
Modeling the Impact of Global Warming on Ecosystem Dynamics: A Compartmental Approach to Sustainability


Abstract
1. Introduction
2. Materials and Methods
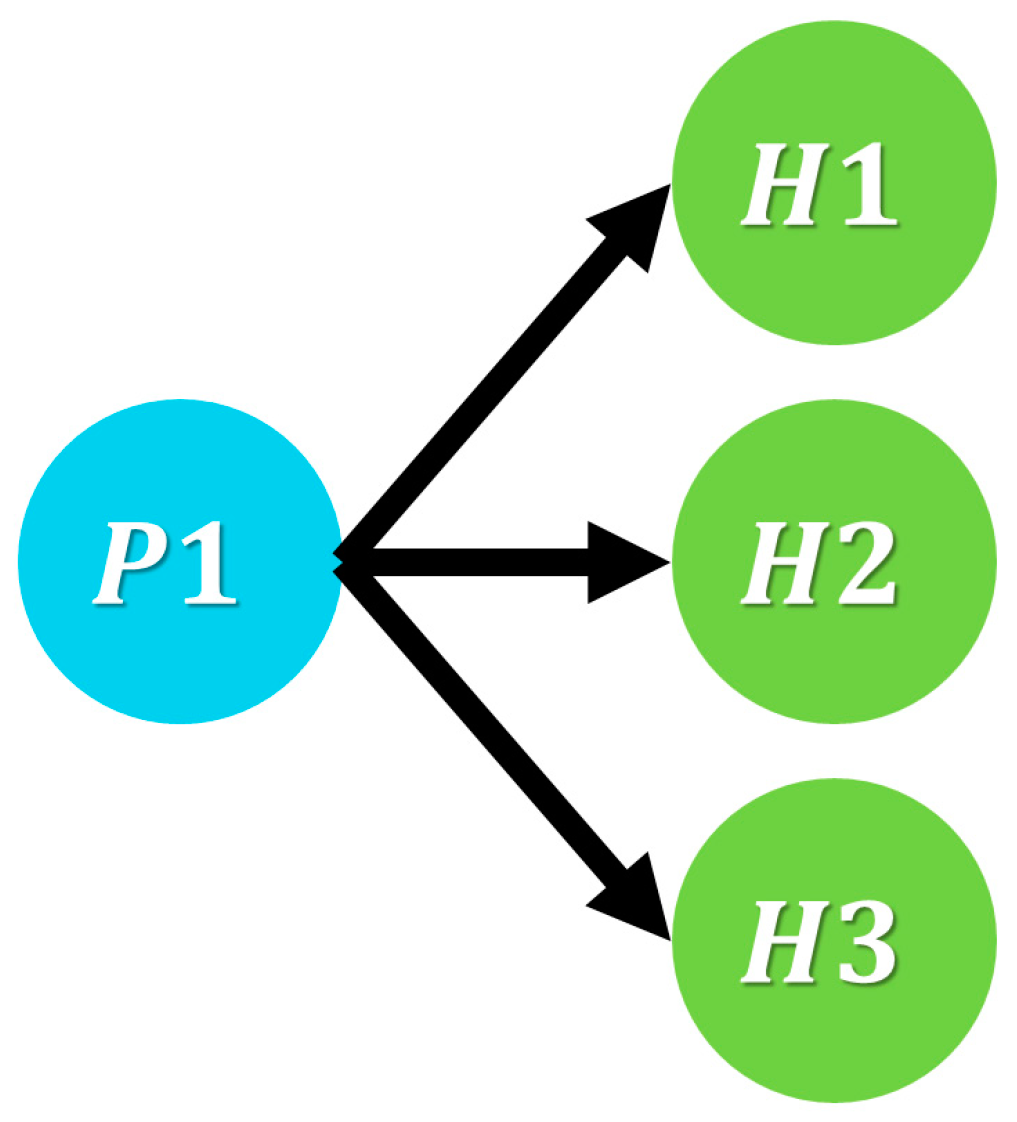
2.1. Basic Multi Predator–Prey Model
2.2. Multi-Level Predator–Prey Model
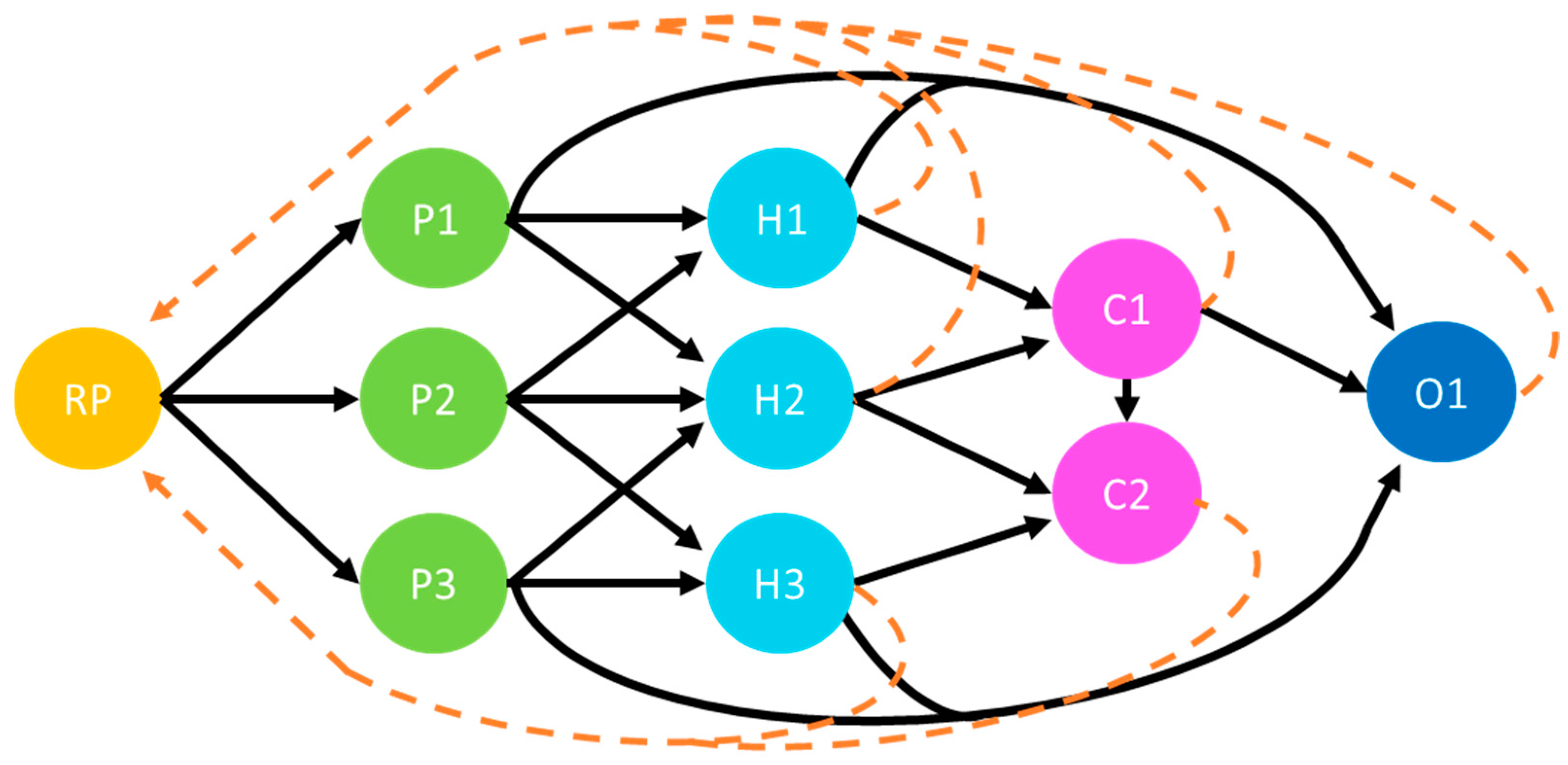
2.3. Generalized Sustainability Socio–Ecological Model (GSSEM)
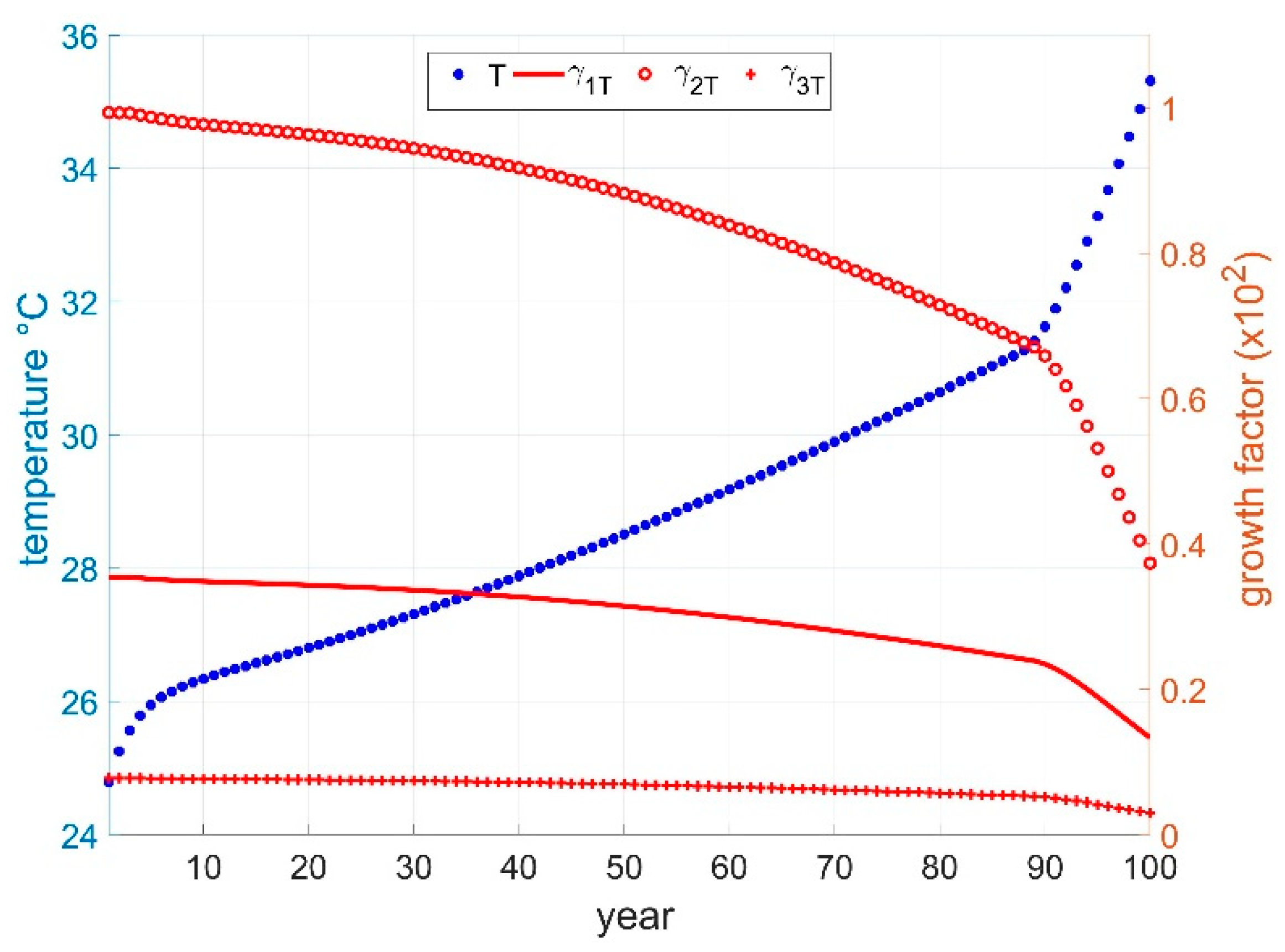
2.4. Global Warming Approach
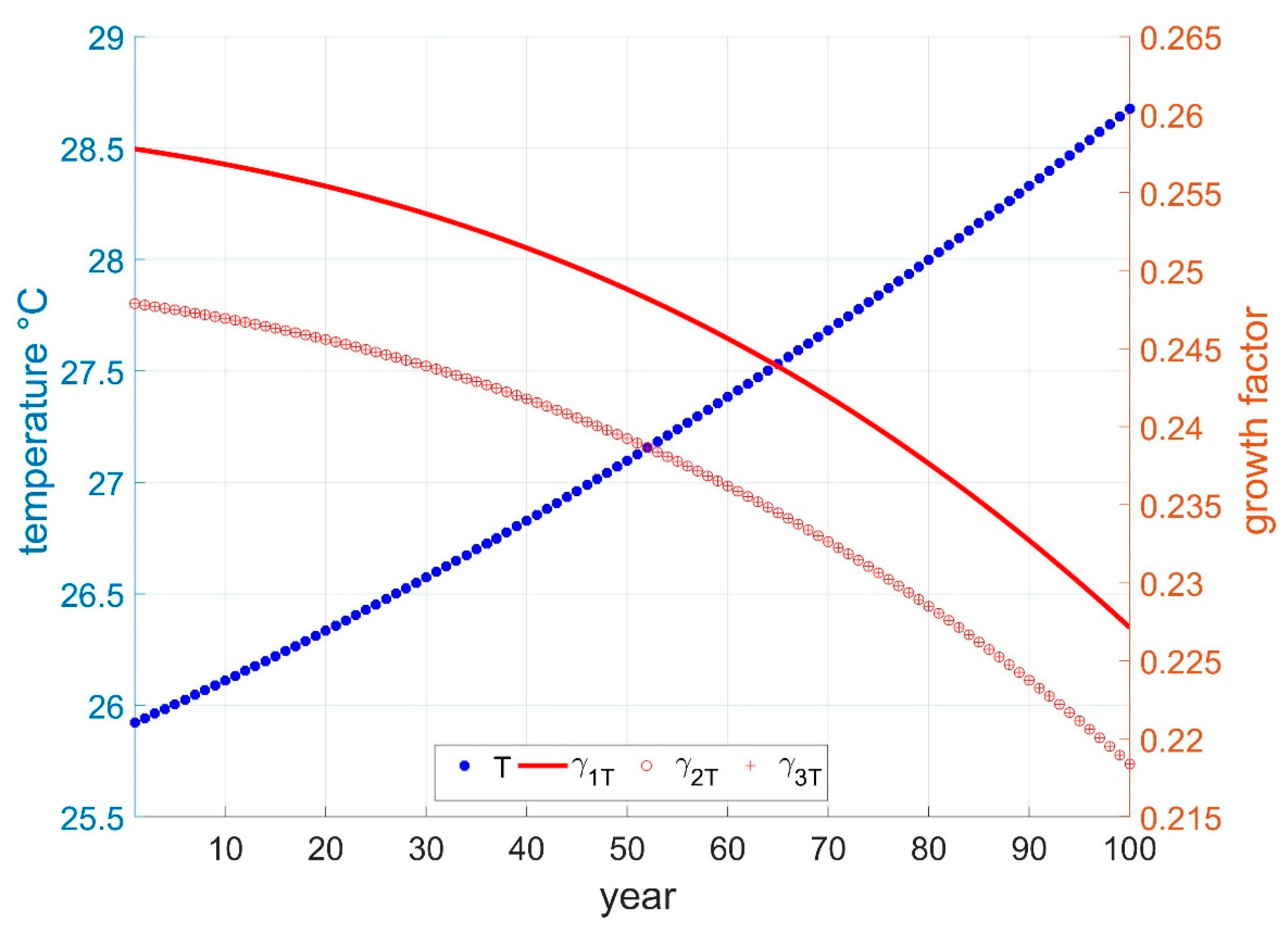
2.4.1. Global Temperature and Time
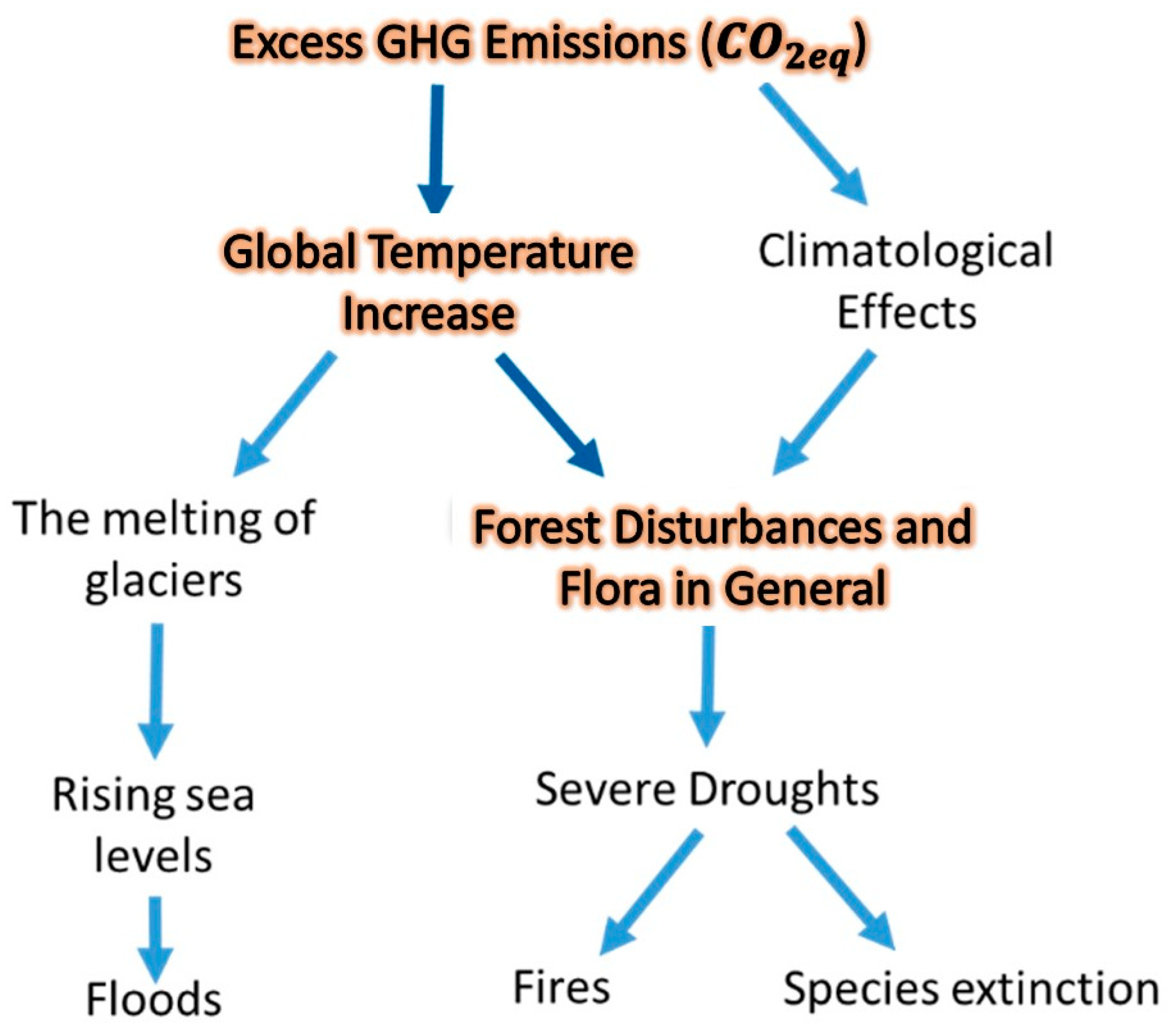
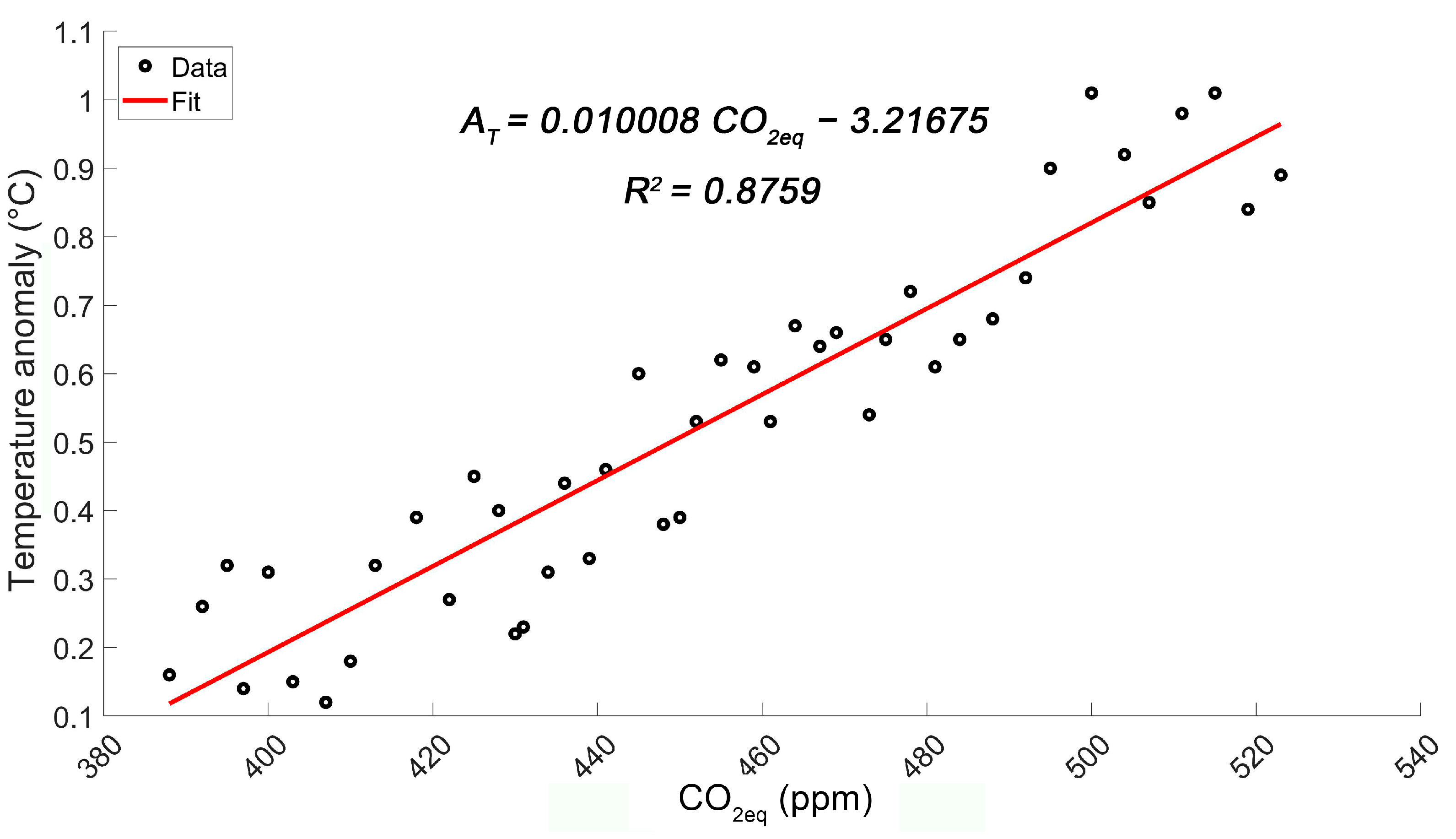
2.4.2. Global Temperature and GHG
2.4.3. Global Temperature and Development
3. Results and Discussion
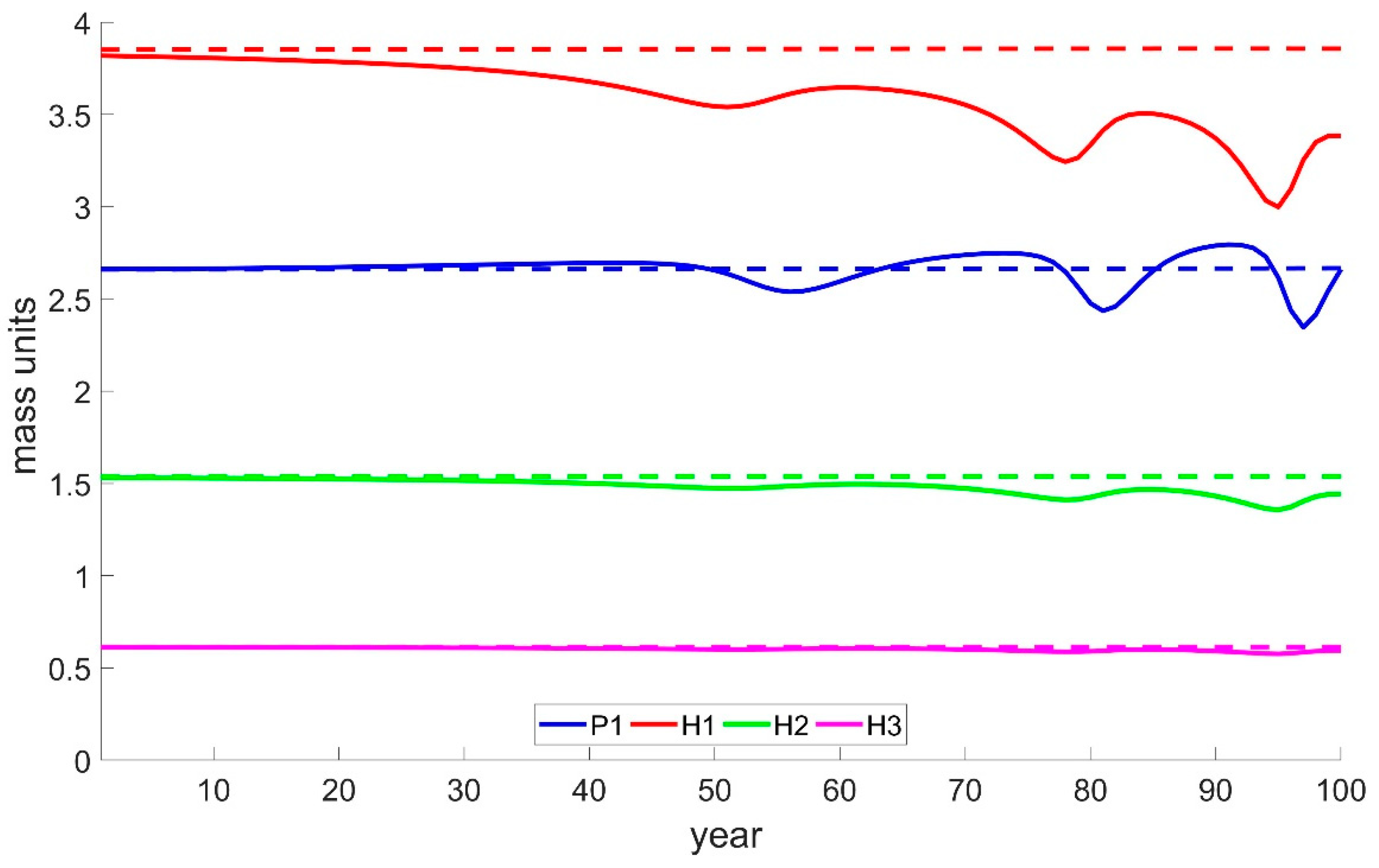
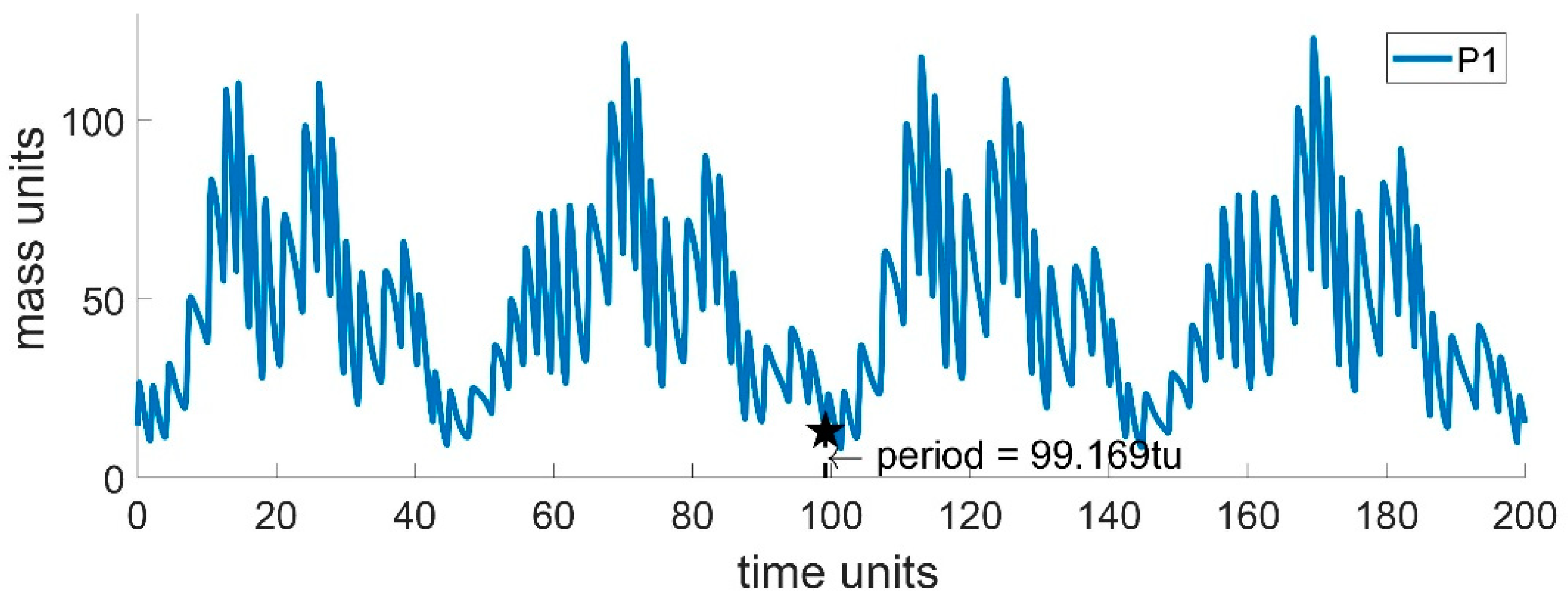
3.1. Case Study 1: Basic Multi Predator–Prey Model
3.2. Case Study 2: Basic Multi Predator–Prey Model
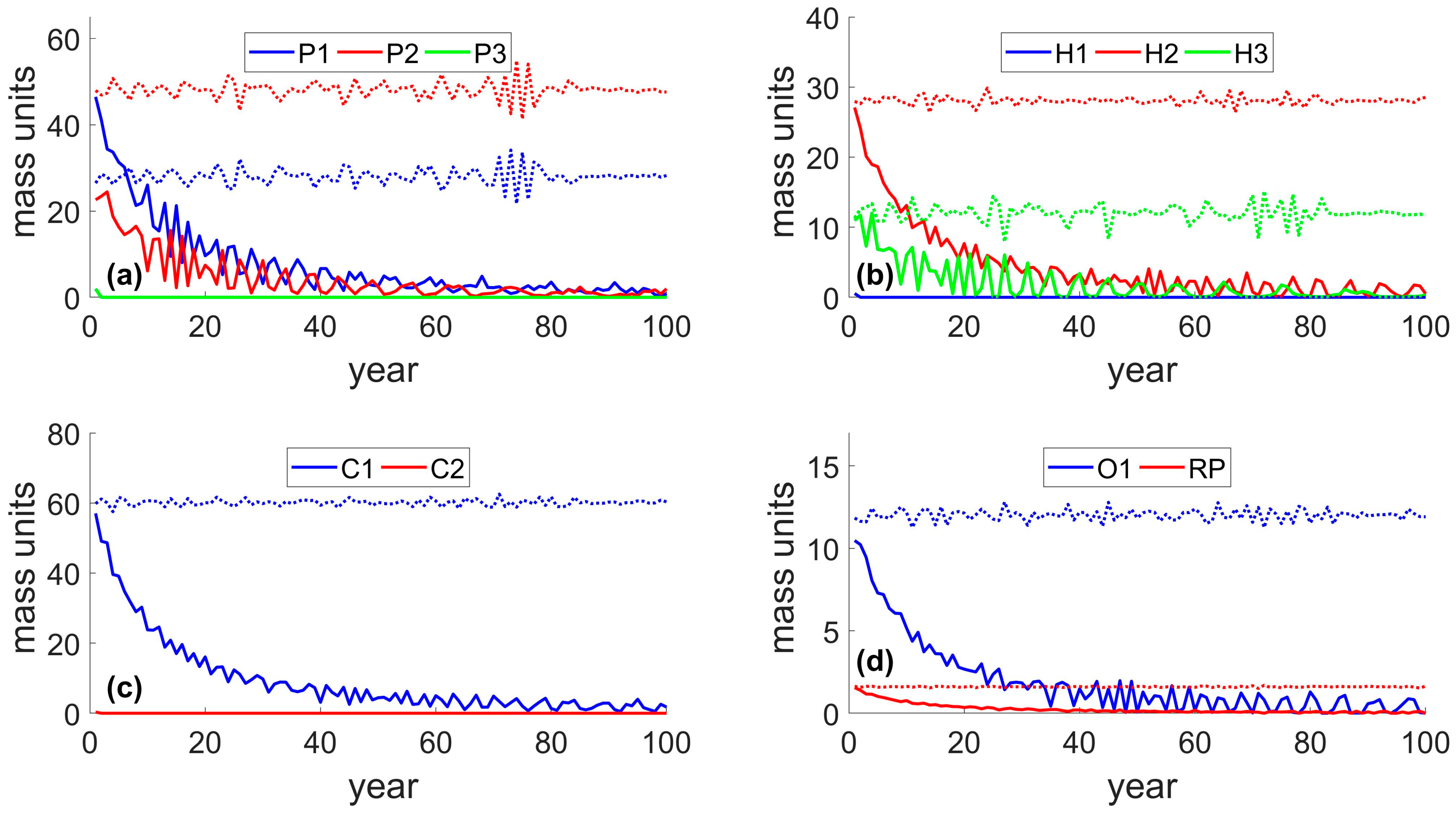
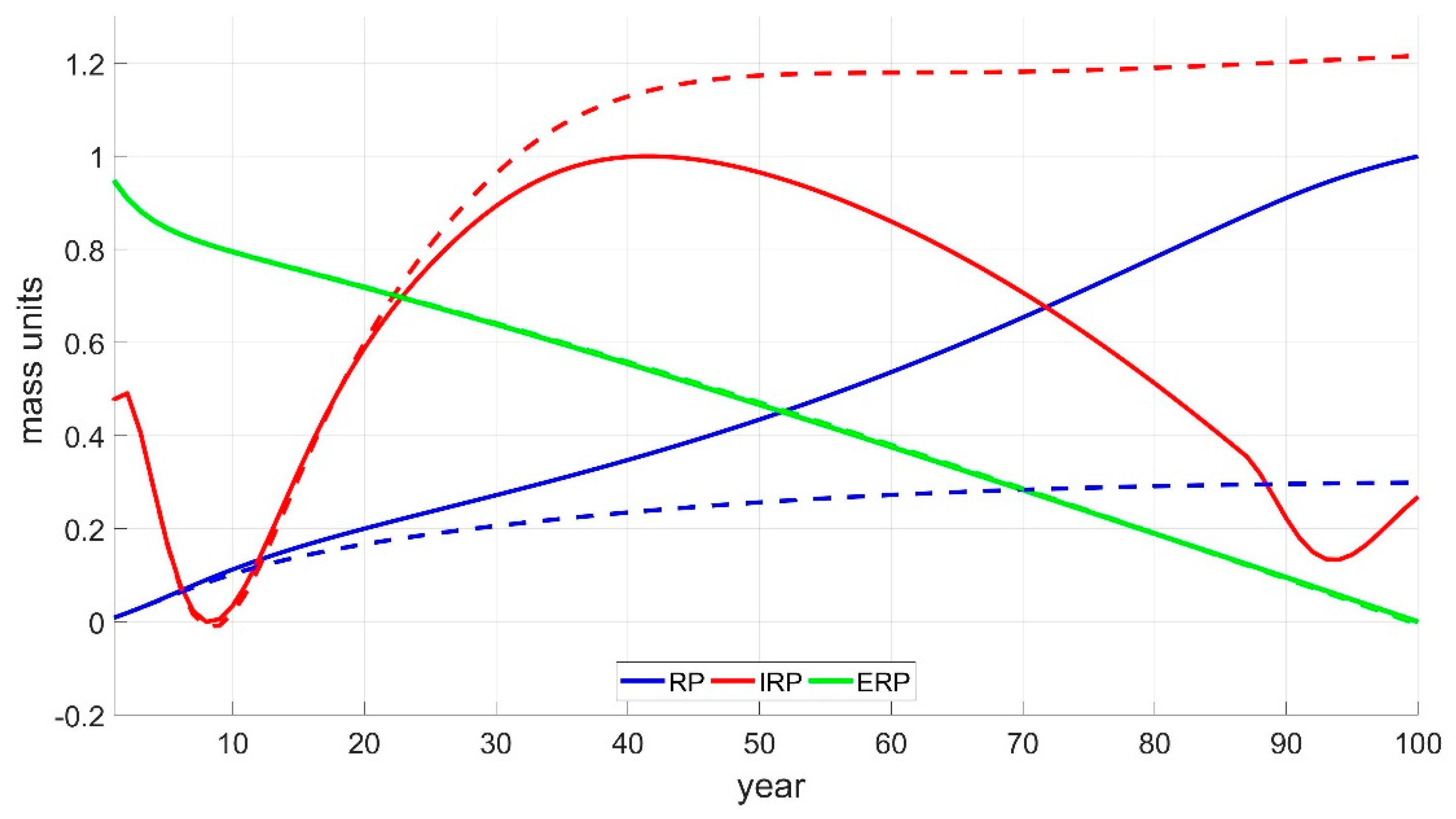
3.3. Case Study 3: Generalized Sustainability Socio–Ecological Model (GSSEM)
3.4. Overview of Ecosystem Responses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Abbreviation | Full Term | Explanation |
| GHG | Greenhouse Gas | Gases that trap heat in the atmosphere, contributing to global warming. |
| CO2eq | Carbon Dioxide Equivalent | A metric measure used to compare emissions from various GHGs based on their global warming potential. |
