Effect of Acute Seawater Temperature Increase on the Survival of a Fish Ectoparasite

 , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Gnathiid Collection
2.3. Experimental Protocol
2.3.1. Great Barrier Reef
2.3.2. Philippines
2.4. Statistical Analyses
2.4.1. Great Barrier Reef
2.4.2. Philippines
2.5. Ethics
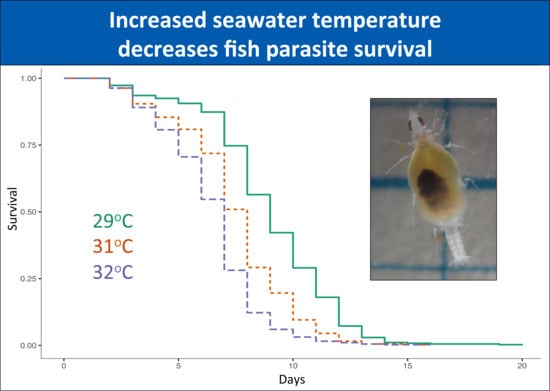
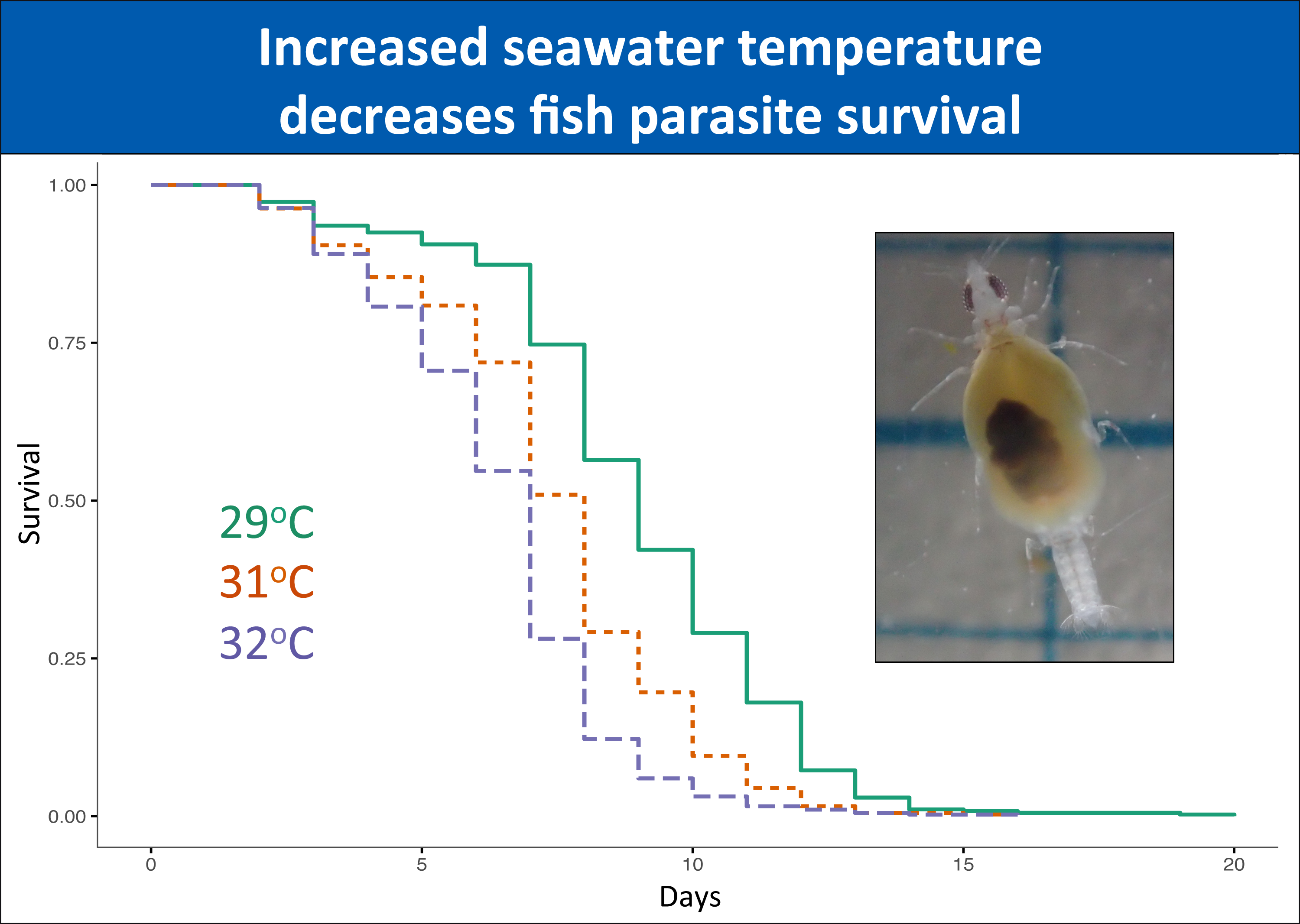
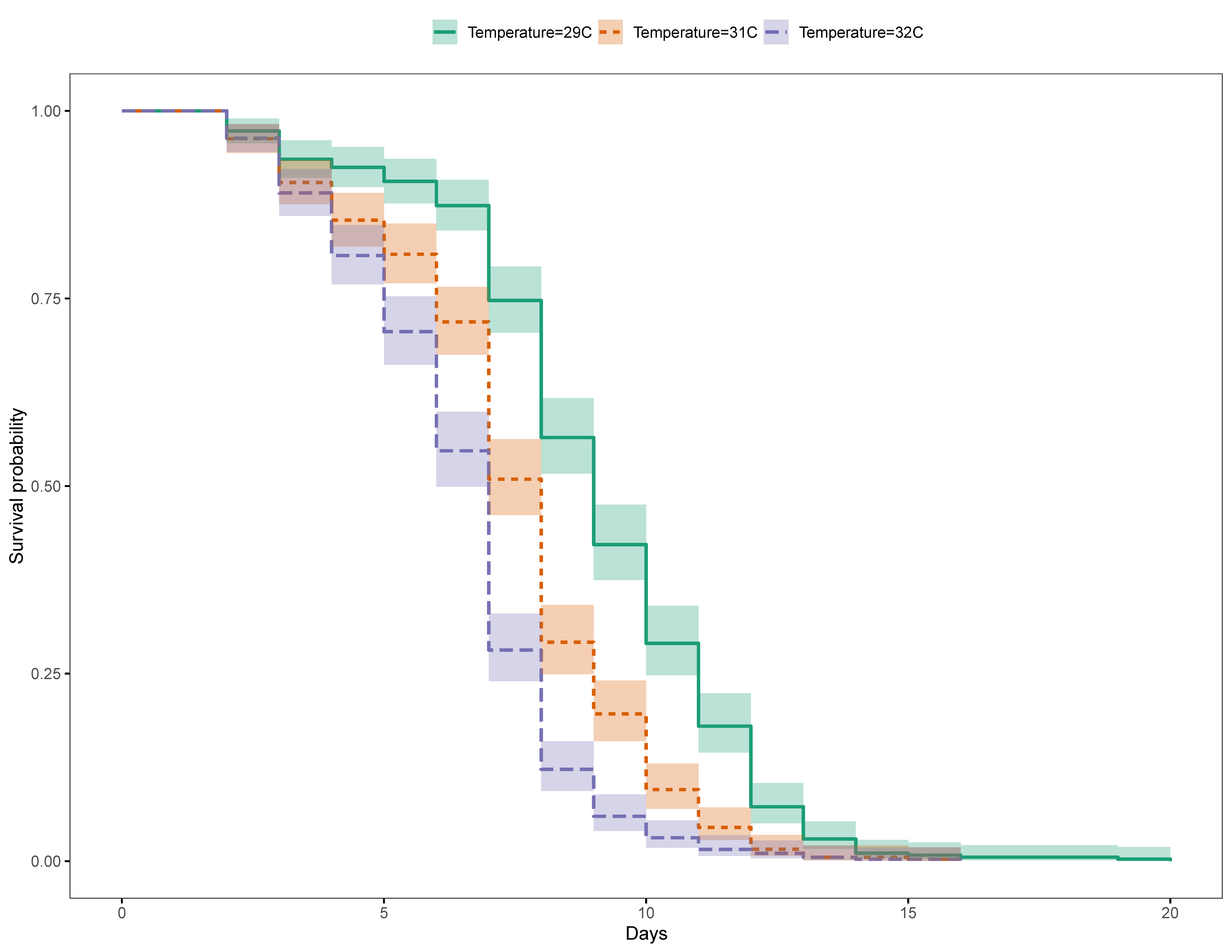
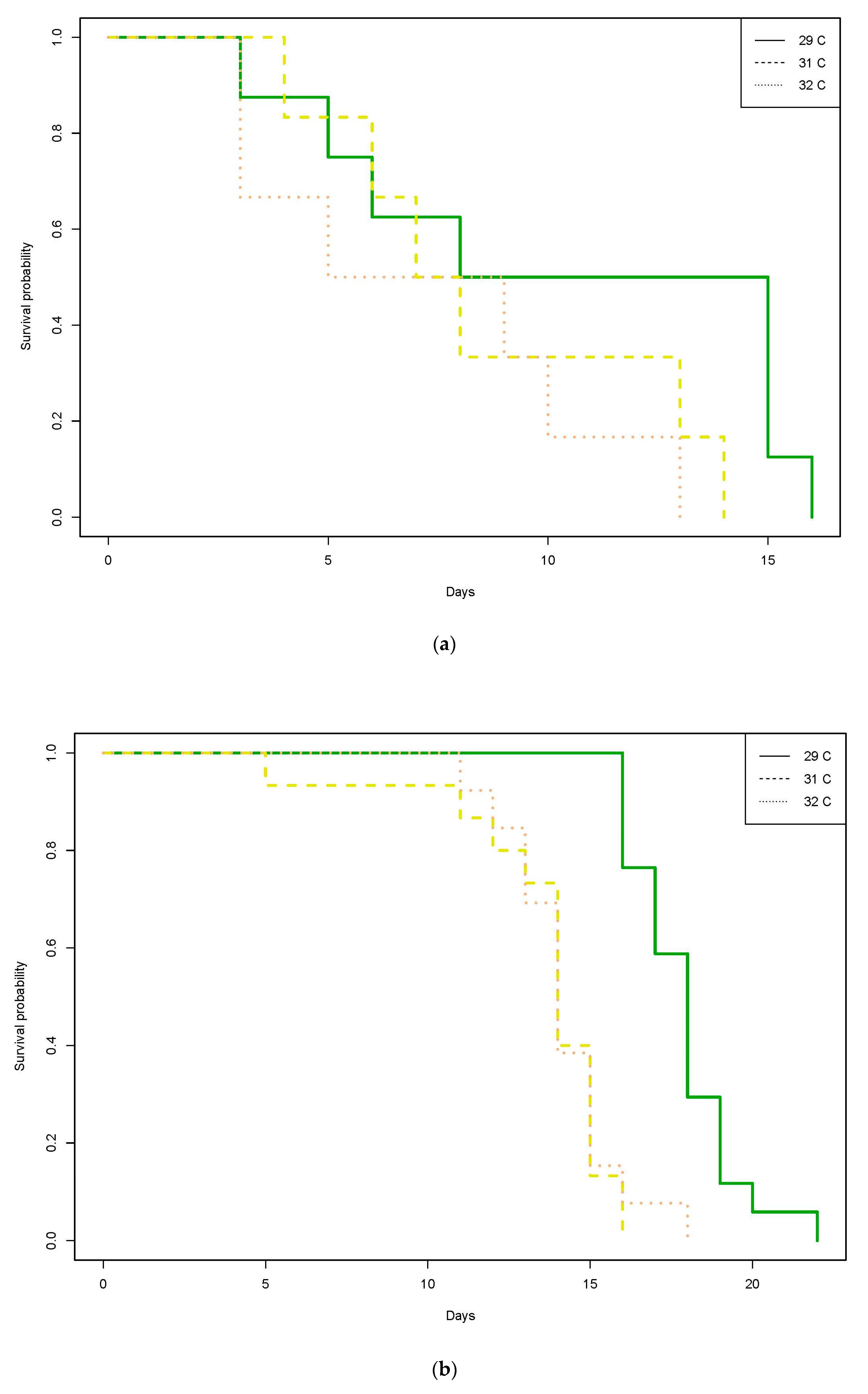
3. Results
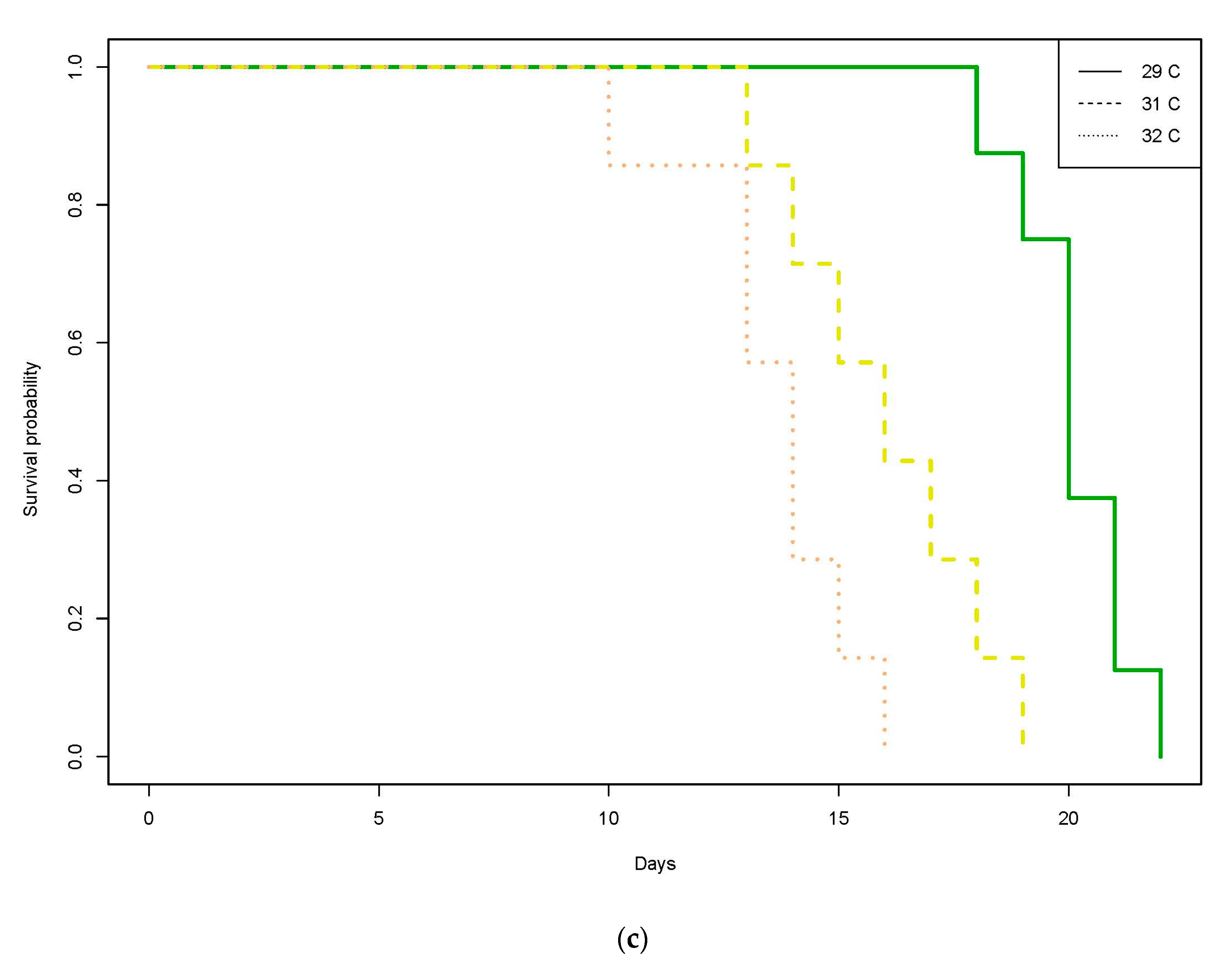
3.1. Great Barrier Reef
3.1.1. Unfed Gnathiids
3.1.2. Fed Gnathiids
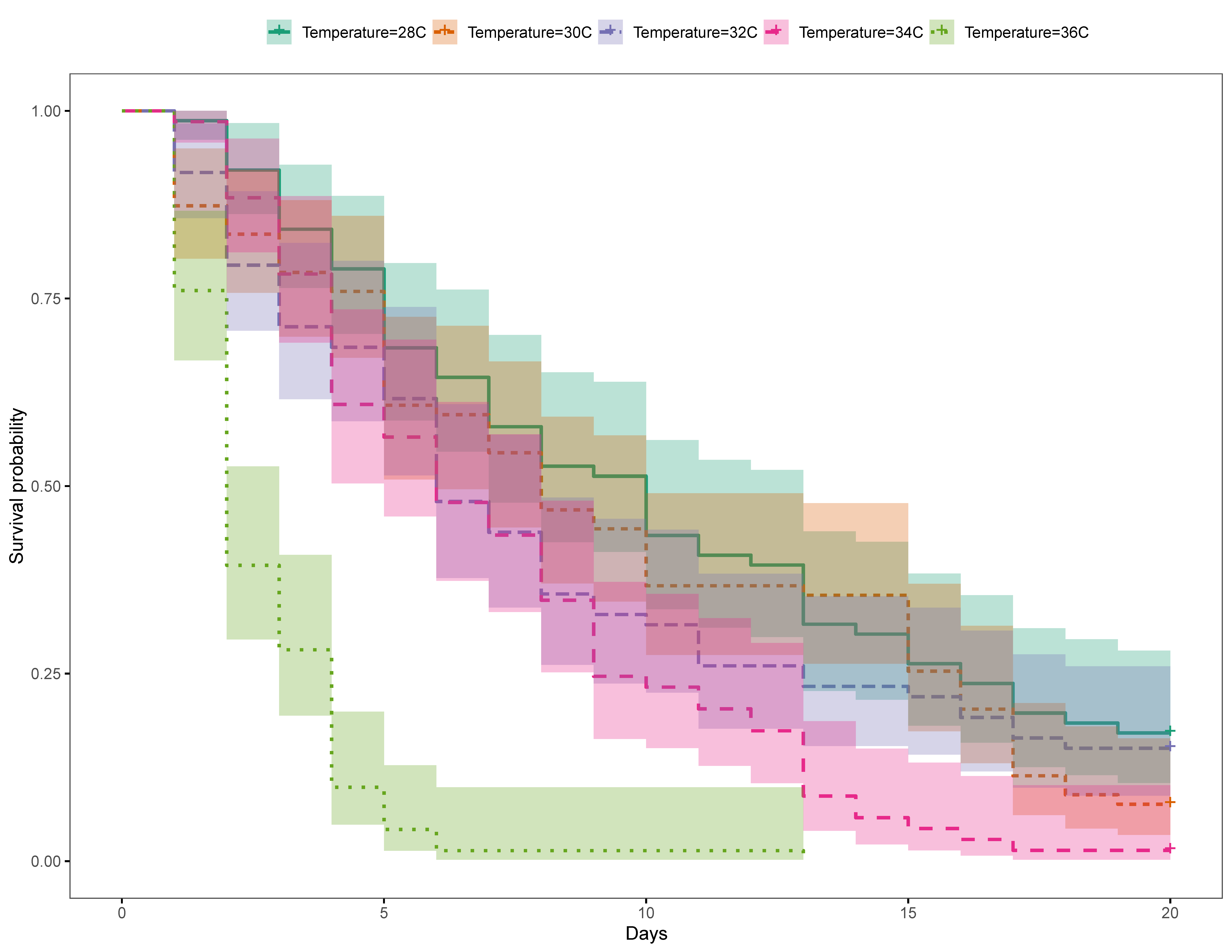
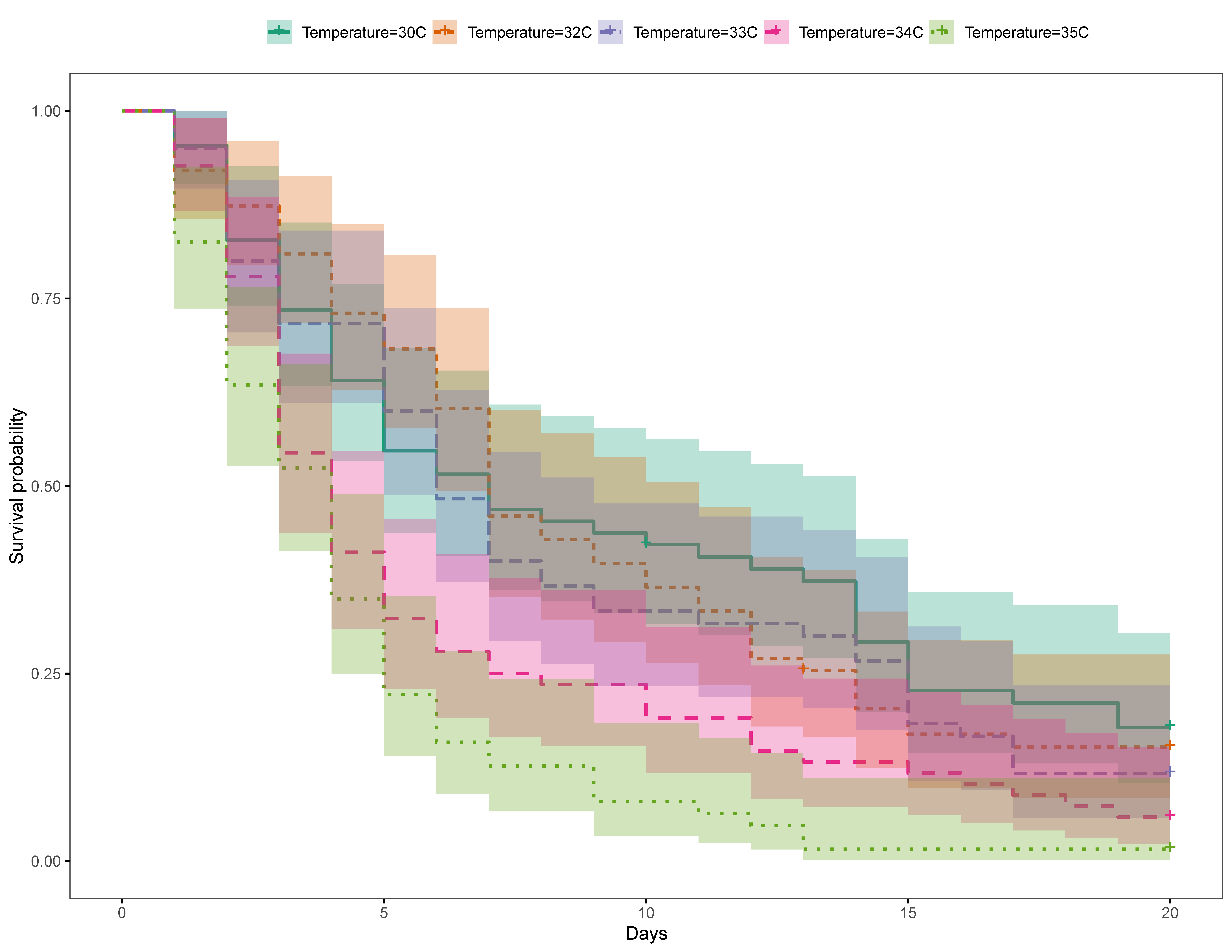
3.2. Philippines
3.2.1. Trial 1
3.2.2. Trial 2
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bindoff, N.L.; Cheung, W.W.L.; Kairo, J.G.; Arístegui, J.; Guinder, V.A.; Hilmi, N.; Jiao, N.; Karim, M.S.; Levin, L.; O’Donoghue, S.; et al. Changing Ocean, Marine Ecosystems, and Dependent Communities; ETH Zurich: Zurich, Switzerland, 2019; pp. 477–587. [Google Scholar]
- Huey, R.B.; Stevenson, R.D. Integrating Thermal Physiology and Ecology of Ectotherms: A Discussion of Approaches. Am. Zool. 1979, 19, 357–366. [Google Scholar] [CrossRef] [Green Version]
- Brett, J.R. 3. Temperature, 3.3. Animals, 3.32. Fishes-Functional Reponses. In Marine Ecology: A Comprehensive, Integrated Treatise on Life in Oceans and Coastal Waters; Kinne, O., Ed.; Wiley-Interscience: Hoboken, NJ, USA, 1984; Volume 1, pp. 515–560. [Google Scholar]
- Kinne, O. 3. Temperature, 3.3. Animals, 3.31. Invertebrates. In Marine Ecology: A Comprehensive, Integrated Treatise on Life in Oceans and Coastal Waters; Kinne, O., Ed.; Wiley-Interscience: Hoboken, NJ, USA, 1984; Volume 1, pp. 407–514. [Google Scholar]
- Newell, R.C.; Branch, G.M. The influence of temperature on the maintenance of metabolic energy balance in marine invertebrates. Adv. Mar. Biol. 1980, 17, 329–396. [Google Scholar] [CrossRef]
- Houde, E.D. Comparative growth, mortality, and energetics of marine fish larvae: Temperature and implied latitudinal effects. Fish. Bull. 1989, 87, 471–495. [Google Scholar]
- Schmidt-Nielsen, K. Animal Physiology: Adaptation and Environment; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Marsden, I.D. Effect of temperature on the microdistribution of the isopod Sphaeroma rugicauda from a saltmarsh habitat. Mar. Biol. 1976, 38, 117–128. [Google Scholar] [CrossRef]
- McNamara, J.C.; Moreira, P.S.; Moreira, G.S. The effect of salinity on the upper thermal limits of survival and metamorphosis during larval development in Macrobrachium amazonicum (Heller) (Decapoda, Palaemonidae). Crustaceana 1986, 50, 231–238. [Google Scholar] [CrossRef]
- Marcogliese, D.J. The impact of climate change on the parasites and infectious diseases of aquatic animals. Oie Rev. Sci. Tech. 2008, 27, 467–484. [Google Scholar] [CrossRef]
- Marcogliese, D.J. The Distribution and Abundance of Parasites in Aquatic Ecosystems in a Changing Climate: More than Just Temperature. Integr. Comp. Biol. 2016, 56, 611–619. [Google Scholar] [CrossRef] [Green Version]
- Munday, P.L.; Jones, G.P.; Pratchett, M.S.; Williams, A.J. Climate change and the future for coral reef fishes. Fish Fish. 2008, 9, 261–285. [Google Scholar] [CrossRef]
- Przeslawski, R.; Ahyong, S.; Byrne, M.; Wörheide, G.; Hutchings, P. Beyond corals and fish: The effects of climate change on noncoral benthic invertebrates of tropical reefs. Glob. Chang. Biol. 2008, 14, 2773–2795. [Google Scholar] [CrossRef]
- Donelson, J.M.; Munday, P.L.; McCormick, M.I.; Pankhurst, N.W.; Pankhurst, P.M. Effects of elevated water temperature and food availability on the reproductive performance of a coral reef fish. Mar. Ecol. Prog. Ser. 2010, 401, 233–243. [Google Scholar] [CrossRef] [Green Version]
