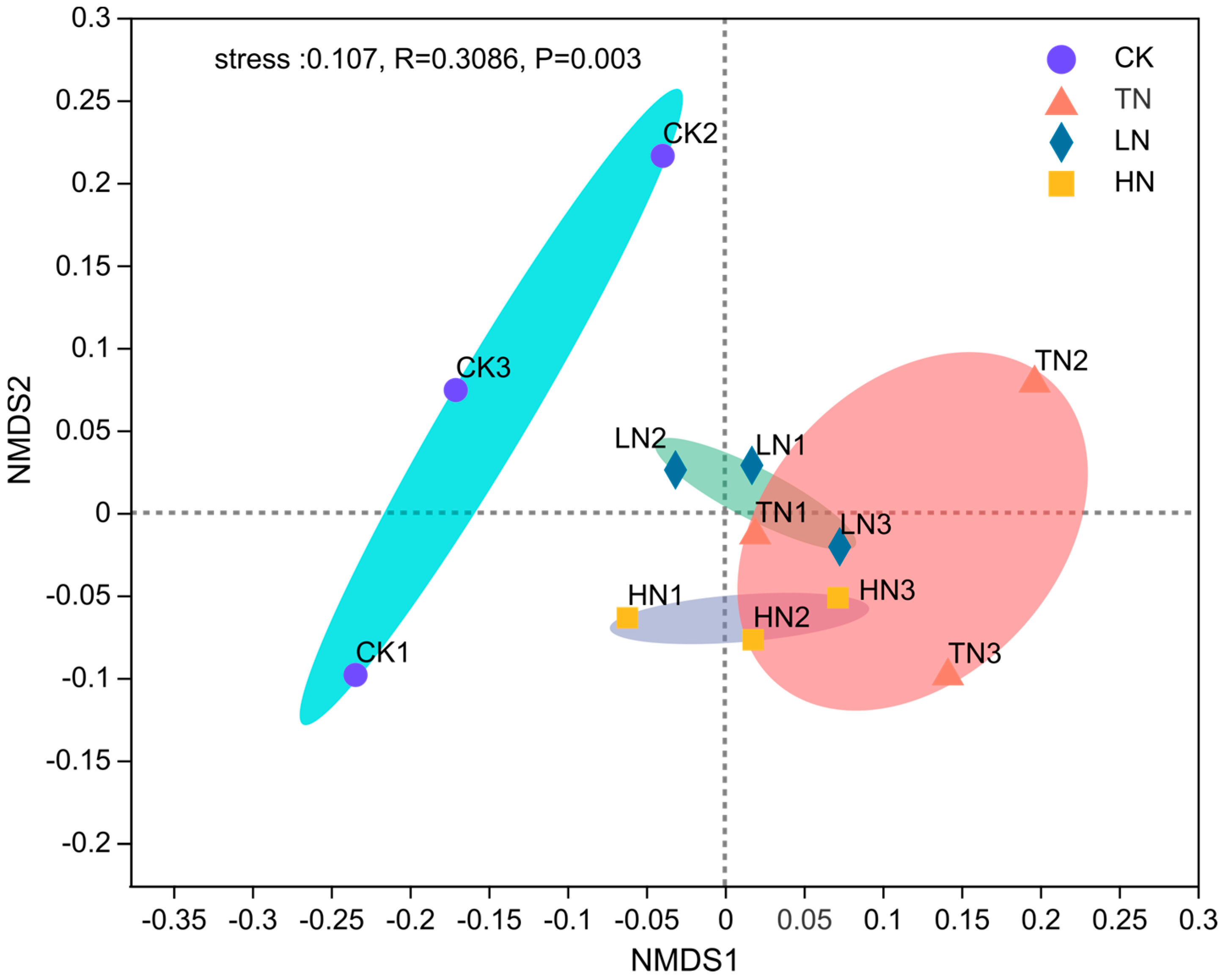
3.2. Effects of N Application on Bacterial Community Structure
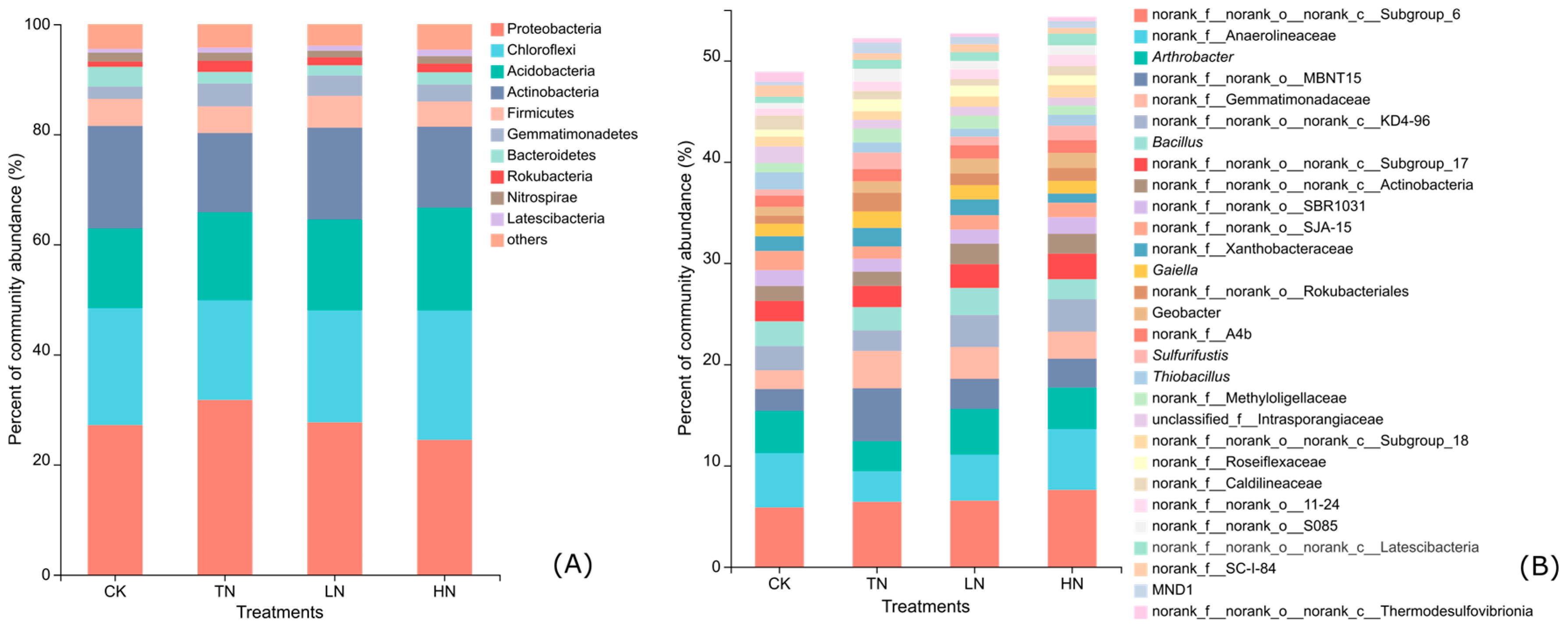
3.3. Soil Bacterial Community Composition
3.4. The Effects of Soil Properties on Bacterial Communities
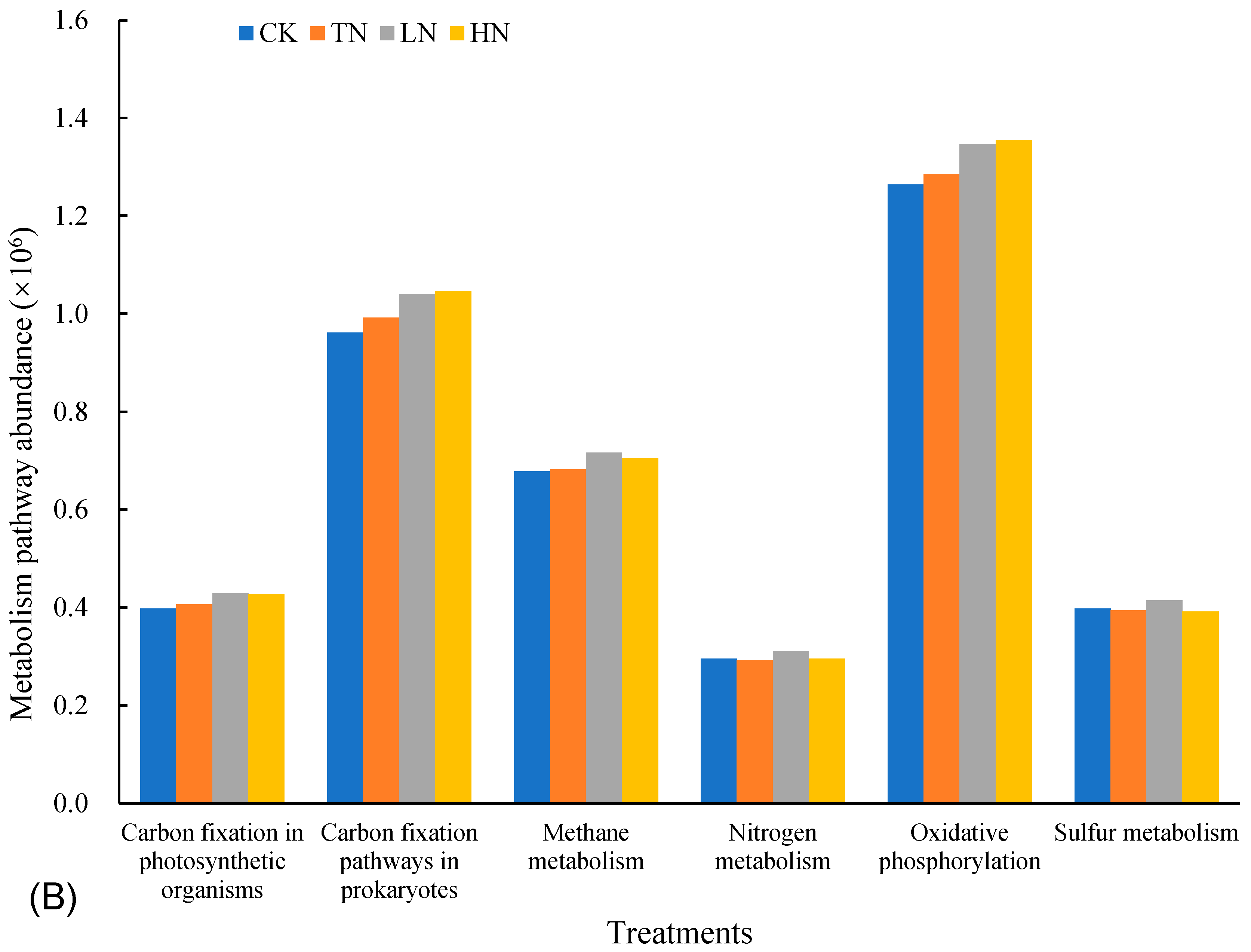
3.5. Functional Prediction of Microbial Groups
4. Discussions
4.1. Soil Physicochemical Parameters as Influenced by N Fertilizer Reduction
4.2. Shifts in Bacterial Diversity and Community Composition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, F.; Cui, Z.; Chen, X.; Ju, X.; Shen, J.; Chen, Q.; Liu, X.; Zhang, W.; Mi, G.; Fan, M.; et al. Integrated nutrient management for food security and environmental quality in China. Adv. Agron. 2012, 116, 1–40. [Google Scholar]
- Li, S.; Tian, Y.; Wu, K.; Ye, Y.; Yu, J.; Zhang, J.; Liu, Q.; Hu, M.; Li, H.; Tong, Y.; et al. Modulating plant growth-metabolism coordination for sustainable agriculture. Nature 2018, 560, 595–600. [Google Scholar] [CrossRef]
- Ju, X.T.; Xing, G.X.; Chen, X.P.; Zhang, S.L.; Zhang, L.J.; Liu, X.J.; Cui, Z.L.; Yin, B.; Christie, P.; Zhu, Z.L.; et al. Reducing environmental risk by improving N management in intensive Chinese agricultural systems. Proc. Natl. Acad. Sci. USA 2009, 106, 3041–3046. [Google Scholar] [CrossRef]
- Shi, X.; Zheng, Y.; Lei, Y.; Xue, W.; Yan, G.; Liu, X.; Cai, B.; Tong, D.; Wang, J. Air quality benefits of achieving carbon neutrality in China. Sci. Total Environ. 2021, 795, 14878. [Google Scholar] [CrossRef]
- Li, Y.; Wu, W.; Yang, J.; Cheng, K.; Smith, P.; Sun, J.; Xu, X.; Yue, Q.; Pan, G. Exploring the environmental impact of crop production in China using a comprehensive footprint approach. Sci. Total Environ. 2022, 824, 153898. [Google Scholar] [CrossRef]
- Choudhury, A.T.M.A.; Kennedy, I.R. Nitrogen fertilizer losses from rice soils and control of environmental pollution problems. Commun. Soil Sci. Plant Anal. 2005, 36, 1625–1639. [Google Scholar] [CrossRef]
