
Relationship between Body and Otolith Morphological Characteristics of Sabre Squirrelfish (Sargocentron spiniferum) from the Southern Red Sea: Difference between Right and Left Otoliths


 ,
,  ,
, 
Abstract
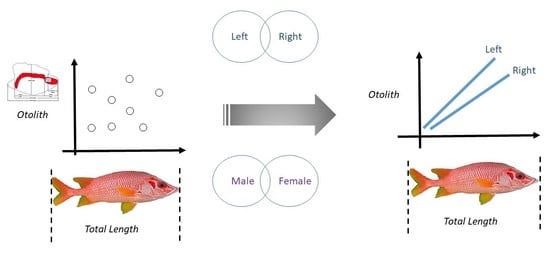
:
1. Introduction
2. Materials and Methods
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Randall, J.; Greenfield, D. Holocentridae: Squirrelfishes (soldierfishes). In FAO Species Identification Guide for Fishery Purposes: The Living Marine Resources of the Western Central Pacific; FAO: Rome, Italy, 1999; pp. 2225–2256. [Google Scholar]
- Randall, J.E. Zoogeography of shore fishes of the Indo-Pacific region. Zool. Stud. 1998, 37, 227–268. [Google Scholar]
- Lieske, E.; Myers, R. Coral Reef Fishes: Caribbean, Indian Ocean and Pacific Ocean Including the Red Sea, USA. Princeton University Press: Princeton, NJ, USA, 1994. [Google Scholar]
- Kuiter, R.H.; Tonozuka, T. Pictorial Guide to Indonesian Reef Fishes; Zoonetics: Sydney, Australia, 2001. [Google Scholar]
- Dubois, M. Effets Combinés de la Pêche et des Perturbations Naturelles sur la Dynamique des Ecosystèmes Coralliens. Ph.D. Thesis, École Pratique des Hautes Études, Paris, France, 2019. [Google Scholar]
- Mohammad, A.S.; Mehanna, S.F.; Osman, Y.A.A.; El-Mahdy, S.M. Age, growth and population parameters of the spiny squirrelfish, Sargocentron spiniferum (Forsskål, 1775) from Shalateen fishing area, Red Sea, Egypt. Egypt. J. Aquat. Biol. Fisheries 2020, 32, 469–480. [Google Scholar] [CrossRef] [Green Version]
- Sea around US: Fisheries, Ecosystems & Biodiversity. Available online: http://www.seaaroundus.org/data/#/msy/30606507 (accessed on 23 July 2021).
- Vitale, F.; Worsøe Clausen, L.; Chonchúir, G.N. Handbook of fish age estimation protocols and validation methods. ICES Coop. Res. Rep. 2019, 346, 1–180. [Google Scholar]
- Panfili, J.; de Pontual, H.; Troadec, J.-P.; Wright, P.J. Manual of Fish Sclerochronology; Ifremer-IRD: Brest, France, 2002; pp. 1–463. [Google Scholar]