- Zarco-Perello, S.; Pratchett, M.; Liao, V. Temperature-growth performance curves for a coral reef fish, Acanthochromis polyacanthus. Galaxea J. Coral Reef Stud. 2012, 14, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.-C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengård, B.; et al. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355, eaai9214. [Google Scholar] [CrossRef] [PubMed]
- Animals and Temperature: Phenotypic and Evolutionary Adaptation; Johnston, I.A.; Bennett, A.F. (Eds.) Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Hoey, A.S.; Howells, E.; Johansen, J.L.; Hobbs, J.-P.A.; Messmer, V.; McCowan, D.M.; Wilson, S.K.; Pratchett, M.S. Recent Advances in Understanding the Effects of Climate Change on Coral Reefs. Diversity 2016, 8, 12. [Google Scholar] [CrossRef] [Green Version]
- Fields, P.A.; Graham, J.B.; Rosenblatt, R.H.; Somero, G.N. Effects of expected global climate change on marine faunas. Trends Ecol. Evol. 1993, 8, 361–367. [Google Scholar] [CrossRef]
- Jennings, S.; Brander, K. Predicting the effects of climate change on marine communities and the consequences for fisheries. J. Mar. Syst. 2010, 79, 418–426. [Google Scholar] [CrossRef]
- Kordas, R.L.; Harley, C.D.G.; O’Connor, M.I. Community ecology in a warming world: The influence of temperature on interspecific interactions in marine systems. J. Exp. Mar. Biol. Ecol. 2011, 400, 218–226. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O. Climate change, coral bleaching and the future of the world’s coral reefs. Mar. Freshw. Res. 1999. [Google Scholar] [CrossRef] [Green Version]
- Bellwood, D.R.; Hughes, T.P. Regional-scale assembly rules and biodiversity of coral reefs. Science 2001, 292, 1532–1534. [Google Scholar] [CrossRef] [Green Version]
- Lough, J.M. Small change, big difference: Sea surface temperature distributions for tropical coral reef ecosystems, 1950-2011. J. Geophys. Res. Ocean. 2012, 117. [Google Scholar] [CrossRef] [Green Version]
- Hughes, T.P.; Kerry, J.T.; Baird, A.H.; Connolly, S.R.; Dietzel, A.; Eakin, C.M.; Heron, S.F.; Hoey, A.S.; Hoogenboom, M.O.; Liu, G.; et al. Global warming transforms coral reef assemblages. Nature 2018, 556, 492–496. [Google Scholar] [CrossRef]
- Lough, J.M.; Anderson, K.D.; Hughes, T.P. Increasing thermal stress for tropical coral reefs: 1871-2017. Sci. Rep. 2018, 8, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tewksbury, J.J.; Huey, R.B.; Deutsch, C.A. ECOLOGY: Putting the Heat on Tropical Animals. Science 2008, 320, 1296–1297. [Google Scholar] [CrossRef]
- Sunday, J.M.; Bates, A.E.; Dulvy, N.K. Global analysis of thermal tolerance and latitude in ectotherms. Proc. R. Soc. B: Biol. Sci. 2011, 278, 1823–1830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habary, A.; Johansen, J.L.; Nay, T.J.; Steffensen, J.F.; Rummer, J.L. Adapt, move or die - how will tropical coral reef fishes cope with ocean warming? Glob. Chang. Biol. 2017, 23, 566–577. [Google Scholar] [CrossRef]
- Lough, J.M. 10th Anniversary Review: A changing climate for coral reefs. J. Environ. Monit. 2008, 10, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Kerry, J.T.; Álvarez-Noriega, M.; Álvarez-Romero, J.G.; Anderson, K.D.; Baird, A.H.; Babcock, R.C.; Beger, M.; Bellwood, D.R.; Berkelmans, R.; et al. Global warming and recurrent mass bleaching of corals. Nature 2017, 543, 373–377. [Google Scholar] [CrossRef]
- Hughes, T.P.; Anderson, K.D.; Connolly, S.R.; Heron, S.F.; Kerry, J.T.; Lough, J.M.; Baird, A.H.; Baum, J.K.; Berumen, M.L.; Bridge, T.C.; et al. Spatial and temporal patterns of mass bleaching of corals in the Anthropocene. Science 2018, 359, 80–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliver, J.K.; Berkelmans, R.; Eakin, C.M. Coral Bleaching in Space and Time. In Coral Bleaching: Patterns, Processes, Causes and Consequences; van Oppen, M.J.H., Lough, J.M., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 27–49. [Google Scholar] [CrossRef]
