1. Introduction
The island of Guam was invaded by
Aulacaspis yasumatsui Takagi (Hemiptera, Diaspididae) in 2003 [
1,
2]. Mortality among the host tree
Cycas micronesica K.D. Hill population was immediate [
3,
4]. The International Union for Conservation of Nature (IUCN) listed this insular cycad species as Endangered in 2006 [
5], and it was added to the United States’ Endangered Species Act (ESA) in 2015 [
6].
The infestation by this non-native specialist armored scale insect generated numerous negative outcomes for the host plant on Guam. Within three years, seedling mortality was 100%, juvenile mortality was 97%, and adult mortality was 63% [
4]. Ongoing plant mortality due to
A. yasumatsui herbivory and other biotic threats continued, and the mortality reached 96% by 2020 [
4]. The fact that this cycad species was the most abundant tree in Guam’s forests in 2002 [
7] underscores the compelling nature of this case study.
This example from Guam adds to a growing body of information on how invasive species threaten native organisms on islands. Indeed, invasive species are a major driver of biodiversity loss on islands, where their impacts are often more severe than on continents [
8,
9,
10]. The many previously studied cases have accumulated knowledge about the control of invasive species specific to islands [
11]. International conferences have been convened to assimilate what has been learned to conserve threatened native island species through the better management of island invasives [
12,
13]. Mammals [
14], plants [
15], and insects [
16] have been studied independently, and in conjunction with each other, in the many case studies of island invasions. Herbivorous insect invasions, in particular, exert extreme consequences on their host populations, sometimes eliminating the host [
17]. The costs incurred while responding to island invasions indicate that the greatest costs are associated with insect invasions, followed by plant invasions, and then mammal invasions [
18].
Prior to this invasion of Guam,
C. micronesica seedlings were the dominant component of the forest floor, with 1600 seedlings per ha reported within the areas of occupancy [
4]. The gestalt appearance of the Guam forests drastically changed as a result of the immediate culling of the seedlings. We also noticed that all the newly emerging seedlings during the early years of the invasion were immediately killed by the ubiquitous
A. yasumatsui pressures; therefore, we predicted long-lasting negative outcomes for the surviving cycad population, due to the sustained lack of regeneration and recruitment [
19]. Following this prediction, we determined the influence of chronic herbivory on
C. micronesica microstrobili traits, to begin to understand how this specialist scale herbivore damages regeneration [
20]. During the years immediately after the invasion, the volume of the male cones declined to 24% of the pre-invasion volume, with some increase in volume during subsequent years. However, as of 2021, the male cone volume remained stunted at 57% of the pre-invasion size. The male cones of this cycad tree are crucial for sustaining the
Anatrachyntis Meyrick (Lepidoptera, Cosmopterigidae) pollinator population, by serving as a venue for sexual encounters and ovipositioning, and then enabling the life history stages of larval development and recruitment to the pupal stage [
1,
21]. Following the invasion, the number of
Anatrachyntis pupae counted from each cone declined to 64% of the pre-invasion numbers [
20], with no recovery in the ensuing years.
Using height increment metrics, an estimated 70 y of demographic depth of the Guam
C. micronesica population was culled from the surviving tree population between 2003 and 2020 [
22]. Every added year of ongoing failed conservation actions delay a recovery of the reproductive output and the recruitment of long-lived seedlings, which will cause even more years of loss of demographic depth. Clearly, new knowledge about
C. micronesica reproductive efforts and regeneration potential is urgently needed to inform ongoing conservation efforts to mitigate the lack of regeneration caused by the
A. yasumatsui invasion.
A detailed look at how the female trees of this dioecious gymnosperm have responded to this consequential invasion is required to augment what we have learned about male tree behaviors. Our objectives were to address this need by quantifying the long-term post-invasion changes in the megastrobilus size metrics and seed set in the east-coast karst forests of northern Guam.