| GSSEM | Generalized Sustainability Socio–Ecological Model | Compartmental model that integrates ecological, social, and economic factors to analyze sustainability. |
| T | Temperature | Global temperature in the ecosystem in °C. |
| t | Time | Time as a variable in the equations. |
| Y | Compartment | General representation of a compartment |
| α | Birth Rate | A parameter representing the birth rate of compartments. |
| ϒ | Growth Rate | A parameter representing the growth rate of species due to interaction with others. |
| µ | Mortality Rate | A parameter representing the mortality rate of compartments. |
| P1, P2, P3 | Plant Compartments | Represents different types of plants in the ecosystems. |
| H1, H2, H3 | Herbivore Compartments | Represents different herbivore species in the ecosystems. |
| C1, C2 | Carnivore Compartments | Represents different carnivore species in the ecosystems. |
| HH | Human Compartment | Represents the humanity mass or resource. |
| IS | Industrial Sector | Represents the sector responsible for manufacturing and services within the human ecosystem. |
| EP | Energy Production | The sector involved in producing energy for human consumption. |
| RP | Resource Pool | Represents abiotic resources such as water, nutrients, and sunlight. |
| IRP | Inaccessible Resource Pool | Resources that are not directly usable by humans and gradually reintegrate into the environment. |
| ERP | Energy Resource Pool | Pool of finite non-renewable energy resources, including oil, natural gas, and coal. |
