
Goal-Directed Travel in the Nocturnal Javan Slow Loris (Nycticebus javanicus)

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Species
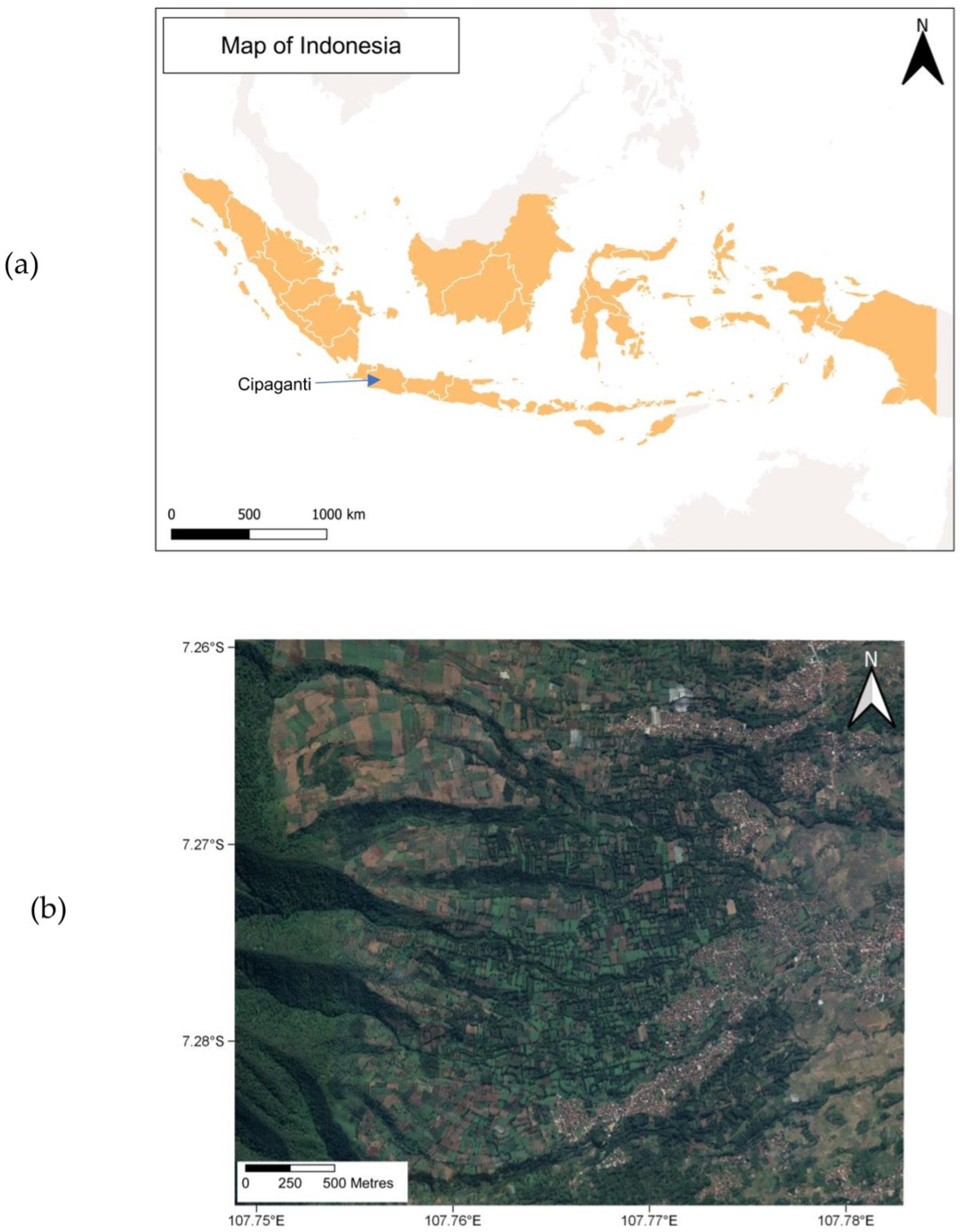
2.2. Study Site and Data Collection
2.3. Data Analysis
3. Results
3.1. Nightly Route Length
3.2. Visited Tree Diversity
3.3. Revisited Gum-Producing Trees
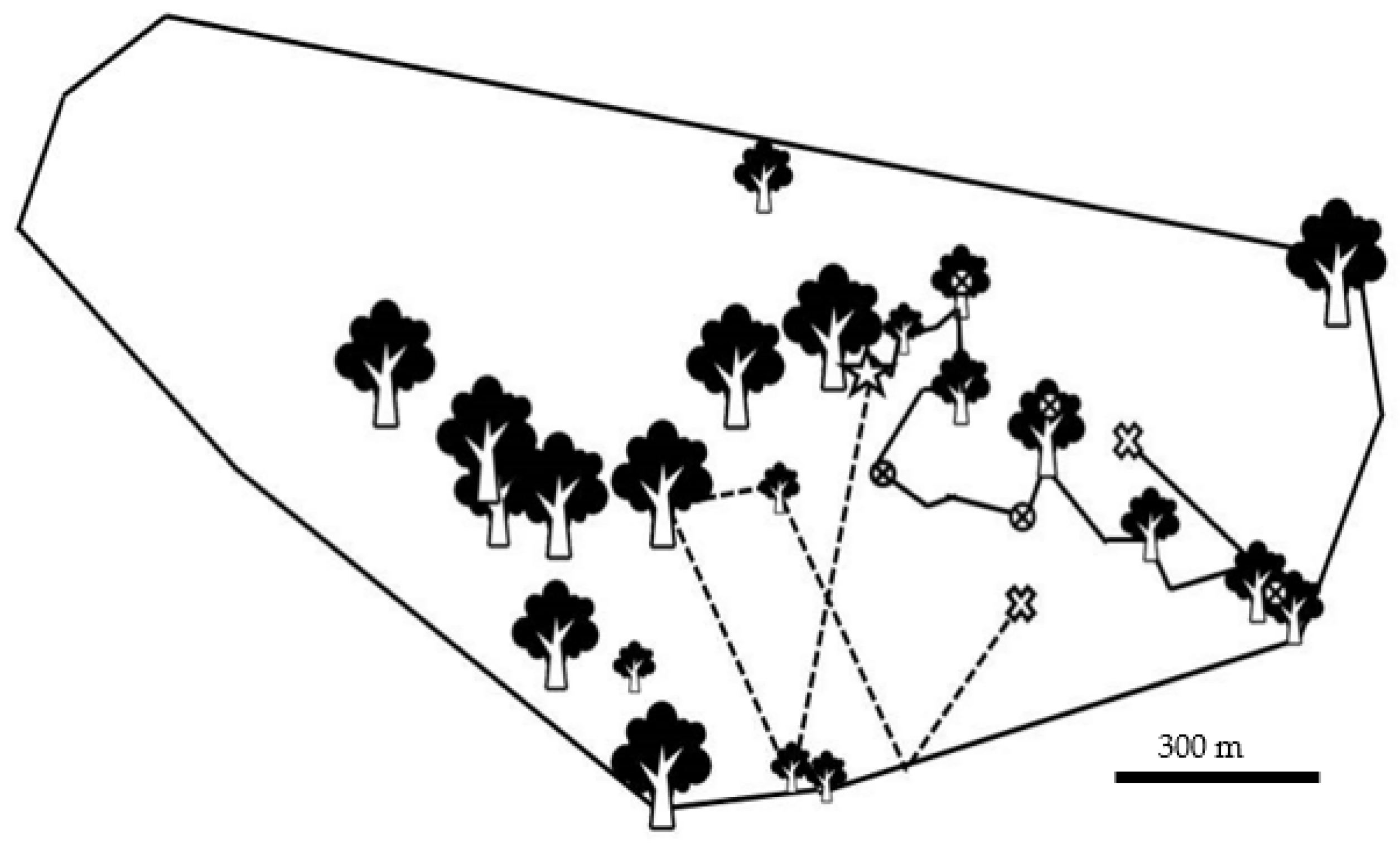
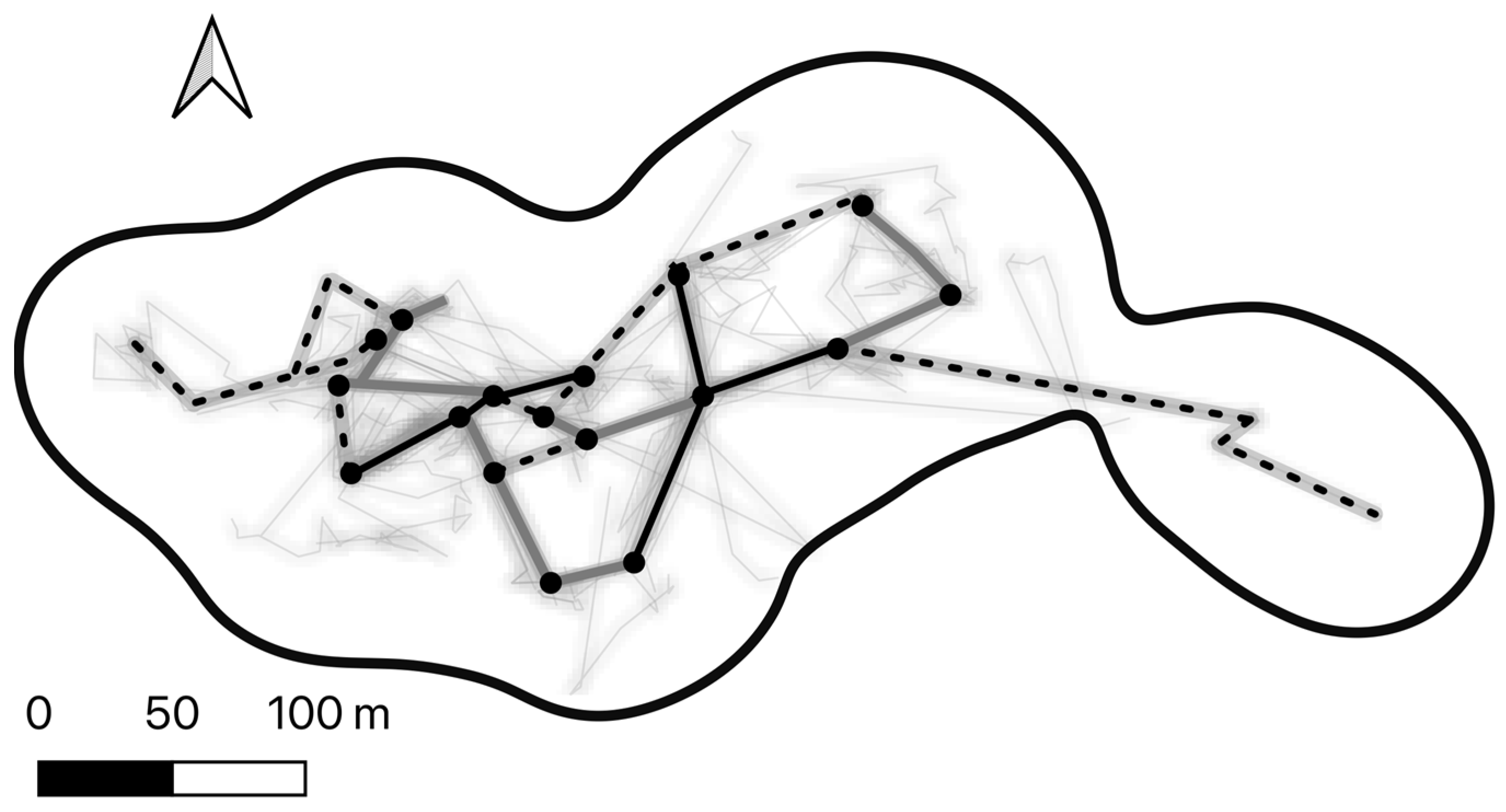
3.4. Directional Changes and Route Overlap
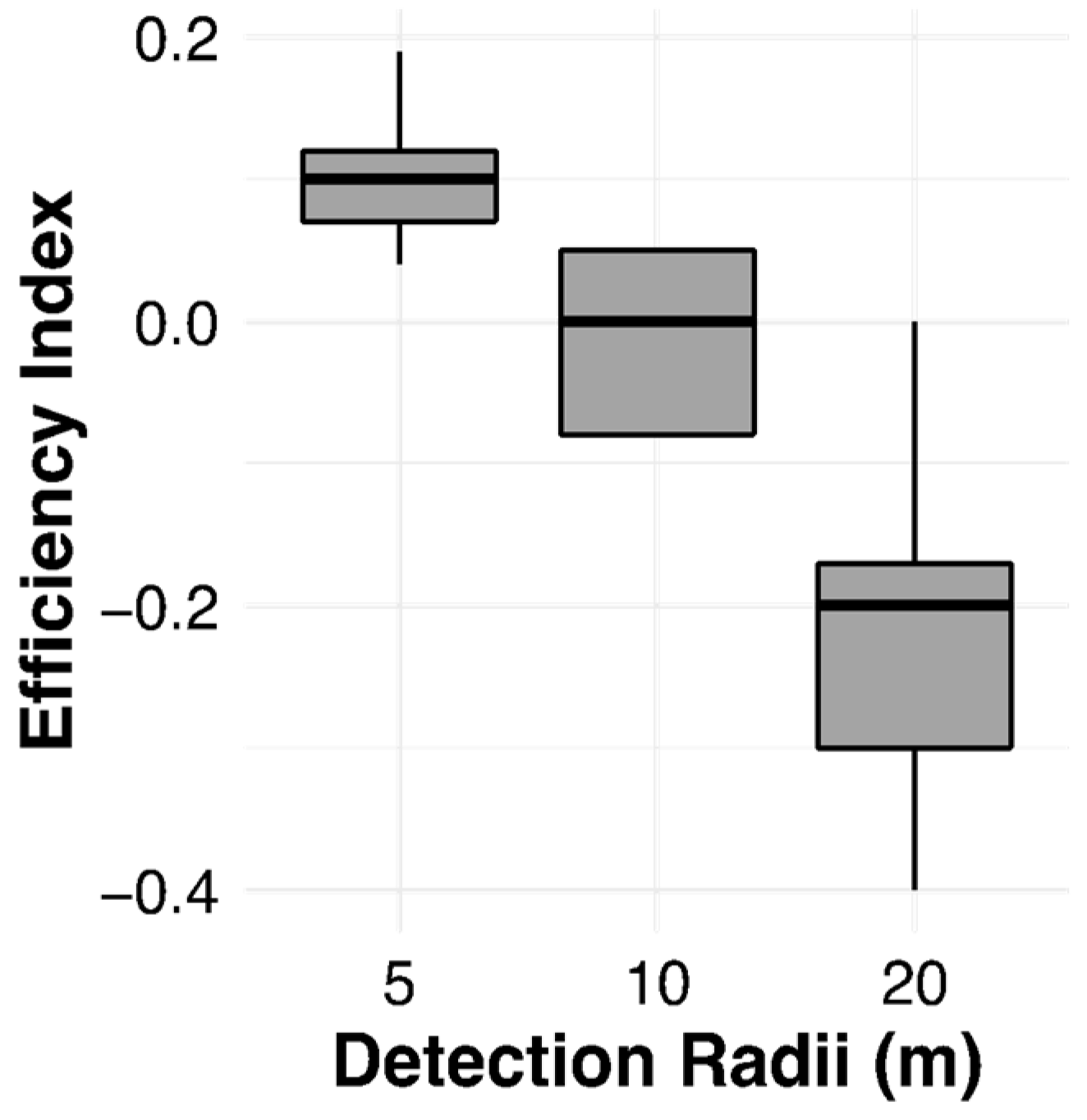
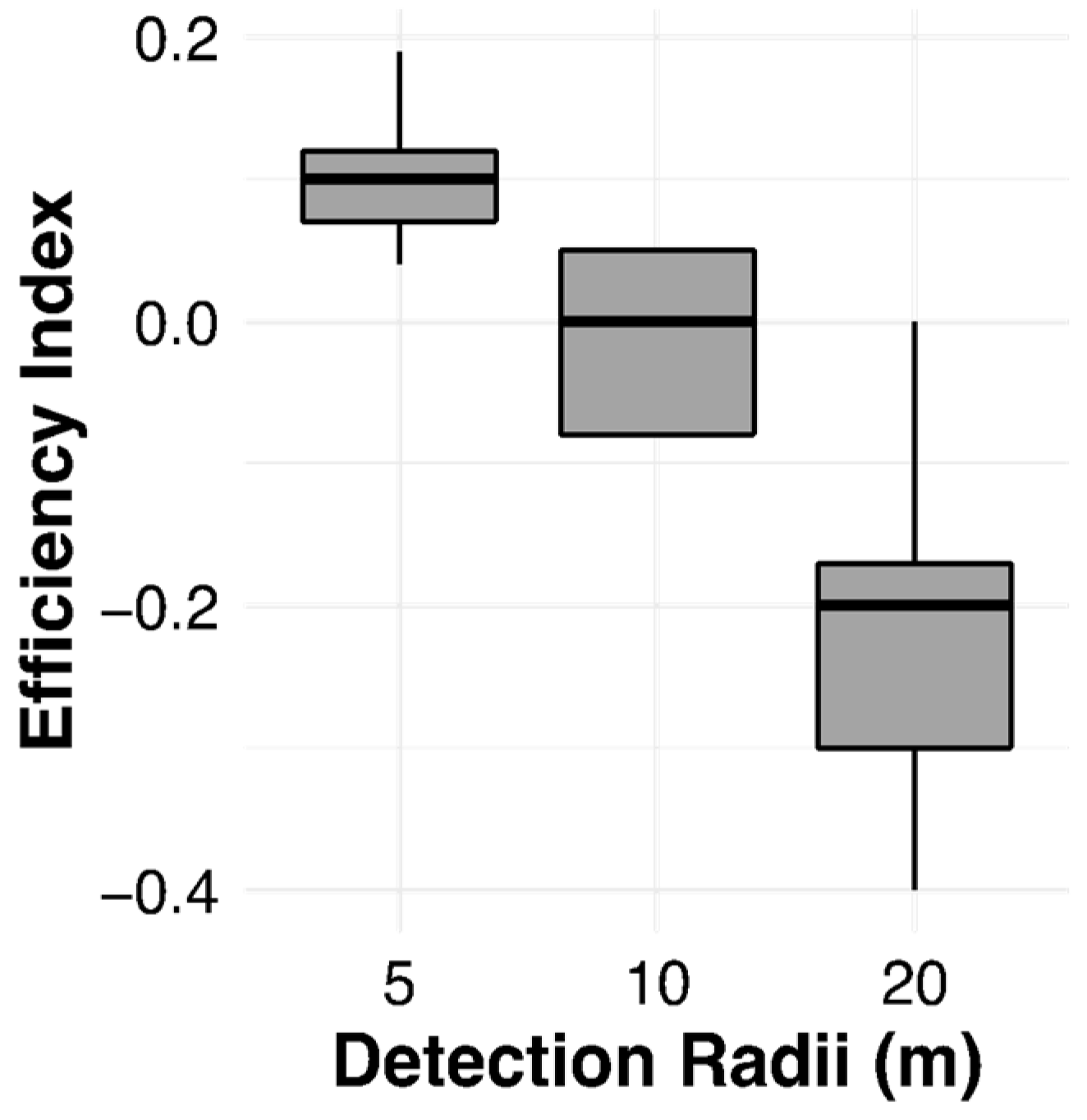
3.5. The Efficiency Index (EI)
4. Discussion
5. Translocation Implications
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Di Fiore, A.; Suarez, S.A. Route-based travel and shared routes in sympatric spider and woolly monkeys: Cognitive and evolutionary implications. Anim. Cogn. 2007, 10, 317–329. [Google Scholar] [CrossRef]
- Mueller, T.; Fagan, W.F. Search and navigation in dynamic environments–from individual behaviors to population distributions. Oikos 2008, 117, 654–664. [Google Scholar] [CrossRef]
- Lührs, M.L.; Dammhahn, M.; Kappeler, P.M.; Fichtel, C. Spatial memory in the grey mouse lemur (Microcebus murinus). Anim. Cogn. 2009, 12, 599–609. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, M.E. Mantled howler monkey spatial foraging decisions reflect spatial and temporal knowledge of resource distributions. Anim. Cogn. 2016, 19, 387–403. [Google Scholar] [CrossRef] [PubMed]
- Sanders, A.E.; Holtzer, R.; Lipton, R.B.; Hall, C.; Verghese, J. Egocentric and exocentric navigation skills in older adults. J. Gerontol. A Biol. Sci. Med. Sci. 2008, 63, 1356–1363. [Google Scholar] [CrossRef] [PubMed]
- Garber, P.A.; Dolins, F.L. Primate spatial strategies and cognition: Introduction to this special issue. Am. J. Primatol. 2014, 76, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Hills, T.T.; Butterfill, S. From foraging to autonoetic consciousness: The primal self as a consequence of embodied prospective foraging. Curr. Zool. 2015, 61, 368–381. [Google Scholar] [CrossRef]
- Gautestad, A.O.; Mysterud, A. The Lévy flight foraging hypothesis: Forgetting about memory may lead to false verification of Brownian motion. Mov. Ecol. 2013, 1, 9. [Google Scholar] [CrossRef]