2. Materials and Methods
Our field work began in 2001 and continued through 2022. We employed the same habitats that were used for the microstrobili measurements, and the site characteristics have been previously described [
20]. This east-coast terrain historically contained high density areas of occupancy, with the presence of
C. micronesica individuals from the seedling stage to more than 5 m in height. The field measurements were performed during November–December of each year. The habitats were invaded by
A. yasumatsui in 2005, and by early 2006 the standing crown of leaves had been killed on every stem. For years, each cohort of newly added leaves was immediately infested, and the trees rarely contained two sequential cohorts of living leaves.
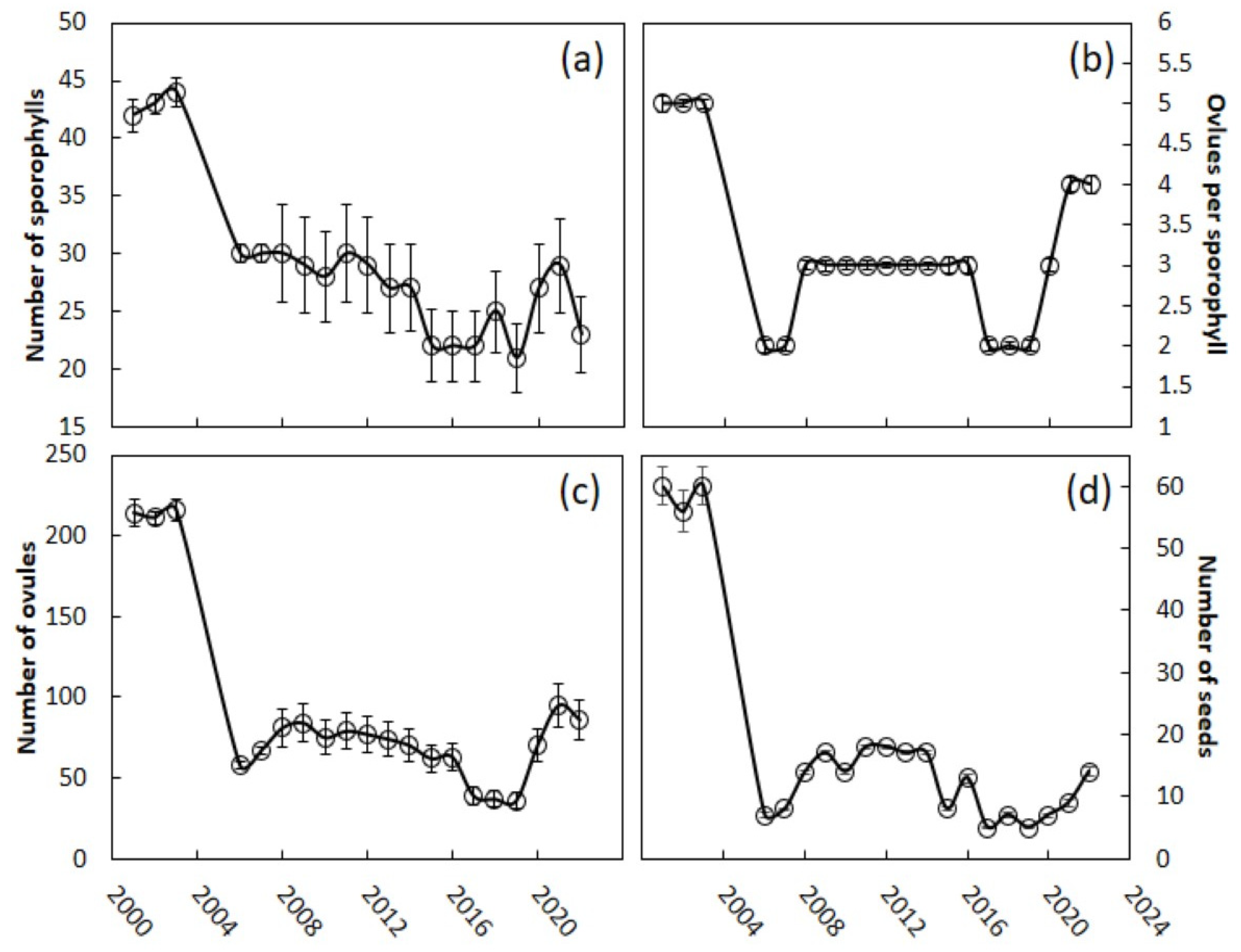
The peak season for
C. micronesica megastrobili emergence before the invasion was April, with ovule receptivity occurring about one month later [
23]. Determining the seed set in November─December was unambiguous for these naked gymnosperm ovules, due to the rapid increase in volume of the fertilized ovules [
24], while the undeveloped ovules remained attached to the megasporophylls without a substantial increase in size (
Figure 1a). The seed sarcotesta changes in color from green to bronze after about one year, so restricting our measurements to the trees with bright green seeds ensured our data were limited to the current year megastrobili. The megasporophylls were positively geotropic and relaxed at the time of our measurements (
Figure 1b). A total of 50 megastrobili were measured each calendar year to quantify the various size metrics. The number of seeds and undeveloped ovules were counted and recorded for each individual sporophyll and the number of sporophylls was counted. The stem height was measured from the soil surface to the top of the cataphylls that terminate the stem apex.
