
Proteins in Scalp Hair of Preschool Children

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Hair Protein Extraction
2.2. Proteomics Method
2.3. Generation of Age-Associated Proteomic Libraries
2.4. Human Scalp Hair Shaft Proteoforms Validation Studies
2.5. Statistical Analysis
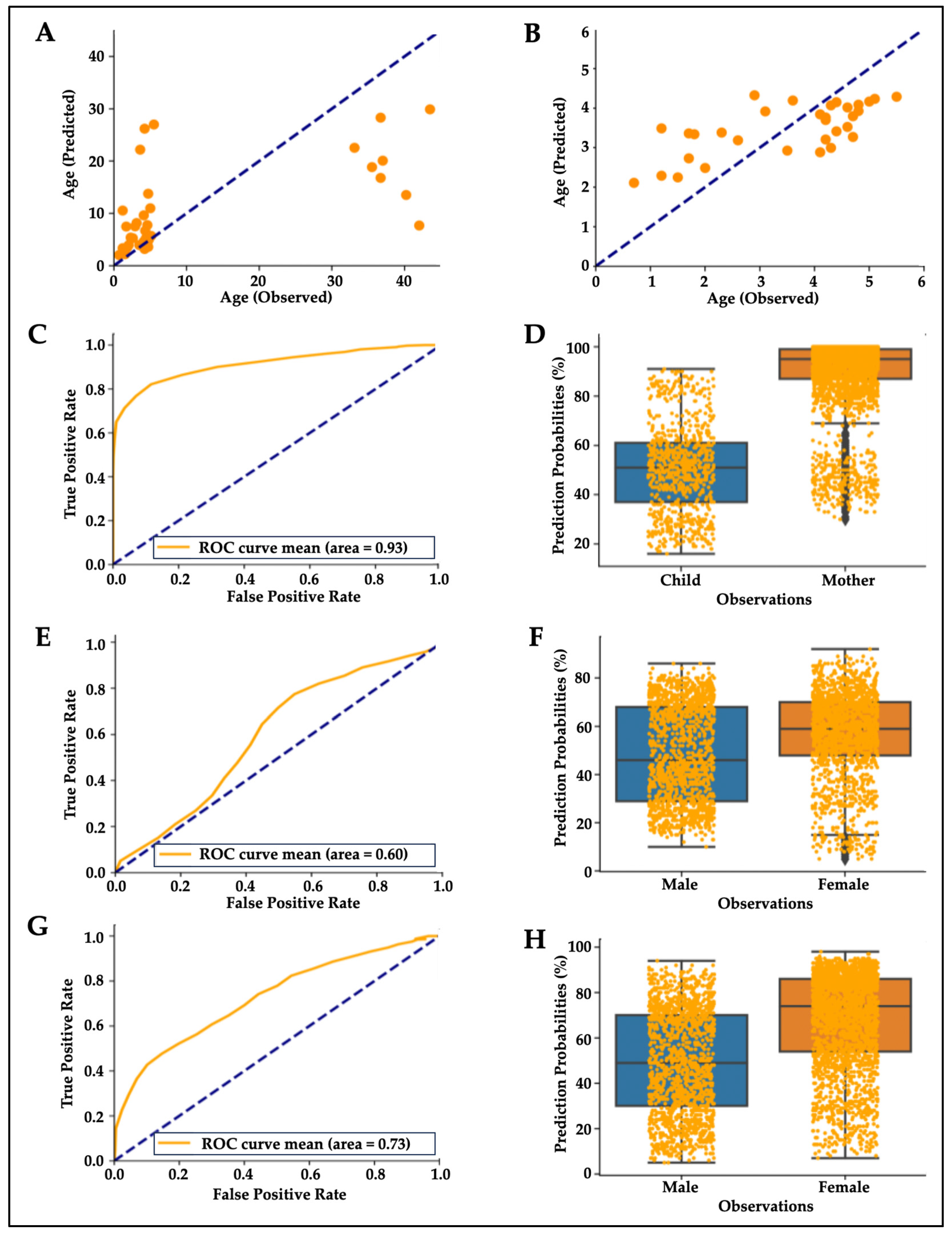
3. Results
3.1. Features of Hair Proteins
3.2. Hair Protein Profiles in Individuals and Families
3.3. Age- and Sex-Related Differences in Hair Proteins
3.4. Top Contributors to Hair Protein Variability
3.5. Biological Role(s) of the Strongest Contributors to Hair Protein Variability
3.6. ELISA Validation of Other Non-Structural Hair Proteins
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bitsko, R.H.; Holbrook, J.R.; Robinson, L.R.; Kaminski, J.W.; Ghandour, R.; Smith, C.; Peacock, G. Health Care, Family, and Community Factors Associated with Mental, Behavioral, and Developmental Disorders in Early Childhood—United States, 2011–2012. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Lopez, M.; Ruiz, M.O.; Rovnaghi, C.R.; Tam, G.K.; Hiscox, J.; Gotlib, I.H.; Barr, D.A.; Carrion, V.G.; Anand, K.J.S. The social ecology of childhood and early life adversity. Pediatr. Res. 2021, 89, 353–367. [Google Scholar] [CrossRef] [PubMed]
- Dubowitz, H.; Kressly, S.J. Documenting Psychosocial Problems in Children’s Electronic Health Records. JAMA Pediatr. 2023, 177, 881–882. [Google Scholar] [CrossRef] [PubMed]
- Rovnaghi, C.R.; Rigdon, J.; Roue, J.M.; Ruiz, M.O.; Carrion, V.G.; Anand, K.J.S. Longitudinal Trajectories of Hair Cortisol: Hypothalamic-Pituitary-Adrenal Axis Dysfunction in Early Childhood. Front. Pediatr. 2021, 9, 740343. [Google Scholar] [CrossRef]
- Anand, K.J.S.; Rigdon, J.; Rovnaghi, C.R.; Qin, F.; Tembulkar, S.; Bush, N.; LeWinn, K.; Tylavsky, F.A.; Davis, R.; Barr, D.A.; et al. Measuring socioeconomic adversity in early life. Acta Paediatr. 2019, 108, 1267–1277. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.A.; Scott, R.D.; Bhutta, Z.A.; Harris, N.B.; Danese, A.; Samara, M. Adversity in childhood is linked to mental and physical health throughout life. BMJ 2020, 371, m3048. [Google Scholar] [CrossRef]
- Palmer, F.B.; Anand, K.J.; Graff, J.C.; Murphy, L.E.; Qu, Y.; Volgyi, E.; Rovnaghi, C.R.; Moore, A.; Tran, Q.T.; Tylavsky, F.A. Early adversity, socioemotional development, and stress in urban 1-year-old children. J. Pediatr. 2013, 163, 1733–1739.e1731. [Google Scholar] [CrossRef]