- Byrne, R.W. How monkeys Find their way. Leadership, coordination, and cognitive maps of African baboons. In On the Move: How and Why Animals Travel in Groups; Boinski, S.B., Garber, P.A., Eds.; University of Chicago Press: Chicago, IL, USA, 2000; pp. 491–518. [Google Scholar]
- Garber, P.A. Evidence for the use of spatial, temporal and social information by some primate foragers. In On the Move: How and Why Animals Travel in Groups; Boinski, S.B., Garber, P.A., Eds.; University of Chicago Press: Chicago, IL, USA, 2000; pp. 261–298. [Google Scholar]
- Piaget, J.; Inhelder, B. The Child’s Concept of Space; Routledge & Paul: London, UK, 1956. [Google Scholar]
- Normand, E.; Boesch, C. Sophisticated Euclidean maps in forest chimpanzees. Anim. Behav. 2009, 77, 1195–1201. [Google Scholar] [CrossRef]
- Watkins, B.; de Guinea, M.; Poindexter, S.A.; Ganzhorn, J.U.; Donati, G.; Eppley, T.M. Routes matter: The effect of seasonality on bamboo lemur navigational strategies. Anim. Behav. 2022, 186, 137–149. [Google Scholar] [CrossRef]
- Presotto, A.; Izar, P. Spatial reference of black capuchin monkeys in Brazilian Atlantic Forest: Egocentric or allocentric? Anim. Behav. 2010, 80, 125–132. [Google Scholar] [CrossRef]
- Asensio, N.; Brockelman, W.Y.; Malaivijitnond, S.; Reichard, U.H. Gibbon travel routes are goal oriented. Anim. Cogn. 2011, 14, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Rolland, E.; Trull, S. Spatial mapping memory: Methods used to determine the existence and type of cognitive maps in arboreal mammals. Mammal Rev. 2022, 52, 96–111. [Google Scholar] [CrossRef]
- Trapanese, C.; Meunier, H.; Masi, S. What, where and when: Spatial foraging decisions in primates. Biol. Rev. 2019, 94, 483–502. [Google Scholar] [CrossRef]
- Green, S.J.; Boruff, B.J.; Grueter, C.C. From ridge tops to ravines: Landscape drivers of chimpanzee ranging patterns. Anim. Behav. 2020, 163, 51–60. [Google Scholar] [CrossRef]
- Green, S.J.; Boruff, B.J.; Bonnell, T.R.; Grueter, C.C. Chimpanzees use least-cost routes to out-of-sight goals. Curr. Biol. 2020, 30, 4528–4533. [Google Scholar] [CrossRef]