- Tuset, V.; Lozano, I.; González, J.; Pertusa, J.; GarcíaDíaz, M. Shape indices to identify regional differences in otolith morphology of comber, Serranus cabrilla (L., 1758). J. Appl. Ichthyol. 2003, 19, 88–93. [Google Scholar] [CrossRef]
- Mehanna, S.; Jawad, L.; Ahmed, Y.; Abu ElRegal, M.; Dawood, D. Relationships between fish size and otolith measurements for Chlorurus sordidus (Forsskål, 1775) and Hipposcarus harid (Forsskål, 1775) from the Red Sea coast of Egypt. J. Appl. Ichthyol. 2016, 32, 356–358. [Google Scholar] [CrossRef]
- Tuset, V.M.; Lombarte, A.; Gonzalez, J.; Pertusa, J.; Lorente, M. Comparative morphology of the sagittal otolith in Serranus spp. J. Fish Biol. 2003, 63, 1491–1504. [Google Scholar] [CrossRef]
- Campana, S.E.; Casselman, J.M. Stock discrimination using otolith shape analysis. Can. J. Fish. Aquat. Sci. 1993, 50, 1062–1083. [Google Scholar] [CrossRef]
- Cardinale, M.; Doering-Arjes, P.; Kastowsky, M.; Mosegaard, H. Effects of sex, stock, and environment on the shape of known-age Atlantic cod (Gadus morhua) otoliths. Can. J. Fish. Aquat. Sci. 2004, 61, 158–167. [Google Scholar] [CrossRef]
- Rooker, J.R.; Secor, D.H.; DeMetrio, G.; Kaufman, A.J.; Ríos, A.B.; Tiina, V. Evidence of trans-Atlantic movement and natal homing of bluefin tuna from stable isotopes in otoliths. Mar. Ecol.-Prog. Ser. 2008, 368, 231–239. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, S.; Yazicioglu, O.; Saygin, S.A.; Polat, N. Relationships of otolith dimensions with body length of European perch, Perca fluviatilis L., 1758 from Lake Ladik, Turkey. Pakistan J. Zool. 2014, 46, 1231–1238. [Google Scholar]
- Zischke, M.T.; Litherland, L.; Tilyard, B.R.; Stratford, N.J.; Jones, E.L.; Wang, Y.-G. Otolith morphology of four mackerel species (Scomberomorus spp.) in Australia: Species differentiation and prediction for fisheries monitoring and assessment. Fish. Res. 2016, 176, 39–47. [Google Scholar] [CrossRef]
- Zorica, B.; Sinovi, G.; Kel, V. Preliminary data on the study of otolith morphology of five pelagic fish species from the Adriatic Sea (Croatia). Acta Adriat. 2010, 51, 89–96. [Google Scholar]
