1. Introduction
The marine environment has been substantially altered by human activities, with consequent adverse effects on wildlife, including cetaceans. Cetaceans are long-lived, near the top of the food chain, and are sentinels to oceanic change [
1]. Monitoring them is, therefore, important. Mortalities, pathology, and disease are at the core of a comprehensive monitoring program, and there has been an increased effort in these fields since about 1990 [
2]. When studying the pathology of wild cetaceans, researchers have usually been limited to opportunistically collected, beach-washed carcasses [
3,
4] and sometimes specimens from fisheries bycatch [
5,
6,
7,
8]. Both data sources have limitations and benefits: cetaceans obtained from fisheries are likely to be a more ‘natural’ part of the population because they have died as a result of human activities, not disease, and are usually in a fresh condition. Beach-washed animals, on the other hand, tend to be more decomposed and often die as a result of disease [
9].
Studies on the mortality and pathology of stranded cetaceans are available for many parts of the world [
3,
9,
10,
11]. Some have reported on mass strandings [
11] and anthropogenically mediated mortalities [
3,
9,
12,
13]. Pathological changes in stranded cetaceans have been the subject of many papers. For example, bacteria and fungi of marine mammals were reviewed by Higgins [
14] and prominent emerging infectious diseases worldwide were reviewed by Van Bressem et al. [
15]. The latter concluded that inshore and estuarine cetaceans were more at risk than those living offshore.
Monitoring cetacean mortalities in Australia has primarily been carried out on a state-by-state basis, with most studies documenting species and distribution, rather than cause of death and the associated pathological changes [
16,
17]. Within Australia, South Australia (SA) has the most comprehensive stranding record, and this includes information on the circumstances associated with events [
16,
18,
19]. Separating the ‘cause’ from the ‘circumstance’ of death was an important advancement in documenting cetacean events in SA [
19] because it distinguished the primary cause of death (e.g., exsanguination/aspiration) from the circumstances surrounding the event (e.g., entanglement). In the SA classification, pathology results obtained during necropsy were combined with observations by collectors and others present at an event, thus providing a thorough picture of a stranding. Publications describing injuries and pathological changes related to human-induced marine mammal mortalities [
20,