- Normand, E.; Ban, S.D.; Boesch, C. Forest chimpanzees (Pan troglodytes verus) remember the location of numerous fruit trees. Anim. Cogn. 2009, 12, 797–807. [Google Scholar] [CrossRef]
- Menzel, C.R.; Savage-Rumbaugh, E.S.; Menzel, E.W. Bonobo (Pan paniscus) spatial memory and communication in a 20-hectare forest. Int. J. Primatol. 2002, 23, 601–619. [Google Scholar] [CrossRef]
- Salmi, R.; Presotto, A.; Scarry, C.J.; Hawman, P.; Doran-Sheehy, D.M. Spatial cognition in western gorillas (Gorilla gorilla): An analysis of distance, linearity, and speed of travel routes. Anim. Cogn. 2020, 23, 545–557. [Google Scholar] [CrossRef]
- Erhart, E.M.; Overdorff, D.J. Group leadership and feeding priority in wild Propithecus diadema edwardsi and Eulemur fulvus rufus. Am. J. Primatol. 1998, 45, 178–179. [Google Scholar]
- Joly, M.; Zimmermann, E. First evidence for relocation of stationary food resources during foraging in a strepsirhine primate (Microcebus murinus). Am. J. Primatol. 2007, 69, 1045–1052. [Google Scholar] [CrossRef] [PubMed]
- Joly, M.; Zimmermann, E. Do solitary foraging nocturnal mammals plan their routes? Biol. Lett. 2011, 7, 638–640. [Google Scholar] [CrossRef] [PubMed]
- Teichroeb, J.A.; Vining, A.Q. Navigation strategies in three nocturnal lemur species: Diet predicts heuristic use and degree of exploratory behavior. Anim. Cogn. 2019, 22, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Nekaris, K.A.I.; Poindexter, S.; Reinhardt, K.D.; Sigaud, M.; Cabana, F.; Wirdateti, W.; Nijman, V. Coexistence between Javan Slow Lorises (Nycticebus javanicus) and humans in a dynamic agroforestry landscape in west Java, Indonesia. Int. J. Primatol. 2017, 38, 303–320. [Google Scholar] [CrossRef]
- Nekaris, K.A.I. Extreme primates: Ecology and evolution of Asian lorises. Evol. Anthropol. Issues News Rev. 2014, 23, 177–187. [Google Scholar] [CrossRef]
- Cabana, F.; Dierenfeld, E.S.; Wirdateti, W.; Donati, G.; Nekaris, K.A.I. Exploiting a readily available but hard to digest resource: A review of exudativorous mammals identified thus far and how they cope in captivity. Integr. Zool. 2018, 13, 94–111. [Google Scholar] [CrossRef]