- Skirving, W.J.; Heron, S.F.; Marsh, B.L.; Liu, G.; De La Cour, J.L.; Geiger, E.F.; Eakin, C.M. The relentless march of mass coral bleaching: A global perspective of changing heat stress. Coral Reefs 2019, 38, 547–557. [Google Scholar] [CrossRef] [Green Version]
- Hughes, T.P.; Kerry, J.T.; Baird, A.H.; Connolly, S.R.; Chase, T.J.; Dietzel, A.; Hill, T.; Hoey, A.S.; Hoogenboom, M.O.; Jacobson, M.; et al. Global warming impairs stock–recruitment dynamics of corals. Nature 2019, 568, 387–390. [Google Scholar] [CrossRef]
- Raymundo, L.J.; Burdick, D.; Hoot, W.C.; Miller, R.M.; Brown, V.; Reynolds, T.; Gault, J.; Idechong, J.; Fifer, J.; Williams, A. Successive bleaching events cause mass coral mortality in Guam, Micronesia. Coral Reefs 2019, 38, 677–700. [Google Scholar] [CrossRef]
- Mora, C.; Ospína, A.F. Tolerance to high temperatures and potential impact of sea warming on reef fishes of Gorgona Island (tropical eastern Pacific). Mar. Biol. 2001, 139, 765–769. [Google Scholar] [CrossRef]
- Booth, D.; Beretta, G. Changes in a fish assemblage after a coral bleaching event. Mar. Ecol. Prog. Ser. 2002, 245, 205–212. [Google Scholar] [CrossRef] [Green Version]
- Spalding, M.D.; Jarvis, G.E. The impact of the 1998 coral mortality on reef fish communities in the Seychelles. Mar. Pollut. Bull. 2002, 44, 309–321. [Google Scholar] [CrossRef]
- Jones, G.P.; McCormick, M.I.; Srinivasan, M.; Eagle, J.V. Coral decline threatens fish biodiversity in marine reserves. Proc. Natl. Acad. Sci. USA 2004, 101, 8251–8253. [Google Scholar] [CrossRef] [Green Version]
- Ospina, A.F.; Mora, C. Effect of body size on reef fish tolerance to extreme low and high temperatures. Environ. Biol. Fishes 2004, 70, 339–343. [Google Scholar] [CrossRef]
- Roessig, J.M.; Woodley, C.M.; Cech, J.J.; Hansen, L.J. Effects of global climate change on marine and estuarine fishes and fisheries. Rev. Fish Biol. Fish. 2004, 14, 251–275. [Google Scholar] [CrossRef]
- Graham, N.A.J.; Wilson, S.K.; Jennings, S.; Polunin, N.V.C.; Bijoux, J.P.; Robinson, J. Dynamic fragility of oceanic coral reef ecosystems. Proc. Natl. Acad. Sci. USA 2006, 103, 8425–8429. [Google Scholar] [CrossRef] [Green Version]
- Garpe, K.C.; Yahya, S.A.S.; Lindahl, U.; Öhman, M.C. Long-term effects of the 1998 coral bleaching event on reef fish assemblages. Mar. Ecol. Prog. Ser. 2006, 315, 237–247. [Google Scholar] [CrossRef]
- Wilson, S.K.; Graham, N.A.J.; Pratchett, M.S.; Jones, G.P.; Polunin, N.V.C. Multiple disturbances and the global degradation of coral reefs: Are reef fishes at risk or resilient? Glob. Chang. Biol. 2006, 12, 2220–2234. [Google Scholar] [CrossRef]
- Pratchett, M.S.; Munday, P.; Wilson, S.K.; Graham, N.A.J.; Cinner, J.; Bellwood, D.R.; Jones, G.P.; Polunin, N.V.C.; McClanahan, T.R. Effects Of Climate-Induced Coral Bleaching On Coral-Reef Fishes‚ Ecological And Economic Consequences. In Oceanography and Marine Biology: An Annual Reviewiology; Gibson, R.N., Atkinson, R.J.A., Gordon, J.D.M., Eds.; Taylor & Francis: Milton Park, UK, 2008; pp. 257–302. [Google Scholar]
- Clark, T.D.; Roche, D.G.; Binning, S.A.; Speers-Roesch, B.; Sundin, J. Maximum thermal limits of coral reef damselfishes are size dependent and resilient to near-future ocean acidification. J. Exp. Biol. 2017, 220, 3519–3526. [Google Scholar] [CrossRef] [Green Version]
- Triki, Z.; Wismer, S.; Levorato, E.; Bshary, R. A decrease in the abundance and strategic sophistication of cleaner fish after environmental perturbations. Glob. Chang. Biol. 2018, 24, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Plaisance, L.; Knowlton, N.; Paulay, G.; Meyer, C. Reef-associated crustacean fauna: Biodiversity estimates using semi-quantitative sampling and DNA barcoding. Coral Reefs 2009, 28, 977–986. [Google Scholar] [CrossRef] [Green Version]
- Plaisance, L.; Caley, M.J.; Brainard, R.E.; Knowlton, N. The diversity of coral reefs: What are we missing? PLoS ONE 2011, 6, e25026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandl, S.J.; Tornabene, L.; Goatley, C.H.R.; Casey, J.M.; Morais, R.A.; Côté, I.M.; Baldwin, C.C.; Parravicini, V.; Schiettekatte, N.M.D.; Bellwood, D.R. Demographic dynamics of the smallest marine vertebrates fuel coral reef ecosystem functioning. Science 2019, 364, 1189–1192. [Google Scholar] [CrossRef] [PubMed]