- Erema, V.V.; Yakovchik, A.Y.; Kashtanova, D.A.; Bochkaeva, Z.V.; Ivanov, M.V.; Sosin, D.V.; Matkava, L.R.; Yudin, V.S.; Makarov, V.V.; Keskinov, A.A.; et al. Biological Age Predictors: The Status Quo and Future Trends. Int. J. Mol. Sci. 2022, 23, 15103. [Google Scholar] [CrossRef]
- Anand, K.J.S.; Rovnaghi, C.R.; Rigdon, J.; Qin, F.; Tembulkar, S.; Murphy, L.E.; Barr, D.A.; Gotlib, I.H.; Tylavsky, F.A. Demographic and psychosocial factors associated with hair cortisol concentrations in preschool children. Pediatr. Res. 2020, 87, 1119–1127. [Google Scholar] [CrossRef]
- Timmers, I.; Quaedflieg, C.W.E.M.; Hsu, C.; Healthcote, L.; Rovnaghi, C.R.; Simons, L. The interaction between stress and chronic pain through the lens of threat learning. Neurosci. Biobehav. Rev. 2019, 107, 641–655. [Google Scholar] [CrossRef]
- Reh, R.K.; Dias, B.G.; Nelson, C.A., 3rd; Kaufer, D.; Werker, J.F.; Kolb, B.; Levine, J.D.; Hensch, T.K. Critical period regulation across multiple timescales. Proc. Natl. Acad. Sci. USA 2020, 117, 23242–23251. [Google Scholar] [CrossRef]
- Brown, T.T.; Jernigan, T.L. Brain development during the preschool years. Neuropsychol. Rev. 2012, 22, 313–333. [Google Scholar] [CrossRef]
- Jernigan, T.L.; Brown, T.T.; Hagler, D.J., Jr.; Akshoomoff, N.; Bartsch, H.; Newman, E.; Thompson, W.K.; Bloss, C.S.; Murray, S.S.; Schork, N.; et al. The Pediatric Imaging, Neurocognition, and Genetics (PING) Data Repository. Neuroimage 2016, 124, 1149–1154. [Google Scholar] [CrossRef]
- Tozzi, L.; Garczarek, L.; Janowitz, D.; Stein, D.J.; Wittfeld, K.; Dobrowolny, H.; Lagopoulos, J.; Hatton, S.N.; Hickie, I.B.; Carballedo, A.; et al. Interactive impact of childhood maltreatment, depression, and age on cortical brain structure: Mega-analytic findings from a large multi-site cohort. Psychol. Med. 2020, 50, 1020–1031. [Google Scholar] [CrossRef]
- Warrier, V.; Stauffer, E.M.; Huang, Q.Q.; Wigdor, E.M.; Slob, E.A.W.; Seidlitz, J.; Ronan, L.; Valk, S.L.; Mallard, T.T.; Grotzinger, A.D.; et al. Genetic insights into human cortical organization and development through genome-wide analyses of 2,347 neuroimaging phenotypes. Nat. Genet. 2023, 55, 1483–1493. [Google Scholar] [CrossRef]
- Ruiz, M.O.; Rovnaghi, C.R.; Tembulkar, S.; Qin, F.; Truong, L.; Shen, S.; Anand, K.J.S. Linear hair growth rates in preschool children. Pediatr. Res. 2023, 95, 359–366. [Google Scholar] [CrossRef]