- Sadighzadeh, Z.; Tuset, V.M.; Valinassab, T.; Dadpour, M.R.; Lombarte, A. Comparison of different otolith shape descriptors and morphometrics for the identification of closely related species of Lutjanus spp. from the Persian Gulf. Mar. Biol. Res. 2012, 8, 802–814. [Google Scholar] [CrossRef]
- Morat, F.; Letourneur, Y.; Nérini, D.; Banaru, D.; Batjakas, I.E. Discrimination of red mullet populations (Teleostean, Mullidae) along multi-spatial and ontogenetic scales within the Mediterranean basin on the basis of otolith shape analysis. Aquat. Living Resour. 2012, 25, 27–39. [Google Scholar] [CrossRef]
- Tuset, V.M.; Rosin, P.L.; Lombarte, A. Sagittal otolith shape used in the identification of fishes of the genus Serranus. Fish. Res. 2006, 81, 316–325. [Google Scholar] [CrossRef]
- Škeljo, F.; Ferri, J. The use of otolith shape and morphometry for identification and size-estimation of five wrasse species in predator-prey studies. J. Appl. Ichthyol. 2011, 28, 524–530. [Google Scholar] [CrossRef]
- Jawad, L.A.; Hoedemakers, K.; Ibáñez, A.; Ahmed, Y.; Abu ElRegal, M.; Mehanna, S. Morphology study of the otoliths of the parrotfish, Chlorurus sordidus (Forsskål, 1775) and Hipposcarus harid (Forsskål, 1775) from the Red Sea coast of Egypt (Family: Scaridae). J. Mar. Biol. Assoc. United Kingd. 2017, 98, 819–828. [Google Scholar] [CrossRef] [Green Version]
- Disspain, M.C.F.; Ulm, S.; Gillanders, B.M. Otoliths in archaeology: Methods, applications and future prospects. J. Archaeol. Sci. Rep. 2016, 6, 623–632. [Google Scholar] [CrossRef]
- Mahé, K.; Bellamy, E.; Delpech, J.P.; Lazard, C.; Salaun, M.; Vérin, Y.; Coppin, F.; Travers-Trolet, M. Evidence of a relationship between weight and total length of marine fish in the North-eastern Atlantic Ocean: Physiological, spatial and temporal variations. J. Mar. Biol. Assoc. UK 2018, 98, 617–625. [Google Scholar] [CrossRef] [Green Version]
- Mahé, K.; Ider, D.; Massaro, A.; Hamed, O.; Jurado-Ruzafa, A.; Gonçalves, P.; Anastasopoulou, A.; Jadaud, A.; Mytilineou, C.; Elleboode, R. Directional bilateral asymmetry in otolith morphology may affect fish stock discrimination based on otolith shape analysis. ICES J. Mar. Sci. 2018, 76, 232–243. [Google Scholar] [CrossRef]