- De Meeûs, T.; Renaud, F. Parasites within the new phylogeny of eukaryotes. Trends Parasitol. 2002, 18, 247–250. [Google Scholar] [CrossRef]
- Rohde, K. Ecology of Marine Parasites; University of Queensland Press: St Lucia, Australia, 1982; p. 245. [Google Scholar]
- Dobson, A.; Lafferty, K.D.; Kuris, A.M.; Hechinger, R.F.; Jetz, W. Homage to linnaeus: How many parasites? How many hosts? Light Evol. 2009, 2, 63–82. [Google Scholar] [CrossRef] [Green Version]
- Hatcher, M.J.; Dunn, A.M. Parasites in Ecological Communities: From Interactions to Ecosystems; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar] [CrossRef] [Green Version]
- Okamura, B.; Hartigan, A.; Naldoni, J. Extensive uncharted biodiversity: The parasite dimension. Integr. Comp. Biol. 2018, 56, 1132–1145. [Google Scholar] [CrossRef]
- Hudson, P.J.; Dobson, A.P.; Lafferty, K.D. Is a healthy ecosystem one that is rich in parasites? Trends Ecol. Evol. 2006, 21, 381–385. [Google Scholar] [CrossRef]
- Kuris, A.M.; Hechinger, R.F.; Shaw, J.C.; Whitney, K.L.; Aguirre-Macedo, L.; Boch, C.A.; Dobson, A.P.; Dunham, E.J.; Fredensborg, B.L.; Huspeni, T.C.; et al. Ecosystem energetic implications of parasite and free-living biomass in three estuaries. Nature 2008, 454, 515–518. [Google Scholar] [CrossRef]
- Hatcher, M.J.; Dick, J.T.A.; Dunn, A.M. Diverse effects of parasites in ecosystems: Linking interdependent processes. Front. Ecol. Environ. 2012, 10, 186–194. [Google Scholar] [CrossRef]
- Hatcher, M.J.; Dick, J.T.A.; Dunn, A.M. Parasites that change predator or prey behaviour can have keystone effects on community composition. Biol. Lett. 2014, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohde, K. Ecology and biogeography of marine parasites. Adv. Mar. Biol. 2002, 43, 1–86. [Google Scholar] [PubMed]
- Rohde, K. Species diversity of parasites on the Great Barrier Reef. Z. Für Parasitenkd. 1976, 50, 93–94. [Google Scholar] [CrossRef]
- Poulin, R.; Blasco-Costa, I.; Randhawa, H.S. Integrating parasitology and marine ecology: Seven challenges towards greater synergy. J. Sea Res. 2016, 113, 3–10. [Google Scholar] [CrossRef]
- Marcogliese, D.J. Implications of climate change for parasitism of animals in the aquatic environment. Can. J. Zool. 2001, 79, 1331–1352. [Google Scholar] [CrossRef]
- Sikkel, P.C.; Richardson, M.A.; Sun, D.; Narvaez, P.; Feeney, W.E.; Grutter, A.S. Changes in abundance of fish-parasitic gnathiid isopods associated with warm-water bleaching events on the northern Great Barrier Reef. Coral Reefs 2019, 38, 721–730. [Google Scholar] [CrossRef]
- Sikkel, P.C.; Welicky, R.L. The Ecological Significance of Parasitic Crustaceans. In Parasitic Crustacea; Springer: London, UK, 2019; pp. 421–477. [Google Scholar]
- Claar, D.C.; Wood, C.L. Pulse Heat Stress and Parasitism in a Warming World. Trends Ecol. Evol. 2020, 35, 704–715. [Google Scholar] [CrossRef]
- Gehman, A.-L.M.; Hall, R.J.; Byers, J.E. Host and parasite thermal ecology jointly determine the effect of climate warming on epidemic dynamics. Proc. Natl. Acad. Sci. USA 2018, 115, 744–749. [Google Scholar] [CrossRef] [Green Version]
- Brazenor, A.K.; Hutson, K.S. Effects of temperature and salinity on the life cycle of Neobenedenia sp. (Monogenea: Capsalidae) infecting farmed barramundi (Lates calcarifer). Parasitol. Res. 2015, 114, 1875–1886. [Google Scholar] [CrossRef]
- Grutter, A.S. Spatial and temporal variations of the ectoparasites of seven reef fish species from Lizard Island and Heron Island, Australia. Mar. Ecol. Prog. Ser. 1994, 115, 21–30. [Google Scholar] [CrossRef]