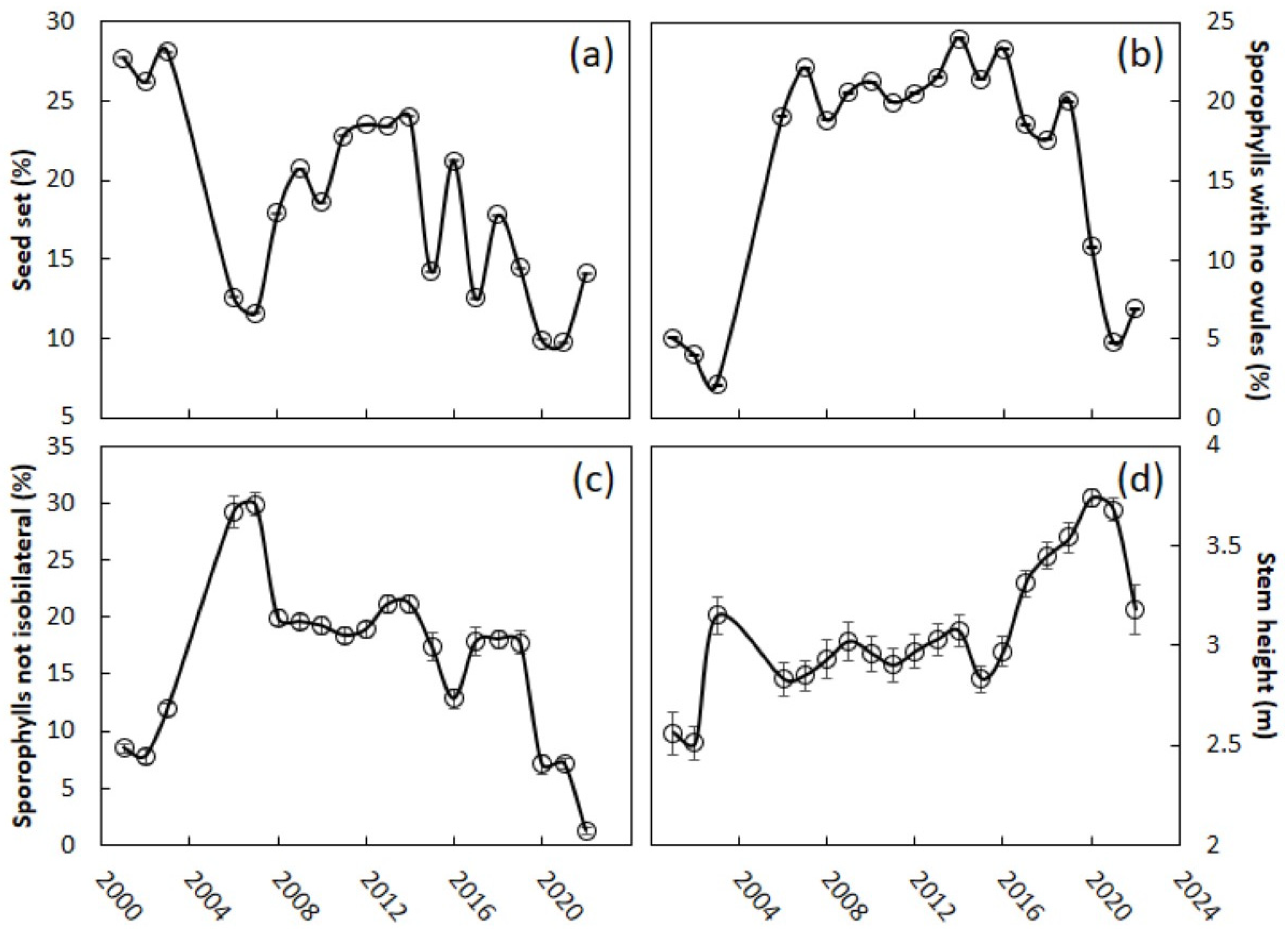
For the measure of reproductive effort, we added undeveloped ovules and seeds to determine the total number of ovules that were produced within each strobilus. For the measure of reproductive output, we added the number of seeds for each strobilus. The seed set per strobilus was calculated as a percentage. The number of ovules per sporophyll was calculated for each strobilus. Megasporophylls are leaf homologs [
25], and their function is to support ovules and seeds. However, some megasporophylls contain no ovules (
Figure 2a). The number of sporophylls which contained no ovules was counted. Megasporophylls are typically isobilateral with two opposing ovules at each sporophyll node, but shortly after the invasion many sporophylls displayed an odd number of ovules (
Figure 2b). The number of sporophylls in this category was counted. We then calculated the percentage of sporophylls within these two categories for each megastrobilus.
Statistical analyses were performed using R (Version 3.6.3), to determine the differences between the pre-invasion years and the post-invasion years, or across years within each group. For all the variables, we first tested the data for normality and homogeneity of variances between the two groups, or within the groups. All the reproductive variables associated with the megastrobilus and stem height measurements indicated that the data were not normally distributed (Shapiro–Wilk test, W < 0.98, p < 0.0027), and the variances were not equal (folded f test, p < 0.013). To compare any two samples, the benchmark pre-invasion years with the post-invasion years, or the pre-invasion years with a subset of the post-invasion years, we used a Kruskal–Wallis rank-sum, two-tailed test. For the derived proportion variables, we used logistic regression (with a binomial distribution) to test for differences in the pre- versus post-invasion years’ differences. For example, for a seed