- Nicolaides, N.; Rothman, S. Studies on the chemical composition of human hair fat. II. The overall composition with regard to age, sex and race. J. Invest. Dermatol. 1953, 21, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Adeola, H.A.; Van Wyk, J.C.; Arowolo, A.; Ngwanya, R.M.; Mkentane, K.; Khumalo, N.P. Emerging Diagnostic and Therapeutic Potentials of Human Hair Proteomics. Proteomics Clin. Appl. 2018, 12, 48. [Google Scholar] [CrossRef] [PubMed]
- Tobin, D.J. The Human Hair Fibre. In Hair in Toxicology: An Important Bio-Monitor; Tobin, D.J., Ed.; Royal Society of Chemistry (U.K.): Cambridge, UK, 2005; pp. 34–56, Chapter 2. [Google Scholar]
- Villain, M.; Cirimele, V.; Kintz, P. Hair analysis in toxicology. Clin. Chem. Lab. Med. 2004, 42, 1265–1272. [Google Scholar] [CrossRef]
- Lee, Y.J.; Rice, R.H.; Lee, Y.M. Proteome analysis of human hair shaft: From protein identification to posttranslational modification. Mol. Cell Proteom. 2006, 5, 789–800. [Google Scholar] [CrossRef]
- Laatsch, C.N.; Durbin-Johnson, B.P.; Rocke, D.M.; Mukwana, S.; Newland, A.B.; Flagler, M.J.; Davis, M.G.; Eigenheer, R.A.; Phinney, B.S.; Rice, R.H. Human hair shaft proteomic profiling: Individual differences, site specificity and cuticle analysis. PeerJ 2014, 2, e506. [Google Scholar] [CrossRef]
- Carlson, T.L.; Moini, M.; Eckenrode, B.A.; Allred, B.M.; Donfack, J. Protein extraction from human anagen head hairs 1-millimeter or less in total length. Biotechniques 2018, 64, 170–176. [Google Scholar] [CrossRef]
- Wu, P.W.; Mason, K.E.; Durbin-Johnson, B.P.; Salemi, M.; Phinney, B.S.; Rocke, D.M.; Parker, G.J.; Rice, R.H. Proteomic analysis of hair shafts from monozygotic twins: Expression profiles and genetically variant peptides. Proteomics 2017, 17, 462. [Google Scholar] [CrossRef]
- Parker, G.J.; Leppert, T.; Anex, D.S.; Hilmer, J.K.; Matsunami, N.; Baird, L.; Stevens, J.; Parsawar, K.; Durbin-Johnson, B.P.; Rocke, D.M.; et al. Demonstration of Protein-Based Human Identification Using the Hair Shaft Proteome. PLoS ONE 2016, 11, e0160653. [Google Scholar] [CrossRef]
- Tawfik, D.S.; Rovnaghi, C.; Profit, J.; Cornell, T.T.; Anand, K.J.S. Prevalence of burnout and its relation to the neuroendocrine system among pediatric residents during the early COVID-19 pandemic: A pilot feasibility study. Compr. Psychoneuroendocrinol. 2023, 14, 100174. [Google Scholar] [CrossRef]
- Elias, J.E.; Gygi, S.P. Target-decoy search strategy for increased confidence in large-scale protein identifications by mass spectrometry. Nat. Methods 2007, 4, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef] [PubMed]
- Anand, K.J.S.; Leib, R.D.; Rovnaghi, C.R.; Singhal, K. PRIDE Repository, ProteomeXchange: 22 December 2019.