21] provided an important baseline for assignment to anthropogenic circumstances. Knowing the circumstance of death for cetaceans is critical for conserving species and populations, because management agencies require such data to monitor trends in mortality. This is especially true for mitigating anthropogenically mediated mortalities, such as entanglements and illegal killings. For example, in a previous study, 42% of cetacean mortalities with known circumstances of death in SA were classified as anthropogenic [
16].
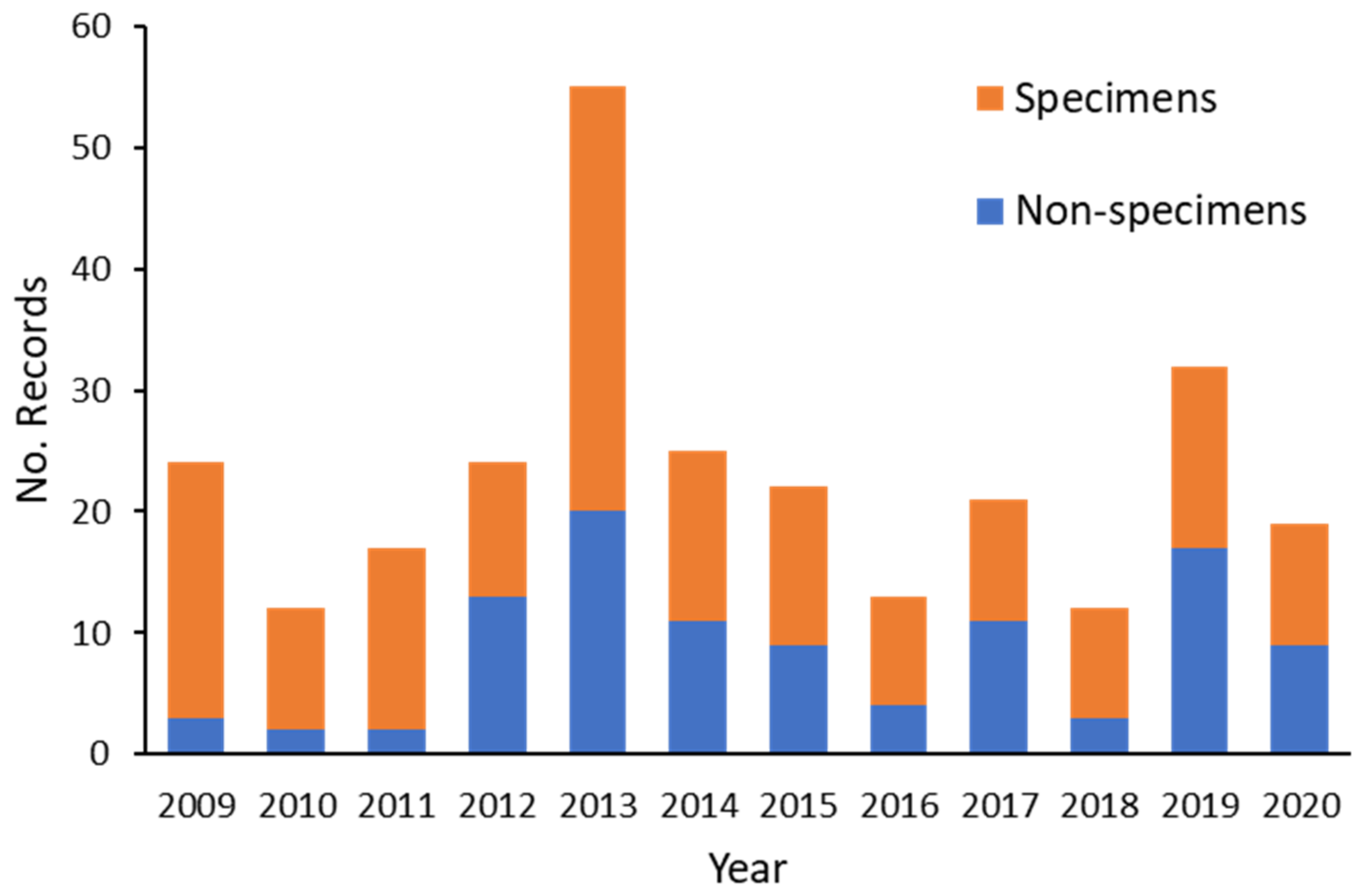
Documenting cetacean strandings and mortalities in SA has been greatly facilitated by the setting up of an intense program of necropsy and state-wide reporting established in about 1990. This yielded data for hundreds of cetaceans (primarily dolphins), including those inhabiting the St Vincent Gulf Bioregion (SVG). A series of publications on the pathology of dolphins in SA are available, including lung nematodes associated with severe pathological lesions [
22], types and prevalence of skeletal pathologies [
23], and forensic investigations [
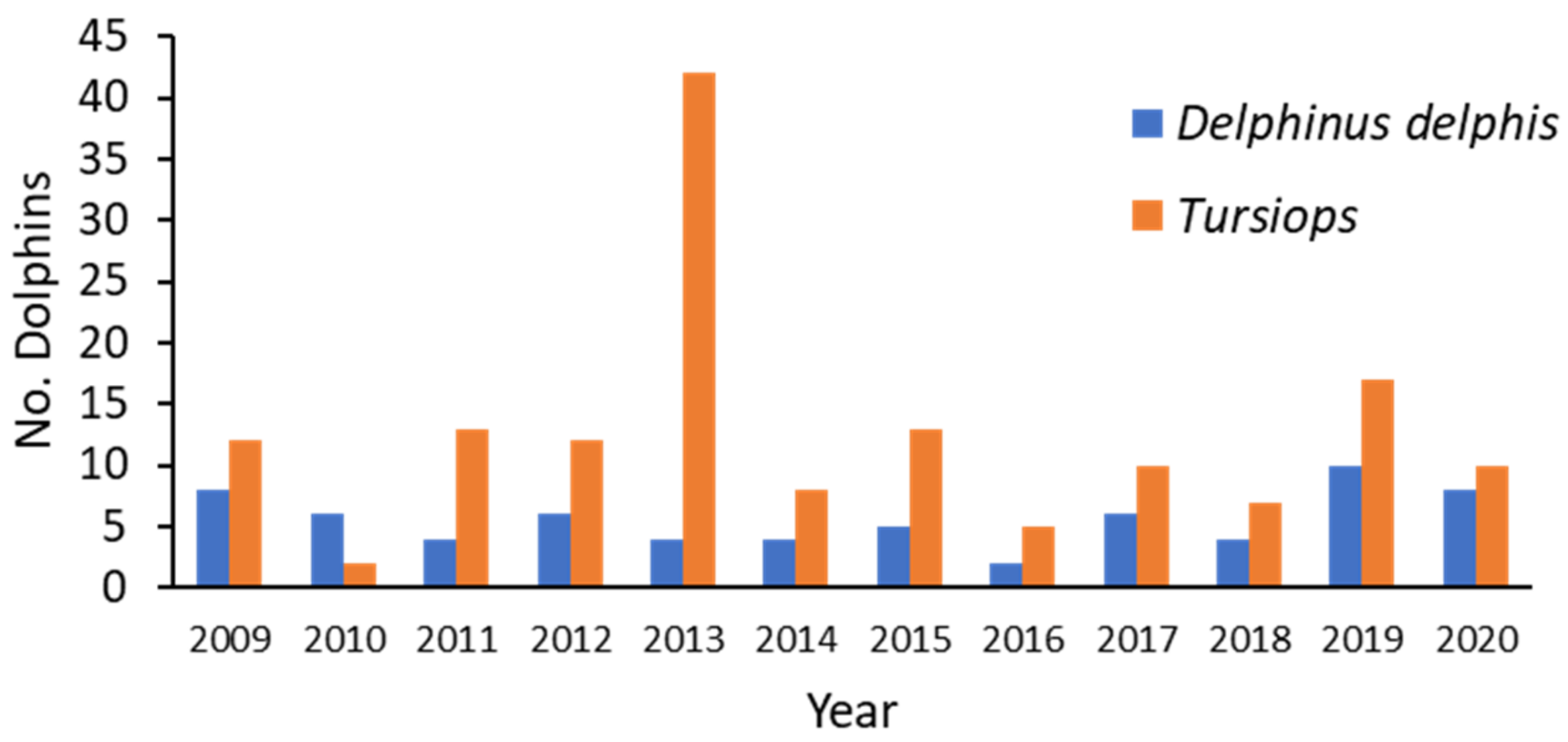
24]. An unusual mortality event (UME) occurred in the SVG during 2013, which primarily involved
Tursiops aduncus (Indo-Pacific bottlenose dolphin) and cetacean morbillivirus [
25].
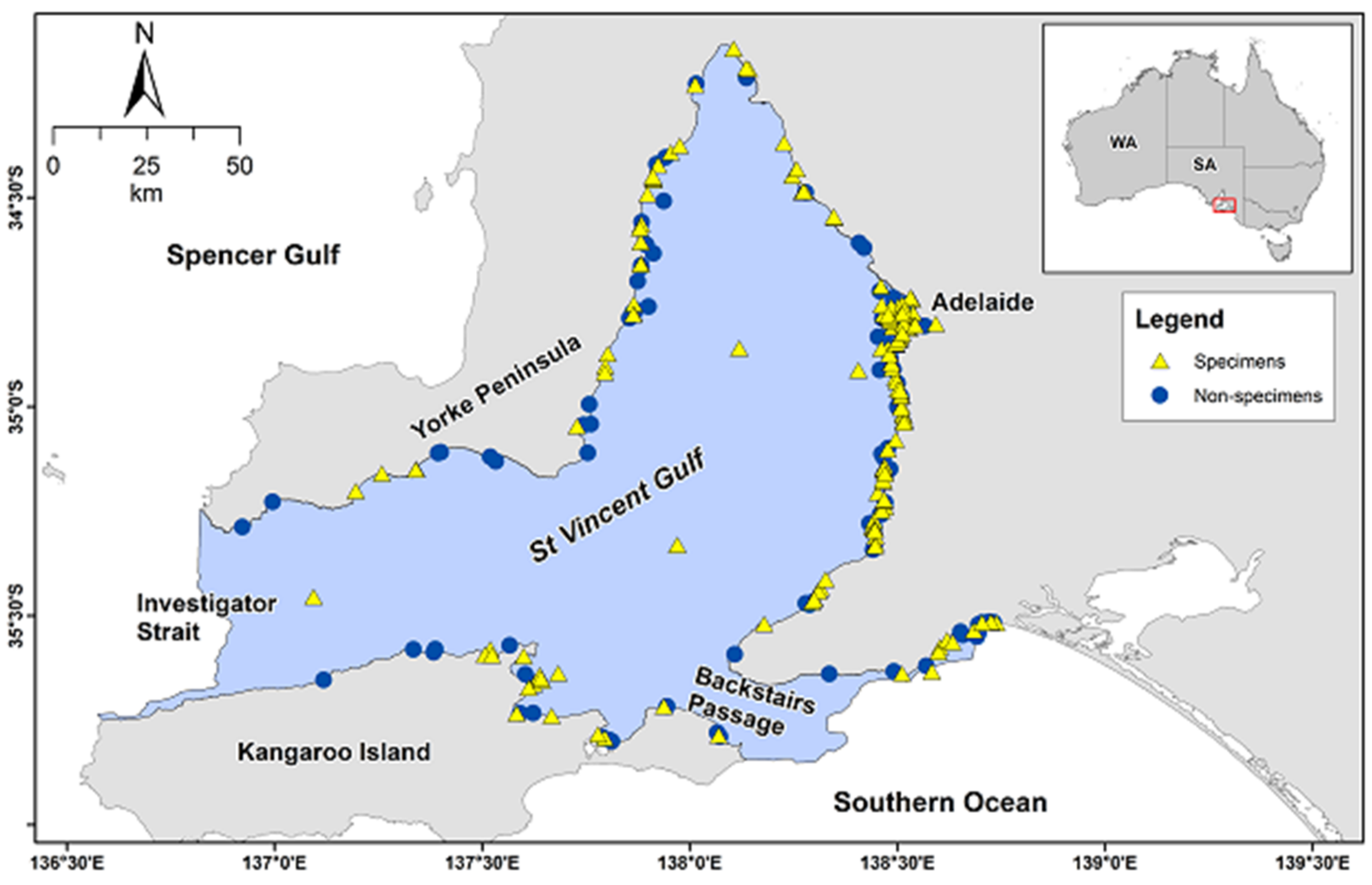
The SVG is of special interest with regard to cetacean mortalities, because it is a semi-enclosed inverse estuary [
26] with restricted flushing, and the largest city in the State is located on its eastern shore (
Figure 1). These characteristics may lead to high levels of toxic contaminants in resident dolphins [
27]. There is a potential flow-on effect on the health of dolphins, particularly the Indo-Pacific bottlenose dolphin, because it feeds on benthic organisms [
28], which accumulate toxins. As a result of concerns for the conservation of this species living in an urban environment, the Adelaide Dolphin Sanctuary (ADS) was created in the Adelaide metropolitan area in 2005. A study on dolphin mortalities in ADS [
29] during 1987–2013 found that disease was the most frequently recorded circumstance of death. Increased funding was made available in 2009 to intensify the study of cetacean mortalities in the SVG, including the ADS.
The aim of this study was to identify trends in the mortalities of SVG cetaceans, including life history and pathology, and compare these with previous studies conducted in SA and elsewhere. Emphasis was placed on dolphins because they made up a large proportion of the records. All records of strandings and other events were included, although necropsied carcasses yielded the most comprehensive data. Major trends in pathology and the circumstance of death of dolphins were documented, including infectious diseases and pathogens associated with them, as well as non-infectious diseases.
