
Biocatalytic Profiling of Free and Immobilized Partially Purified Alkaline Protease from an Autochthonous Bacillus aryabhattai Ab15-ES



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Partial Purification of Protease
2.3. Immobilization of Partially Purified Protease in Calcium Alginate Gel Beads
2.4. Determination of Immobilization Efficiency
2.5. Protease Assay
2.6. Characterization of Free and Immobilized Protease
2.6.1. Effect of Temperature and pH on Free and Immobilized Protease Activity
2.6.2. Effect of Temperature and pH on Free and Immobilized Protease Stability
2.6.3. Kinetics Properties of Free and Immobilized Protease
2.6.4. Storage Stability of Free and Immobilized Protease
2.6.5. Reusability of Immobilized Protease
2.6.6. Scanning Electron Microscopy of Beads with Immobilized Protease
3. Results and Discussions
3.1. Partial Purification of Protease
3.2. Immobilization of Protease in Calcium Alginate Gel Beads
3.3. Scanning Electron Microscopic Analysis
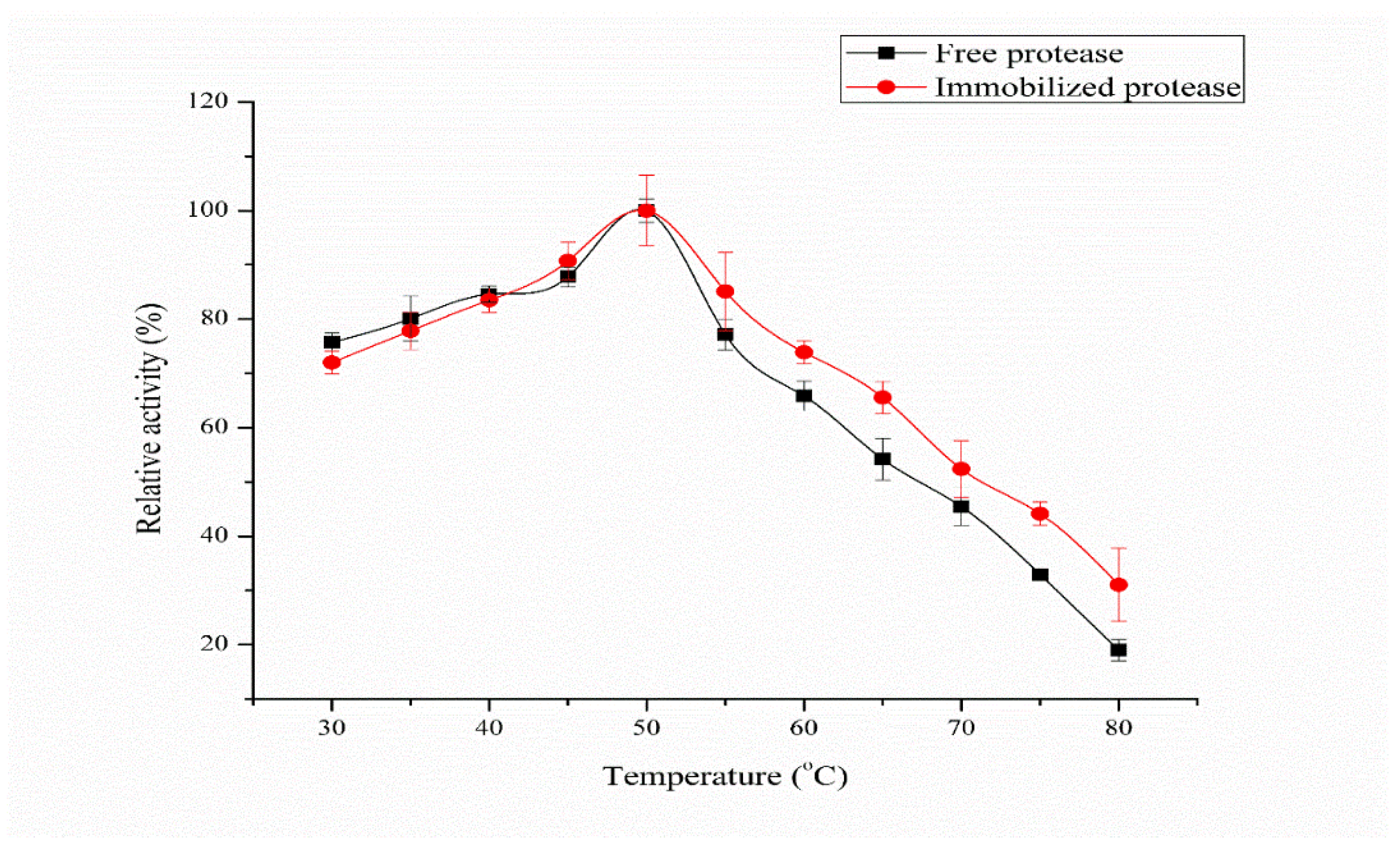
3.4. Effect of Temperature and pH on Free and Immobilized Protease Activity
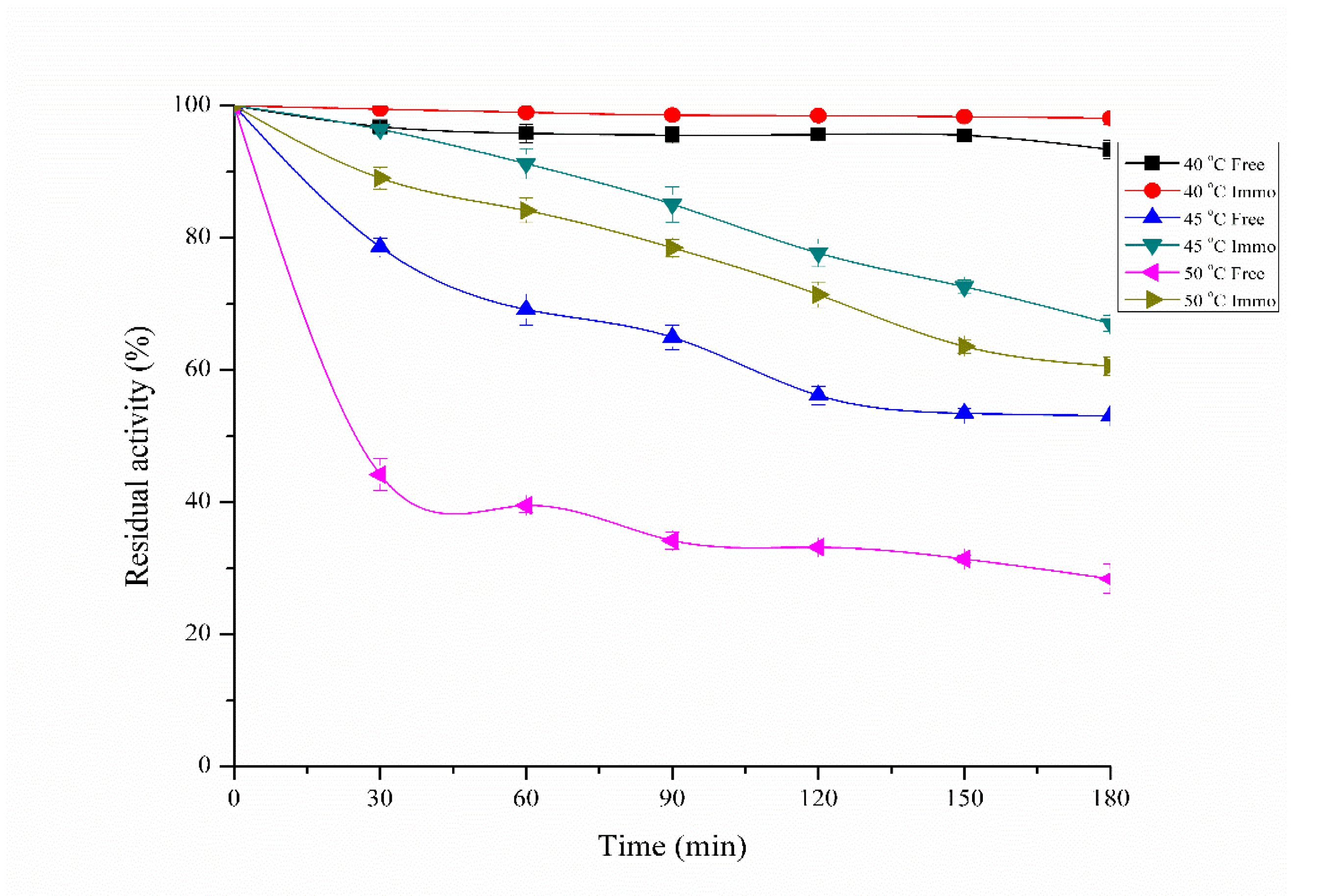
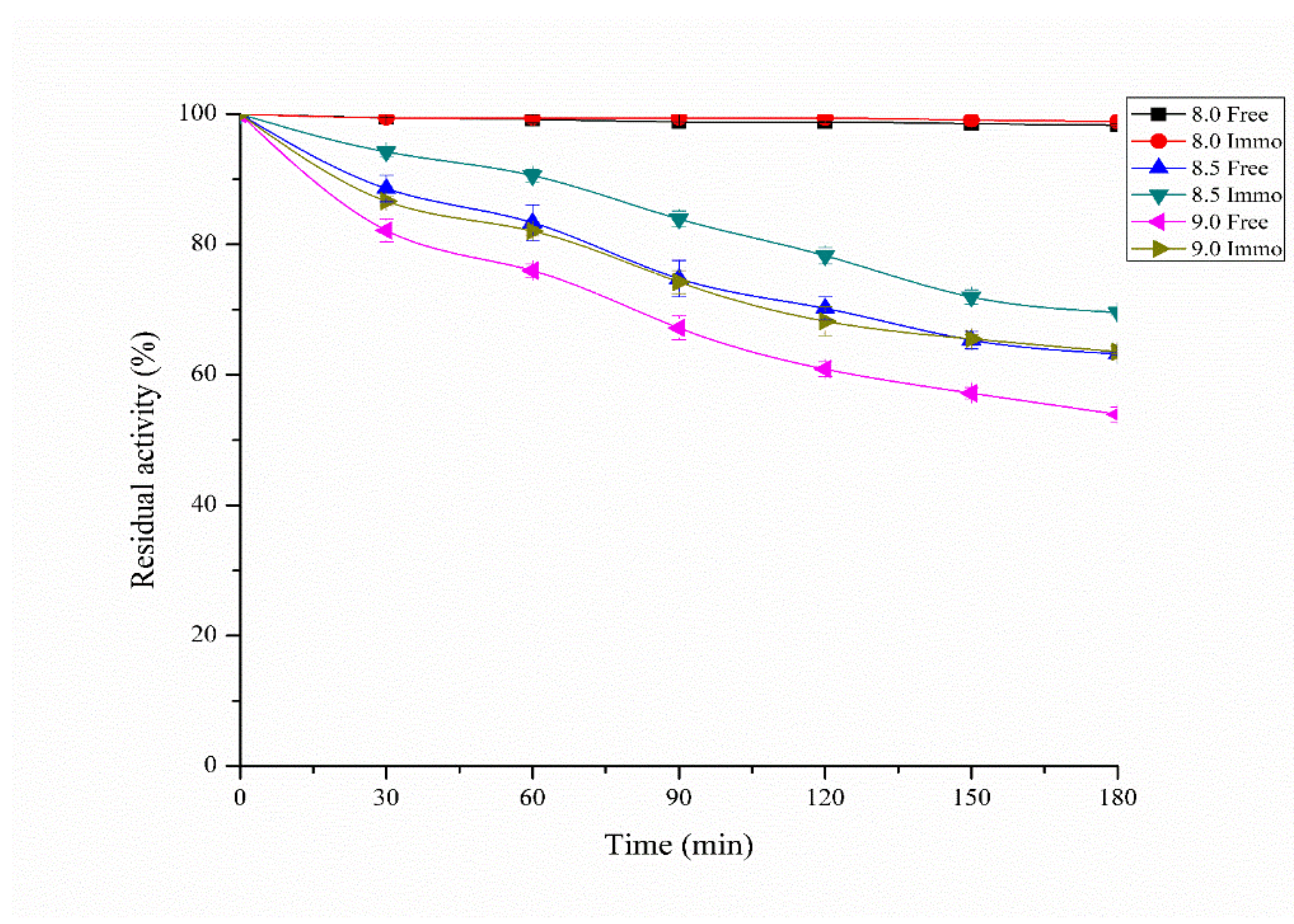
3.5. Effect of Temperature and pH on Free and Immobilized Protease Stability
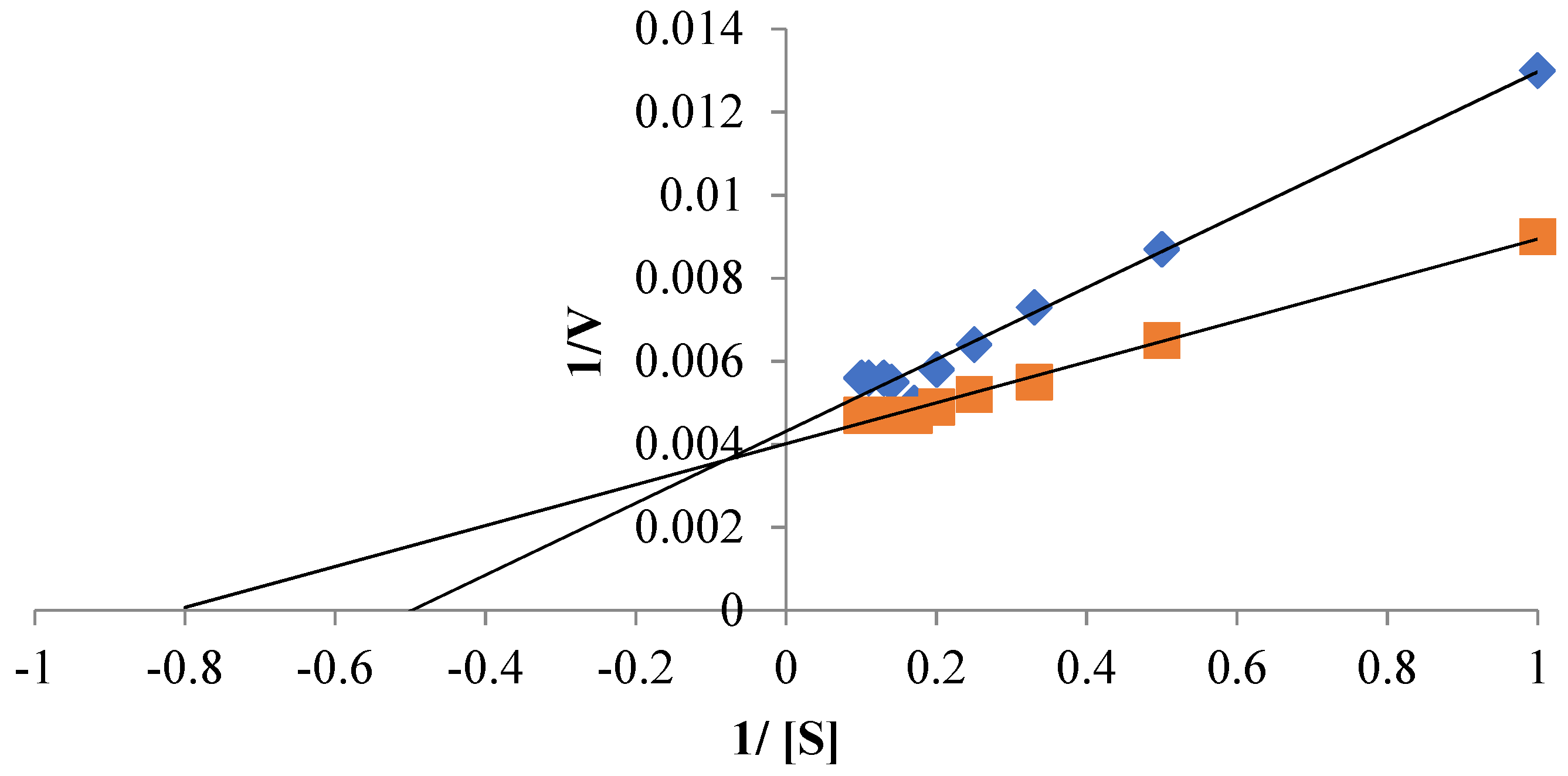
3.6. Kinetics Properties of Free and Immobilized Protease
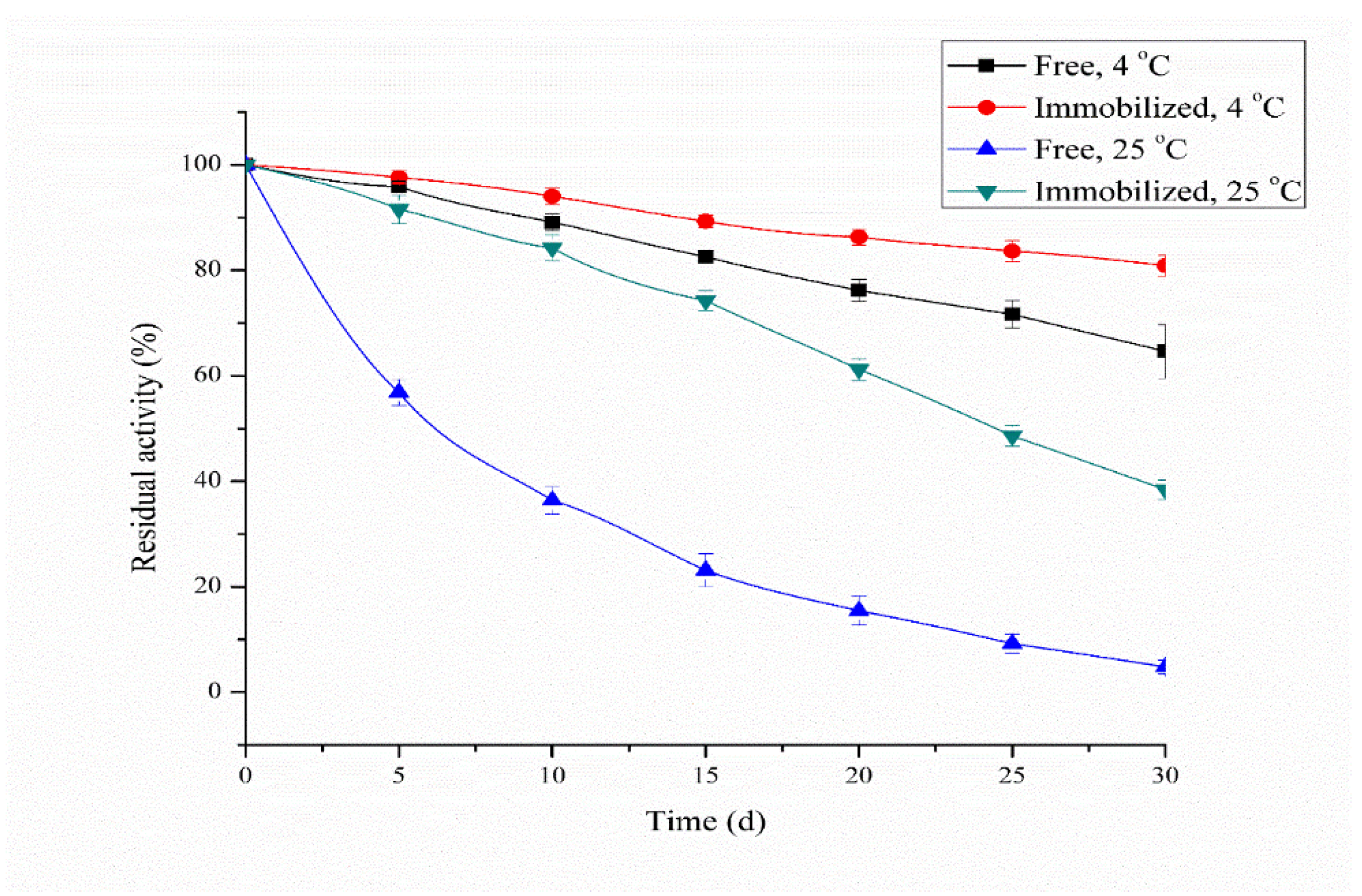
3.7. Storage Stability of Free and Immobilized Protease
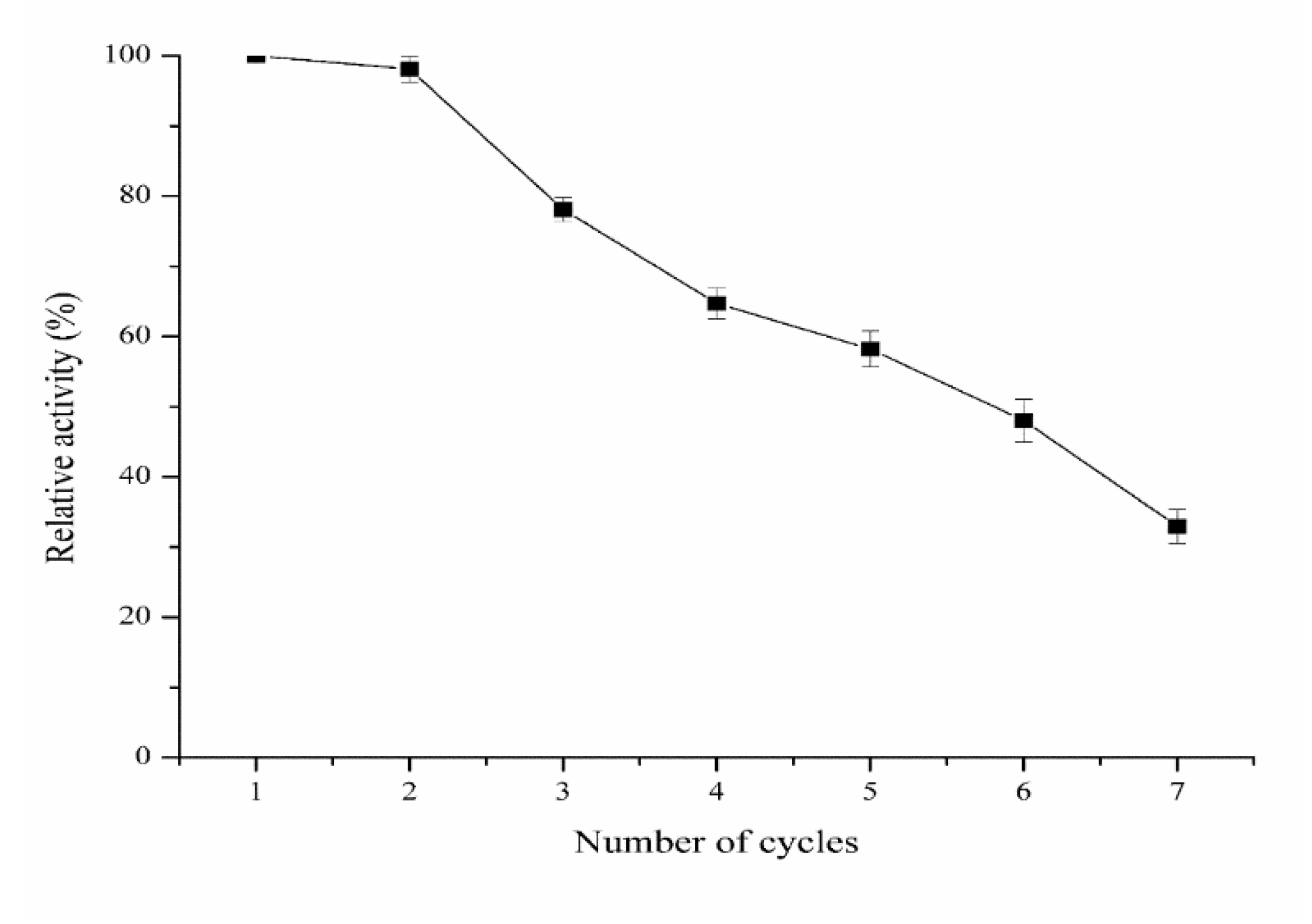
3.8. Reusability of Immobilized Protease
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ellaiah, P.; Srinivasulu, B.; Adinarayana, K. A review on microbial alkaline proteases. J. Sci. Ind. Res. 2002, 61, 690–704. [Google Scholar]
- Gupta, A.; Khare, S.K. Enhanced production and characterization of a solvent-stable protease from solvent tolerant Pseudomonas aeruginosa PseA. Enzym. Microb. Technol. 2007, 42, 11–16. [Google Scholar] [CrossRef]
- Shankar, S.; Rao, M.; Laxman, R.S. Purification and characterization of an alkaline protease by a new strain of Beauveria sp. Process Biochem. 2011, 46, 579–585. [Google Scholar] [CrossRef]
- Adetunji, A.I.; Olaniran, A.O. Statistical modelling and optimization of protease production by an autochthonous Bacillus aryabhattai Ab15-ES: A response surface methodology approach. Biocatal. Agric. Biotechnol. 2020, 24, 101528. [Google Scholar] [CrossRef]
- Sundararajan, S.; Kannan, C.N.; Chittibabu, S. Alkaline protease from Bacillus cereus VITSN04: Potential application as a dehairing agent. J. Biosci. Bioeng. 2011, 111, 128–133. [Google Scholar] [CrossRef]
- Zambare, V.; Nilegaonkar, S.; Kanekar, P. A novel extracellular protease from Pseudomonas aeruginosa MCMB-327: Enzyme production and its partial characterization. New Biotechnol. 2011, 28, 173–181. [Google Scholar] [CrossRef]
- Annamalai, N.; Rajeswari, M.V.; Balasubramanian, T. Extraction, purification and application of thermostable and halostable alkaline protease from Bacillus alveayuensis CAS 5 using marine wastes. Food Bioprod. Process. 2014, 92, 335–342. [Google Scholar] [CrossRef]