- Xu, P.; Jiang, M.; Jiang, Y.; Khan, I.; Zhou, W.; Wu, H.; Wu, X.; Shaaban, M.; Lu, J.; Hu, R. Prior nitrogen fertilization stimulated N2O emission from rice cultivation season under a rapeseed-rice production system. Plant Soil 2022, 471, 685–696. [Google Scholar] [CrossRef]
- Chen, J.; Huang, Y.; Tang, Y. Quantifying economically and ecologically optimum nitrogen rates for rice production in south-eastern China. Agric. Ecosyst. Environ. 2011, 142, 195–204. [Google Scholar] [CrossRef]
- Wang, L.; Huang, D. Nitrogen and phosphorus losses by surface runoff and soil microbial communities in a paddy field with different irrigation and fertilization managements. PLoS ONE 2021, 16, e0254227. [Google Scholar] [CrossRef]
- Hua, W.; Luo, P.; An, N.; Cai, F.; Zhang, S.; Chen, K.; Yang, J.; Han, X. Manure application increased crop yields by promoting nitrogen use efficiency in the soils of 40-year soybean-maize rotation. Sci. Rep. 2020, 10, 14882. [Google Scholar] [CrossRef]
- Philippot, L.; Chenu, C.; Kappler, A.; Rillig, M.C.; Fierer, N. The interplay between microbial communities and soil properties. Nat. Rev. Microbiol. 2023, 22, 226–239. [Google Scholar] [CrossRef]
- Gorfer, M.; Borruso, L.; Deltedesco, E.; Gichuhi, E.W.; Menge, D.M.; Makihara, D.; Praeg, N.; Cesco, S.; Mimmo, T.; Merbold, L.; et al. The effect of environmental parameters and fertilization practices on yield and soil microbial diversity in a Kenyan paddy rice field. Appl. Soil Ecol. 2022, 176, 104495. [Google Scholar] [CrossRef]
- Wang, J.; Rhodes, G.; Huang, Q.; Shen, Q. Plant growth stages and fertilization regimes drive soil fungal community compositions in a wheat-rice rotation system. Biol. Fertil. Soils 2018, 54, 731–742. [Google Scholar] [CrossRef]
- Cardenas-Alegria, O.V.; Dantas, C.W.D.; Lobato, E.M.S.G.; Da Silva, S.P.; Moraes, C.D.S.; Cruz, A.C.R.; Ramos, R.T.J.; Carneiro, A.R. Influence of soil management for soybean production under microbial diversity in amazon soils. Front. Environ. Sci. 2022, 10, 1024610. [Google Scholar] [CrossRef]