- Deutsch, E.W.; Csordas, A.; Sun, Z.; Jarnuczak, A.; Perez-Riverol, Y.; Ternent, T.; Campbell, D.S.; Bernal-Llinares, M.; Okuda, S.; Kawano, S.; et al. The ProteomeXchange consortium in 2017: Supporting the cultural change in proteomics public data deposition. Nucleic Acids Res. 2017, 45, D1100–D1106. [Google Scholar] [CrossRef]
- Slominski, R.; Rovnaghi, C.R.; Anand, K.J. Methodological Considerations for Hair Cortisol Measurements in Children. Ther. Drug Monit. 2015, 37, 812–820. [Google Scholar] [CrossRef]
- Wold, S.; Esbensen, K.; Geladi, P. Principal component analysis. WIREs Comput. Stat. 2010, 2, 433–459. [Google Scholar] [CrossRef]
- Hotelling, H. Analysis of a complex of statistical variables into principal components. J. Educ. Psychol. 1933, 24, 417–441. [Google Scholar] [CrossRef]
- Jolliffe, I.T. Principal Component Analysis; Springer: New York, NY, USA, 1986. [Google Scholar]
- Linderman, G.C.; Steinerberger, S. Clustering with t-SNE, provably. SIAM J. Math. Data Sci. 2019, 1, 313–332. [Google Scholar] [CrossRef]
- Van der Maaten, L.; Hinton, G. Visualizing data using t-SNE. J. Mach. Learn. Res. 2008, 9, 2579–2605. [Google Scholar]
- Jylhävä, J.; Pedersen, N.L.; Hägg, S. Biological Age Predictors. EbioMedicine 2017, 21, 29–36. [Google Scholar] [CrossRef]
- Barthelemy, N.R.; Bednarczyk, A.; Schaeffer-Reiss, C.; Jullien, D.; Van Dorsselaer, A.; Cavusoglu, N. Proteomic tools for the investigation of human hair structural proteins and evidence of weakness sites on hair keratin coil segments. Anal. Biochem. 2012, 421, 43–55. [Google Scholar] [CrossRef]
- Gomes, J.; Au, F.; Basak, A.; Cakmak, S.; Vincent, R.; Kumarathasan, P. Maternal blood biomarkers and adverse pregnancy outcomes: A systematic review and meta-analysis. Crit. Rev. Toxicol. 2019, 49, 461–478. [Google Scholar] [CrossRef]
- Handelman, S.K.; Romero, R.; Tarca, A.L.; Pacora, P.; Ingram, B.; Maymon, E.; Chaiworapongsa, T.; Hassan, S.S.; Erez, O. The plasma metabolome of women in early pregnancy differs from that of non-pregnant women. PLoS ONE 2019, 14, e0224682. [Google Scholar] [CrossRef]
- Romero, R.; Erez, O.; Maymon, E.; Chaemsaithong, P.; Xu, Z.; Pacora, P.; Chaiworapongsa, T.; Done, B.; Hassan, S.S.; Tarca, A.L. The maternal plasma proteome changes as a function of gestational age in normal pregnancy: A longitudinal study. Am. J. Obstet. Gynecol. 2017, 217, 67.e61–67.e67. [Google Scholar] [CrossRef]
- Lara, J.; Cooper, R.; Nissan, J.; Ginty, A.T.; Khaw, K.T.; Deary, I.J.; Lord, J.M.; Kuh, D.; Mathers, J.C. A proposed panel of biomarkers of healthy ageing. BMC Med. 2015, 13, 222. [Google Scholar] [CrossRef] [PubMed]
- Deelen, J.; Kettunen, J.; Fischer, K.; van der Spek, A.; Trompet, S.; Kastenmuller, G.; Boyd, A.; Zierer, J.; van den Akker, E.B.; Ala-Korpela, M.; et al. A metabolic profile of all-cause mortality risk identified in an observational study of 44,168 individuals. Nat. Commun. 2019, 10, 3346. [Google Scholar] [CrossRef] [PubMed]
- Breiman, L. Random Forest. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Ebstein, R.P.; Knafo, A.; Mankuta, D.; Chew, S.H.; Lai, P.S. The contributions of oxytocin and vasopressin pathway genes to human behavior. Horm. Behav. 2012, 61, 359–379. [Google Scholar] [CrossRef]