set, the number of seeds was used as the dependent variable compared to total number of ovules for each year, and the pre- versus post-invasion years was the independent variable in the model. The reliability of the models was determined by the residual deviance as an indicator of overdispersion. If over-dispersed, we fitted the data to a quasibinomial distribution.
4. Discussion
The 2003 invasion of Guam by
A. yasumatsui [
1,
2] has caused immeasurable negative ecosystem changes, partly because the naïve insular host tree species was the most abundant tree at the time, [
7] and therefore served as a foundation species [
26]. The influences of this invasion on
C. micronesica mortality have been well-documented and reported [
3,
4]. By 2013, this species was no longer on the list of the 20 most abundant tree species in Guam [
27], and by 2020 the population reached 96% mortality [
4]. In order to foster effective species recovery, conservationists need to enumerate every distinct threat, then understand the driving mechanisms that underlie each threat, and how the disparate threats interact. We have shown that continued plant mortality is not the only negative outcome of this insect invasion, but the continued failure of
C. micronesica to regenerate and recruit is also critical. Our historical predictions of sustained constraints on
C. micronesica regeneration following this invasion [
19] are now more fully understood. Our results reveal that a reduction in female reproductive effort (the number of ovules per megastrobilus) and reproductive output (the number of seeds per megastrobilus) occurred immediately after the invasion, and these important measures of plant fecundity have shown few signs of recovery among the surviving tree population of 2022. Considering the extent of population-level mortality and the unhealthy appearance of the surviving trees, the sustained production of megastrobili throughout the living population is surprising.