- Poindexter, S.A.; Nekaris, K.A.I. Vertical clingers and gougers: Rapid acquisition of adult limb proportions facilitates feeding behaviours in young Javan slow lorises (Nycticebus javanicus). Mammal Biol. 2017, 87, 40–49. [Google Scholar] [CrossRef]
- Reinhardt, K.D.; Wirdateti, W.; Nekaris, K.A.I. Climate-mediated activity of the Javan Slow Loris, Nycticebus javanicus. AIMS Environ. Sci. 2016, 3, 249–260. [Google Scholar] [CrossRef]
- Karimloo, L.; Campera, M.; Imron, M.A.; Rakholia, S.; Mehta, A.; Hedger, K.; Nekaris, K.A.I. Habitat Use, Terrestriality and Feeding Behaviour of Javan Slow Lorises in Urban Areas of a Multi-Use Landscape in Indonesia. Land 2023, 12, 1349. [Google Scholar] [CrossRef]
- Rode-Margono, E.J.; Nijman, V.; Wirdateti, N.K.; Nekaris, K.A.I. Ethology of the critically endangered Javan slow loris Nycticebus javanicus E. Geoffroy Saint-Hilaire in West Java. Asian Prim. 2014, 4, 27–41. [Google Scholar]
- Altmann, J. Observational study of behavior: Sampling methods. Behaviour 1974, 49, 227–266. [Google Scholar] [CrossRef] [PubMed]
- Nekaris, K.A.I. Activity budget and positional behavior of the Mysore slender loris (Loris tardigradus lydekkerianus): Implications for slow climbing locomotion. Folia Primatol. 2001, 72, 228–241. [Google Scholar] [CrossRef] [PubMed]
- Byrne, R.W.; Noser, R.; Bates, L.A.; Jupp, P.E. How did they get here from there? Detecting changes of direction in terrestrial ranging. Anim. Behav. 2009, 77, 619–631. [Google Scholar] [CrossRef]
- Janson, C.H. Experimental evidence for spatial memory in foraging wild capuchin monkeys, Cebus apella. Anim. Behav. 1998, 55, 1229–1243. [Google Scholar] [CrossRef]
- Finnerty, P.B.; Stutz, R.S.; Price, C.J.; Banks, P.B.; McArthur, C. Leaf odour cues enable non-random foraging by mammalian herbivores. J. Anim. Ecol. 2017, 86, 1317–1328. [Google Scholar] [CrossRef]
- Kane, A.; Kendall, C.J. Understanding how mammalian scavengers use information from avian scavengers: Cue from above. J. Anim. Ecol. 2017, 86, 837–846. [Google Scholar] [CrossRef]
- Stutz, R.S.; Croak, B.M.; Proschogo, N.; Banks, P.B.; McArthur, C. Olfactory and visual plant cues as drivers of selective herbivory. Oikos 2017, 126, 259–268. [Google Scholar] [CrossRef]