- Guleria, S.; Walia, A.; Chauhan, A.; Shirkot, C.K. Immobilization of Bacillus amyloliquefaciens SP1 and its alkaline protease in various matrices for effective hydrolysis of casein. 3 Biotech 2016, 6, 208. [Google Scholar] [CrossRef] [Green Version]
- Deng, A.H.; Wu, J.; Zhang, Y.; Zhang, G.Q.; Wen, T.Y. Purification and characterization of a surfactant-stable high-alkaline protease from Bacillus sp. B001. Bioresour. Technol. 2010, 101, 7100–7106. [Google Scholar] [CrossRef]
- Jisha, V.N.; Smitha, R.B.; Pradeep, S.; Sreedevi, S.; Unni, K.N.; Sajith, S.; Priji, P.; Josh, M.S.; Benjamin, S. Versatility of microbial proteases. Adv. Enzym. Res. 2013, 1, 39–51. [Google Scholar] [CrossRef]
- Sawant, R.; Nagendran, S. Protease: An enzyme with multiple industrial applications. World J. Pharm. Pharm. Sci. 2014, 3, 568–579. [Google Scholar]
- Herman, R.A.; Ayepa, E.; Zhang, W.-X.; Li, Z.-N.; Zhu, X.; Ackah, M.; Yuan, S.-S.; You, S.; Wang, J. Molecular modification and biotechnological applications of microbial aspartic proteases. Crit. Rev. Biotechnol. 2023. [Google Scholar] [CrossRef] [PubMed]
- Adetunji, A.I.; Olaniran, A.O. Treatment of lipid-rich wastewater using a mixture of free or immobilized bioemulsifier and hydrolytic enzymes from indigenous bacterial isolates. Desalination Water Treat. 2018, 132, 274–280. [Google Scholar] [CrossRef] [Green Version]
- Rao, M.B.; Tanksale, A.M.; Ghatge, M.S.; Deshpande, V.V. Molecular and biotechnological aspects of microbial proteases. Microbiol. Mol. Biol. Rev. 1998, 62, 597–635. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.; Beg, Q.K.; Khan, S.; Chauhan, B. An overview of fermentation, downstream processing and properties of microbial alkaline proteases. Appl. Microbiol. Biotechnol. 2002, 60, 381–395. [Google Scholar] [PubMed]
- Kumar, C.G.; Takagi, H. Microbial alkaline proteases: From a bioindustrial viewpoint. Biotechnol. Adv. 1999, 17, 561–594. [Google Scholar] [CrossRef]
- Haddar, A.; Bougatef, A.; Agrebi, R.; Sellami-Kamoun, A.; Nasri, M. A novel surfactant-stable alkaline serine-protease from a newly isolated Bacillus mojavensis A21: Purification and characterization. Process Biochem. 2009, 44, 29–35. [Google Scholar] [CrossRef]
- Raval, V.R.; Pillai, S.; Rawal, C.M.; Singh, S.P. Biochemical and structural characterization of a detergent-stable serine alkaline protease from seawater haloalkaliphilic bacteria. Process Biochem. 2014, 49, 955–962. [Google Scholar] [CrossRef]
- Tang, X.Y.; Wu, B.; Ying, H.J.; He, B.F. Biochemical properties and potential applications of a solvent-stable protease from a highly-yield protease producer Pseudomonas aeruginosa PT 121. Appl. Biochem. Biotechnol. 2010, 160, 1017–1031. [Google Scholar] [CrossRef]
- Devi, P.R.; VijayaRaghavan, P.; Vasudheven, I.; Joshua, L.; VijaKumar, M. Purification and characterization of protease from Rhizopus oligosporus. Int. J. Biol. Technol. 2011, 2, 46–49. [Google Scholar]