- Kuypers, M.M.M.; Marchant, H.K.; Kartal, B. The microbial nitrogen-cycling network. Nat. Rev. Microbiol. 2018, 16, 263–276. [Google Scholar] [CrossRef]
- Cao, X.; Zhang, J.; Yu, Y.; Ma, Q.; Kong, Y.; Pan, W.; Wu, L.; Jin, Q. Alternate wetting-drying enhances soil nitrogen availability by altering organic nitrogen partitioning in rice-microbe system. Geoderma 2022, 424, 115993. [Google Scholar] [CrossRef]
- Ma, Q.; Qian, Y.; Yu, Q.; Cao, Y.; Tao, R.; Zhu, M.; Ding, J.; Li, C.; Guo, W.; Zhu, X. Controlled-release nitrogen fertilizer application mitigated N losses and modified microbial community while improving wheat yield and N use efficiency. Agric. Ecosyst. Environ. 2023, 349, 108445. [Google Scholar] [CrossRef]
- Xie, Y.; Wang, Z.; Cheng, X.; Qiu, R.; Hamoud, Y.A.; Hong, C.; Zong, X.; Wang, Y.; Agathokleous, E.; Guo, X. Dissecting the combined effects of cultivar, fertilization, and irrigation on rhizosphere bacterial communities and nitrogen productivity in rice. Sci. Total Environ. 2022, 835, 155534. [Google Scholar] [CrossRef]
- Fan, F.; Yang, Q.; Li, Z.; Wei, D.; Cui, X.A.; Liang, Y. Impacts of organic and inorganic fertilizers on nitrification in a cold climate soil are linked to the bacterial ammonia oxidizer community. Microb. Ecol. 2011, 62, 982–990. [Google Scholar] [CrossRef]
- Chen, Z.; Hou, H.; Zheng, Y.; Qin, H.; Zhu, Y.; Wu, J.; Wei, W. Influence of fertilisation regimes on a nosZ-containing denitrifying community in a rice paddy soil. J. Sci. Food Agric. 2012, 92, 1064–1072. [Google Scholar] [CrossRef]
- Zhu, J.; Peng, H.; Ji, X.; Li, C.; Li, S. Effects of reduced inorganic fertilization and rice straw recovery on soil enzyme activities and bacterial community in double-rice paddy soils. Eur. J. Soil Biol. 2019, 94, 103116. [Google Scholar] [CrossRef]
- Xu, P.; Jiang, M.; Khan, I.; Zhao, J.; Yang, T.; Tu, J.; Hu, R. Available nitrogen and ammonia-oxidizing archaea in soil regulated N2O emissions regardless of rice planting under a double rice crop**-fallow system. Agric. Ecosyst. Environ. 2022, 340, 108166. [Google Scholar] [CrossRef]