| AT | Anomaly of Temperature | The deviation of the current global temperature from a historical average or baseline. |
| tu | Time Unit | A generic unit of time used in the simulations of the ecosystems. |
| mu | Mass Units | Arbitrary units used to represent the mass of compartments in the simulations. |
| W | Wage | Represents the per capita wage. |
References
- Rockström, J.; Steffen, W.; Noone, K.; Persson, Å.; Chapin, F.S., III; Lambin, E.F.; Lenton, T.M.; Scheffer, M.; Folke, C.; Schellnhuber, H.J.; et al. A safe operating space for humanity. Nature 2009, 461, 472–475. [Google Scholar] [CrossRef]
- Steffen, W.; Richardson, K.; Rockström, J.; Cornell, S.E.; Fetzer, I.; Bennett, E.M.; Biggs, R.; Carpenter, S.R.; De Vries, W.; De Wit, C.A.; et al. Planetary boundaries: Guiding human development on a changing planet. Science 2015, 347, 1259855. [Google Scholar] [CrossRef]
- Varotsos, C.; Efstathiou, M. Has global warming already arrived? J. Atmos. Solar-Terr. Phys. 2019, 182, 31–38. [Google Scholar] [CrossRef]
- Hughes, L. Biological consequences of global warming: Is the signal already apparent? Trends Ecol. Evol. 2000, 15, 56–61. [Google Scholar] [CrossRef]
- Coudrain, A.; Francou, B.; Kundzewicz, Z.W. Glacier shrinkage in the Andes and consequences for water resources—Editorial. Hydrol. Sci. J. 2005, 50, 925–932. [Google Scholar] [CrossRef]