- Mugdha, A.; Usha, M. Enzymatic treatment of wastewater containing dyestuffs using different delivery systems. Sci. Rev. Chem. Commun. 2012, 2, 31–40. [Google Scholar]
- Karam, J.; Nicell, J.A. Potential applications of enzymes in waste treatment. J. Chem. Technol. Biotechnol. 1997, 69, 141–153. [Google Scholar] [CrossRef]
- Anwar, A.; Ul Qader, S.A.; Raiz, A.; Iqbal, S.; Azhar, A. Calcium alginate: A support material for immobilization of proteases from newly isolated strain of Bacillus subtilis KIBGE-HAS. World Appl. Sci. J. 2009, 7, 1281–1286. [Google Scholar]
- Kumar, D.; Kumar, V.; Verma, A.K.; Singh, P.; Dubey, A. Characterization and immobilization of partially purified alkaline protease extracted from rhizospheric soil bacterium, Bacillus megaterium strain EN-2 and Bacillus subtilis strain EN-3. Afr. J. Microbiol. Res. 2014, 8, 33–39. [Google Scholar]
- Sahin, S.; Ozmen, I.; Kir, E. Purification, immobilization and characterization of protease from local Bacillus subtilis M-11. Asia-Pac. J. Chem. Eng. 2015, 10, 241–247. [Google Scholar] [CrossRef]
- Massolini, G.; Calleri, E. Immobilized trypsin systems coupled on-line to separation methods: Recent developments and analytical applications. J. Sep. Sci. 2005, 28, 7–21. [Google Scholar] [CrossRef]
- Adetunji, A.I.; Olaniran, A.O. Immobilization and characterization of lipase from an indigenous Bacillus aryabhattai SE3-PB isolated from lipid-rich wastewater. Prep. Biochem. Biotechnol. 2018, 48, 898–905. [Google Scholar] [CrossRef]
- Hasirci, N.; Aksoy, S.; Tumturk, H. Activation of poly (dimer acid-co-alkyl polyamine) particles for covelent immobilization of α-amylase. React. Funct. Polym. 2006, 66, 1546–1551. [Google Scholar] [CrossRef]
- Gangadharan, D.; Nampoothiri, K.M.; Sivaramakrishnan, S.; Pandey, A. Immobilized bacterial α-amylase for effective hydrolysis of raw and soluble starch. Food Res. Int. 2009, 42, 436–442. [Google Scholar] [CrossRef]
- Adetunji, A.I. Treatment of Lipid-Rich Wastewater Using Free and Immobilized Bioemulsifier and Hydrolytic Enzymes from Indigenous Bacterial Isolates. Ph.D. Thesis, University of KwaZulu-Natal, Durban, South Africa, 2017. [Google Scholar]
- Bonine, B.M.; Polizelli, P.P.; Bonilla-Rodriguez, G.O. Immobilization of a plant lipase from Pachira aquatica in alginate and alginate/PVA beads. Enzym. Res. 2014, 2014, 7. [Google Scholar] [CrossRef] [Green Version]
- Datta, S.; Rene, C.L.; Rajaram, Y.R.S. Enzyme immobilization: An overview on techniques and support materials. 3 Biotech 2013, 3, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamad, N.R.; Marzuki, N.H.C.; Buang, N.A.; Huyop, F.; Abdul Wahab, R. An overview of technologies for immobilization of enzymes and surface analysis techniques for immobilized enzymes. Biotechnol. Biotechnol. Equip. 2015, 29, 205–220. [Google Scholar] [CrossRef] [PubMed]
- Tumturk, H.; Demirel, G.; Altinok, H.; Aksoy, S.; Hasirci, N. Immobilization of glucose isomerase in surface-modified alginate gel beads. J. Food Biochem. 2008, 32, 234–246. [Google Scholar] [CrossRef]