We recently reported similar negative outcomes for male
C. micronesica trees, where the microstrobilus size metrics declined immediately after the invasion [
20]. While both of the tree sexes had a large reduction in strobilus size traits (76% reduction in microstrobilus volume after the scale invasion, and an 88% reduction in megastrobilus seed number), microstrobilus volume has increased in recent years, but the megastrobilus size traits have not shown signs of recovery. Moreover, the number of
Anatrachyntis pupae counted in
C. micronesica microstrobili throughout the years since the invasion has revealed that some cones contained few to no pupae in recent years [
20]. This development indicates that sustained observations of pollinator recruitment by conservationists are warranted to foresee a possible future decline in the pollinator population, as this moth relies on
C. micronesica microstrobili for ovipositioning, larval development, and recruitment to the pupal stage [
21,
28]. Our collective observations of population-level behaviors indicate that the reduced reproductive effort of the female trees and the rapid mortality of seedlings immediately after emergence comprise the major reasons for the sustained reduction in
C. micronesica regeneration following this consequential island invasion.
The construction of a sporophyll with no ovules (the blank sporophylls) or a reduced number of ovules (the sporophylls with odd numbers of ovules) may be one means of reducing the costs of reproduction. This plant behavior deserves further study, as it may be a universal response to all abiotic and biotic forms of female cycad plant stress. One approach to reduce the reproductive costs for these female trees may be to increase the window of time between megastrobilus initiation events [
29]. This behavior would give the tree time to increase the stored pool of nonstructural resources required to support a new strobilus to maturity. However, this behavior would also mean the individual would remain uninvolved in population-level gamete exchange for extended periods of time. A second approach to reduce the costs of reproduction may be to initiate strobili more often, but limit the reproductive effort by constraining the sink activity of the most costly structures, in this case the number of mature seeds. The tree is not in full control of the percent seed set, as that is also under the control of pollinator behavior and population size. However, the maximum number of potential seeds within a strobilus is under the plant’s control. A reduction in the number of ovules per strobilus following the stress of the invasion could have been achieved by retaining the pre-invasion mean of five ovules per sporophyll, then reducing the number of sporophylls per megastrobilus. The female trees in Guam relied partly on this behavior (29% initial reduction), but these trees relied more heavily on reducing the number of ovules per sporophyll (60% initial reduction) as the means of reducing the total ovule count per strobilus. These findings may indicate a limited genetic control over the plasticity of sporophyll number per strobilus, and a greater genetic control over the plasticity of ovule number per sporophyll. These issues deserve further study.
These phenomena coalesce to indicate that even if conservation efforts or fortuitous events begin to partly mitigate the ongoing mortality of the surviving C. micronesica population, recovery of regeneration may not ensue without further research and intervention. Therefore, proactive conservation actions may be required to mitigate the sustained constraints on plant fecundity and the recruitment of long-lived seedlings. Our findings argue for an immediate focus on more research to inform these conservation efforts. We consider four avenues of immediate relevant research, which should be pursued in synchronicity to ensure that each agendum is cross-informed by the other agenda.
First, a lepidoptera taxonomist is urgently needed to clarify the identity of the pollinators. Cycad pollination mutualists are also typically consumers of male cone tissue.
Cycas micronesica is the only cycad species known to rely exclusively on a micro-lepidopteran cone borer, as other cycad species rely on beetles and thrips as their mutualist. Our morphology observations indicate that the
Anatrachyntis taxon found on Guam is the same as that found with the
C. micronesica population on Rota Island, but distinct from that found on Yap Island. However, the taxon has not been unambiguously identified at the species level for any of these three insular populations. This must be corrected. The Guam moth is likely a two-island endemic insect, and is at risk of co-extinction [
26]. The moth cannot be considered for listing under the Endangered Species Act if the binomial remains ambiguous, yet to date, no funds have been appropriated to correct this conservation emergency. Appropriating the services of a taxonomist to use every approach required to provide the binomial for the Guam, Rota, and Yap
Anatrachyntis populations is urgent. If listed, this animal could be provided new conservation support, and this would synergize the conservation support being provided to