- Rohlf, F. tpsDig 2.10; Department of Ecology and Evolution, State University of New York: Stony Brook, NY, USA, 2006. [Google Scholar]
- Russ, J. Computer-Assisted Microscopy.-The Measurement and Analysis of Image; Plenum Press Corp: New York, NY, USA, 1990; pp. 71–79. [Google Scholar]
- Pavlov, D.A.; Emelyanova, N.G.; Ha, V.T.; Thuan, L.T.B. Otolith morphology, age, and growth of freckled goatfish Upeneus tragula (Mullidae) in the coastal zone of Vietnam. J. Ichthyol. 2015, 55, 363–372. [Google Scholar] [CrossRef]
- Mahe, K.; Evano, H.; Mille, T.; Muths, D.; Bourjea, J. Otolith shape as a valuable tool to evaluate the stock structure of swordfish Xiphias gladius in the Indian Ocean. Afr. J. Mar. Sci. 2016, 38, 457–464. [Google Scholar] [CrossRef]
- Kabakli, F.; Ergüden, D. Relationships between Fish Length and Otolith Dimensions of Redcoat, Sargocentron rubrum (Forsskal, 1775) in the Southeastern Mediterranean Sea, Turkey. Turk. J. Marit. Mar. Mar. Sci. 2018, 4, 156–162. [Google Scholar]
- Osman, A.G.M.; Farrag, M.M.; Mehanna, S.F.; Osman, Y.A. Use of otolithic morphometrics and ultrastructure for comparing between three goatfish species (family: Mullidae) from the northern Red Sea, Hurghada, Egypt. Iran. J. Fish. Sci. 2020, 19, 814–832. [Google Scholar]
- Fox, J.; Weisberg, S. Multivariate linear models in R. An R Companion to Applied Regression; Sage Publications: Los Angeles, CA, USA, 2011. [Google Scholar]
- R Development Core Team. R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Torres, G.J.; Lombarte, A.; Morales-Nin, B. Variability of the sulcus acusticus in the sagittal otolith of the genus Merluccius (Merlucciidae). Fish. Res. 2000, 46, 5–13. [Google Scholar] [CrossRef]
- Harvey, J.T.; Loughlin, T.R.; Perez, M.A.; Oxman, D.S. Relationship Between Fish Size and Otolith Length for 63 Species of Fishes from the Eastern North Pacific Ocean; (NOAA Technical Report NMFS, 150); NOAA/National Marine Fisheries Service: Seattle, WA, USA, 2000. [Google Scholar]