- Muscogiuri, G.; Barrea, L.; Annunziata, G.; Vecchiarini, M.; Orio, F.; Di Somma, C.; Colao, A.; Savastano, S. Water intake keeps type 2 diabetes away? Focus on copeptin. Endocrine 2018, 62, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekharan, B.; Montllor-Albalate, C.; Colin, A.E.; Andersen, J.L.; Jang, Y.C.; Reddi, A.R. Cu/Zn Superoxide Dismutase (Sod1) regulates the canonical Wnt signaling pathway. Biochem. Biophys. Res. Commun. 2021, 534, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Une, M.; Yamakawa, M.; Watanabe, Y.; Uchino, K.; Honda, N.; Adachi, M.; Nakanishi, M.; Umezawa, A.; Kawata, Y.; Nakashima, K.; et al. SOD1-interacting proteins: Roles of aggregation cores and protein degradation systems. Neurosci. Res. 2021, 170, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Nam, M.K.; Seong, Y.; Jeong, G.H.; Yoo, S.A.; Rhim, H. HtrA2 regulates alpha-Synuclein-mediated mitochondrial reactive oxygen species production in the mitochondria of microglia. Biochem. Biophys. Res. Commun. 2023, 638, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Ganne, A.; Balasubramaniam, M.; Griffin, W.S.T.; Shmookler Reis, R.J.; Ayyadevara, S. Glial Fibrillary Acidic Protein: A Biomarker and Drug Target for Alzheimer’s Disease. Pharmaceutics 2022, 14, 71354. [Google Scholar] [CrossRef] [PubMed]
- Griffin, S.T.G.; Stanley, L.C.; Yeralan, O.; Rovnaghi, C.R.; Marshak, D.R. Cytokines in Human Neurodegenerative Disease; Conn, P.M., Ed.; Academic Press, Inc.: San Diego, CA, USA, 1993; Volume 17, p. 295. [Google Scholar]
- Cruz, C.F.; Fernandes, M.M.; Gomes, A.C.; Coderch, L.; Marti, M.; Mendez, S.; Gales, L.; Azoia, N.G.; Shimanovich, U.; Cavaco-Paulo, A. Keratins and lipids in ethnic hair. Int. J. Cosmet. Sci. 2013, 35, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Franbourg, A.; Hallegot, P.; Baltenneck, F.; Toutain, C.; Leroy, F. Current research on ethnic hair. J. Am. Acad. Dermatol. 2003, 48, S115–S119. [Google Scholar] [CrossRef]
- Horvath, A.L. Solubility of structurally complicated materials: 3. Hair. Sci. World J. 2009, 9, 255–271. [Google Scholar] [CrossRef]
- Marti, M.; Barba, C.; Manich, A.M.; Rubio, L.; Alonso, C.; Coderch, L. The influence of hair lipids in ethnic hair properties. Int. J. Cosmet. Sci. 2016, 38, 77–84. [Google Scholar] [CrossRef]
- Wilson, A.S.; Tobin, D.J. Aging Hair; Springer: Berlin, Germany, 2010; pp. 249–261. [Google Scholar]
- Biasiolo, A.; Tono, N.; Ruvoletto, M.; Quarta, S.; Turato, C.; Villano, G.; Beneduce, L.; Fassina, G.; Merkel, C.; Gatta, A.; et al. IgM-linked SerpinB3 and SerpinB4 in sera of patients with chronic liver disease. PLoS ONE 2012, 7, e40658. [Google Scholar] [CrossRef]
- Markovina, S.; Wang, S.; Henke, L.E.; Luke, C.J.; Pak, S.C.; DeWees, T.; Pfeifer, J.D.; Schwarz, J.K.; Liu, W.; Chen, S.; et al. Serum squamous cell carcinoma antigen as an early indicator of response during therapy of cervical cancer. Br. J. Cancer 2018, 118, 72–78. [Google Scholar] [CrossRef]
- van Kempen, P.M.; Noorlag, R.; Swartz, J.E.; Bovenschen, N.; Braunius, W.W.; Vermeulen, J.F.; Van Cann, E.M.; Grolman, W.; Willems, S.M. Oropharyngeal squamous cell carcinomas differentially express granzyme inhibitors. Cancer Immunol. Immunother. 2016, 65, 575–585. [Google Scholar] [CrossRef]
- Charlesworth, A.; Chiaverini, C.; Chevrant-Breton, J.; DelRio, M.; Diociaiuti, A.; Dupuis, R.P.; El Hachem, M.; Le Fiblec, B.; Sankari-Ho, A.M.; Valhquist, A.; et al. Epidermolysis bullosa simplex with PLEC mutations: New phenotypes and new mutations. Br. J. Dermatol. 2013, 168, 808–814. [Google Scholar] [CrossRef]
- Paumard-Hernandez, B.; Calvete, O.; Inglada Perez, L.; Tejero, H.; Al-Shahrour, F.; Pita, G.; Barroso, A.; Carlos Trivino, J.; Urioste, M.; Valverde, C.; et al. Whole exome sequencing identifies PLEC, EXO5 and DNAH7 as novel susceptibility genes in testicular cancer. Int. J. Cancer 2018, 143, 1954–1962. [Google Scholar] [CrossRef]
- Winter, L.; Turk, M.; Harter, P.N.; Mittelbronn, M.; Kornblum, C.; Norwood, F.; Jungbluth, H.; Thiel, C.T.; Schlotzer-Schrehardt, U.; Schroder, R. Downstream effects of plectin mutations in epidermolysis bullosa simplex with muscular dystrophy. Acta Neuropathol. Commun. 2016, 4, 44. [Google Scholar] [CrossRef]