- Ölcer, Z.; Tanriseven, A. Co-immobilization of dextransucrase and dextranase in alginate. Process Biochem. 2010, 45, 1645–1651. [Google Scholar] [CrossRef]
- Jamal, F.; Qidwai, T.; Singh, D.; Pandey, P.K. Biocatalytic activity of immobilized pointed gourd (Trichosanthes dioica) peroxidase-concanavalin A complex on calcium alginate pectin gel. J. Mol. Catal. B Enzym. 2012, 74, 125–131. [Google Scholar] [CrossRef]
- Mahajan, R.; Gupta, V.K.; Sharma, J. Comparison and suitability of gel matrix for entrapping higher content of enzymes for commercial applications. Indian J. Pharm. Sci. 2010, 72, 223–228. [Google Scholar]
- Buchholz, K.; Kasche, V.; Bornscheuer, U.T. Characterization of immobilized catalyst. In Biocatalysts and Enzyme Technology; Buchholz, K., Kasche, V., Bornscheuer, U.T., Eds.; Boschstraẞe: Weinheim, Germany, 2012; pp. 411–448. [Google Scholar]
- Blandino, A.; Macias, M.; Cantero, D. Glucose oxidase release from calcium alginate gel capsules. Enzym. Microb. Technol. 2000, 27, 319–324. [Google Scholar] [CrossRef]
- Sabotic, J.; Kos, J. Microbial and fungal protease inhibitors- current and potential applications. Appl. Microbiol. Biotechnol. 2012, 93, 1351–1375. [Google Scholar] [CrossRef]
- Gupta, R.; Beg, Q.K.; Lorenz, P. Bacterial alkaline proteases: Molecular approaches and industrial applications. Appl. Microbiol. Biotechnol. 2002, 59, 15–32. [Google Scholar]
- Devi, M.K.; Banu, A.R.; Gnanaprabhal, G.R.; Pradeep, B.V.; Palaniswamy, M. Purification, characterization of alkaline protease enzyme from native isolate Aspergillus niger and its compatibility with commercial detergents. Indian J. Sci. Technol. 2008, 1, 1–6. [Google Scholar] [CrossRef]
- Vijayaraghavan, P.; Vincent, S.G.P. Cow dung as a novel, inexpensive substrate for the production of a halo-tolerant alkaline protease from Halomonas sp. PVI for eco-friendly applications. Biochem. Eng. J. 2012, 69, 57–60. [Google Scholar] [CrossRef]
- Coêlho, D.F.; Saturnino, T.P.; Fernandes, F.F.; Mazzola, P.G.; Silveira, E.; Tambourgi, E.B. Azocasein substrate for determinaXtion of proteolytic activity: Reexamining a traditional method using bromelain samples. Biomed. Res. Int. 2016, 2016, 8409183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aissaoui, N.; Marzouki, M.N.; Abidi, F. Purification and biochemical characterization of a novel intestinal protease from Scorpaena notata. Int. J. Food Prop. 2017, 20, S2151–S2165. [Google Scholar]
- Iversen, S.L.; Jǿrgensen, M.H. Azocasein assay for alkaline protease in complex fermentation broth. Biotechnol. Tech. 1995, 9, 573–576. [Google Scholar] [CrossRef]
- Wilson, P.; Remigio, Z. Production and characterization of protease enzyme produced by a novel moderate thermophilic bacterium (EP1001) isolated from an alkaline hot spring, Zimbabwe. Afr. J. Microbiol. Res. 2012, 6, 5542–5551. [Google Scholar]
- Jaouadi, B.; Ellouz-Chaabouni, S.; Rhimi, M.; Bejar, S. Biochemical and molecular characterization of a detergent-stable serine alkaline protease from Bacillus pumilus CBS with high catalytic efficiency. Biochimie 2008, 90, 1291–1305. [Google Scholar] [CrossRef]