- Grutter, A.S. Parasite removal rates by the cleaner wrasse Labroides dimidiatus. Mar. Ecol. Prog. Ser. 1996, 130, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Grutter, A.S.; Poulin, R. Cleaning of Coral Reef Fishes by the Wrasse Labroides dimidiatus: Influence of Client Body Size and Phylogeny. Copeia 1998, 1998, 1447707. [Google Scholar] [CrossRef]
- Smit, N.J.; Davies, A.J. The curious life-style of the parasitic stages of Gnathiid isopods. Adv. Parasitol. 2004, 58, 289–391. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K. Life history of gnathiid isopods—Current knowledge and and future directions. Plankton Benthos Res. 2007, 2, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lafferty, K.D.; Kuris, A.M. Trophic strategies, animal diversity and body size. Trends Ecol. Evol. 2002, 17, 507–513. [Google Scholar] [CrossRef]
- Raffel, T.R.; Martin, L.B.; Rohr, J.R. Parasites as predators: Unifying natural enemy ecology. Trends Ecol. Evol. 2008, 23, 610–618. [Google Scholar] [CrossRef]
- Grutter, A.S. Parasite infection rather than tactile stimulation is the proximate cause of cleaning behaviour in reef fish. Proc. R. Soc. B Biol. Sci. 2001, 268, 1361–1365. [Google Scholar] [CrossRef] [Green Version]
- Grutter, A.S. Feeding ecology of the fish ectoparasite Gnathia sp. (Crustacea: Isopoda) from the Great Barrier Reef, and its implications for fish cleaning behaviour. Mar. Ecol. Prog. Ser. 2003, 259, 295–302. [Google Scholar] [CrossRef]
- Sikkel, P.C.; Cheney, K.L.; Côté, I.M. In situ evidence for ectoparasites as a proximate cause of cleaning interactions in reef fish. Anim. Behav. 2004, 68, 241–247. [Google Scholar] [CrossRef]
- Sikkel, P.C.; Herzlieb, S.E.; Kramer, D.L. Compensatory cleaner-seeking behavior following spawning in female yellowtail damselfish. Mar. Ecol. Prog. Ser. 2005, 296, 1–11. [Google Scholar] [CrossRef]
- Cheney, K.L.; Côté, I.M. Mutualism or parasitism? The variable outcome of cleaning symbioses. Biol. Lett. 2005, 1, 162–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binning, S.A.; Roche, D.G.; Grutter, A.S.; Colosio, S.; Sun, D.; Miest, J.; Bshary, R. Cleaner wrasse indirectly affect the cognitive performance of a damselfish through ectoparasite removal. Proc. R. Soc. B Biol. Sci. 2018, 285. [Google Scholar] [CrossRef] [PubMed]
- Sellers, J.C.; Holstein, D.M.; Botha, T.L.; Sikkel, P.C. Lethal and sublethal impacts of a micropredator on post-settlement Caribbean reef fishes. Oecologia 2019, 189, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.M.; Grutter, A.S. Parasitic isopods (Gnathia sp.) reduce haematocrit in captive blackeye thicklip (Labridae) on the Great Barrier Reef. J. Fish Biol. 2005, 66, 860–864. [Google Scholar] [CrossRef]
- Hayes, P.M.; Smit, N.J.; Grutter, A.S.; Davies, A.J. Unexpected response of a captive blackeye thicklip, Hemigymnus melapterus (Bloch), from Lizard Island, Australia, exposed to juvenile isopods Gnathia aureamaculosa Ferreira & Smit. J. Fish Dis. 2011, 34, 563–566. [Google Scholar] [CrossRef]
- Grutter, A.S.; Pickering, J.L.; McCallum, H.; McCormick, M.I. Impact of micropredatory gnathiid isopods on young coral reef fishes. Coral Reefs 2008, 27, 655–661. [Google Scholar] [CrossRef]
- Jones, C.M.; Grutter, A.S. Reef-based micropredators reduce the growth of post-settlement damselfish in captivity. Coral Reefs 2008, 27, 677–684. [Google Scholar] [CrossRef]
- Penfold, R.; Grutter, A.S.; Kuris, A.M.; McCormick, M.I.; Jones, C.M. Interactions between juvenile marine fish and gnathiid isopods: Predation versus micropredation. Mar. Ecol. Prog. Ser. 2008, 357, 111–119. [Google Scholar] [CrossRef]
- Artim, J.M.; Sellers, J.C.; Sikkel, P.C. Micropredation by gnathiid isopods on settlement stage reef fish in the eastern Caribbean Sea. Bull. Mar. Sci. 2015, 91, 479–487. [Google Scholar] [CrossRef]
- Grutter, A.S.; Blomberg, S.P.; Fargher, B.; Kuris, A.M.; McCormick, M.I.; Warner, R.R. Size-related mortality due to gnathiid isopod micropredation correlates with settlement size in coral reef fishes. Coral Reefs 2017, 36, 549–559. [Google Scholar] [CrossRef]