C. micronesica.
Second, a source-sink physiologist is needed to better understand the nonstructural carbohydrate relations that define the timing and size of cycad growth events [
30,
31,
32]. Traditional knowledge of the production of starch from
Cycas revoluta Thunb. stem tissue exploited the fact that male tree stems contain more starch than female tree stems, and the greatest yield of starch can be obtained by harvesting immediately prior to a primary growth event [
30]. New
C. micronesica leaf and microstrobilus initiation and growth decreased the nonstructural carbohydrate content of subtending stems, confirming the reliance on stored stem resources for new growth [
32]. The depletion of nonstructural carbohydrates preceded
C. revoluta plant mortality during the
A. yasumatsui infestations [
33]. Guam’s chronic
A. yasumatsui infestations have created a situation where the unrelenting hemipteran feeding behaviors, coupled with the resource-depleting primary growth events of the host trees, appear to have caused the surviving trees to be functioning at a constant deficit of nonstructural resources. Indeed, the post-invasion
C. micronesica population on Guam has a reduced starch and sugar content of the stems [
34] and seed gametophytes [
35] as a result of the invasion. The deficit of resources stored in
C. micronesica organs is the probable cause of the reduced reproductive effort for the male and female trees, and the deficit in stored resources in the megagametophytes may partly explain the reduced germination and early seedling survival of the seeds harvested from scale-infested trees [
36]. The 4% of Guam’s pre-invasion population of
C. micronesica that remains alive [
4] is not thriving, but instead is in survival mode. A source-sink physiologist would be able to more fully understand how endogenous resource pools are involved in the ongoing constraints on plant survival, fecundity, regeneration, and recruitment.
Third, a plant anatomist will be required to understand the changes that have occurred to the stem apices. Pachycaulous cycad stems grow in diameter without the benefit of a secondary vascular cambium, and rely on a large primary thickening meristem (PTM) to construct most of the radial growth [
30]. Healthy arborescent cycad stems exhibit minimal change in diameter for the full vertical span of the stem when compared to lignophyte trees (
Figure 5a). A conspicuous decrease in the apical stem diameter of Guam’s surviving
C. micronesica trees has occurred in recent years as the trees have responded to the resource-limited growing conditions since the
A. yasumatsui invasion (
Figure 5b). A plant anatomist is needed to understand how this behavior has influenced the size of the PTM. To our knowledge, this is the first case study of a population-level response of an apparent reduction in PTM size as a result of chronic biotic threats. If a reduction in PTM size is confirmed by an anatomist, the reasons behind the 44% decline in height increment [
22], the reduced microstrobilus size [
20], and the reduced megastrobilus size (reported herein) may become better understood. Moreover, we have observed a recent change of leaf morphology and allometry, such that petiole length has increased, rachis length has decreased, and leaflet number has declined. These overall smaller leaves mimic juvenile leaf traits. We predict these reductions in leaf size may benefit the
C. micronesica population by reducing the drag coefficient of the canopies during future tropical cyclones. Clearly, a plant anatomist is urgently needed to interpret how this unusual apical stem behavior may influence the recovery of reproductive effort and fecundity, and how the resulting changes in leaf traits may influence tropical cyclone damage. What are the long-term consequences of this abnormal decrease in stem apex diameter? If the Guam population begins to recover in health due to conservation interventions, will the PTM grow to the size that characterized the trees prior to 2005? If so, will the stratum characterizing the 2005–2023 height growth remain constrained in diameter for the life of the tree, or will the tissues within this stratum be able to use diffuse mitotic activity to expand to the pre-invasion diameter?
Fourth, a pollination biologist will be required to manage female plant fecundity. Prior to the invasion, we confirmed that in many Guam habitats the C. micronesica seed set was rarely greater than 30%. This limited seed set in Guam is in sharp contrast to the 80% seed set and the 90+ seeds per strobilus we have measured in Yap (T.E.M., unpublished). These observations indicate that the Guam Anatrachyntis taxon may be less efficient as a pollinator than the Yap Anatrachyntis taxon, or that other herbivore threats on Guam interact with the pollinators in a manner that is absent on Yap. They also indicate that the Guam trees could probably support many more seeds per strobilus than occurs under non-stressful conditions. The sustained reduction in the natural seed set of the surviving Guam trees may be reversed by a pollination biologist using controlled pollinations. If a source-sink physiologist confirms the recovery of the nonstructural carbohydrate content of the surviving stems, these management actions may improve species recovery by increasing tree fecundity without stressing the trees.