- Sellami-Kamoun, A.; Haddar, A.; Ali, N.E.; Ghorbel-Frikha, B.; Kanoun, S.; Nasri, M. Stability of thermostable alkaline protease from Bacillus licheniformis RP1 in commercial solid laundry detergent formulations. Microbiol. Res. 2008, 163, 299–306. [Google Scholar] [CrossRef]
- Mothe, T.; Sultanpuran, V.R. Production, purification and characterization of a thermotolerant alkaline serine protease from a novel species of Bacillus caseinilyticus. 3 Biotech 2016, 6, 53. [Google Scholar] [CrossRef] [Green Version]
- Ghafoor, A.; Hasnain, S. Purification and characterization of an extracellular protease from Bacillus subtilis EAG-2 strain isolated from ornamental plant nursery. Pol. J. Microbiol. 2010, 59, 107–112. [Google Scholar] [CrossRef]
- Maruthiah, T.; Esakkiraj, P.; Prabakaran, G.; Palavesam, A.; Immanuel, G. Purification and characterization of moderately halophilic alkaline serine protease from marine Bacillus subtilis AP-MSU 6. Biocatal. Agric. Biotechnol. 2013, 2, 116–119. [Google Scholar] [CrossRef]
- Rajkumar, R.; Jayappriyan, K.R.; Rengasamy, R. Purification and characterization of a protease produced by Bacillus megaterium RRM2: Application in detergent and dehairing industries. J. Basic Microbiol. 2011, 51, 614–624. [Google Scholar] [CrossRef]
- Benkiar, A.; Nadia, Z.J.; Badis, A.; Rebzani, F.; Soraya, B.T.; Rekik, H.; Naili, B.; Ferradji, F.Z.; Bejar, S.; Jaouadi, B. Biochemical and molecular characterization of a thermos- and detergent-stable alkaline serine keratinolytic protease from Bacillus circulans strain DZ100 for detergent formulations and feather-biodegradation process. Int. Biodeterior. Biodegrad. 2013, 83, 129–138. [Google Scholar] [CrossRef]
- Joo, H.S.; Kumar, C.G.; Park, G.C.; Paik, S.R.; Chang, C.S. Bleach-resistant alkaline protease produced by a Bacillus sp. isolated from the Korean polychaete, Periserrula leucophryna. Process Biochem. 2004, 39, 1441–1447. [Google Scholar] [CrossRef]
- Sharma, K.M.; Kumar, R.; Vats, S.; Gupta, A. Production, partial purification and characterization of alkaline protease from Bacillus aryabhattai K3. Int. J. Adv. Pharm. Biol. Chem. 2014, 3, 290–298. [Google Scholar]
- Pathak, A.P.; Rathod, M.G.; Mahabole, M.P.; Khairnar, R.S. Enhanced catalytic activity of Bacillus aryabhattai P1 protease by modulation with nanoactivator. Heliyon 2020, 6, e04053. [Google Scholar] [CrossRef]
- Sheldon, R.A.; van Pelt, S. Enzyme immobilization in biocatalysis: Why, what, and how. Chem. Soc. Rev. 2013, 42, 6223–6235. [Google Scholar] [CrossRef] [Green Version]
- Secades, P.; Guijarro, J.A. Purification and characterization of an extracellular protease from the fish pathogen Yersinia ruckeri and effect of culture conditions on production. Appl. Environ. Microbiol. 1999, 65, 3969–3975. [Google Scholar] [CrossRef] [Green Version]
- Lowry, O.H.; Rosebrough, N.; Farr, A.L.; Rondall, R.L. Protein measurement with the Folin-phenol reagent. J. Biol. Chem. 1951, 193, 265–273. [Google Scholar] [CrossRef]