- Che, S.; Zhao, B.; Li, Y.; Liang, Y.; Wei, L.; Lin, Z.; Hu, S.; Bing, S. Review grain yield and nitrogen use efficiency in rice production regions in China. J. Integr. Agric. 2015, 14, 2456–2466. [Google Scholar] [CrossRef]
- Bao, S. Soil Agro-Chemistrical Analysis, 3rd ed.; China Agriculture Press: Beijing, China, 2007; pp. 268–270, 389–391. [Google Scholar]
- Dennis, K.L.; Wang, Y.; Blatner, N.R.; Wang, S.; Abdulrahman, S.; Trudeau, E.; Roers, A.; Weaver, C.T.; Lee, J.J.; Gilbert, J.A.; et al. Adenomatous polyps are driven by microbe-instigated focal inflammation and are controlled by IL-10-producing T cells. Cancer Res. 2013, 73, 5905–5913. [Google Scholar] [CrossRef]
- Wang, J.; Ma, Y.; Di, L.; Qian, X.; Wang, G. Straw incorporation with nitrogen amendment shapes bacterial community structure in an iron-rich paddy soil by altering nitrogen reserves. Microorganisms 2021, 9, 988. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Goebel, B. Taxonomic note: A place for DNA-DNA reassociation and 16S rRNA sequence analysis in the present species definition in bacteriology. Int. J. Syst. Bacteriol. 1994, 44, 846–849. [Google Scholar] [CrossRef]
- Faith, D.P.; Minchin, P.R.; Belbin, L. Compsitional dissimilarity as a robust measure of ecogical distance. Vegetatio 1987, 69, 57–68. [Google Scholar] [CrossRef]
- Louca, S.; Parfrey, L.W.; Doebeli, M. Decoupling function and taxonomy in the global ocean microbiome. Science 2016, 353, 1272–1277. [Google Scholar] [CrossRef]
- Anas, M.; Liao, F.; Verma, K.K.; Sarwar, M.A.; Mahmood, A.; Chen, Z.; Li, Q.; Zeng, X.; Liu, Y.; Li, Y. Fate of nitrogen in agriculture and environment: Agronomic, eco-physiological and molecular approaches to improve nitrogen use efficiency. Biol. Res. 2020, 53, 47. [Google Scholar] [CrossRef]
- Chen, L.; Xie, H.; Wang, G.; Yuan, L.; Qian, X.; Wang, W.; Xu, Y.; Zhang, W.; Zhang, H.; Liu, L.; et al. Reducing environmental risk by improving crop management practices at high crop yield levels. Field Crops Res. 2021, 265, 108123. [Google Scholar] [CrossRef]