The list of invasive herbivore threats to Guam’s
C. micronesica population is substantial [
37]. In addition to the numerous invasive herbivores, the stem borer
Acalolepta marianarum Aurivillius (Coleoptera: Cerambycidae) relies on
C. micronesica stems for ovipositioning and larval development [
1]. Irruptions of this native stem borer occurred after the
A. yasumatsui invasion, due to reductions in tree health from the unprecedented alien insect feeding behaviors [
38]. These developments exemplify how each herbivore possesses the potential to magnify the
A. yasumatsui damage into an invasional meltdown [
26]. Until adequate biological control organisms are successfully established on Guam, most alternative conservation actions, such as tree salvage and transplant projects, will probably not be successful [
39]. The transplanted trees will likely die during the years after the project is completed and deemed a success. The long list of biological threats also illuminates the urgency of funding the four recommended research thrusts, as continued delays in mitigating the invasive pest damage indicate that an irruption of any one of the herbivore threats could occur at any time, leading to an unprecedented increase in
C. micronesica mortality rates.
As discussed previously, both the male and female trees exhibited negative responses in reproductive behavior after the invasion. However, there are notable differences between the sexes in the recovery of some traits. These differential responses of the male versus female trees represent a distinct example of sexual dimorphism. Other examples of
C. micronesica sexual dimorphism have been reported. More branching among the male trees occurred when compared to the female trees [
40]. The frequency of strobilus production was greater for the male trees than for the female trees [
23,
28]. The height increment of the male trees exceeded that of the female trees [
22]. Based on branching patterns, more female trees were killed following the
A. yasumatsui invasion than male trees [
40]. The chronic
A. yasumatsui infestations reduced the height increment of the male trees to a greater extent than that of the female trees [
22]. Understanding these and other behaviors that portray forms of sexual dimorphism may aid conservationists in developing a more effective
C. micronesica recovery plan. Our results also indicate that cycads may be an ideal model species for studying how stressful conditions [
41] and other global change factors [
42] may influence diecious plant species in a manner that differs from hermaphrodite plant species.
We report herein an increase in female tree height throughout the years of this study, as we did for the male trees [
20]. This finding does not indicate there were recruitment increments into large size categories due to population-level growth events. Instead, this metric captures the fact that more of the smaller individuals of the host tree have died in response to the chronic
A. yasumatsui herbivory before the larger individuals have died [
3,
4]. Our annual search for appropriate megastrobili was random in this study, and as the insular population’s mean height increased as each year passed, the random sample of current-year megastrobili was conducted among the plant population that gradually increased in mean height.
This study adds to a growing body of literature documenting the multi-decadal changes to the
C. micronesica population. Reaching an adequate level of understanding of the threats to the recovery of an endangered tree species requires long-term research by knowledgeable scientists. When germane research is conducted, conservationists benefit from an adaptive management approach, wherein every year of decision-making is supported by more of the accumulated new knowledge. In addition to this paper, our sustained research has resulted in several other publications based on long-term studies [
4,
20,
22,
43].
Benchmarking is an important endeavor for establishing knowledge of how global change factors influence biodiversity [
44,
45]. During an era of rapid change, if the research community fails to characterize various organism and population traits before substantial changes ensue, the opportunity to benchmark could be lost. Our pre-invasion megastrobili traits reported herein serve as one more benchmark for future conservationists to determine if the
C. micronesica population will ever recover to a healthy pre-invasion status. We have previously reported that the pre-invasion height increment behaviors [
22] and the pre-invasion microstrobili traits [
20] may also serve as benchmarks for future
C. micronesica conservation assessments of species recovery.
The threats causing tree extinctions are occurring on a global scale. The most common threat to tree conservation is habitat conversion for various uses, such as agriculture or urban development [
46]. Our case study is an example of an alternative threat to trees, that of invasive herbivorous insects. As new invasions begin to threaten more tree species globally, our study may serve as an example to inform the mitigation efforts of future tree species extinction threats. Plant mortality during the initial three to four years after the invasion was so rapid [
3,
4], the conservation community was in triage mode without any local research to inform decisions. However, the research since those early years has been substantial, the greatest conservation intervention needs (biological control) are understood, and published recommendations from scientists with germane expertise have been numerous.