- Fossen, I.; Albert, O.T.; Nilssen, E.M. Improving the precision of ageing assessments for long rough dab by using digitised pictures and otolith measurements. Fish. Res. 2003, 60, 53–64. [Google Scholar] [CrossRef]
- Lychakov, D.V.; Rebane, Y.T.; Lombarte, A.; Fuiman, L.; Takabayashi, A. Fish otolith asymmetry: Morphometry and modeling. Hear. Res. 2006, 219, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Morat, F.; Banaru, D.; Mérigot, B.; Batjakas, I.E.; Betoulle, S.; Vignon, M.; Lecomte-Finiger, R.; Letourneur, Y. Relationships between fish length and otolith length for nine teleost fish species from the Mediterranean basin, Kerguelen Islands, and Pacific Ocean. Cybium 2008, 32, 265–269. [Google Scholar]
- Pavlov, D. Differentiation of three species of the genus Upeneus (Mullidae) based on otolith shape analysis. J. Ichthyol. 2016, 56, 37–51. [Google Scholar] [CrossRef]
- Mahé, K. Sources de Variation de la Forme des Otolithes: Implications Pour la Discrimination des Stocks de Poissons. Ph.D. Thesis, Université du Littoral Côte d’Opale, Boulogne-sur-Mer, France, 2019. [Google Scholar]
- Mahé, K.; MacKenzie, K.; Ider, D.; Massaro, A.; Hamed, O.; Jurado-Ruzafa, A.; Gonçalves, P.; Anastasopoulou, A.; Jadaud, A.; Mytilineou, C.; et al. Directional bilateral asymmetry in fish otolith: A potential tool to evaluate stock boundaries? Symmetry 2021, 13, 987. [Google Scholar] [CrossRef]
- Hilbig, R.; Knie, M.; Shcherbakov, D.; Anken, R.H. Analysis of behaviour and habituation of fish exposed to diminished gravity in correlation to inner ear stone formation—A sounding rocket experiment (TEXUS 45). In Proceedings of the 20th ESA Symposium on European Rocket and Balloon Programmes and Related Research, Hyere, France, 22–26 May 2011. [Google Scholar]
- Lemberget, T.; Mccormick, M.I. Replenishment success linked to fluctuating asymmetry in larval fish. Oecologia 2009, 159, 83–93. [Google Scholar] [CrossRef]
- Gagliano, M.; McCormick, M.I. Feeding history influences otolith shape in tropical fish. Mar. Ecol. Prog. Ser. 2004, 278, 291–296. [Google Scholar] [CrossRef]