- Cunningham, E.; Janson, C. Integrating information about location and value of resources by white-faced saki monkeys (Pithecia pithecia). Anim. Cogn. 2007, 10, 293–304. [Google Scholar] [CrossRef]
- Dominy, N.J.; Ross, C.F.; Smith, T.D. Evolution of the special senses in primates: Past, present, and future. Anat. Rec. Part A 2004, 281, 1078–1082. [Google Scholar] [CrossRef]
- de Guinea, M.; Estrada, A.; Nekaris, K.; Van Belle, S. Arboreal route navigation in a Neotropical mammal: Energetic implications associated with tree monitoring and landscape attributes. Mov. Ecol. 2019, 7, 39. [Google Scholar] [CrossRef]
- Janmaat, K.R.; Ban, S.D.; Boesch, C. Chimpanzees use long-term spatial memory to monitor large fruit trees and remember feeding experiences across seasons. Anim. Behav. 2013, 86, 1183–1205. [Google Scholar] [CrossRef]
- Cabana, F.; Dierenfeld, E.; Wirdateti, W.; Donati, G.; Nekaris, K.A.I. Trialling nutrient recommendations for slow lorises (Nycticebus spp.) based on wild feeding ecology. J. Anim. Physiol. Anim. Nutr. 2018, 102, e1–e10. [Google Scholar] [CrossRef] [PubMed]
- Berger-Tal, O.; Bar-David, S. Recursive movement patterns: Review and synthesis across species. Ecosphere 2015, 6, 1–12. [Google Scholar] [CrossRef]
- Bracis, C.; Bildstein, K.L.; Mueller, T. Revisitation analysis uncovers spatio-temporal patterns in animal movement data. Ecography 2018, 41, 1801–1811. [Google Scholar] [CrossRef]
- Porter, L.M.; Garber, P.A. Foraging and spatial memory in wild Weddell’s saddleback tamarins (Saguinus fuscicollis weddelli) when moving between distant and out-of-sight goals. Int. J. Primatol. 2013, 28, 1035–1058. [Google Scholar] [CrossRef]
- McLean, K.A.; Trainor, A.M.; Asner, G.P.; Crofoot, M.C.; Hopkins, M.E.; Campbell, C.J.; Jansen, P.A. Movement patterns of three arboreal primates in a Neotropical moist forest explained by LiDAR-estimated canopy structure. Landsc. Ecol. 2016, 31, 1849–1862. [Google Scholar] [CrossRef]
- De Raad, A.L.; Hill, R.A. Topological spatial representation in wild chacma baboons (Papio ursinus). Anim. Cogn. 2019, 22, 397–412. [Google Scholar] [CrossRef]