4. Discussion
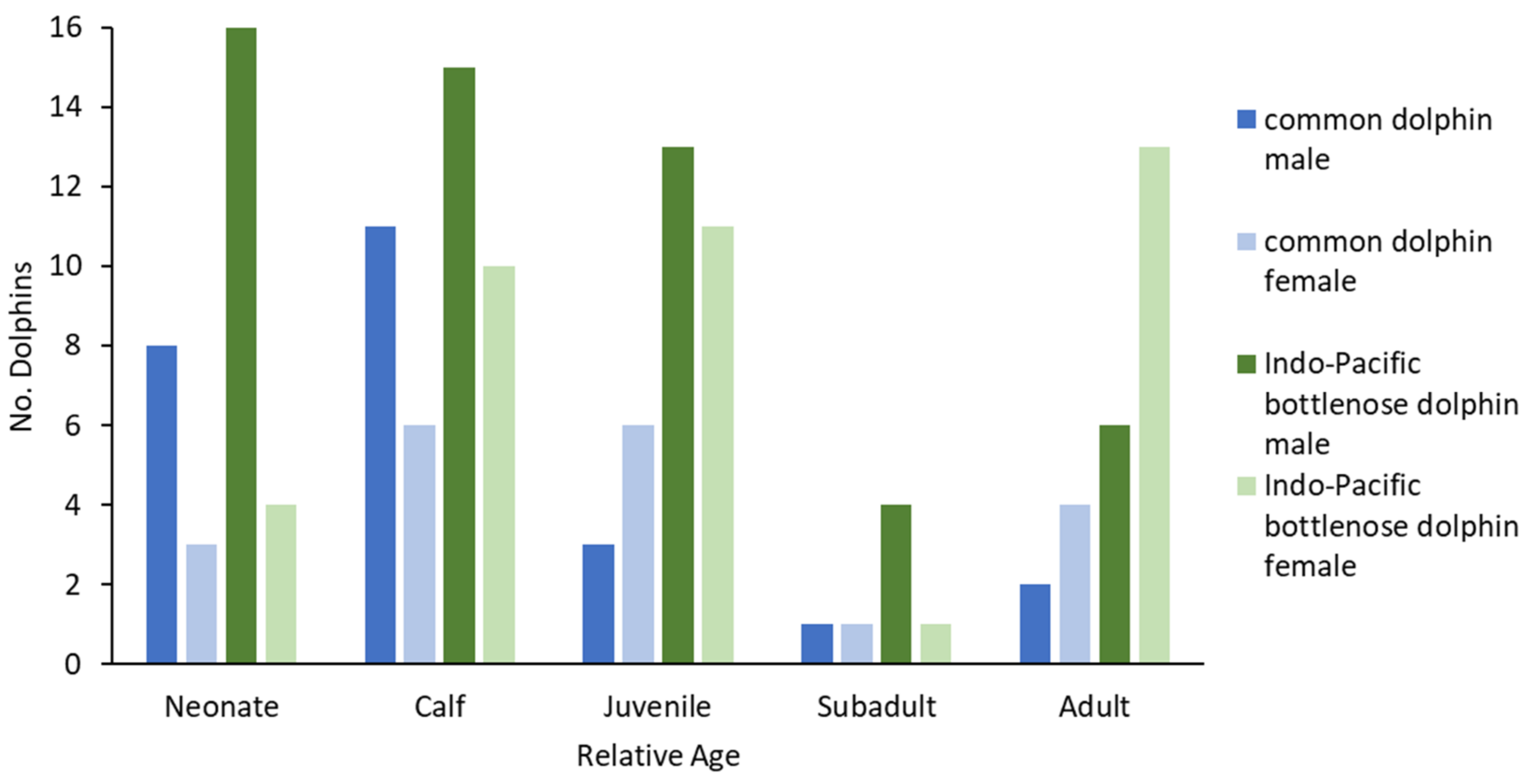
One of the unique aspects of the SA studies of stranded cetaceans, including that in the SVG during 2009–2020, is that specimens have been collected over a long period of time and archived for further investigation. This is especially important for assessing the maturity of individuals using skeletons and reproductive organs [
35]. In addition, a broad range of biological and pathological data were collected. The study also made a significant contribution to the pathogens identified in Australian cetaceans. Although not specifically referred to as ‘circumstances of death’, many studies have reported on similar categories, such as disease, live stranding, neonatal death, mother–calf separation, malnutrition, predation, and anthropogenic cases [
3,
9,
12,
13,
36]. The categories established in SA have been in use for over 25 years, and it is hoped that this system will set a benchmark for other Australian studies.
Human-induced mortality and injury of cetaceans have been the focus of many studies worldwide, because such data are required by wildlife management agencies. The highest rate of anthropogenic cases (65%) was recorded in Brazil [
10] and the lowest (10%) at Cape Cod, USA [
11]. In the Canary Islands, Spain, there was an apparent reduction in anthropogenic mortalities between two studies of six (33%) and seven (19%) years [