- Curtis, L.M.; Grutter, A.S.; Smit, N.J.; Davies, A.J. Gnathia aureamaculosa, a likely definitive host of Haemogregarina balistapi and potential vector for Haemogregarina bigemina between fishes of the Great Barrier Reef, Australia. Int. J. Parasitol. 2013, 43, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Honma, Y.; Chiba, A. Pathological changes in the branchial chamber wall of stingrays, Dasyatis spp., associated with the presence of juvenile gnathiids (Isopoda, Crustacea). Fish Pathol. 1991, 26, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Grutter, A.S. Spatiotemporal Variation and Feeding Selectivity in the Diet of the Cleaner Fish Labroides dimidiatus. Copeia 1997, 1997, 1447754. [Google Scholar] [CrossRef]
- Grutter, A.S. Cleaning symbioses from the parasites’ perspective. Parasitology 2002, 124, 65–81. [Google Scholar] [CrossRef]
- Grutter, A.S.; Blomberg, S.P.; Box, S.; Bshary, R.; Ho, O.; Madin, E.M.P.; McClure, E.C.; Meekan, M.G.; Murphy, J.M.; Richardson, M.A.; et al. Changes in local free-living parasite populations in response to cleaner manipulation over 12 years. Oecologia 2019, 190, 783–797. [Google Scholar] [CrossRef]
- Demairé, C.; Triki, Z.; Binning, S.A.; Glauser, G.; Roche, D.G.; Bshary, R. Reduced access to cleaner fish negatively impacts the physiological state of two resident reef fishes. Mar. Biol. 2020, 167, 48. [Google Scholar] [CrossRef]
- Grutter, A.; Bejarano, S.; Cheney, K.; Goldizen, A.; Sinclair-Taylor, T.; Waldie, P. Effects of the cleaner fish Labroides dimidiatus on grazing fishes and coral reef benthos. Mar. Ecol. Prog. Ser. 2020, 643, 99–114. [Google Scholar] [CrossRef]
- Hughes, T.P.; Kerry, J.T.; Simpson, T. Large-scale bleaching of corals on the Great Barrier Reef. Ecology 2018, 99, 501. [Google Scholar] [CrossRef] [Green Version]
- Arceo, H.O.; Quibilan, M.C.; Aliño, P.M.; Lim, G.; Licuanan, W.Y. Coral bleaching in Philippine reefs: Coincident evidences with mesoscale thermal anomalies. Bull. Mar. Sci. 2001, 69, 579–593. [Google Scholar]
- Raymundo, L.; Maypa, A. Recovery of the Apo Island Marine Reserve, Philippines, 2 years after the El Niño bleaching event. Coral Reefs 2002, 21, 260–261. [Google Scholar] [CrossRef]
- Magdaong, E.T.; Fujii, M.; Yamano, H.; Licuanan, W.Y.; Maypa, A.; Campos, W.L.; Alcala, A.C.; White, A.T.; Apistar, D.; Martinez, R. Long-term change in coral cover and the effectiveness of marine protected areas in the Philippines: A meta-analysis. Hydrobiologia 2014, 733, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Grutter, A.S.; Feeney, W.E.; McClure, E.C.; Narvaez, P.; Smit, N.J.; Sun, D.; Sikkel, P.C. Practical methods for culturing parasitic gnathiid isopods. Int. J. Parasitol. 2020, 50, 825–837. [Google Scholar] [CrossRef] [PubMed]
- Sikkel, P.C.; Tuttle, L.J.; Cure, K.; Coile, A.M.; Hixon, M.A. Low Susceptibility of Invasive Red Lionfish (Pterois volitans) to a Generalist Ectoparasite in Both Its Introduced and Native Ranges. PLoS ONE 2014, 9, e95854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, T.R.N.; Sikkel, P.C. Habitat associations of fish-parasitic gnathiid isopods in a shallow reef system in the central Philippines. Mar. Biodivers. 2017, 49, 83–96. [Google Scholar] [CrossRef]
- Artim, J.M.; Sikkel, P.C. Comparison of sampling methodologies and estimation of population parameters for a temporary fish ectoparasite. Int. J. Parasitol. Parasites Wildl. 2016, 5, 145–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graba-Landry, A.C.; Loffler, Z.; McClure, E.C.; Pratchett, M.S.; Hoey, A.S. Impaired growth and survival of tropical macroalgae (Sargassum spp.) at elevated temperatures. Coral Reefs 2020, 39, 475–486. [Google Scholar] [CrossRef]
- Pacific Ocean: Great Barrier Reef: Sea Water Temperature Compared to Long Term Averages. Available online: http://data.aims.gov.au/aimsrtds/yearlytrends.xhtml (accessed on 13 November 2018).