- Lineweaver, H.; Burk, D. The determination of enzyme dissociation constants. J. Am. Chem. Soc. 1934, 56, 658–666. [Google Scholar] [CrossRef]
- Purwanto, M.G.M. The role and efficiency of ammonium sulphate precipitation in purification process of papain crude extract. Procedia Chem. 2016, 18, 127–131. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.; Cha, D.; Cheng, G.; Peng, Q.; Shen, P. Purification and characterization of a thermostable protease from a newly isolated Geobacillus sp. YMTC 1049. Enzym. Microb. Technol. 2007, 40, 1592–1597. [Google Scholar] [CrossRef]
- Geethanjali, S.; Subash, A. Optimization and immobilization of purified Labeo rohita visceral protease by entrapment method. Enzym. Res. 2013, 2013, 874050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farag, A.M.; Hassan, M.A. Purification, characterization and immobilization of a keratinase from Aspergillus oryzae. Enzym. Microb. Technol. 2004, 34, 85–93. [Google Scholar] [CrossRef]
- Gupta, K.A.; Kumar, V.; Dubey, A.; Verma, A.K. Kinetic characterization and effect of immobilized thermostable β-glucosidase in alginate gel beads on sugarcane juice. ISRN Biochem. 2014, 2014, 178498. [Google Scholar]
- Thu, T.T.M.; Krasaekoopt, W. Encapsulation of protease from Aspergillus oryzae and lipase from Thermomyces lanuginoseus using alginate and different copolymer types. Agric. Nat. Res. 2016, 50, 155–161. [Google Scholar]
- Sun, J.; Liu, J.; Liu, Y.; Li, Z. Optimization of entrapping conditions of nitrifying bacteria and selection of entrapping agent. In Proceedings of the 2nd International Conference on Environmental Science and Technology (IPCBEE), Singapore, 26–28 February 2011; IACSIT Press: Chengdu, China, 2011. [Google Scholar]
- Konsoula, Z.; Kyriakides, M.L. Thermostable α-amylase production by Bacillus subtilis entrapped in calcium alginate gel capsules. Enzym. Microb. Technol. 2006, 39, 690–696. [Google Scholar] [CrossRef]
- Elibol, M.; Moreira, A.R. Production of extracellular alkaline protease by immobilization of the marine bacterium Teredinobacter turnirae. Process Biochem. 2003, 38, 1445–1450. [Google Scholar] [CrossRef]
- Zhan, J.F.; Jiang, S.T.; Pan, L.J. Immobilization of phospholipase A1 using polyvinyl alcohol-alginate matrix and evaluation of the effects of immobilization. Braz. J. Chem. Eng. 2013, 30, 721–728. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, G.; Singh, K.; Talat, M.; Srivastava, O.N.; Kayastha, A.M. Functionalized graphene sheets as immobilization matrix for Fenugreek β-amylase: Enzyme kinetics and stability studies. PLoS ONE 2014, 9, e113408. [Google Scholar] [CrossRef] [Green Version]
- He, S.; Song, D.; Chen, M.; Cheng, H. Immobilization of lipases on magnetic collagen fibers and its applications for short-chain ester synthesis. Catalysts 2017, 7, 178. [Google Scholar] [CrossRef] [Green Version]
- Tükel, S.S.; Hürrem, F.; Yildirim, D.; Alptekin, O. Preparation of crosslinked enzyme aggregates (CLEA) of catalase and its characterization. J. Mol. Catal. B Enzym. 2013, 97, 252–257. [Google Scholar] [CrossRef]