- Swan, C.M.; Palmer, M.A. Preferential feeding by an aquatic consumer mediates non-additive decomposition of speciose leaf litter. Oecologia 2006, 149, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Vignon, M.; Morat, F. Environmental and genetic determinant of otolith shape revealed by a non-indigenous tropical fish. Mar. Ecol. Prog. Ser. 2010, 411, 231–241. [Google Scholar] [CrossRef]
- Bose, A.P.H.; Zimmermann, H.; Winkler, G.; Kaufmann, A.; Strohmeier, T.; Koblmüller, S.; Sefc, K.M. Congruent geographic variation in saccular otolith shape across multiple species of African cichlids. Sci. Rep. 2020, 10, 12820. [Google Scholar] [CrossRef] [PubMed]
- Schulz-Mirbach, T.; Ladich, F.; Plath, M.; Heß, M. Enigmatic ear stones: What we know about the functional role and evolution of fish otoliths. Biol. Rev. 2019, 94, 457–482. [Google Scholar] [CrossRef]
- Burke, N.; Brophy, D.; King, P.A. Otolith shape analysis: Its application for discriminating between stocks of Irish Sea and Celtic Sea herring (Clupea harengus) in the Irish Sea. ICES J. Mar. Sci. 2008, 65, 1670–1675. [Google Scholar] [CrossRef] [Green Version]
- Vignon, M. Short-term stress for long-lasting otolith morphology-brief embryological stress disturbance can reorient otolith ontogenetic trajectory. Can. J. Fish. Aquat. Sci. 2018, 75, 1713–1722. [Google Scholar] [CrossRef]




{kind=link}
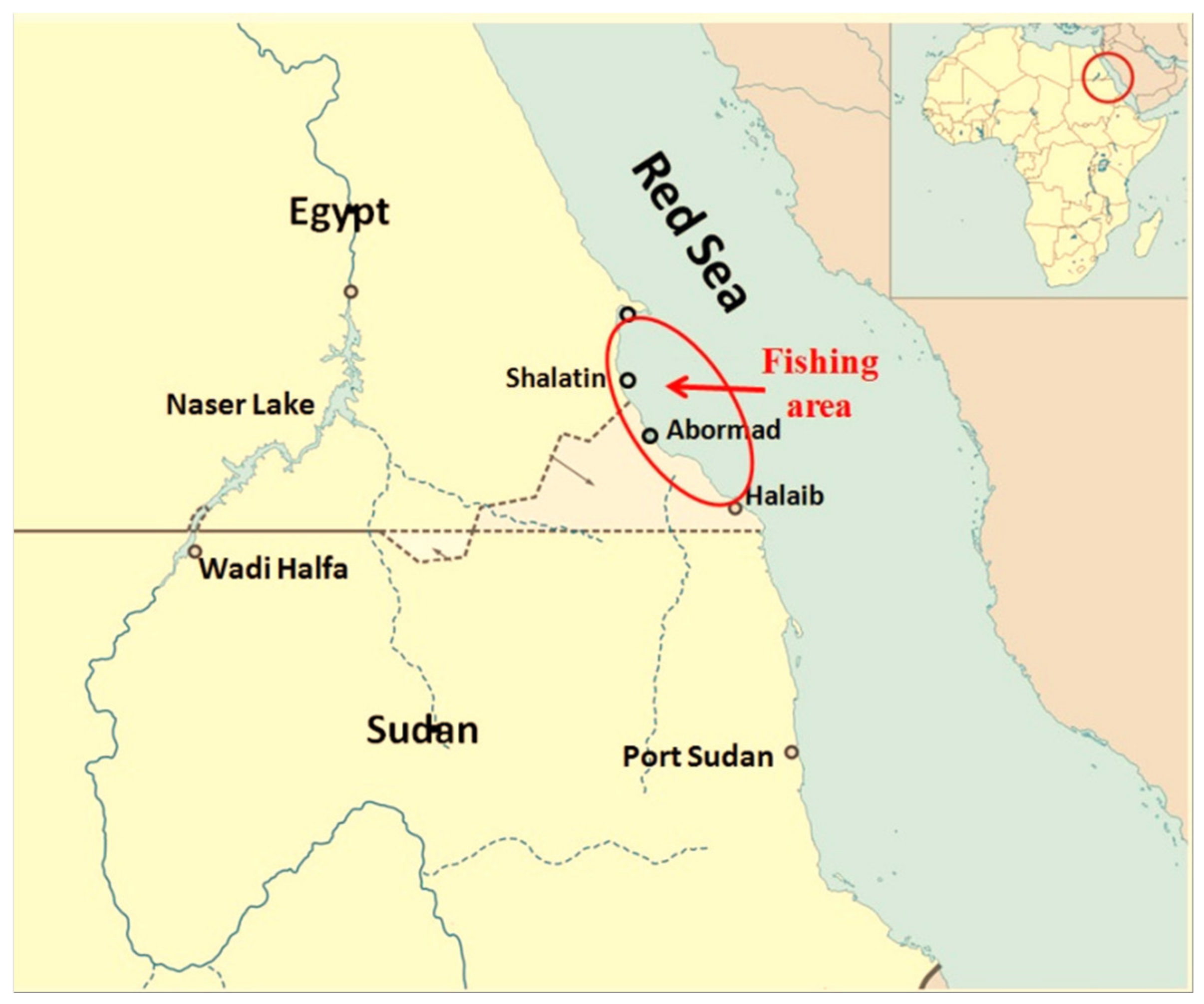{kind=link}
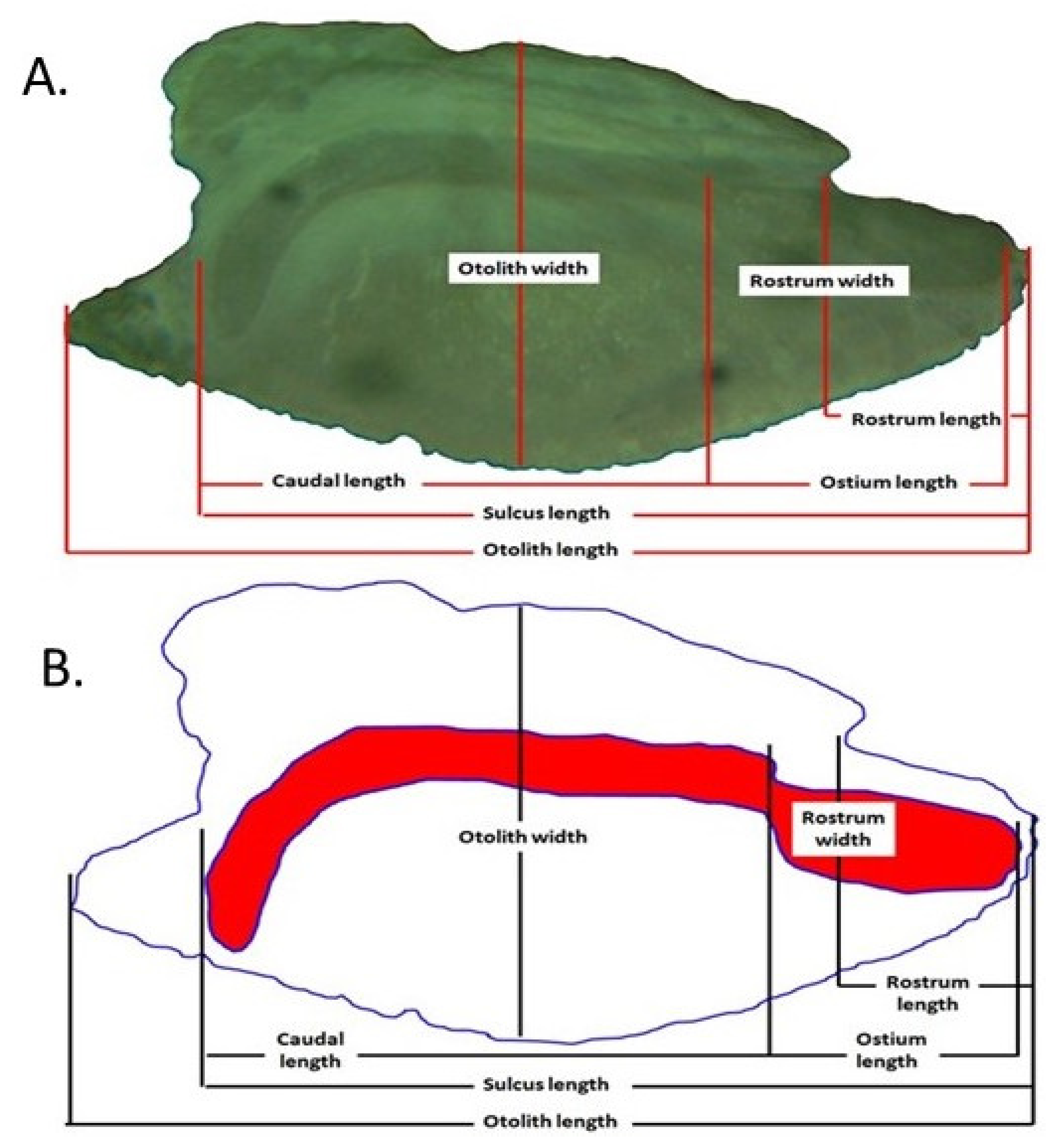{kind=link}
{kind=link}
| Size Parameters | Shape Indices | |
|---|---|---|
| Length: OL | Circularity: | CI = OP/OA2 |
| Width: OH | Ellipticity: | EL = (OL − OH)/(OL + OH) |
| Perimeter: OP | Roundness: | RO = 4 OA/πOL2 |
| Area: OA | Aspect ratio: | AR = OL/OH |
| Weight: OW | Rectangularity: | RE = OA/(OL · OH) |
| Sulcus: OSL | Form-factor: | FF = 4 πOA/OP2 |
| Ostium: OOs | Compactness: | CO = OP2/OA |
| Cauda: OCu | Squareness: | SQ = OA/(OL · OW) |
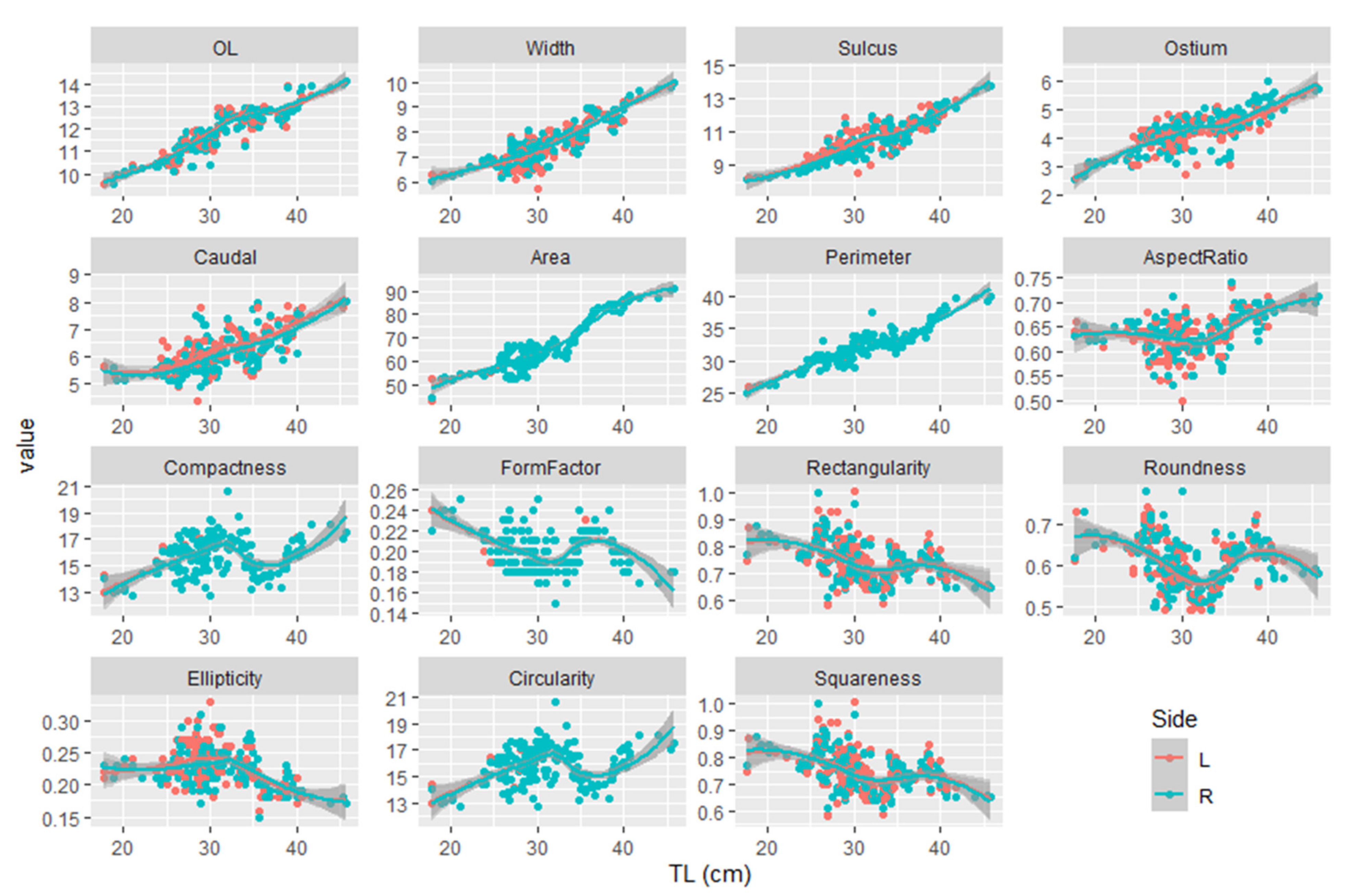
| Otolith Descriptor | TL | Sex Effect | Side Effect | Allometric Equations between Otolith Descriptor and TL |