- Barel, O.; Shorer, Z.; Flusser, H.; Ofir, R.; Narkis, G.; Finer, G.; Shalev, H.; Nasasra, A.; Saada, A.; Birk, O.S. Mitochondrial complex III deficiency associated with a homozygous mutation in UQCRQ. Am. J. Hum. Genet. 2008, 82, 1211–1216. [Google Scholar] [CrossRef]
- Eckhaus, J.; Lawrence, K.M.; Helbig, I.; Bui, M.; Vadlamudi, L.; Hopper, J.L.; Scheffer, I.E.; Berkovic, S.F. Genetics of febrile seizure subtypes and syndromes: A twin study. Epilepsy Res. 2013, 105, 103–109. [Google Scholar] [CrossRef]
- Lagae, L. What’s new in: “genetics in childhood epilepsy”. Eur. J. Pediatr. 2008, 167, 715–722. [Google Scholar] [CrossRef]
- Munakata, K.; Uemura, M.; Takemasa, I.; Ozaki, M.; Konno, M.; Nishimura, J.; Hata, T.; Mizushima, T.; Haraguchi, N.; Noura, S.; et al. SCGB2A1 is a novel prognostic marker for colorectal cancer associated with chemoresistance and radioresistance. Int. J. Oncol. 2014, 44, 1521–1528. [Google Scholar] [CrossRef]
- Mauvais-Jarvis, F.; Bairey Merz, N.; Barnes, P.J.; Brinton, R.D.; Carrero, J.J.; DeMeo, D.L.; De Vries, G.J.; Epperson, C.N.; Govindan, R.; Klein, S.L.; et al. Sex and gender: Modifiers of health, disease, and medicine. Lancet 2020, 396, 565–582. [Google Scholar] [CrossRef]
- Wright, F.A.; Sullivan, P.F.; Brooks, A.I.; Zou, F.; Sun, W.; Xia, K.; Madar, V.; Jansen, R.; Chung, W.; Zhou, Y.H.; et al. Heritability and genomics of gene expression in peripheral blood. Nat. Genet. 2014, 46, 430–437. [Google Scholar] [CrossRef]
- Stoeckli, E.T. Understanding axon guidance: Are we nearly there yet? Development 2018, 145, 151415. [Google Scholar] [CrossRef]
- Sutor, B.; Hagerty, T. Involvement of gap junctions in the development of the neocortex. Biochim. Biophys. Acta 2005, 1719, 59–68. [Google Scholar] [CrossRef]
- Lee, S.E.; West, K.P., Jr.; Cole, R.N.; Schulze, K.J.; Wu, L.S.; Yager, J.D.; Groopman, J.; Christian, P. General intelligence is associated with subclinical inflammation in Nepalese children: A population-based plasma proteomics study. Brain Behav. Immun. 2016, 56, 253–263. [Google Scholar] [CrossRef]
- Lakshmi Priya, M.D.; Geetha, A. A biochemical study on the level of proteins and their percentage of nitration in the hair and nail of autistic children. Clin. Chim. Acta 2011, 412, 1036–1042. [Google Scholar] [CrossRef]
- Breen, M.S.; Ozcan, S.; Ramsey, J.M.; Wang, Z.; Ma’ayan, A.; Rustogi, N.; Gottschalk, M.G.; Webster, M.J.; Weickert, C.S.; Buxbaum, J.D.; et al. Temporal proteomic profiling of postnatal human cortical development. Transl. Psychiatry 2018, 8, 267. [Google Scholar] [CrossRef]
- Carlyle, B.C.; Kitchen, R.R.; Kanyo, J.E.; Voss, E.Z.; Pletikos, M.; Sousa, A.M.M.; Lam, T.T.; Gerstein, M.B.; Sestan, N.; Nairn, A.C. A multiregional proteomic survey of the postnatal human brain. Nat. Neurosci. 2017, 20, 1787–1795. [Google Scholar] [CrossRef]
- Johnson, A.A.; Shokhirev, M.N.; Wyss-Coray, T.; Lehallier, B. Systematic review and analysis of human proteomics aging studies unveils a novel proteomic aging clock and identifies key processes that change with age. Ageing Res. Rev. 2020, 60, 101070. [Google Scholar] [CrossRef]
- Pedlar, C.R.; Newell, J.; Lewis, N.A. Blood Biomarker Profiling and Monitoring for High-Performance Physiology and Nutrition: Current Perspectives, Limitations and Recommendations. Sports Med. 2019, 49, 185–198. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family Code | Subject | Age (Months) | Age (Years) | Gender | Race | Ethnicity | # of Hair Proteins | Peptide Spectral Matches |
|---|---|---|---|---|---|---|---|---|
| F107 | Mother | 450.6 | 37.6 | F | White | NH | 819 | 6.533 |