- Cabana, F.; Dierenfeld, E.; Wirdateti, W.; Donati, G.; Nekaris, K.A.I. The seasonal feeding ecology of the Javan slow loris (Nycticebus javanicus). Am. J. Phys. Anthropol. 2017, 162, 768–781. [Google Scholar] [CrossRef]
- Collins, R.; Nekaris, K.A.I. Release of greater slow lorises, confiscated from the pet trade, to Batutegi Protected Forest, Sumatra, Indonesia. In Global Re-Introduction Perspectives; IUCN Reintroduction Specialist Group: Abu Dhabi, United Arab Emirates, 2008; pp. 192–195. [Google Scholar]
- Kenyon, M.; Streicher, U.; Loung, H.; Tran, T.; Tran, M.; Vo, B.; Cronin, A. Survival of reintroduced pygmy slow loris Nycticebus pygmaeus in South Vietnam. Endanger Species Res. 2014, 25, 185–195. [Google Scholar] [CrossRef]
- Fuller, G.; Eggen, W.F.; Wirdateti, W.; Nekaris, K.A.I. Welfare impacts of the illegal wildlife trade in a cohort of confiscated greater slow lorises, Nycticebus coucang. J. Appl. Anim. Wel. Sci. 2018, 21, 224–238. [Google Scholar] [CrossRef]
- Streicher, U.; Nadler, T. Re-introduction of pygmy lorises in Vietnam. Reintroduction News 2003, 23, 37–40. [Google Scholar]
- Poindexter, S.; Khoa, D.; Nekaris, K. Ranging patterns of reintroduced pygmy slow lorises (Nycticebus pygmaeus) in Cuc Phuong National Park, Vietnam. Vietnam. J. Primatol. 2017, 2, 37–49. [Google Scholar]
- Moore, R.S.; Nekaris, K.A.I. Compassionate conservation, rehabilitation and translocation of Indonesian slow lorises. Endanger Species Res. 2014, 26, 93–102. [Google Scholar] [CrossRef]
- Campera, M.; Brown, E.; Imron, M.A.; Nekaris, K.A.I. Unmonitored releases of small animals? The importance of considering natural dispersal, health, and human habituation when releasing a territorial mammal threatened by wildlife trade. Bioll. Conserv. 2020, 242, 108404. [Google Scholar] [CrossRef]
- Minckley, W.L. Translocation as a tool for conserving imperiled fishes: Experiences in western United States. Biol. Conser. 1995, 72, 297–309. [Google Scholar] [CrossRef]
- IUCN Species Survival Commission (SSC). Guidelines for Reintroductions and Other Conservation Translocations; IUCN: Gland, Switzerland, 1989; p. 57. [Google Scholar]