|---|---|---|---|---|
| OL | 0.482 | 0.306 | 0.010 | Left side OL = 2.833 × TL0.418 |
| Right side OL = 2.631 × TL0.439 | ||||
| OH | 0.274 | 0.791 | 0.148 | OH = 1.091 × TL0.563 |
| Sulcus length | 0.078 | 0.534 | 0.336 | SU = 1.549 × TL0.554 |
| Ostium length | 0.008 | 0.823 | 0.621 | OS = 0.441 × TL0.656 |
| Caudal length | <0.001 | 0.201 | <0.001 | Left side CA = 1.154 × TL0.491 |
| Right side CA = 1.119 × TL0.454 | ||||
| OA | < 0.001 | 0.133 | 0.446 | OA = 4.625 × TL0.777 |
| OP | 0.007 | 0.114 | 0.986 | OP = 7.104 × TL0.441 |
| AR | 0.973 | 0.325 | 0.853 | AR = 0.400 × TL0.135 |
| C | 0.023 | 0.402 | 0.703 | C = 10.909 × TL0.105 |
| FF | 0.001 | 0.345 | 0.388 | FF = 0.288 × TL−0.105 |
| RE | 0.301 | 0.413 | 0.642 | RE = 1.552 × TL−0.215 |
| RO | 0.523 | 0.222 | 0.263 | RO = 0.790 × TL−0.08 |
| EL | 0.574 | 0.289 | 0.091 | EL = 0.638 × TL−0.308 |
| CI | <0.001 | 0.402 | 0.802 | CI = 10.909 × TL−0.105 |
| SQ | <0.001 | 0.413 | <0.001 | Left side SQ = 1.532 × TL−0.211 |
| Right side SQ = 1.573 × TL−0.219 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osman, Y.A.A.; Mahé, K.; El-Mahdy, S.M.; Mohammad, A.S.; Mehanna, S.F. Relationship between Body and Otolith Morphological Characteristics of Sabre Squirrelfish (Sargocentron spiniferum) from the Southern Red Sea: Difference between Right and Left Otoliths. Oceans 2021, 2, 624-633. https://doi.org/10.3390/oceans2030035
Osman YAA, Mahé K, El-Mahdy SM, Mohammad AS, Mehanna SF. Relationship between Body and Otolith Morphological Characteristics of Sabre Squirrelfish (Sargocentron spiniferum) from the Southern Red Sea: Difference between Right and Left Otoliths. Oceans. 2021; 2(3):624-633. https://doi.org/10.3390/oceans2030035
Chicago/Turabian StyleOsman, Yassein A. A., Kélig Mahé, Samia M. El-Mahdy, Ashraf S. Mohammad, and Sahar F. Mehanna. 2021. "Relationship between Body and Otolith Morphological Characteristics of Sabre Squirrelfish (Sargocentron spiniferum) from the Southern Red Sea: Difference between Right and Left Otoliths" Oceans 2, no. 3: 624-633. https://doi.org/10.3390/oceans2030035