- Elewa, A.M.; Abdelhady, A.A. Past, present, and future mass extinctions. J. Afr. Earth Sci. 2020, 162, 103678. [Google Scholar] [CrossRef]
- Prăvălie, R. Major perturbations in the Earth’s forest ecosystems. Possible implications for global warming. Earth-Sci. Rev. 2018, 185, 544–571. [Google Scholar] [CrossRef]
- Lippmann, R.; Babben, S.; Menger, A.; Delker, C.; Quint, M. Development of Wild and Cultivated Plants under Global Warming Conditions. Curr. Biol. 2019, 29, R1326–R1338. [Google Scholar] [CrossRef]
- Sun, Q.; Miao, C.; Hanel, M.; Borthwick, A.G.; Duan, Q.; Ji, D.; Li, H. Global heat stress on health, wildfires, and agricultural crops under different levels of climate warming. Environ. Int. 2019, 128, 125–136. [Google Scholar] [CrossRef]
- Fattorini, S. Climate Change and Extinction Events. In Encyclopedia of Geology, Volume 1–6, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2021; Volume 5, pp. 585–595. [Google Scholar] [CrossRef]
- Rodriguez-Gonzalez, P.T.; Rico-Martinez, R.; Rico-Ramirez, V. Effect of feedback loops on the sustainability and resilience of human-ecosystems. Ecol. Model. 2020, 426, 109018. [Google Scholar] [CrossRef]
- Zhou, T.; Huang, X.; Zhang, N. Does the high-speed railway make cities more carbon efficient? Evidence from the perspective of the spatial spillover effect. Environ. Impact Assess. Rev. 2023, 101, 107137. [Google Scholar] [CrossRef]
- Gao, D.; Mo, X.; Xiong, R.; Huang, Z. Tax Policy and Total Factor Carbon Emission Efficiency: Evidence from China’s VAT Reform. Int. J. Environ. Res. Public Health 2022, 19, 9257. [Google Scholar] [CrossRef]