3,
9]. The proportion of anthropogenic cases in SA (whole State) was 42% [
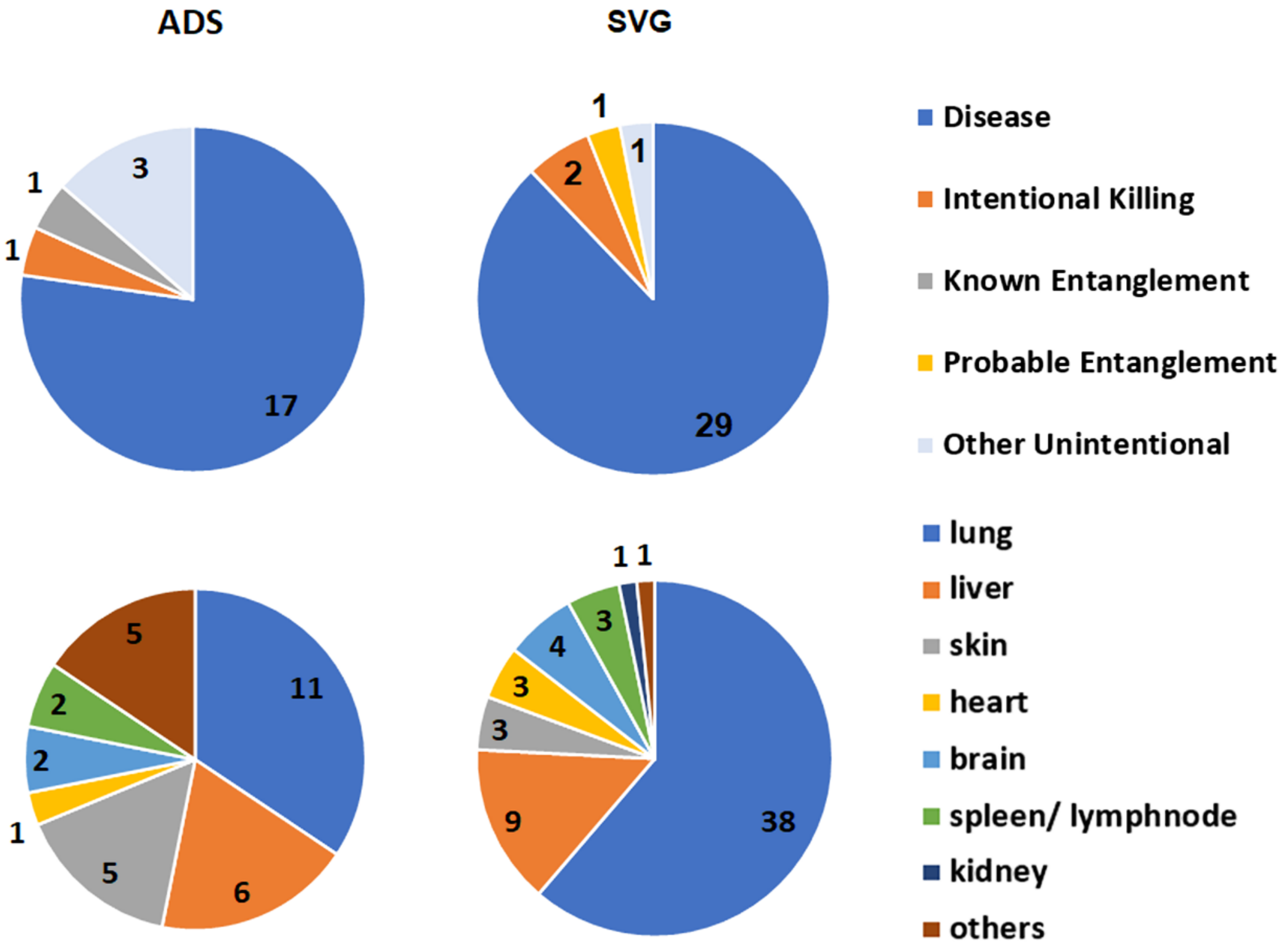
16], and that in the present study of the SVG was 21%
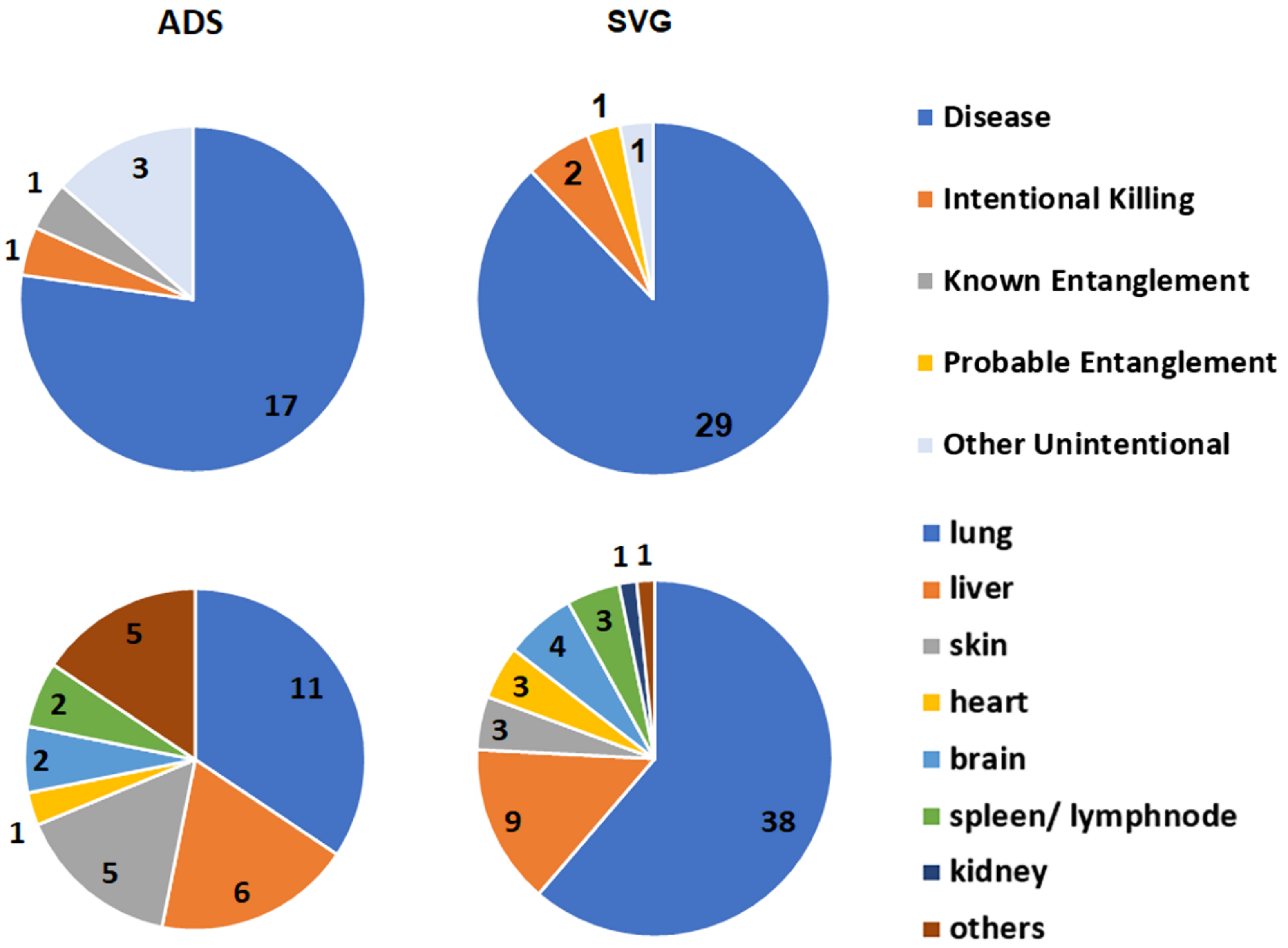
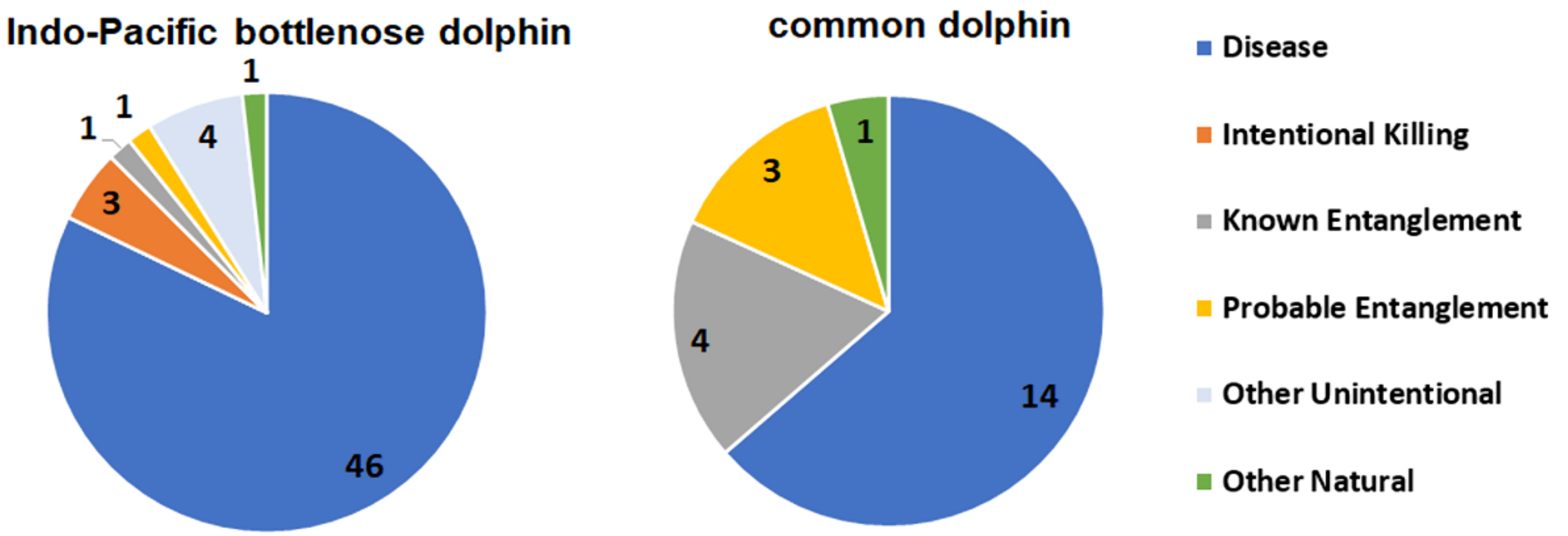
Anthropogenic cases of mortality and injury in cetaceans include intentional (e.g., shooting) and unintentional mortalities (e.g., vessel collision, entanglement/entrapment, and foreign bodies in the digestive tract) [
3,
4,
9,
20]. Worldwide, entanglement remains one of the major causes of anthropogenic mortality for cetaceans [
3,
20,
37,
38]. In Australia, there has been an increase in entanglements, primarily of odontocetes, since 2000, and populations of inshore small cetaceans are believed to be at risk of decline [
39]. South Australia follows the same pattern of high rates of entanglement in fishing gear [
16,
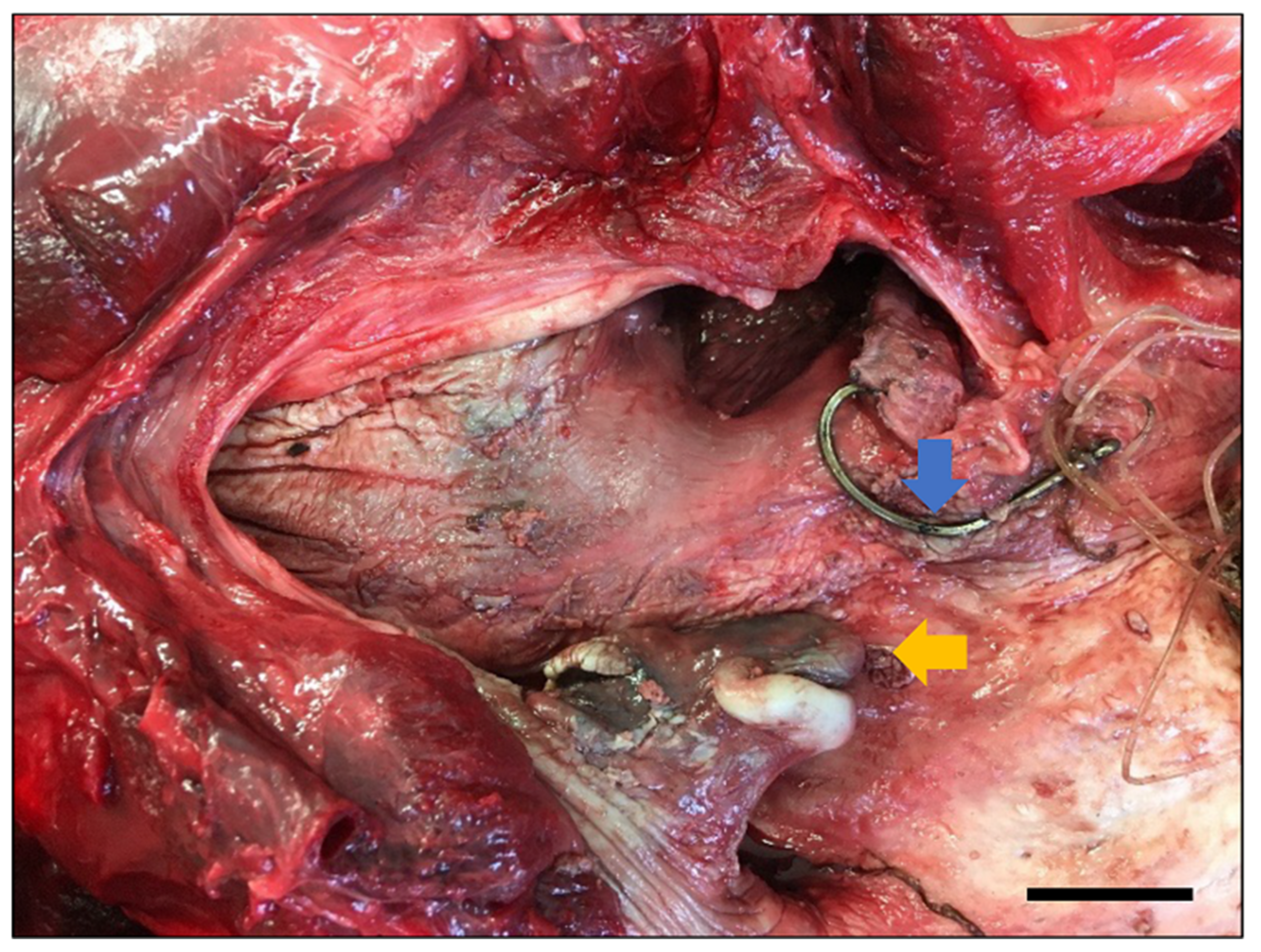
19], with 13.4% of records in this category in the SVG study. Compared with Indo-Pacific bottlenose dolphins, common dolphins had a higher proportion of fatal entanglements (6% vs. 32%), and of these, Known Entanglements were collected only from the SA Sardine Fishery. Probable Entanglements of common dolphins recorded in the SVG study were likely to be associated