- Griffith, B.; Scott, J.M.; Carpenter, J.W.; Reed, C. Translocation as a species conservation tool: Status and strategy. Science 1989, 245, 477–480. [Google Scholar] [CrossRef]
- Wolf, C.M.; Garland, T., Jr.; Griffith, B. Predictors of avian and mammalian translocation success: Reanalysis with phylogenetically independent contrasts. Biol. Conserv. 1998, 86, 243–255. [Google Scholar] [CrossRef]
- Berger-Tal, O.; Blumstein, D.T.; Swaisgood, R.R. Conservation translocations: A review of common difficulties and promising directions. Anim. Conserv. 2020, 23, 121–131. [Google Scholar] [CrossRef]
- Tuberville, T.D.; Clark, E.E.; Buhlmann, K.A.; Gibbons, J.W. Translocation as a conservation tool: Site fidelity and movement of repatriated gopher tortoises (Gopherus polyphemus). In Animal Conservation Forum; Cambridge University Press: Cambridge, UK, 2005; Volume 8, pp. 349–358. [Google Scholar]
- Ebrahimi, M.; Bull, C.M. Determining the success of varying short-term confinement time during simulated translocations of the endangered pygmy bluetongue lizard (Tiliqua adelaidensis). Amphibia-Reptilia 2013, 34, 31–39. [Google Scholar] [CrossRef]
- Kenward, R.E.; Hodder, K.H. Red squirrels (Sciurus vulgaris) released in conifer woodland: The effects of source habitat, predation and interactions with grey squirrels (Sciurus carolinensis). J. Zool. 1998, 244, 23–32. [Google Scholar] [CrossRef]
- Le Gouar, P.; Mihoub, J.B.; Sarrazin, F. Dispersal and habitat selection: Behavioural and spatial constraints for animal translocations. In Reintroduction Biology: Integrating Science and Management; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2012; pp. 138–164. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HR (KDE) | HR (MCP) | Full-Night | Half-Night | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| N | Mean | Max | Min | N | Mean | Max | Min | ||||
| Male | AC | 3.27 | 12.4 | - | - | - | - | 6 | 250.6 | 686.1 | 107.9 |
| AL | 6.6 | 9.5 | 5 | 733.2 | 1121.7 | 485.5 | 25 | 275.8 | 1582.6 | 73.2 | |
| AZ | 5.0 | 8.0 | 6 | 417.8 | 684.6 | 133.6 | 12 | 190.4 | 307.1 | 96.3 | |
| FE | 12.7 | 14.4 | 3 | 668.1 | 811.4 | 414.6 | 20 | 257.5 | 627.7 | 93.7 | |
| RA | 6.2 | 20.7 | 1 | 516.5 | 516.5 | 516.5 | 17 | 192.5 | 317.9 | 61 | |