- Sinha, E. Identifying enablers and outcomes of circular economy for sustainable development: A systematic literature review. Bus. Strat. Dev. 2022, 5, 232–244. [Google Scholar] [CrossRef]
- Mamun, A.; Boubaker, S.; Nguyen, D.K. Green finance and decarbonization: Evidence from around the world. Financ. Res. Lett. 2022, 46, 102807. [Google Scholar] [CrossRef]
- Martinho, V.J.P.D. European Union farming systems: Insights for a more sustainable land use. Land Degrad. Dev. 2022, 33, 527–544. [Google Scholar] [CrossRef]
- Foley, J.A.; DeFries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Chapin, F.S.; Coe, M.T.; Daily, G.C.; Gibbs, H.K.; et al. Global consequences of land use. Science 2005, 309, 570–574. [Google Scholar] [CrossRef]
- Ostrom, E. A General framework for analyzing sustainability of social-ecological systems. Science 2009, 325, 419–422. [Google Scholar] [CrossRef]
- Boccara, N. Modeling Complex Systems; Springer Nature: Dordrecht, The Netherlands, 2010; ISBN 9781441965615. [Google Scholar]
- Yang, Y. Sustainability Analysis of Enterprise Performance Management Driven by Big Data and Internet of Things. Sustainability 2023, 15, 4839. [Google Scholar] [CrossRef]