with this fishery, since it is the primary net fishery in the gulf. Of the dolphins that died, Known Entanglements of Indo-Pacific bottlenose dolphins were related only to recreational fishing gear in the ADS (
Figure A4), as has been reported by Byard et al. [
25].
Intentional Killing of SA dolphins has been concerning in the past [
16,
19]. No cases were recorded in the ADS during 2005–2013 [
29]. In the SVG study, three Indo-Pacific bottlenose dolphin mortalities were recorded with shotgun pellets along the Adelaide metropolitan coast during 2009–2020.
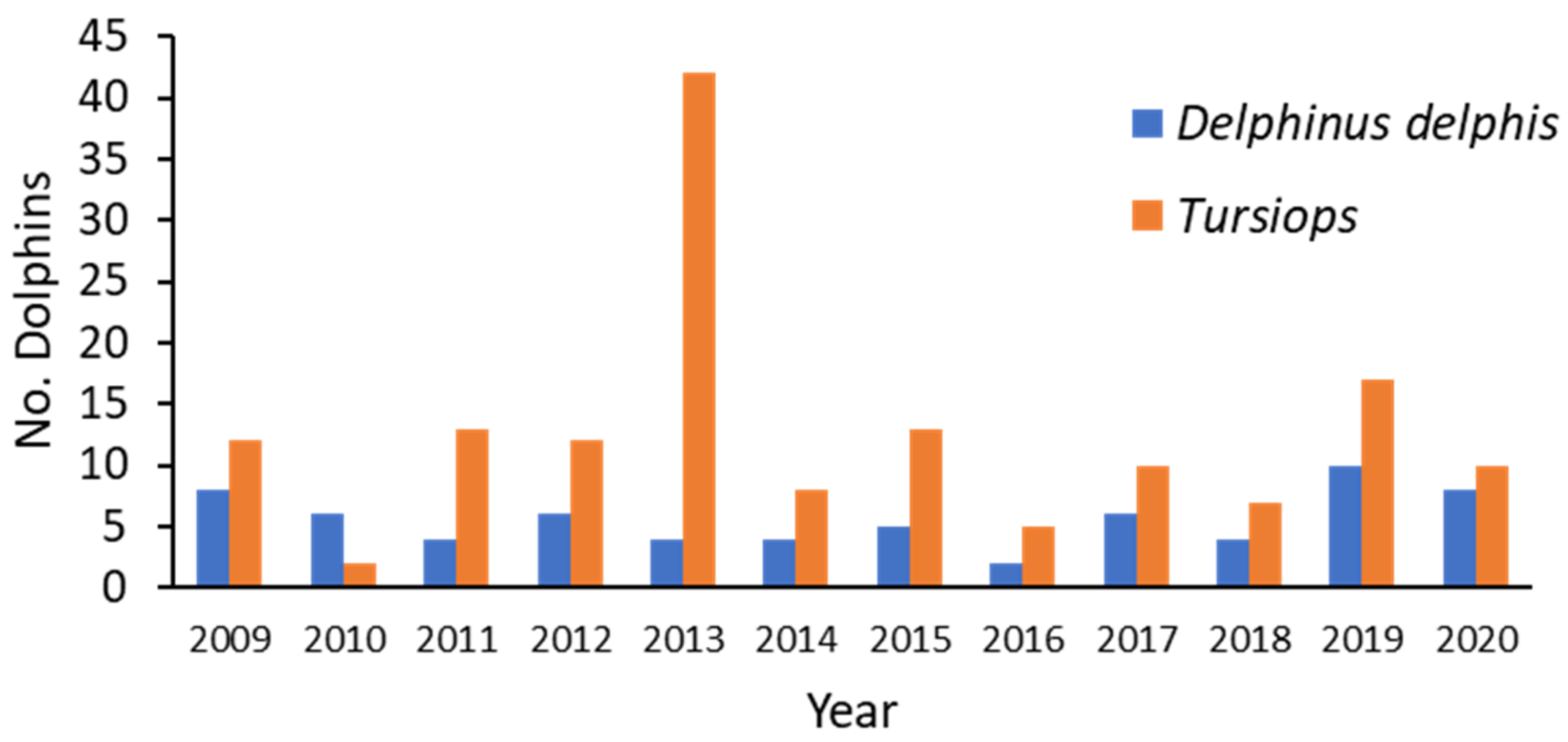
When the circumstance of death for the 2009–2020 SVG study was compared with that of the previous period (1990–2008), the only clear difference in proportion was for Disease, where it increased from 38% in the first period to 73% in the second (
Table A3(a), (b)). The reasons for such a disparity are unlikely to be related to an increase in disease per se, but probably to the effort applied in the more recent study. From 2009, increased funding enabled more histological preparations and diagnostic testing for microorganisms. In addition, consistent specialized evaluation of disease was provided by the lead author (IT). Anthropogenic cases remained proportionately the same when the time periods were compared for SVG (22 and 21%).