| Child1 | 27.6 | 2.3 | F | White | NH | 568 | 3.949 | |
| Child2 | 58.0 | 4.8 | M | White | Other | 464 | 2.873 | |
| F123 | Mother | 447.1 | 37.3 | F | White | NH | 809 | 10.370 |
| Child1 | 24.0 | 2 | F | White | NH | 499 | 3.728 | |
| Child2 | 52.4 | 4.4 | M | White | NH | 573 | 5.078 | |
| F134 | Mother | 431.8 | 35.9 | F | White | NA | 684 | 5.445 |
| Child1 | 20.9 | 1.74 | M | Mixed | NA | 387 | 2.760 | |
| Child2 | 67.6 | 5.6 | F | Mixed | NA | 759 | 6.872 | |
| F142 | Mother | 447.3 | 37.3 | F | Asian | NA | 650 | 8.370 |
| Child1 | 20.1 | 1.7 | M | Mixed | NA | 581 | 6.208 | |
| Child2 | 50.6 | 4.2 | M | Mixed | NA | 226 | 2.353 | |
| F183 | Mother | 530.0 | 44.2 | F | Asian | NH | 1090 | 10.527 |
| Child1 | 8.5 | 0.7 | M | Asian | NH | 314 | 2.331 | |
| Child2 | 44.0 | 3.7 | M | Asian | NH | 1010 | 8.065 | |
| F218 | Mother | 504 | 42 | F | White | H | 609 | 4.144 |
| Child1 | 58.5 | 4.9 | F | White | H | 524 | 3.107 | |
| Child2 | 35.2 | 2.9 | F | White | H | 631 | 4.475 | |
| F271 | Mother | 402.6 | 33.6 | F | White | NH | 769 | 7.525 |
| Child1 | 15.1 | 1.3 | F | White | NH | 557 | 7.161 | |
| Child2 | 42.5 | 3.5 | F | White | NH | 600 | 5.615 | |
| F286 | Mother | 489.8 | 40.8 | F | White | NH | 616 | 9.209 |
| Child1 | 22.0 | 1.8 | M | White | NH | 403 | 4.727 | |
| Child2 | 52.5 | 4.4 | F | White | NH | 614 | 6.061 | |
| F346 | Child | 50.3 | 4.2 | M | White | NA | 475 | 3.429 |
| F192 | Child | 38.1 | 3.2 | F | Other | H | 283 | 1.731 |
| F132 | Child | 51.4 | 4.3 | F | White | NA | 272 | 1.892 |
| F363 | Child | 56.6 | 4.7 | M | Mixed | NH | 270 | 1.914 |
| F281 | Child | 53.5 | 4.5 | M | Mixed | Mixed | 406 | 3.192 |
| F173 | Child | 51.3 | 4.3 | F | Other | Other | 835 | 7.168 |
| F380 | Child | 14.8 | 1.2 | M | Asian | NA | 237 | 1.814 |
| F159 | Child | 62.7 | 5.2 | F | White | NH | 485 | 3.830 |
| F179 | Child | 53.0 | 4.4 | F | Asian | Other | 494 | 2.926 |
| F149 | Child | 61.3 | 5.1 | F | Mixed | NH | 698 | 5.733 |
| F106 | Child | 56.7 | 4.7 | F | Asian | NH | 668 | 6.390 |
| F153 | Child | 57.1 | 4.8 | M | Asian | NA | 275 | 2.549 |
| F256 | Child | 55.8 | 4.7 | M | Mixed | Mixed | 638 | 7.016 |
| F190 | Child | 31.5 | 2.6 | F | Asian | Other | 527 | 7.460 |
| F104 | Child | 50.2 | 4.2 | M | White | NH | 672 | 8.084 |
| F113 | Child | 17.9 | 1.5 | F | White | NH | 441 | 3.640 |
| Entrez Gene Name | Gene Symbol: Human | Expr Log Ratio | p-Value | Location | Type(s) |
|---|---|---|---|---|---|
| Involucrin | IVL | –2.85 | 0.0576 | Cytoplasm | other |
| Serpin family B4 | SERPINB4 | –2.452 | 0.0009 *** | Cytoplasm | other |
| Actin binding protein | POF1B | –2.097 | 0.0151 * | Membrane | other |
| Plectin | PLEC | –1.886 | 0.0004 *** | Cytoplasm | other |
| Alpha-2-macroglobulin like 1 | A2ML1 | –1.858 | 0.0042 ** | Cytoplasm | other |
| H3 clustered histone 1 | HIST1H3A | –1.743 | 0.0038 ** | Nucleus | other |
| Ubiquinol-cytochrome c reductase complex III | UQCRQ | –1.716 | 0.0007 *** | Cytoplasm | enzyme |
| Adenosylhomocysteinase | AHCY | –1.472 | 0.0040 ** | Cytoplasm | enzyme |
| Heat shock protein family A (Hsp70-1A) | HSPA1A | –1.35 | 0.0569 | Cytoplasm | enzyme |
| H2B clustered histone 9 | HIST1H2BH | –1.17 | 0.5070 | Nucleus | other |
| Histidine ammonia-lyase | HAL | –1.087 | 0.0851 | Cytoplasm | enzyme |
| COPI coat complex subunit zeta 1 | COPZ1 | –0.931 | 0.158 | Cytoplasm | transporter |
| Eukaryotic translation initiation factor 3A | EIF3A | –0.8 | 0.0567 | Cytoplasm | other |
| Tubulin alpha 1c | TUBA1C | –0.526 | 0.262 | Cytoplasm | other |
| Casein beta | CSN2 | –0.269 | 0.491 | Extracellular | kinase |
| ATP citrate lyase | ACLY | –0.249 | 0.0954 | Cytoplasm | enzyme |
| Protein disulfide isomerase A3 | PDIA3 | –0.051 | 0.884 | Cytoplasm | peptidase |