- Nisal, A.; Diwekar, U.; Hanumante, N.; Shastri, Y.; Cabezas, H. Integrated model for food-energy-water (FEW) nexus to study global sustainability: The main generalized global sustainability model (GGSM). PLoS ONE 2022, 17, e0267403. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, N.; Pinheiro, S.; Magalhães, S.; Tarroso, P.; Vincent, A. Predicting the impacts of climate change on the distribution of European syngnathids over the next century. Front. Mar. Sci. 2023, 10, 1138657. [Google Scholar] [CrossRef]
- Diwekar, U.; Nisal, A.; Shastri, Y.; Cabezas, H. Optimal Control for Deriving Policies for Global Sustainability. Comput. Aided Chem. Eng. 2023, 52, 2471–2476. [Google Scholar] [CrossRef]
- Flensborg, L.C.; A Maureaud, A.; Bravo, D.N.; Lindegren, M. An indicator-based approach for assessing marine ecosystem resilience. ICES J. Mar. Sci. 2023, 80, 1487–1499. [Google Scholar] [CrossRef]
- Pindyck, R.S. The Use and Misuse of Models for Climate Policy. Rev. Environ. Econ. Policy 2017, 11, 100–114. [Google Scholar] [CrossRef]
- Weyant, J. Some Contributions of Integrated Assessment Models of Global Climate Change. Rev. Environ. Econ. Policy 2017, 11, 115–137. [Google Scholar] [CrossRef]
- Stehfest, E.; van Vuuren, D.; Kram, T.; Bouwman, L. Integrated Assessment of Global Environmental Change with IMAGE 3.0: Model Description and Policy Applications. The Hague: PBL Netherlands Environmental Assessment Agency; 2014. Available online: https://www.pbl.nl/sites/default/files/downloads/pbl-2014-integrated_assessment_of_global_environmental_change_with_image30_735.pdf (accessed on 27 August 2024).