| TO | 4.5 | 9.3 | 4 | 317.9 | 377 | 289 | 13 | 189.9 | 355.3 | 63.1 | |
| Female | EN | 3.6 | 7.7 | 2 | 578.6 | 731.9 | 425.3 | 7 | 332.2 | 471.9 | 198.5 |
| LU | 6.3 | 9.73 | 4 | 513.5 | 695.8 | 328.4 | 20 | 162.3 | 406.2 | 23.3 | |
| MA | 3.6 | 6.4 | 6 | 626.4 | 1264 | 360.9 | 14 | 253.4 | 511.7 | 83.6 | |
| OE | 4.1 | 5.4 | 1 | 428.2 | 428.2 | 428.2 | 12 | 202.7 | 418.4 | 73 | |
| SH | 2.4 | 3.8 | 7 | 613.9 | 871.8 | 400.5 | 14 | 238 | 401.7 | 113 | |
| SI | 4.12 | 11.0 | - | - | - | - | 11 | 328 | 523.6 | 156.8 | |
| TE | 4.9 | 6.5 | 5 | 555.7 | 968 | 380.2 | 21 | 205 | 450.9 | 33.6 | |
| Total | 541.4 | 239.4 | |||||||||
| Sex | ID | Number of CP | Route Overlap | # Unique Sites (Revisited) | Percentage Revisited |
|---|---|---|---|---|---|
| Female | LU | 14 | 35% | 37 (28) | 76% |
| TE | 13 | 44% | 60 (29) | 48% | |
| MA | 11 | 32% | 38 (24) | 63% | |
| EN | 10 | 30% | 21 (9) | 43% | |
| SI | 9 | 25% | 36 (15) | 42% | |
| OE | 8 | 39% | 34 (17) | 50% | |
| SH | 5 | 22% | 35 (15) | 49% | |
| Male | AL | 18 | 37% | 40 (32) | 80% |
| FE | 15 | 19% | 67 (44) | 66% | |
| RA | 12 | 18% | 25 (15) | 60% | |
| TO | 11 | 16% | 35 (18) | 52% | |
| AZ | 8 | 28% | 29 (13) | 45% | |
| AC | 6 | 32% | 14 (5) | 36% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poindexter, S.A.; Nijman, V.; Imron, M.A.; Nekaris, K.A.-I. Goal-Directed Travel in the Nocturnal Javan Slow Loris (Nycticebus javanicus). Ecologies 2023, 4, 568-579. https://doi.org/10.3390/ecologies4030037
Poindexter SA, Nijman V, Imron MA, Nekaris KA-I. Goal-Directed Travel in the Nocturnal Javan Slow Loris (Nycticebus javanicus). Ecologies. 2023; 4(3):568-579. https://doi.org/10.3390/ecologies4030037
Chicago/Turabian StylePoindexter, Stephanie A., Vincent Nijman, Muhammed Ali Imron, and K. Anne-Isola Nekaris. 2023. "Goal-Directed Travel in the Nocturnal Javan Slow Loris (Nycticebus javanicus)" Ecologies 4, no. 3: 568-579. https://doi.org/10.3390/ecologies4030037
APA StylePoindexter, S. A., Nijman, V., Imron, M. A., & Nekaris, K. A.-I. (2023). Goal-Directed Travel in the Nocturnal Javan Slow Loris (Nycticebus javanicus). Ecologies, 4(3), 568-579. https://doi.org/10.3390/ecologies4030037