| Scinderin | SCIN | 0.028 | 0.221 | Cytoplasm | other |
| Alström syndrome protein 1 | ALMS1 | 0.18 | 0.572 | Cytoplasm | other |
| Histone H3.4 | HIST3H3 | 0.64 | 0.153 | Nucleus | other |
| Myeloperoxidase | MPO | 0.925 | 0.886 | Cytoplasm | enzyme |
| Secretoglobin 2A1 | SCGB2A1 | 5.32 | 0.0008 *** | Extracellular | other |
| Entrez Gene Name | Gene Symbol: Human | Expr Log Ratio | p-Value | Location | Type |
|---|---|---|---|---|---|
| Casein beta | CSN2 | −3.046 | 0.0184 * | Extracellular | kinase |
| Serpin family B4 | SERPINB4 | −1.303 | 0.391 | Cytoplasm | other |
| Secretoglobin family 2A1 | SCGB2A1 | −1.036 | 0.0513 | Extracellular | other |
| Protein disulfide isomerase A3 | PDIA3 | −0.78 | 0.662 | Cytoplasm | peptidase |
| ATP citrate lyase | ACLY | −0.531 | 0.585 | Cytoplasm | enzyme |
| Myeloperoxidase | MPO | −0.493 | 0.581 | Cytoplasm | enzyme |
| Involucrin | IVL | −0.476 | 0.804 | Cytoplasm | other |
| Eukaryotic translation initiation factor 3A | EIF3A | −0.295 | 0.226 | Cytoplasm | other |
| Alpha-2-macroglobulin like 1 | A2ML1 | −0.254 | 0.923 | Cytoplasm | other |
| Scinderin | SCIN | −0.187 | 0.375 | Cytoplasm | other |
| Heat shock protein family A (Hsp70-1A) | HSPA1A | −0.122 | 0.573 | Cytoplasm | enzyme |
| Actin binding protein | POF1B | 0.094 | 0.875 | Membrane | other |
| Histone H3.4 | H3-4 | 0.139 | 0.938 | Nucleus | other |
| Histidine ammonia-lyase | HAL | 0.175 | 0.522 | Cytoplasm | enzyme |
| COPI coat complex zeta 1 | COPZ1 | 0.225 | 0.536 | Cytoplasm | transporter |
| Tubulin alpha 1c | TUBA1C | 0.245 | 0.314 | Cytoplasm | other |
| H3 clustered histone 1 | H3C1 | 0.249 | 0.562 | Nucleus | other |
| Adenosylhomocysteinase | AHCY | 0.333 | 0.202 | Cytoplasm | enzyme |
| Plectin | PLEC | 0.441 | 0.256 | Cytoplasm | other |
| Ubiquinol-cytochrome c reductase complex III | UQCRQ | 1.415 | 0.0976 | Cytoplasm | enzyme |
| H2B clustered histone 9 | H2BC9 | 1.423 | 0.221 | Nucleus | other |
| Alström syndrome protein 1 | ALMS1 | 1.754 | 0.0214 * | Cytoplasm | other |
| Hair Sample Pools Based on Hair Cortisol Concentration | Cortisol ng/mL | AVP pg/mL | Cu/Zn SOD ng/mL | HTRA2 ng/mL | GFAP ng/mL |
|---|---|---|---|---|---|
| Low Child pool cortisol (n = 72) | 40.84 | 14.81 | 0.25 | 7.54 | 0.00 |
| Moderate Child pool cortisol (n = 21) | 60.34 | 11.91 | 0.18 | 4.61 | 0.41 |
| High Child pool cortisol (n = 7) | 190.89 | 7.18 | 0.23 | 9.14 | n/a |
| Low Father pool cortisol (n = 13) | 22.39 | 8.36 | 0.63 | 9.65 | 2.64 |
| Low Mother pool cortisol (n = 39) | 17.24 | 7.88 | 0.49 | 7.71 | 1.45 |
| High Mother pool cortisol (n = 7) | 36.77 | 11.68 | n/a | n/a | n/a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rovnaghi, C.R.; Singhal, K.; Leib, R.D.; Xenochristou, M.; Aghaeepour, N.; Chien, A.S.; Dinakarpandian, D.; Anand, K.J.S. Proteins in Scalp Hair of Preschool Children. Psych 2024, 6, 143-162. https://doi.org/10.3390/psych6010009
Rovnaghi CR, Singhal K, Leib RD, Xenochristou M, Aghaeepour N, Chien AS, Dinakarpandian D, Anand KJS. Proteins in Scalp Hair of Preschool Children. Psych. 2024; 6(1):143-162. https://doi.org/10.3390/psych6010009
Chicago/Turabian StyleRovnaghi, Cynthia R., Kratika Singhal, Ryan D. Leib, Maria Xenochristou, Nima Aghaeepour, Allis S. Chien, Deendayal Dinakarpandian, and Kanwaljeet J. S. Anand. 2024. "Proteins in Scalp Hair of Preschool Children" Psych 6, no. 1: 143-162. https://doi.org/10.3390/psych6010009
APA StyleRovnaghi, C. R., Singhal, K., Leib, R. D., Xenochristou, M., Aghaeepour, N., Chien, A. S., Dinakarpandian, D., & Anand, K. J. S. (2024). Proteins in Scalp Hair of Preschool Children. Psych, 6(1), 143-162. https://doi.org/10.3390/psych6010009