- Water Temperature at Lizard Island Graph. Australian Institute of Marine Science AIMS. Available online: http://data.aims.gov.au/aimsrtds/datatool.xhtml?from=2018-10-15&thru=2018-10-20&period=MONTH&aggregations=AVG&channels=3523 (accessed on 13 November 2018).
- NOAA Coral Reef Watch Program. Available online: https://coralreefwatch.noaa.gov/satellite/vs/philippines.php (accessed on 5 January 2018).
- Therneau, T.M.; Grambsch, P.M. Modeling Survival Data: Extending the Cox Model; Springer: New York, NY, USA, 2000. [Google Scholar]
- Therneau, T. Coxme: Mixed Effects Cox Models. R Package Version 2.2-3. 2015. Available online: http://CRAN.R-project.org/package=coxme (accessed on 17 July 2020).
- Fox, J.; Weisberg, S. Multivariate Linear Models in R. In An R Companion to Applied Regression, 2nd ed.; Thousand Oaks: Los Angeles, CA, USA, 2011; pp. 1–31. [Google Scholar]
- Ganachaud, A.; Sen Gupta, A.S.; Orr, J.C.; Wijffels, S.E.; Ridgway, K.R.; Hemer, M.A.; Maes, C.; Steinberg, C.R.; Tribollet, A.D.; Qiu, B.; et al. Observed and expected changes to the tropical Pacific Ocean. In Vulnerability of Tropical Pacific Fisheries and Aquaculture to Climate Change; Bell, J.D., Johnson, J.E., Hobday, A.J., Eds.; Secretariat of the Pacific Community: Noumea, New Caledonia, 2011; pp. 101–187. [Google Scholar]
- Huey, R.B.; Kingsolver, J.G. Climate Warming, Resource Availability, and the Metabolic Meltdown of Ectotherms. Am. Nat. 2019, 194, E140–E150. [Google Scholar] [CrossRef]
- Smale, D.A.; Wernberg, T.; Oliver, E.C.J.; Thomsen, M.; Harvey, B.P.; Straub, S.C.; Burrows, M.T.; Alexander, L.V.; Benthuysen, J.A.; Donat, M.G.; et al. Marine heatwaves threaten global biodiversity and the provision of ecosystem services. Nat. Clim. Chang. 2019, 9, 306–312. [Google Scholar] [CrossRef] [Green Version]
- Frölicher, T.L.; Fischer, E.M.; Gruber, N. Marine heatwaves under global warming. Nature 2018, 560, 360–364. [Google Scholar] [CrossRef]
- Oliver, E.C.J.; Donat, M.G.; Burrows, M.T.; Moore, P.J.; Smale, D.A.; Alexander, L.V.; Benthuysen, J.A.; Feng, M.; Gupta, A.S.; Hobday, A.J.; et al. Longer and more frequent marine heatwaves over the past century. Nat. Commun. 2018, 9, 1324. [Google Scholar] [CrossRef]
- Compton, T.J.; Rijkenberg, M.J.A.; Drent, J.; Piersma, T. Thermal tolerance ranges and climate variability: A comparison between bivalves from differing climates. J. Exp. Mar. Biol. Ecol. 2007, 352, 200–211. [Google Scholar] [CrossRef] [Green Version]
- Stillman, J.H.; Somero, G.N. A comparative analysis of the upper thermal tolerance limits of eastern pacific porcelain crabs, genus Petrolisthes: Influences of latitude, vertical zonation, acclimation, and phylogeny. Physiol. Biochem. Zool. 2000, 73, 200–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attrill, M.J.; Kelmo, F.; Jones, M.B. Impact of the 1997-98 El Niño event on the coral reef-associated echinoderm assemblage from northern Bahia, northeastern Brazil. Clim. Res. 2004, 26, 151–158. [Google Scholar] [CrossRef] [Green Version]
- Chan, B.K.K.; Morritt, D.; De Pirro, M.; Leung, K.M.Y.; Williams, G.A. Summer mortality: Effects on the distribution and abundance of the acorn barnacle Tetraclita japonica on tropical shores. Mar. Ecol. Prog. Ser. 2006, 328, 195–204. [Google Scholar] [CrossRef]
- Jokiel, P.L.; Coles, S.L. Coral Reefs to elevated temperature. Environ. Conserv. 1990, 8, 155–162. [Google Scholar]
- Mouritsen, K.N.; Poulin, R. Parasitism, climate oscillations and the structure of natural communities. Oikos 2002, 97, 462–468. [Google Scholar] [CrossRef] [Green Version]
- Poulin, R. Global warming and temperature-mediated increases in cercarial emergence in trematode parasites. Parasitology 2006, 132, 143–151. [Google Scholar] [CrossRef]
- Thieltges, D.W.; Fredensborg, B.L.; Studer, A.; Poulin, R. Large-scale patterns in trematode richness and infection levels in marine crustacean hosts. Mar. Ecol. Prog. Ser. 2009, 389, 139–147. [Google Scholar] [CrossRef]
- Pechenik, J.A.; Fried, B. Effect of temperature on survival and infectivity of Echinostoma trivolvis cercariae: A test of the energy limitation hypothesis. Parasitology 1995, 111, 373–378. [Google Scholar] [CrossRef]
- Lyholt, H.C.K.; Buchmann, K. Diplostomum spathaceum:effects of temperature and light on cercarial shedding and infection of rainbow trout. Dis. Aquat. Org. 1996, 25, 169–173. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, A.M. The influence of temperature on the survival and infectivity of the cercariae of Echinoparyphium recurvatum (Digenea: Echinostomatidae). Parasitology 1999, 118, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Toledo, R.; Muñoz-Antoli, C.; Pérez, M.; Esteban, J.G. Survival and infectivity of Hypoderaeum conoideum and Euparyphium albuferensis cercariae under laboratory conditions. J. Helminthol. 1999, 73, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Mouritsen, K.N. The Hydrobia ulvae—Maritrema subdolum association: Influence of temperature, salinity, light, water-pressure and secondary host exudates on cercarial emergence and longevity. J. Helminthol. 2002, 76, 341–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thieltges, D.W.; Rick, J. Effect of temperature on emergence, survival and infectivity of cercariae of the marine trematode Renicola roscovita (Digenea: Renicolidae). Dis. Aquat. Org. 2006, 73, 63–68. [Google Scholar] [CrossRef]
- Leiva, N.V.; Manríquez, P.H.; Aguilera, V.M.; González, M.T. Temperature and pCO2 jointly affect the emergence and survival of cercariae from a snail host: Implications for future parasitic infections in the Humboldt Current system. Int. J. Parasitol. 2019, 49, 49–61. [Google Scholar] [CrossRef]
- Franzova, V.A.; MacLeod, C.D.; Wang, T.; Harley, C.D.G. Complex and interactive effects of ocean acidification and warming on the life span of a marine trematode parasite. Int. J. Parasitol. 2019, 49, 1015–1021. [Google Scholar] [CrossRef]
- Conley, D.C.; Curtis, M.A. Effects of temperature and photoperiod on the duration of hatching, swimming, and copepodid survival of the parasitic copepod Salmincola edwardsii. Can. J. Zool. 1993, 71, 972–976. [Google Scholar] [CrossRef]
- Hirazawa, N.; Takano, R.; Hagiwara, H.; Noguchi, M.; Narita, M. The influence of different water temperatures on Neobenedenia girellae (Monogenea) infection, parasite growth, egg production and emerging second generation on amberjack Seriola dumerili (Carangidae) and the histopathological effect of this parasite on fish skin. Aquaculture 2010, 299, 2–7. [Google Scholar] [CrossRef]
- Mahmoud, N.E.; Fahmy, M.M.; Abuowarda, M.M.; Zaki, M.; Ismail, E.M.; Ismael, E.S. Mediterranean sea fry; a source of isopod infestation problem in Egypt with reference to the effect of salinity and temperature on the survival of Livoneca redmanii (Isopoda: Cymothoidae) juvenile stages. J. Egypt. Soc. Parasitol. 2019, 49, 235–242. [Google Scholar]