- Mohana, K.M.; Pooja, S.; Govindhasamy, G.; Arasu, A.; Rajagopalan, B. Precision nitrogen management in aerobic system for maximising paddy (Oryza sativa L.) yields: A review. Crop Res. 2023, 58, 107–115. [Google Scholar]
- Bai, Z.; Huang, J.; Zhu, L.; Cao, X.; Zhu, C.; Zhong, C.; Jin, Q.; Zhang, J. Effects of N application strategies on N leaching loss in paddy soil and N use characteristics in different super hybrid rice cultivars. Paddy Water Environ. 2020, 18, 27–41. [Google Scholar] [CrossRef]
- Liu, W.; Bao, S.; Lu, Y.; Zhang, Q.; Geng, Y.; Shao, X.; Guo, L. Effects of dense planting with less nitrogen fertilization on rice yield and nitrogen use efficiency in northeast china. Int. J. Plant Prod. 2021, 15, 625–634. [Google Scholar] [CrossRef]
- Oldroyd, G.E.D.; Leyser, O. A plant’s diet, surviving in a variable nutrient environment. Science 2020, 368, 45. [Google Scholar] [CrossRef]
- Li, B.B.; Roley, S.S.; Duncan, D.S.; Guo, J.R.; Quensen, J.F.; Yu, H.Q.; Tiedje, J.M. Long-term excess nitrogen fertilizer increases sensitivity of soil microbial community to seasonal change revealed by ecological network and metagenome analyses. Soil Biol. Biochem. 2021, 160, 108349. [Google Scholar] [CrossRef]
- Wang, X.; Feng, J.; Ao, G.K.L.; Qin, W.K.; Han, M.G.; Shen, Y.W.; Liu, M.L.; Chen, Y.; Zhu, B. Globally nitrogen addition alters soil microbial community structure, but has minor effects on soil microbial diversity and richness. Soil Biol. Biochem. 2023, 179, 108980. [Google Scholar] [CrossRef]
- Marshall, D.J.; Cameron, H.J.; Loreau, H. Relationships between intrinsic population growth rate, carrying capacity and metabolism in microbial populations. ISME J. 2023, 17, 2140–2143. [Google Scholar] [CrossRef]
- Ouyang, Y.; Norton, J.M. Short-Term nitrogen fertilization affects microbial community composition and nitrogen mineralization functions in an agricultural soil. Appl. Environ. Microbiol. 2020, 86, e02278-19. [Google Scholar] [CrossRef]