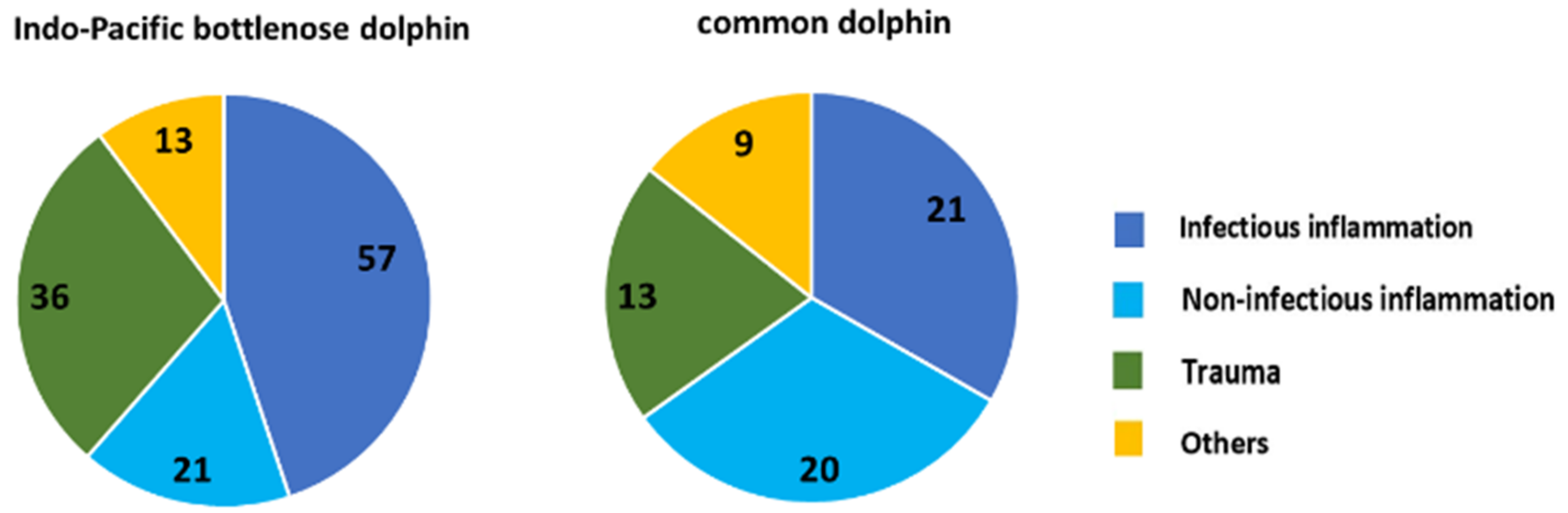
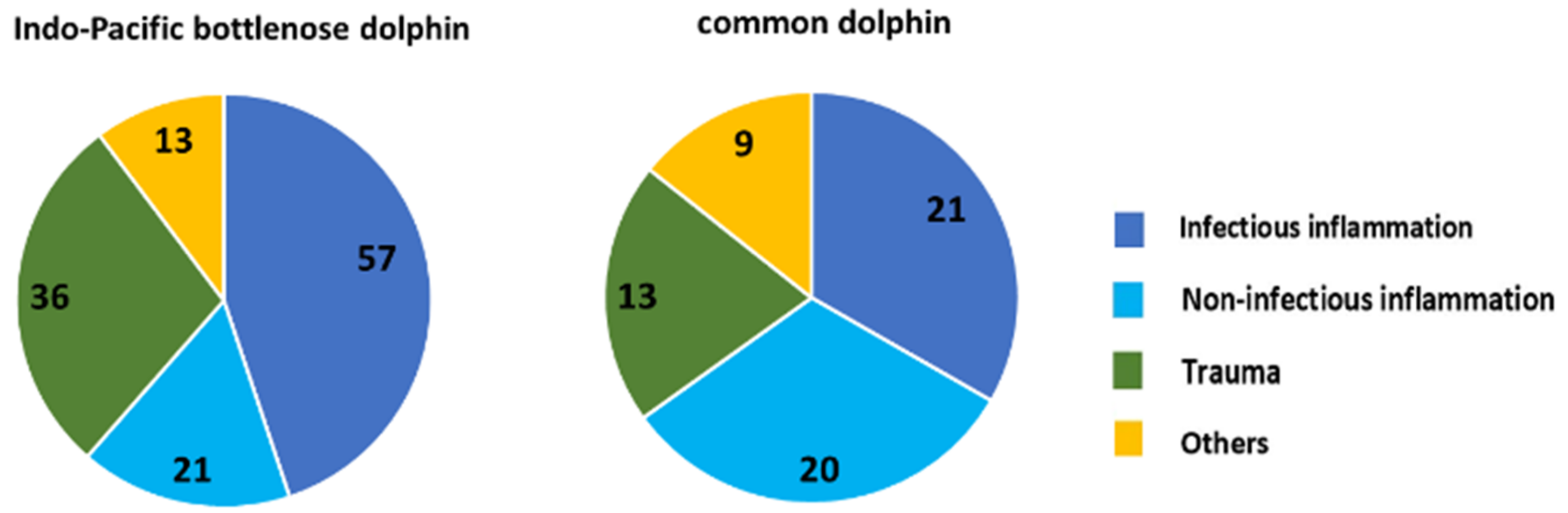
Infectious disease, particularly pneumonia, has been the most common pathological lesion reported for stranded cetaceans [
3,
10,
40,
41], as was the case in the SVG study. The etiology of pneumonia includes viruses, bacteria, fungi, and parasites (nematodes) with prevalence varying according to the host species, geographic distribution of the pathogen, and environmental factors [
42]. An additional compounding factor is whether the pathogen is primary or secondary [
41]. To date, there has been no broad-ranging study of infectious disease in Australian cetaceans [
2]. The SVG study found that the patterns of infection in Indo-Pacific bottlenose dolphins varied between the enclosed and altered habitat of the ADS and the greater gulf. In the ADS, infections included many organs (mainly the lungs, liver, and skin), while in other parts of the SVG, they were mostly in the lungs.
Worldwide, emerging infectious diseases of cetaceans include morbillivirus and
Brucella [
15], both of which are known to contribute to high mortality of cetaceans [
43,
44]. Morbillivirus has been recorded on several occasions in Australian cetaceans, including UMEs [
25]. It is often associated with pneumonia and lesions that are secondary bacterial infections [
45,
46]. Bronchopneumonia has been reported in morbillivirus cases of
Tursiops in SA [
22], Western Australia (WA) [
47], and Queensland [
48]. Secondary infections involving bacteria and fungi were also reported in SA and WA. Morbilliviruses can also cause immunosuppression and can lead to opportunistic bacterial infections in dolphins [
47,
49,
50]. Lymphoid depletion was often reported in dolphins with morbillivirus infection in the lymph organs [
50]. The increase in parasitic infections and skin lesions seen in 2013 seemed to be due, in part, to an immunocompromised state as a result of lymphoid depletion.
Brucellosis is a zoonotic disease [
51] and a potential human health threat.
Brucella spp. was first isolated from small cetaceans, including the common dolphin and common bottlenose dolphin in the UK [
52] and USA [
53], and subsequently many other world regions. Although
Brucella has not been identified in wild dolphins in Australia, although it has been reported in captive common bottlenose dolphins from Queensland [
54]. There are no published cases of identified
Brucella in SA, but skeletal lesions suggestive of
Brucella infection have been found in Indo-Pacific bottlenose dolphins from both gulfs [
23], although this has not been confirmed. Guzman-Verri et al. [
55] also reported skeletal lesions consistent with
Brucella infection. There was no sign of brucellosis in the visceral organs of the cetaceans examined in the present study, but a serological survey is being conducted by the University of Adelaide (pers. comm. A.L. Chaber and L. Woolford, May 2022) to confirm its presence or absence in the state.