- Evans, N.A. The influence of environmental temperature upon transmission of the cercariae of Echinostoma liei (Digenea: Echinostomatidae). Parasitology 1985, 90, 269–275. [Google Scholar] [CrossRef]
- Lo, C.T.; Lee, K.M. Pattern of Emergence and the Effects of Temperature and Light on the Emergence and Survival of Heterophyid Cercariae (Centrocestus formosanus and Haplorchis pumilio). J. Parasitol. 1996, 82, 347. [Google Scholar] [CrossRef] [PubMed]
- Fried, B.; Ponder, E.L. Effects of temperature on survival, infectivity and in vitro encystment of the cercariae of Echinostoma caproni. J. Helminthol. 2003, 77, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Tucker, C.S.; Sommerville, C.; Wootten, R. The effect of temperature and salinity on the settlement and survival of copepodids of Lepeophtheirus salmonis (Krøyer, 1837) on Atlantic salmon, Salmo salar L. J. Fish Dis. 2000, 23, 309–320. [Google Scholar] [CrossRef]
- Peck, L.S.; Webb, K.E.; Bailey, D.M. Extreme sensitivity of biological function to temperature in Antarctic marine species. Funct. Ecol. 2004, 18, 625–630. [Google Scholar] [CrossRef] [Green Version]
- Pörtner, H.O.; Knust, R. Climate change affects marine fishes through the oxygen limitation of thermal tolerance. Science 2007, 315, 95–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyrowitsch, D.; Christensen, N.; Hindsbo, O. Effects of temperature and host density on the snail-finding capacity of cercariae of Echinostoma caproni (Digenea: Echinostomatidae). Parasitology 1991, 102, 391–395. [Google Scholar] [CrossRef]
- Cech, J.J.; Moyle, P.B. Fishes: An Introduction to Ichthyology, 5th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 2004. [Google Scholar]
- Gagliano, M.; McCormick, M.I.; Meekan, M.G. Temperature-induced shifts in selective pressure at a critical developmental transition. Oecologia 2007, 152, 219–225. [Google Scholar] [CrossRef]
- Nilsson, G.E.; Crawley, N.; Lunde, I.G.; Munday, P.L. Elevated temperature reduces the respiratory scope of coral reef fishes. Glob. Chang. Biol. 2009, 15, 1405–1412. [Google Scholar] [CrossRef]
- Nay, T.J.; Johansen, J.L.; Habary, A.; Steffensen, J.F.; Rummer, J.L. Behavioural thermoregulation in a temperature-sensitive coral reef fish, the five-lined cardinalfish (Cheilodipterus quinquelineatus). Coral Reefs 2015, 34, 1261–1265. [Google Scholar] [CrossRef]
- Jones, C.M.; Miller, T.L.; Grutter, A.S.; Cribb, T.H. Natatory-stage cymothoid isopods: Description, molecular identification and evolution of attachment. Int. J. Parasitol. 2008, 38, 477–491. [Google Scholar] [CrossRef]
- Shodipo, M.O.; Gomez, R.D.C.; Welicky, R.L.; Sikkel, P.C. Apparent kleptoparasitism in fish—parasitic gnathiid isopods. Parasitol. Res. 2018, 118, 653–655. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, M.D.; Artim, J.D.; Hendrick, G.C.; Packard, A.J.; Sikkel, P.C. Fish-Parasitic Gnathiid Isopods Metamorphose Following Invertebrate-Derived Meal. J. Parasitol. 2019, 105, 793. [Google Scholar] [CrossRef] [PubMed]
- Ota, Y.; Hoshino, O.; Hirose, M.; Tanaka, K.; Hirose, E. Third-stage larva shifts host fish from teleost to elasmobranch in the temporary parasitic isopod, Gnathia trimaculata (Crustacea; Gnathiidae). Mar. Biol. 2012, 159, 2333–2347. [Google Scholar] [CrossRef]
- Sikkel, P.C.; Welicky, R.L.; Artim, J.M.; McCammon, A.M.; Sellers, J.C.; Coile, A.M.; Jenkins, W.G. Nocturnal migration reduces exposure to micropredation in a coral reef fish. Bull. Mar. Sci. 2017, 93, 475–489. [Google Scholar] [CrossRef]
- Artim, J.M.; Sikkel, P.C. Live coral repels a common reef fish ectoparasite. Coral Reefs 2013, 32, 487–494. [Google Scholar] [CrossRef]
- Artim, J.M.; Nicholson, M.D.; Hendrick, G.C.; Brandt, M.; Smith, T.; Sikkel, P.C. Abundance of a Cryptic Generalist Parasite Reflects Degradation of an Ecosystem. Ecosphere 2020, in press. [Google Scholar]
- Dupont, S.; Pörtner, H. Get ready for ocean acidification. Nature 2013, 498, 429. [Google Scholar] [CrossRef]
- Paula, J.R.; Otjacques, E.; Hildebrandt, C.; Grutter, A.S.; Rosa, R. Ocean Acidification Does Not Affect Fish Ectoparasite Survival. Oceans 2020, 1, 3. [Google Scholar] [CrossRef] [Green Version]
- Carlson, C.J.; Burgio, K.R.; Dougherty, E.R.; Phillips, A.J.; Bueno, V.M.; Clements, C.F.; Castaldo, G.; Dallas, T.A.; Cizauskas, C.A.; Cumming, G.S.; et al. Parasite biodiversity faces extinction and redistribution in a changing climate. Sci. Adv. 2017, 3. [Google Scholar] [CrossRef] [Green Version]
- Carlson, C.J.; Cizauskas, C.A.; Burgio, K.R.; Clements, C.F.; Harris, N.C. The More Parasites, the Better? Science 2013, 342, 1041. [Google Scholar] [CrossRef] [Green Version]
- Cizauskas, C.A.; Carlson, C.J.; Burgio, K.R.; Clements, C.F.; Dougherty, E.R.; Harris, N.C.; Phillips, A.J. Parasite vulnerability to climate change: An evidence-based functional trait approach. R. Soc. Open Sci. 2017, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Df | Chisq | Pr(>Chisq) | ||
|---|---|---|---|---|
| Temperature | 2 | 183.76 | <0.0001 | *** |
| Headwidth | 1 | 1.87 | 0.1719 | |
| Stage | 2 | 26.93 | <0.0001 | *** |
| Temperature × Headwidth | 2 | 0.07 | 0.9639 | |
| Temperature × Stage | 4 | 3.06 | 0.5471 | |
| Headwidth × Stage | 2 | 23.85 | <0.0001 | *** |
| Temperature × Headwidth × Stage | 4 | 6.51 | 0.1642 |
| Df | Chisq | Pr(>Chisq) | ||
|---|---|---|---|---|
| Temperature | 2 | 28.2063 | 7.50 × 10−7 | *** |
| Headwidth | 1 | 1.9549 | 0.16206 | |
| Stage | 2 | 21.3179 | 2.35 × 10−5 | *** |
| Temperature × Headwidth | 2 | 6.0753 | 0.04795 | * |
| Temperature × Stage | 4 | 13.6392 | 0.00854 | ** |
| Headwidth × Stage | 2 | 3.0686 | 0.21561 | |
| Temperature × Headwidth × Stage | 4 | 3.3548 | 0.5003 |
| Trial | Df | Chisq | Pr(>Chisq) | |
|---|---|---|---|---|
| 1 | Temperature | 4 | 24.927 | <0.0001 |
| 2 | Temperature | 4 | 8.4374 | 0.07681 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shodipo, M.O.; Duong, B.; Graba-Landry, A.; Grutter, A.S.; Sikkel, P.C. Effect of Acute Seawater Temperature Increase on the Survival of a Fish Ectoparasite. Oceans 2020, 1, 215-236. https://doi.org/10.3390/oceans1040016
Shodipo MO, Duong B, Graba-Landry A, Grutter AS, Sikkel PC. Effect of Acute Seawater Temperature Increase on the Survival of a Fish Ectoparasite. Oceans. 2020; 1(4):215-236. https://doi.org/10.3390/oceans1040016
Chicago/Turabian StyleShodipo, Mary O., Berilin Duong, Alexia Graba-Landry, Alexandra S. Grutter, and Paul C. Sikkel. 2020. "Effect of Acute Seawater Temperature Increase on the Survival of a Fish Ectoparasite" Oceans 1, no. 4: 215-236. https://doi.org/10.3390/oceans1040016
APA StyleShodipo, M. O., Duong, B., Graba-Landry, A., Grutter, A. S., & Sikkel, P. C. (2020). Effect of Acute Seawater Temperature Increase on the Survival of a Fish Ectoparasite. Oceans, 1(4), 215-236. https://doi.org/10.3390/oceans1040016