- Zhao, J.; Ni, T.; Li, J.; Lu, Q.; Fang, Z.Y.; Huang, Q.W.; Zhang, R.F.; Li, R.; Shen, B.; Shen, Q.R. Effects of organic–inorganic compound fertilizer with reduced chemical fertilizer application on crop yields, soil biological activity and bacterial community structure in a rice–wheat cropping system. Appl. Soil Ecol. 2016, 99, 1–12. [Google Scholar] [CrossRef]
- Bose, H.; Sahu, R.P.; Sar, P. Impact of arsenic on microbial community structure and their metabolic potential from rice soils of West Bengal, India. Sci. Total Environ. 2022, 841, 156486. [Google Scholar] [CrossRef]
- Dai, Z.M.; Su, W.Q.; Chen, H.H.; Barberán, A.; Zhao, H.C.; Yu, M.; Yu, L.; Brookes, P.C.; Schadt, C.W.; Chang, S.X.; et al. Long-term nitrogen fertilization decreases bacterial diversity and favors the growth of Actinobacteria and Proteobactria in agro-ecosystems across the globe. Glob. Chang. Biol. 2018, 24, 3452–3461. [Google Scholar] [CrossRef] [PubMed]
- Sansupa, C.; Wahdan, S.F.M.; Hossen, S.; Disayathanoowat, T.; Wubet, T.; Purahong, W. Can We Use Functional Annotation of Prokaryotic Taxa (FAPROTAX) to Assign the Ecological Functions of Soil Bacteria? Appl. Sci. 2021, 11, 688. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | pH | EC μS cm−1 | AN mg kg−1 | NN mg kg−1 | KN mg kg−1 | AP mg kg−1 | AK mg kg−1 | OM g kg−1 |
|---|---|---|---|---|---|---|---|---|
| CK | 6.67 ± 0.07 | 92.8 ± 4.9 b | 5.02 ± 0.41 c | 3.46 ± 0.15 c | 115 ± 3.9 c | 88.8 ± 7.0 a | 149 ± 4.0 a | 21.3 ± 1.9 a |
| TN | 6.73 ± 0.13 | 104.5 ± 7.1 ab | 6.55 ± 0.41 ab | 5.54 ± 0.12 b | 140.7 ± 2.2 b | 78.3 ± 5.6 b | 126.7 ± 7.3 b | 18.1 ± 1.6 b |
| LN | 6.65 ± 0.07 | 112.9 ± 8.8 a | 5.85 ± 0.62 bc | 3.77 ± 0.63 c | 133.7 ± 2.9 b | 77.3 ± 1.1 b | 137.1 ± 11.7 ab | 20.2 ± 1.5 ab |
| HN | 6.783 ± 0.06 | 88 ± 8.7 b | 7.44 ± 0.55 a | 8.19 ± 0.18 a | 160 ± 12 a | 77.9 ± 3.1 b | 133.6 ± 9.6 ab | 18.5 ± 0.9 b |
| Treatments | Chao | ACE | Shannon | Coverage |
|---|---|---|---|---|
| CK | 5256.5 ±754.49 | 5516.6 ± 1328.4 | 6.71 ± 0.19 | 0.963 |
| TN | 5131 ± 519.9 | 5123.3 ± 530.35 | 6.83 ± 0.16 | 0.965 |