Marine mammal bacteria include pathogenic and non-pathogenic types [
14]. The SVG study identified 32 species of pathogenic bacteria, including eight Indo-Pacific bottlenose dolphins with
Vibrio parahaemolyticus and
V. alginolyticus associated with necro-suppurative pneumonia and ulcerative dermatitis. These pathogens have been reported from Italy, where they were associated with meningoencephalitis [
56]. The SVG study also identified
Staphylococcus aureus in the Indo-Pacific bottlenose dolphin (n = 3), and these infections were associated with pneumonia. A study of the common bottlenose dolphin from the west coast of the USA concluded that
Staphylococcus aureus was a significant risk to mortality [
57]. There have been few reports of bacterial species recorded from Australian cetaceans [
58,
59]. Of note is a case of
Streptococcus iniae in a dead common dolphin from the metropolitan coast of Adelaide, which also had sepsis [
60]. This bacterium was identified in the SVG study and was considered the primary cause of death. The total list of pathogenic bacteria recorded in SA now stands at 33 species, including
Corynebacterium ulcerans [
19].
Toxoplasma gondii is a zoonotic organism that is infectious to almost all mammals and birds, with infection rates in humans varying according to country, region, and age. Although the final host is the cat, infection of dolphins [
61] indicates that the marine environment has been impacted.
Toxoplasma gondii infection can be fatal in cetaceans [
61] and has been reported from many world regions [
62,
63,
64]. In Australian cetaceans, this protozoan parasite has been identified in several species of dolphins from WA, Queensland, and New South Wales [
65,
66,
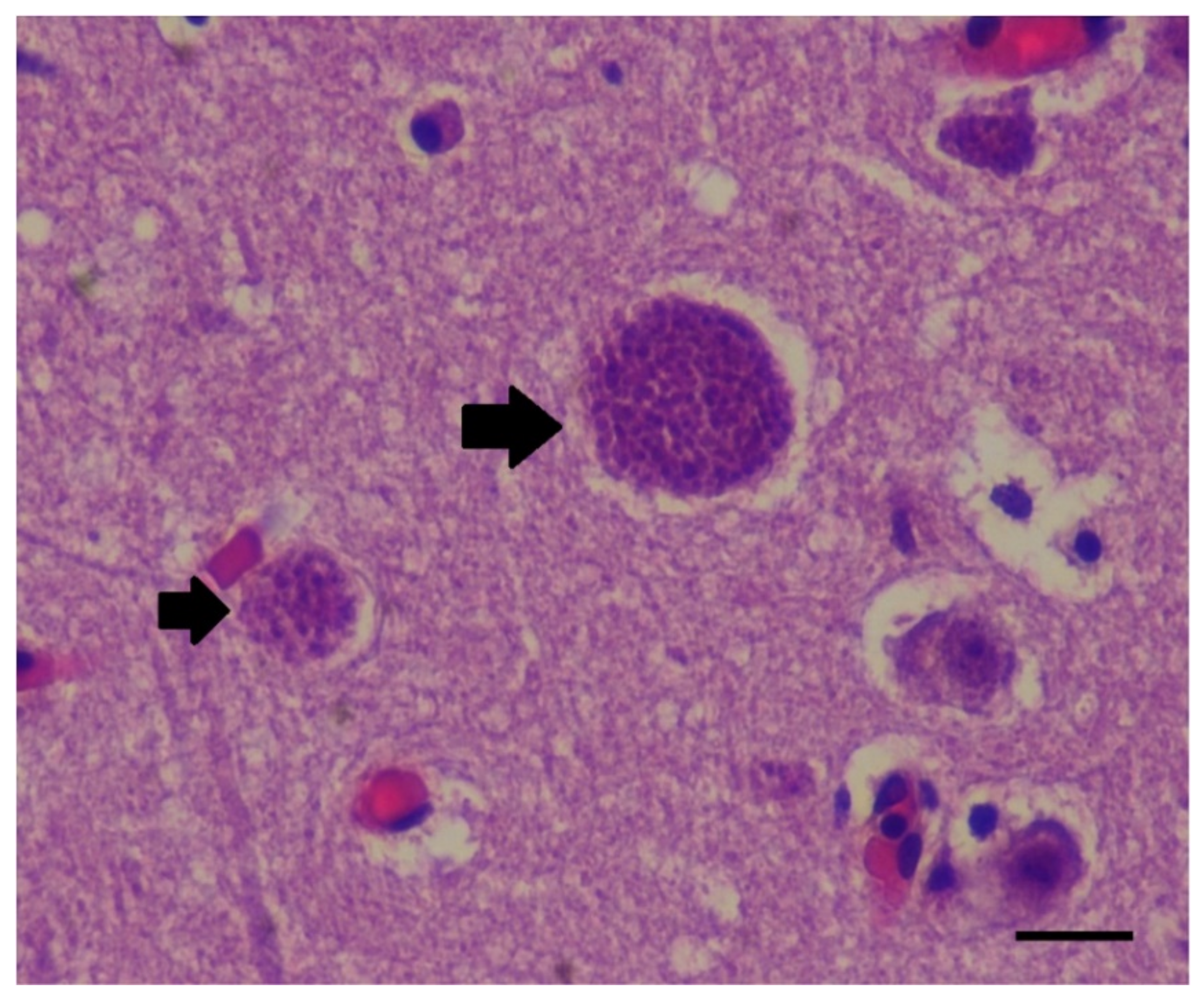
67]. Toxoplasma cysts were morphologically detected and immuno-histologically confirmed in the brain of a dolphin co-infected with morbillivirus during the SA 2013 UME. A serological investigation is underway for
Toxoplasma gondii in Indo-Pacific bottlenose dolphins in SA (pers. comm. R. O’Handley April 2022).
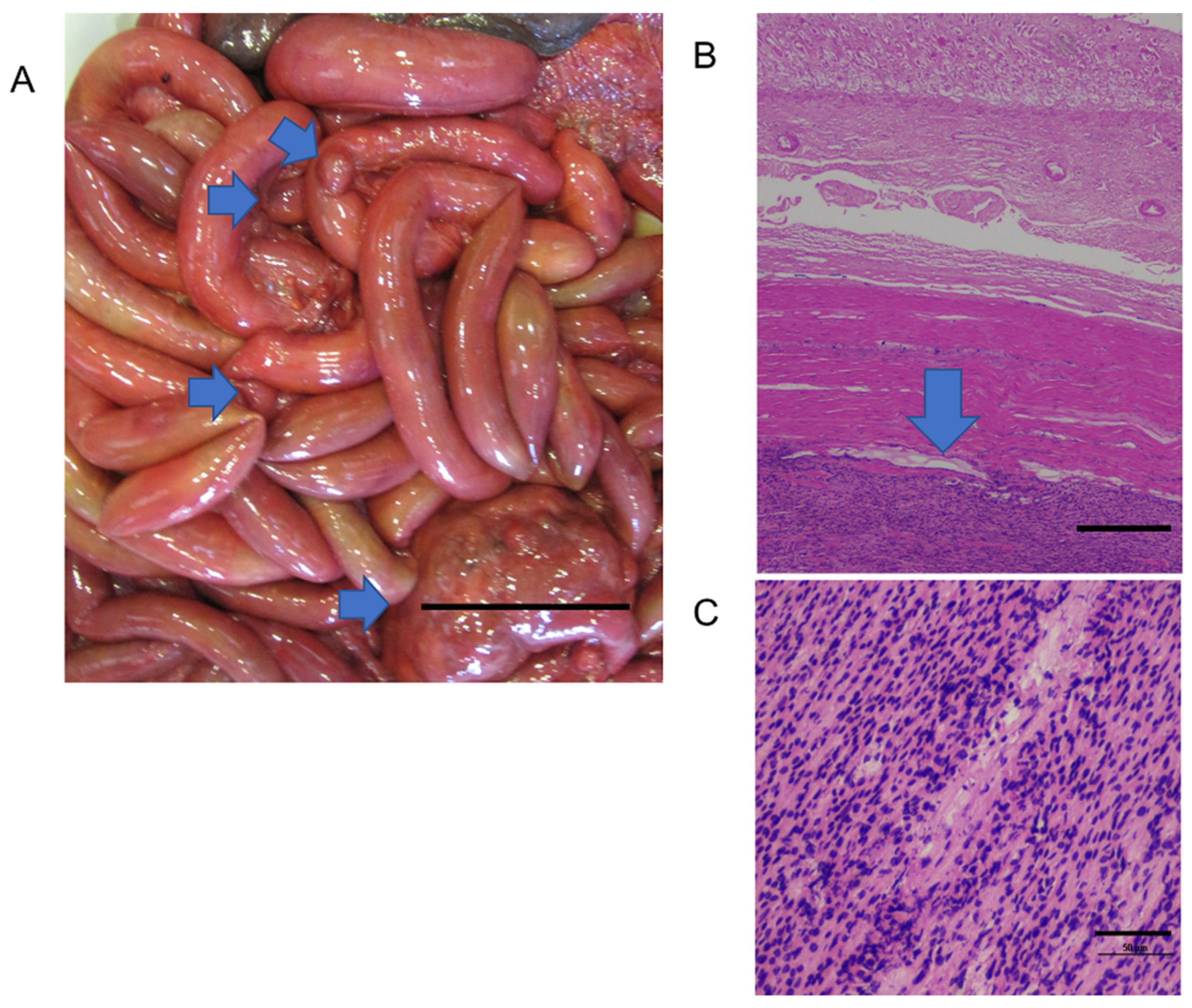
Lung nematode infections have been reported in cetaceans worldwide [
3,
10,
13,
68], including SA [
22]. These are most frequently observed in the bronchi and interstitial tissue of the lung [
10]. Trematode infections of the liver, pancreas, and brain have also been reported in SA (this study) and elsewhere [
3,
10,
44] causing inflammation in the respective tissues [
69].