- Saravanakumar, T.; Palvannan, T.; Kim, D.H.; Park, S.M. Optimized immobilization of peracetic acid producing recombinant acetyl xylan esterase on chitosan coated-Fe3O4 magnetic nanoparticles. Proc. Biochem. 2014, 49, 1920–1928. [Google Scholar] [CrossRef]
- Xiao, A.; Xu, C.; Lin, Y.; Ni, H.; Zhu, Y.; Cai, H. Preparation and characterization of κ-carrageenase immobilized onto magnetic iron oxide nanoparticles. Electr. J. Biotechnol. 2016, 19, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Quioroga, E.; Illanes, C.O.; Ochoa, N.A.; Barberis, S. Performance improvement of araujiain, a cysteine phytoprotease by immobilization within calcium alginate beads. Process Biochem. 2011, 46, 1029–1034. [Google Scholar] [CrossRef]
- Ibrahim, A.S.S.; Al-Salamah, A.A.; El-Toni, A.M.; Almaary, K.S.; El-Tayeb, M.A.; Elbadawi, Y.B.; Antranikian, G. Enhancement of alkaline protease activity and stability via covalent immobilization onto hollow core-mesoporous shell silica nanospheres. Int. J. Mol. Sci. 2016, 17, 184. [Google Scholar] [CrossRef] [Green Version]
- Mateo, C.; Palomo, J.M.; Fernandez-Lorente, G.; Guisan, J.M.; Fernandez-Lafuente, R. Improvement of enzyme activity, stability and selectivity via immobilization techniques. Enzym. Microb. Technol. 2007, 40, 1451–1463. [Google Scholar] [CrossRef]
- Ranjbakhsh, E.; Bordbar, A.K.; Abbasi, M.; Khosropour, A.R.; Shams, E. Enhancement of stability and catalytic activity of immobilized lipase on silica-coated modified magnetite nanoparticles. Chem. Eng. J. 2012, 179, 272–276. [Google Scholar] [CrossRef]
- Ahmed, S.A.; Al-domany, R.A.; El-Shayeb, N.M.A.; Radwan, H.H.; Saleh, A. Optimization, immobilization of extracellular alkaline protease and characterization of its enzymatic properties. Res. J. Agric. Biol. Sci. 2008, 4, 434–446. [Google Scholar]
- Zanphorlin, L.M.; Facchini, F.D.A.; Vasconcelos, F.; Bonugli-Santos, R.C.; Rodrigues, A.; Sette, L.D.; Gomes, E.; Bonilla-Rodrigues, G.O. Production, partial characterization and immobilization in alginate beads of an alkaline protease from a new thermophilic fungus Myceliophthora sp. J. Microbiol. 2010, 48, 331–336. [Google Scholar] [CrossRef]
- Danial, E.N.; Hamza, A.H.; Mahmoud, R.H. Characteristics of immobilized urease on grafted alginate bead systems. Braz. Arch. Biol. Technol. 2015, 58, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Su, R.; Shi, P.; Zhu, M.; Hong, F.; Li, D. Studies on the properties of graphene oxide-alkaline protease bio-composites. Bioresour. Technol. 2012, 115, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.-L.; Huang, Y.-M.; Li, X.-H.; Xu, P.; Wu, H.; Li, N.; Lou, W.-Y.; Zong, M.-H. Preparation and characterization of immobilized lipase from Pseudomonas cepacia onto magnetic cellulose nanocrystals. Sci. Rep. 2016, 6, 20420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Queiroz, A.; Passes, E.; Alves, S.; Silva, G.; Higa, O.; Vítolo, M. Alginate-poly (vinyl alcohol) core-shell microspheres for lipase immobilization. J. Appl. Polym. Sci. 2006, 102, 1553–1560. [Google Scholar] [CrossRef]