- Calvin, K.; Patel, P.; Clarke, L.; Asrar, G.; Bond-Lamberty, B.; Cui, R.Y.; Di Vittorio, A.; Dorheim, K.; Edmonds, J.; Hartin, C.; et al. GCAM v5.1: Representing the linkages between energy, water, land, climate, and economic systems. Geosci. Model. Dev. 2019, 12, 677–698. [Google Scholar] [CrossRef]
- King, C.W. An integrated biophysical and economic modeling framework for long-term sustainability analysis: The HARMONEY model. Ecol. Econ. 2019, 169, 106464. [Google Scholar] [CrossRef]
- Jørgensen, S.E.; Fath, B.D. Fundamentals of Ecological Modelling: Applications in Environmental Management and Research, 4th ed.; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Bertozzi, A.L.; Franco, E.; Mohler, G.; Short, M.B.; Sledge, D. The challenges of modeling and forecasting the spread of COVID-19. Proc. Natl. Acad. Sci. USA 2020, 117, 16732–16738. [Google Scholar] [CrossRef]
- Mwalili, S.; Kimathi, M.; Ojiambo, V.; Gathungu, D.; Mbogo, R. SEIR model for COVID-19 dynamics incorporating the environment and social distancing. BMC Res. Notes 2020, 13, 1–5. [Google Scholar] [CrossRef]
- He, S.; Peng, Y.; Sun, K. SEIR modeling of the COVID-19 and its dynamics. Nonlinear Dyn. 2020, 101, 1667–1680. [Google Scholar] [CrossRef]
- Rodriguez-Gonzalez, P.T.; Rico-Martinez, R.; Rico-Ramirez, V. An integrated stochastic economic-ecological-social model with stratified-population. Ecol. Model. 2018, 368, 15–26. [Google Scholar] [CrossRef]
- Nisal, A.; Diwekar, U.; Hanumante, N.; Shastri, Y.; Cabezas, H.; Ramirez, V.R.; Rodríguez-González, P.T. Evaluation of global techno-socio-economic policies for the FEW nexus with an optimal control based approach. Front. Sustain. 2022, 3, 948443. [Google Scholar] [CrossRef]
- Hanumante, N.; Shastri, Y.; Nisal, A.; Diwekar, U.; Cabezas, H. Water stress-based price for global sustainability: A study using generalized global sustainability model (GGSM). Clean Technol. Environ. Policy 2024, 1–20. [Google Scholar] [CrossRef]
- Jones, M.C.; Grosse, G.; Treat, C.; Turetsky, M.; Anthony, K.W.; Brosius, L. Past permafrost dynamics can inform future permafrost carbon-climate feedbacks. Commun. Earth Environ. 2023, 4, 272. [Google Scholar] [CrossRef]
- Schuur, E.A.; Abbott, B.W.; Commane, R.; Ernakovich, J.; Euskirchen, E.; Hugelius, G.; Grosse, G.; Jones, M.; Koven, C.; Leshyk, V.; et al. Permafrost and Climate Change: Carbon Cycle Feedbacks From the Warming Arctic. Annu. Rev. Environ. Resour. 2022, 47, 343–371. [Google Scholar] [CrossRef]
- Kimmins, H.; Blanco, J.A.; Seely, B.; Welham, C.; Scoullar, K. Forecasting Forest Futures: A Hybrid Modelling Approach to the Assessment of Sustainability of Forest Ecosystems and Their Values; Routledge: Oxfordshire, UK, 2010; pp. 1–281. [Google Scholar] [CrossRef]
- Martin, R.; Schlüter, M. Combining system dynamics and agent-based modeling to analyze social-ecological interactions—An example from modeling restoration of a shallow lake. Front. Environ. Sci. 2015, 3, 66. [Google Scholar] [CrossRef]
- Dong, L.; Huang, Z. Some evidence and new insights for feedback loops of human-nature interactions from a holistic Earth perspective. J. Clean. Prod. 2023, 432, 139667. [Google Scholar] [CrossRef]
- Ripple, W.J.; Wolf, C.; Lenton, T.M.; Gregg, J.W.; Natali, S.M.; Duffy, P.B.; Rockström, J.; Schellnhuber, H.J. Many risky feedback loops amplify the need for climate action. One Earth 2023, 6, 86–91. [Google Scholar] [CrossRef]
- E Visser, M.; Both, C. Shifts in phenology due to global climate change: The need for a yardstick. Proc. R. Soc. B Biol. Sci. 2005, 272, 2561–2569. [Google Scholar] [CrossRef]
- Janni, M.; Maestri, E.; Gullì, M.; Marmiroli, M.; Marmiroli, N. Plant responses to climate change, how global warming may impact on food security: A critical review. Front. Plant Sci. 2023, 14, 1297569. [Google Scholar] [CrossRef]
- I Zandalinas, S.; Balfagón, D.; Gómez-Cadenas, A.; Mittler, R. Plant responses to climate change: Metabolic changes under combined abiotic stresses. J. Exp. Bot. 2022, 73, 3339–3354. [Google Scholar] [CrossRef]
- Schmitz, O.J.; Hambäck, P.A.; Beckerman, A.P. Trophic Cascades in Terrestrial Systems: A Review of the Effects of Carnivore Removals on Plants. Am. Nat. 2000, 155, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, P.; Tucker, C.; Sy, H. Tree density and species decline in the African Sahel attributable to climate. J. Arid. Environ. 2012, 78, 55–64. [Google Scholar] [CrossRef]
- Hasibuan, A.; Supriatna, A.K.; Rusyaman, E.; Biswas, H.A. Harvested Predator–Prey Models Considering Marine Reserve Areas: Systematic Literature Review. Sustainability 2023, 15, 12291. [Google Scholar] [CrossRef]
- Korobeinikov, A. Stability of ecosystem: Global properties of a general predator-prey model. Math. Med. Biol. A J. IMA 2009, 26, 309–321. [Google Scholar] [CrossRef]
- Kotecha, P.; Diwekar, U.; Cabezas, H. Model-based approach to study the impact of biofuels on the sustainability of an ecological system. Clean Technol. Environ. Policy 2013, 15, 21–33. [Google Scholar] [CrossRef]
- Shastri, Y.; Diwekar, U.; Cabezas, H.; Williamson, J. Is sustainability achievable? Exploring the limits of sustainability with model systems. Environ. Sci. Technol. 2008, 42, 6710–6716. [Google Scholar] [CrossRef]
- Chapra, S.C.; Canale, R.P. Numerical Methods for Engineers, 6th ed.; McGraw-Hill: New York, NY, USA, 2010. [Google Scholar]
- Pawlowski, C.W.; Fath, B.D.; Mayer, A.L.; Cabezas, H. Towards a sustainability index using information theory. Energy 2005, 30, 1221–1231. [Google Scholar] [CrossRef]
- Lotka, A.J. Elements of Physical Biology; Williams and Wilkins: Philadelphia, PA, USA, 1925. [Google Scholar]
- Volterra, V. Variazioni e fluttuazioni del numero d’individui in specie animali conviventi. Mem. Della R. Accad. Naz. Dei Lincei 1926, 2, 31–113. [Google Scholar]
- Intergovernmental Panel on Climate Change (IPCC). Impacts of 1.5 °C Global Warming on Natural and Human Systems. In Global Warming of 1.5 °C: IPCC Special Report on Impacts of Global Warming of 1.5 °C Above Pre-Industrial Levels in Context of Strengthening Response to Climate Change, Sustainable Development, and Efforts to Eradicate Poverty; Cambridge University Press: Cambridge, UK, 2022; pp. 175–312. [Google Scholar]
- I. P. on C. C. (IPCC). Climate Change 2021—The Physical Science Basis: Working Group I Contribution to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. In Climate Change 2021—The Physical Science Basis; Cambridge University Press: Cambridge, UK, 2023. [Google Scholar] [CrossRef]
- I. P. on C. C. (IPCC). Climate Change 2022—Impacts, Adaptation and Vulnerability: Working Group II Contribution to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. In Climate Change 2022—Impacts, Adaptation and Vulnerability; Cambridge University Press: Cambridge, UK, 2023. [Google Scholar] [CrossRef]
- NOAA Global Monitoring Laboratory—THE NOAA ANNUAL GREENHOUSE GAS INDEX (AGGI). Available online: https://gml.noaa.gov/aggi/aggi.html (accessed on 27 August 2024).