The constraints on the regeneration and recruitment of
C. micronesica are consequential, but the direct threat of mature plant mortality remains the greatest threat to this gymnosperm tree. The implementation of a coalition of biological control organisms to reduce
A. yasumatsui damage was the greatest
C. micronesica conservation need as soon as the 2003 invasion occurred, was the greatest need when the host species was added to the IUCN Red List in 2006 [
5], was the greatest need when it was listed under the ESA in 2015 [
6], and remains the greatest conservation need today. The introduction and successful establishment of the armored scale predator
Rhyzobius lophanthae Blaisdell (Coleoptera, Coccinellidae) in 2005 [
47] is the probable reason for the surviving
C. micronesica plants today. However, there are numerous limitations of this predator that do not allow the adequate control of
A. yasumatsui (reviewed in [
37,
48]). We concur with previous recommendations [
22,
37,
39,
48] that the funding of a sustained, dedicated effort to establish a coalition of biological control organisms has been and continues to be the greatest conservation action priority needed to enable species recovery. The antecedent few years have been characterized by unprecedented declines in
A. yasumatsui infestations, indicating that a fortuitous irruption of some biological control organism has occurred on Guam. Three years ago we recommended with urgency that the engaged conservation community should enact a dedicated effort to identify this organism [
37]. To our knowledge, this has not occurred to date. This new knowledge is urgently needed for species recovery for two reasons. First, there is no reason for Guam conservationists to assume that this unknown biological control organism will sustain the future biological control of
A. yasumatsui, and its precise identity will be required to monitor this in the future. Second, if a fortuitous invasion of Guam by an unprecedented biological control organism has indeed occurred, this organism needs to be transported and released on Rota to conserve the
C. micronesica population there. The island of Rota was invaded by
A. yasumatsui in 2007 [
2], and the host
C. micronesica population experienced devastation similar to that of Guam (T.E.M., unpublished data). A decline in the
A. yasumatsui herbivory has also occurred recently on Rota Island (T.E.M., personal observations). There is no reason to believe the fortuitous biological control on Rota is due to the same organism exhibiting control on Guam. A survey of what is controlling
A. yasumatsui on Rota is warranted, and if identified, that organism needs to be transported and released on Guam.
We conclude with three caveats. First, we used a fixed geographic range on the northeast coast of Guam for this study. We do not assume these same trends have occurred in other regions of Guam or in Rota, where all
C. micronesica plants have been threatened by
A. yasumatsui. Second, the recent fortuitous control of
A. yasumatsui may be a turning point after which plant mortality will subside. If this does occur, then we predict that increases in stem apex, microstrobilus, and megastrobilus size will occur in the future in response to this turning point. The direct documentation of these changes by funded, competent scientists who understand cycad biology is warranted. Third, Guam is located within the most active tropical cyclone zone globally [
49]. The island has not experienced direct landfall of an intense tropical cyclone since the 2003
A. yasumatsui invasion. Therefore, there are no historical observations that could inform how this form of large-scale abiotic disturbance will interact with the continuing demise of the island’s cycad population caused by insidious biotic threats. The ongoing changes that we have reported for male and female
C. micronesica tree behaviors may be altered in unique ways when the next direct landfall of a tropical cyclone damages the vegetation of the island. Indeed, natural disturbances may profoundly influence the impact of invasive species [
50]. The conservation community will need an embedded plant ecologist with knowledge of tropical cyclones to determine the consequences to species recovery when this occurs.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