- Çevik, E.; Şenel, M.; Abasiyanik, M.F. Immobilization of urease on copper chelated EC-Tribeads and reversible adsorption. Afr. J. Biotechnol. 2011, 10, 6590–6597. [Google Scholar]
- Yahşi, A.; Şahin, F.; Demirel, G.; Tümtürk, H. Binary immobilization of tyrosinase by using alginate gel beads and poly (acrylamide-co-acrylic acid) hydrogels. Int. J. Biol. Macromol. 2005, 36, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Bhushan, B.; Pal, A.; Jain, V. Improved enzyme catalytic characteristics upon glutaraldehyde cross-linking of alginate entrapped xylanase isolated from Aspergillus flavus MTCC 9390. Enzym. Res. 2015, 2015, 210784. [Google Scholar] [CrossRef] [Green Version]
- Li, L.-J.; Xia, W.-J.; Ma, G.-P.; Chen, Y.-L.; Ma, Y.-Y. A study on the enzymatic properties and reuse of cellulase immobilized with carbon nanotubes and sodium alginate. AMB Express 2019, 9, 112. [Google Scholar] [CrossRef]
- Verma, M.L.; Naebe, M.; Barrow, C.J.; Puri, M. Enzyme immobilization on amino-functionalized multi-walled carbon nanaotubes: Structural and biocatalytic characterization. PLoS ONE 2013, 10, e73642. [Google Scholar]
- Pithawala, K.; Mishra, N.; Bahadur, A. Immobilization of urease in alginate, paraffin and lac. J. Serb. Chem. Soc. 2010, 75, 175–183. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Step | Total Activity (U) | Total Protein (mg) | Specific Activity (U/mg) | Recovery (%) | Purification Fold |
|---|---|---|---|---|---|
| Crude enzyme | 24,784 | 190.31 | 130.23 | 100 | 1.00 |
| (NH4)2SO4 precipitation (60%) | 15,114 | 52.90 | 285.71 | 60.98 | 2.19 |
| Sodium Alginate Concentration (%) | Immobilization Efficiency (%) |
|---|---|
| 1 | 22.78 ± 2.07 |
| 2 | 68.76 ± 2.42 |
| 3 | 53.90 ± 3.86 |
| 4 | 45.35 ± 1.62 |
| 5 | 39.74 ± 0.88 |
| CaCl2 Concentration (M) | Immobilization Efficiency (%) |
|---|---|
| 0.1 | 46.81 ± 2.41 |
| 0.2 | 62.96 ± 0.58 |
| 0.3 | 71.06 ± 1.30 |
| 0.4 | 55.92 ± 2.40 |
| 0.5 | 42.87 ± 1.78 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adetunji, A.I.; Olaniran, A.O. Biocatalytic Profiling of Free and Immobilized Partially Purified Alkaline Protease from an Autochthonous Bacillus aryabhattai Ab15-ES. Reactions 2023, 4, 231-245. https://doi.org/10.3390/reactions4020013
Adetunji AI, Olaniran AO. Biocatalytic Profiling of Free and Immobilized Partially Purified Alkaline Protease from an Autochthonous Bacillus aryabhattai Ab15-ES. Reactions. 2023; 4(2):231-245. https://doi.org/10.3390/reactions4020013
Chicago/Turabian StyleAdetunji, Adegoke Isiaka, and Ademola Olufolahan Olaniran. 2023. "Biocatalytic Profiling of Free and Immobilized Partially Purified Alkaline Protease from an Autochthonous Bacillus aryabhattai Ab15-ES" Reactions 4, no. 2: 231-245. https://doi.org/10.3390/reactions4020013
APA StyleAdetunji, A. I., & Olaniran, A. O. (2023). Biocatalytic Profiling of Free and Immobilized Partially Purified Alkaline Protease from an Autochthonous Bacillus aryabhattai Ab15-ES. Reactions, 4(2), 231-245. https://doi.org/10.3390/reactions4020013