| LN | 5214.8 ± 371.76 | 5130.5 ± 349.3 | 6.82 ± 0.05 | 0.965 |
| HN | 5560.9 ± 233.72 | 5574.3 ± 250.7 | 6.99 ± 0.1 | 0.961 |
| Phylum | NN | AN | KN | AP | AK | EC | pH | OM |
|---|---|---|---|---|---|---|---|---|
| Proteobacteria | 0.09 | 0.39 | −0.06 | 0.01 | −0.02 | 0.52 | −0.01 | −0.37 |
| Chloroflexi | 0.08 | −0.05 | 0.24 | −0.17 | 0.20 | −0.32 | 0.06 | −0.46 |
| Acidobacteria | 0.24 | 0.58 * | 0.50 | −0.52 | −0.41 | 0.06 | 0.29 | −0.51 |
| Actinobacteria | −0.45 | −0.08 | −0.43 | 0.19 | 0.23 | 0.24 | −0.37 | 0.22 |
| Firmicutes | −0.36 | −0.13 | −0.33 | 0.25 | 0.33 | 0.12 | −0.40 | 0.33 |
| Gemmatimonadetes | 0.19 | 0.55 | 0.44 | −0.80 ** | −0.69 * | 0.46 | 0.03 | −0.33 |
| Bacteroidetes | −0.10 | −0.56 | −0.17 | 0.23 | 0.33 | −0.53 | 0.07 | 0.17 |
| Rokubacteria | 0.52 | 0.81 ** | 0.59 * | −0.48 | −0.39 | 0.22 | 0.42 | −0.58 * |
| Nitrospirae | 0.43 | 0.01 | 0.30 | 0.00 | 0.13 | −0.06 | 0.09 | −0.43 |
| Latescibacteria | 0.57 | 0.50 | 0.83 *** | −0.76 ** | −0.65 * | −0.15 | 0.50 | −0.57 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qian, X.; Xie, S.; Hu, R.; Zhao, W.; Gu, J.; Huang, W.; Xu, F. The Reduction of Nitrogen Fertilizer Rate Shifted Soil Bacterial Community Structure in Rice Paddies. Soil Syst. 2024, 8, 124. https://doi.org/10.3390/soilsystems8040124
Qian X, Xie S, Hu R, Zhao W, Gu J, Huang W, Xu F. The Reduction of Nitrogen Fertilizer Rate Shifted Soil Bacterial Community Structure in Rice Paddies. Soil Systems. 2024; 8(4):124. https://doi.org/10.3390/soilsystems8040124
Chicago/Turabian StyleQian, Xiaoqing, Shifan Xie, Rui Hu, Wenhui Zhao, Junfei Gu, Wujian Huang, and Fulei Xu. 2024. "The Reduction of Nitrogen Fertilizer Rate Shifted Soil Bacterial Community Structure in Rice Paddies" Soil Systems 8, no. 4: 124. https://doi.org/10.3390/soilsystems8040124
APA StyleQian, X., Xie, S., Hu, R., Zhao, W., Gu, J., Huang, W., & Xu, F. (2024). The Reduction of Nitrogen Fertilizer Rate Shifted Soil Bacterial Community Structure in Rice Paddies. Soil Systems, 8(4), 124. https://doi.org/10.3390/soilsystems8040124