Non–infectious diseases were also reported in the SVG study. Tumors are rarely reported in marine mammals [
70]. Our finding of a tumor in the gastrointestinal tract and liver of an adult common dolphin appeared to be a leiomyoma. Cetacean leiomyoma has been reported in the uterus and intestine of Atlantic white-sided dolphins (
Lagenorhynchus acutus) [
71] and in the pylorus of the common bottlenose dolphin [
72]. The tumors in the Atlantic white-sided dolphin were multicentric, which is consistent with our case, and both species showed similar histopathologic changes.
Suspected jaundice was identified in 10 SVG dolphins (5 Indo-Pacific bottlenose dolphins and 5 common dolphins), all but one of which were neonates or calves. These age groups are more likely to contract physiological jaundice [
73,
74].
Skin lesions of cetaceans have been extensively studied around the world [
75,
76,
77,
78] and are associated with bacteria, viruses, fungi, and algae [
79,
80,
81,
82]. Skin lesions of dolphins can be good indicators of overall body condition [
83] and therefore warrant monitoring. Freshwater dermatitis, associated with low salinity, has been identified in bottlenose dolphins from Gippsland, Victoria, and the Swan River, WA [
84], but this condition has not yet been observed in SA. Lesions that were recorded in the SVG study included a potential case of a lobomycosis-like skin lesion in an Indo-Pacific bottlenose dolphin from the ADS. There are no records of this type of lesion in Australian cetaceans, although it has been found in other parts of the world and can be zoonotic [
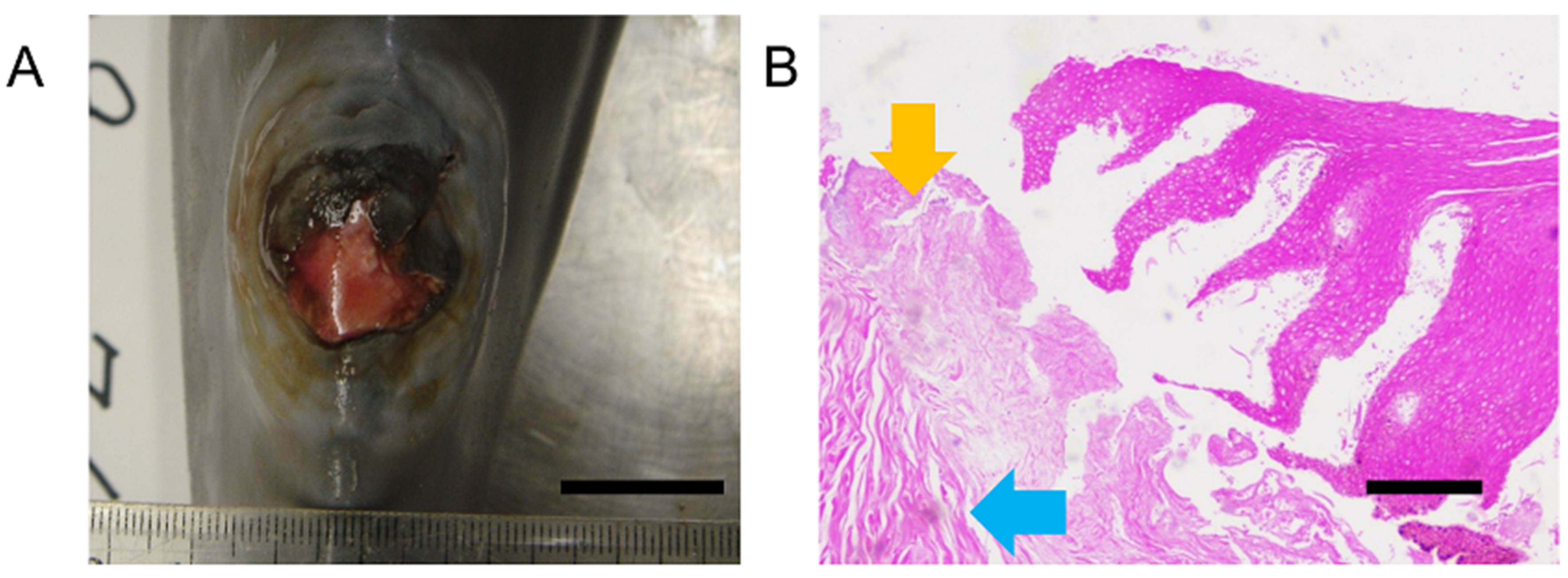
85]. Skin wounds are exposed to microorganisms that can cause infection. In addition, microorganisms found on the teeth of fish that have bitten a cetacean could be pathogenic. As the natural healing of wounds depends on the individual’s immune status, lesions often become chronic when body condition is poor.
The non-dolphin cetacean species recorded in the present study were seasonal visitors and vagrants [
32]. Although monitoring their pathology is less beneficial than that of dolphins, the data gathered will contribute to an overall understanding of their diseases and circumstances of death. The mass stranding of sperm whales in the SVG was unprecedented in modern times. This species is known from the open ocean coast of SA and is almost never recorded in the shallow gulfs [
32]. The circumstances of death for other cetaceans followed the same overall pattern as dolphins, with anthropogenic cases (vessel collision and rope entanglement) of large baleen whales recorded on several occasions. A study of the southern right whale off southern Australia documented multiple cases of entanglement and vessel collision [
86], as have been reported elsewhere [
87,
88]. The probable vessel collision of a fin whale in SVG is consistent with results worldwide reporting that this species is often killed in this manner [
89]. Many of the cases involving other cetaceans were juveniles, and of particular note was a neonatal humpback whale that was stranded alive. It is very rare for this species to give birth in SA waters [
90].
Future Research
Dolphins in the SVG are particularly vulnerable to human-induced threats because they live in a semi-closed system and the human population is likely to increase substantially, thus presenting more risk. It is, therefore, imperative to monitor mortalities in a manner consistent with the methods used in the present study and to archive specimens that can be used in future investigations. Emerging infectious diseases, such as brucellosis, toxoplasmosis, and lobomycosis, should be included in monitoring programs. Monitoring the skin lesions of live and dead dolphins will be vital to monitoring disease because it provides an early warning system for underlying health issues. Long-term, often subtle, effects on population health are difficult to detect, but monitoring toxic contaminants, immune-compromised animals, and reproductive success can be informative.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}