- Data.GISS: GISS Surface Temperature Analysis (GISTEMP v4). Available online: https://data.giss.nasa.gov/gistemp/ (accessed on 27 August 2024).
- Nahar, K.; Hasanuzzaman, M.; Ahamed, K.U.; Hakeem, K.R.; Ozturk, M.; Fujita, M. Plant Responses and Tolerance to High Temperature Stress: Role of Exogenous Phytoprotectants. In Crop Production and Global Environmental Issues; Springer: Berlin/Heidelberg, Germany, 2015; pp. 385–435. [Google Scholar] [CrossRef]
- Paiva, S.L.; Savi, M.A.; Viola, F.M.; Leiroz, A.J. Global warming description using Daisyworld model with greenhouse gases. Biosystems 2014, 125, 1–15. [Google Scholar] [CrossRef]
- Lin, Y.-S.; Medlyn, B.E.; Ellsworth, D.S. Temperature responses of leaf net photosynthesis: The role of component processes. Tree Physiol. 2012, 32, 219–231. [Google Scholar] [CrossRef]
- O’Sullivan, O.S.; Heskel, M.A.; Reich, P.B.; Tjoelker, M.G.; Weerasinghe, L.K.; Penillard, A.; Zhu, L.; Egerton, J.J.G.; Bloomfield, K.J.; Creek, D.; et al. Thermal limits of leaf metabolism across biomes. Glob. Chang. Biol. 2017, 23, 209–223. [Google Scholar] [CrossRef] [PubMed]
- Rocklöv, J.; Ebi, K.; Forsberg, B. Mortality Related to Temperature and Persistent Extreme Temperatures—A Study of Cause-Specific and Age Stratified Mortality. Epidemiology 2011, 22, S13. [Google Scholar] [CrossRef]
- Estes, J.A.; Terborgh, J.; Brashares, J.S.; Power, M.E.; Berger, J.; Bond, W.J.; Carpenter, S.R.; Essington, T.E.; Holt, R.D.; Jackson, J.B.C.; et al. Trophic downgrading of planet earth. Science 2011, 333, 301–306. [Google Scholar] [CrossRef]
- da Gama, J.T. The Role of Soils in Sustainability, Climate Change, and Ecosystem Services: Challenges and Opportunities. Ecologies 2023, 4, 552–567. [Google Scholar] [CrossRef]
- Rodríguez, B.C.; Durán-Zuazo, V.H.; Rodríguez, M.S.; García-Tejero, I.F.; Ruiz, B.G.; Tavira, S.C. Conservation Agriculture as a Sustainable System for Soil Health: A Review. Soil Syst. 2022, 6, 87. [Google Scholar] [CrossRef]
- Davis, A.S.; Hill, J.D.; Chase, C.A.; Johanns, A.M.; Liebman, M. Increasing Cropping System Diversity Balances Productivity, Profitability and Environmental Health. PLoS ONE 2012, 7, e47149. [Google Scholar] [CrossRef] [PubMed]
- Fetene, A.; Yeshitela, K.; Gebremariam, E. The effects of anthropogenic landscape change on the abundance and habitat use of terrestrial large mammals of Nech Sar National Park. Environ. Syst. Res. 2019, 8, 19. [Google Scholar] [CrossRef]
- Said, M.Y.; Ogutu, J.O.; Kifugo, S.C.; Makui, O.; Reid, R.S.; de Leeuw, J. Effects of extreme land fragmentation on wildlife and livestock population abundance and distribution. J. Nat. Conserv. 2016, 34, 151–164. [Google Scholar] [CrossRef]
- Stirling, I.; Derocher, A.E. Effects of climate warming on polar bears: A review of the evidence. Glob. Chang. Biol. 2012, 18, 2694–2706. [Google Scholar] [CrossRef]
- World Population Prospects—Population Division—United Nations. Available online: https://population.un.org/wpp/ (accessed on 27 August 2024).
- Climate Change. Available online: https://www.who.int/news-room/fact-sheets/detail/climate-change-and-health (accessed on 24 October 2024).
- Ripple, W.J.; Wolf, C.; Newsome, T.M.; Barnard, P.; Moomaw, W.R. World Scientists’ Warning of a Climate Emergency. Bioscience 2020, 70, 8–12. [Google Scholar] [CrossRef]
- Xu, Z.; Sheffield, P.E.; Hu, W.; Su, H.; Yu, W.; Qi, X.; Tong, S. Climate Change and Children’s Health—A Call for Research on What Works to Protect Children. Int. J. Environ. Res. Public Health 2012, 9, 3298–3316. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Wang, S.; Hu, Y.; Yue, M.; Zhang, T.; Liu, Y.; Tian, J.; Shang, K. Impact of ambient temperature on morbidity and mortality: An overview of reviews. Sci. Total. Environ. 2017, 586, 241–254. [Google Scholar] [CrossRef] [PubMed]
- Pintor, M.P. The future of the temperature–mortality relationship. Lancet Public Health 2024, 9, e636–e637. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Parameters | Initial Values | ||||||
|---|---|---|---|---|---|---|---|
| Plant | Herbivores | ||||||
| 1.2 | 0.20 | 0.532 | 2 | ||||
| 0.02 | 0.10 | 0.266 | 1.5 | ||||
| 0.02 | 0.05 | 0.133 | 1 | ||||
| 0.02 | 0.5 | ||||||
| Growth Parameters | Initial Values | ||||||
|---|---|---|---|---|---|---|---|
| Plant | Herbivores | Carnivores/Omnivore | 7.93 | ||||
| 0.01 | 0.01 | 14.59 | 0.01 | 14.59 | |||
| 0.01 | 0.01 | 0.01 | 32.89 | ||||
| 0.01 | 0.1 | 0.1 | 13.76 | ||||
| 0.01 | 0.01 | 0.75 | 9.16 | ||||
| 0.01 | 0.01 | Resources | 37.5 | ||||
| 0.01 | 0.1 | 0.75 | 11.47 | ||||
| 0.01 | 0.01 | 0.25 | 26.03 | ||||
| 0.01 | 0.01 | 0.25 | 25.19 | ||||
| 0.01 | 0.1 | 0.26 | 11.46 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tovar-Ortiz, S.A.; Rodriguez-Gonzalez, P.T.; Tovar-Gómez, R. Modeling the Impact of Global Warming on Ecosystem Dynamics: A Compartmental Approach to Sustainability. World 2024, 5, 1077-1100. https://doi.org/10.3390/world5040054
Tovar-Ortiz SA, Rodriguez-Gonzalez PT, Tovar-Gómez R. Modeling the Impact of Global Warming on Ecosystem Dynamics: A Compartmental Approach to Sustainability. World. 2024; 5(4):1077-1100. https://doi.org/10.3390/world5040054
Chicago/Turabian StyleTovar-Ortiz, Sinue A., Pablo T. Rodriguez-Gonzalez, and Rigoberto Tovar-Gómez. 2024. "Modeling the Impact of Global Warming on Ecosystem Dynamics: A Compartmental Approach to Sustainability" World 5, no. 4: 1077-1100. https://doi.org/10.3390/world5040054
APA StyleTovar-Ortiz, S. A., Rodriguez-Gonzalez, P. T., & Tovar-Gómez, R. (2024). Modeling the Impact of Global Warming on Ecosystem Dynamics: A Compartmental Approach to Sustainability. World, 5(4), 1077-1100. https://doi.org/10.3390/world5040054