1. Introduction
Proper thyroid function and thyroid hormone regulation are critical for organismal health as thyroid hormones affect nearly every organ system of the body. Thyroid-stimulating hormone (TSH), or thyrotropin, is the primary hormone stimulating thyroid hormone released from the thyroid gland. Produced by the anterior pituitary in response to thyrotropin-releasing hormone (TRH), TSH stimulates the release of triiodothyronine (T3) and thyroxine (T4) from thyroid follicular cells. The metabolically active T3 is primarily responsible for regulation of growth, development, and metabolism through genomic and non-genomic effects on numerous biochemical pathways. T3 is metabolically active in its free (unbound) form and the availability of free T3 is regulated both by its rate of conversion from the metabolically less active T4 and the degree to which it is bound to the proteins thyroxine-binding globulin, transthyretin, and albumin. In addition, regulation can be achieved by converting T4 into reverse T3 (rT3), which is commonly accepted as an end product of T4 degradation and which can weakly compete with T3 in binding to T3 thyroid nuclear hormone receptors. Given the importance of thyroid hormone to organismal health, the study of thyroid hormone production and regulation has received extensive attention within the field of human medicine.
The bottlenose dolphin (
Tursiops truncatus) is a fully-aquatic mammal that has undergone dramatic physiological adaptations to the thermoregulatory and diving challenges of an aquatic existence. It is also the most commonly held cetacean at marine parks, zoos, and aquaria, establishing a need to understand its normal physiological function for purposes of veterinary care and animal welfare. Thyroid hormones in bottlenose dolphins presumably support the same metabolic functions as observed in humans and other terrestrial mammals, yet there are distinct differences that suggest possible alterations in metabolic regulation and functionality that require understanding in order to facilitate the diagnosis and treatment of thyroid-based metabolic disease. Most notably, rT3 in dolphins (and other odontocetes) occurs at levels that far exceed those observed in humans and other terrestrial mammals [
1,
2,
3], although the reason for the high rT3 levels has not yet been determined. Levels of total thyroxine (tT4), the sum of free and protein-bound forms, also commonly fall within the high-end of the normal range for humans: >90 nM, [
4,
5]. Given these distinctions, there remains little information on the normal variation of thyroid hormones in dolphins and how they are affected by intrinsic (e.g., sex, age, reproductive status) and extrinsic (e.g., seasons, temperature, light cycles) factors. To date, thyroid hormone measurements in bottlenose dolphins have come from wild-caught dolphins [
2,
6] and those held under human care [
2,
7], yet differences in potential environmental factors and intrinsic factors make it difficult to confidently establish thyroid hormone levels and patterns for clinical use.
One approach to assessing thyroid function is through stimulation of the hypothalamic-pituitary-thyroid axis, either through the administration of TRH to stimulate TSH release from the anterior pituitary, or administration of TSH to directly stimulate the production and release of T3 and T4 from the thyroid gland. TSH stimulation tests have been used in belugas (
Delphinapterus leucas) and dolphins [
1,
7,
8]. These studies were limited to one or a few subjects and, in some cases, were performed in wild-caught animals with potentially unknown confounding variables (e.g., underlying health conditions, effects of handling stress). The study reported here builds on a prior TSH stimulation test conducted in a bottlenose dolphin [
7] in order to advance the understanding of dolphin thyroid function and the subsequent dynamics of circulating thyroid hormones post-stimulation. The study increases the sample size of subjects to bolster confidence in the observed hormonal response in hopes of advancing the procedure as a clinical tool in the veterinary care of the bottlenose dolphin.
3. Results
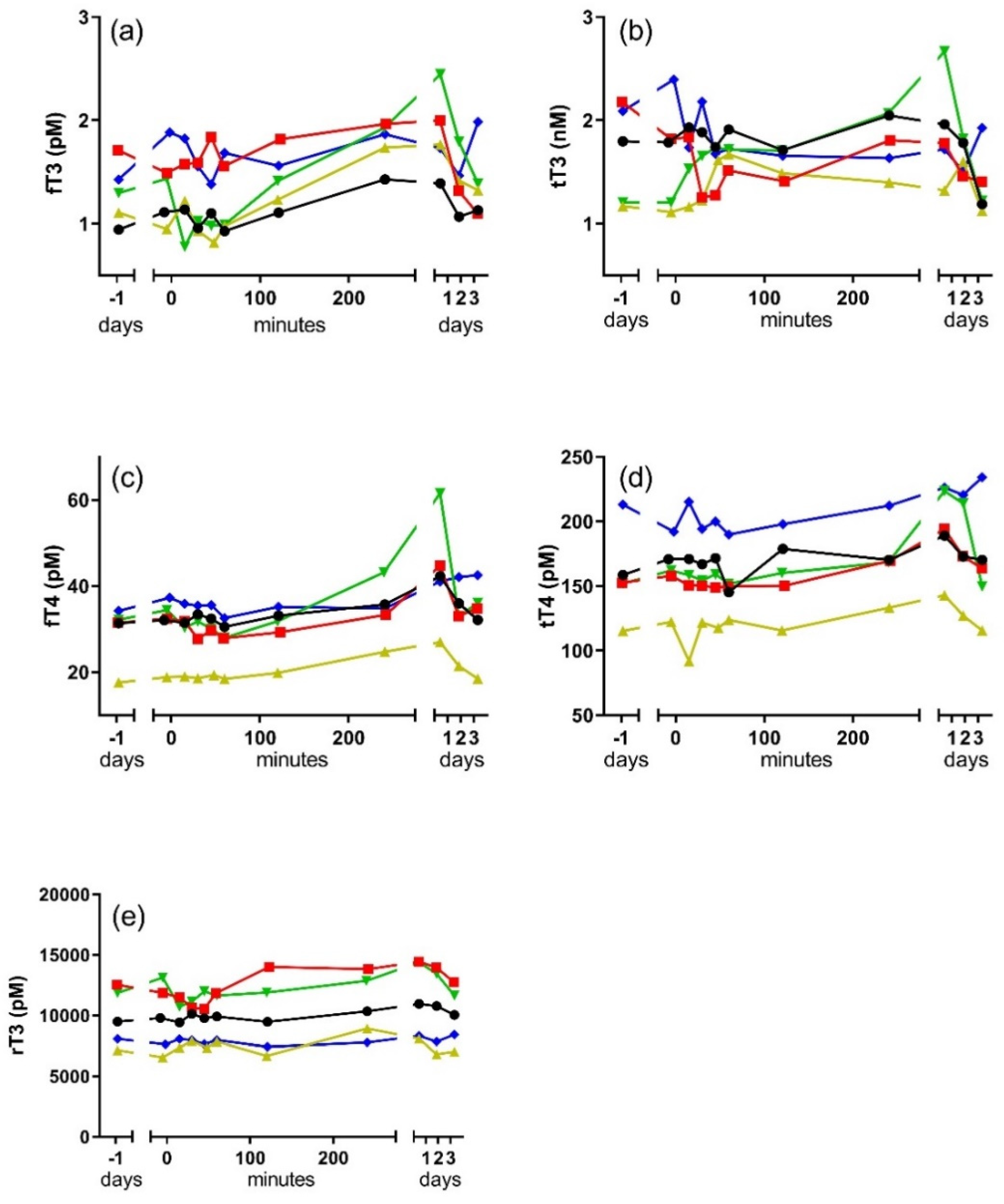
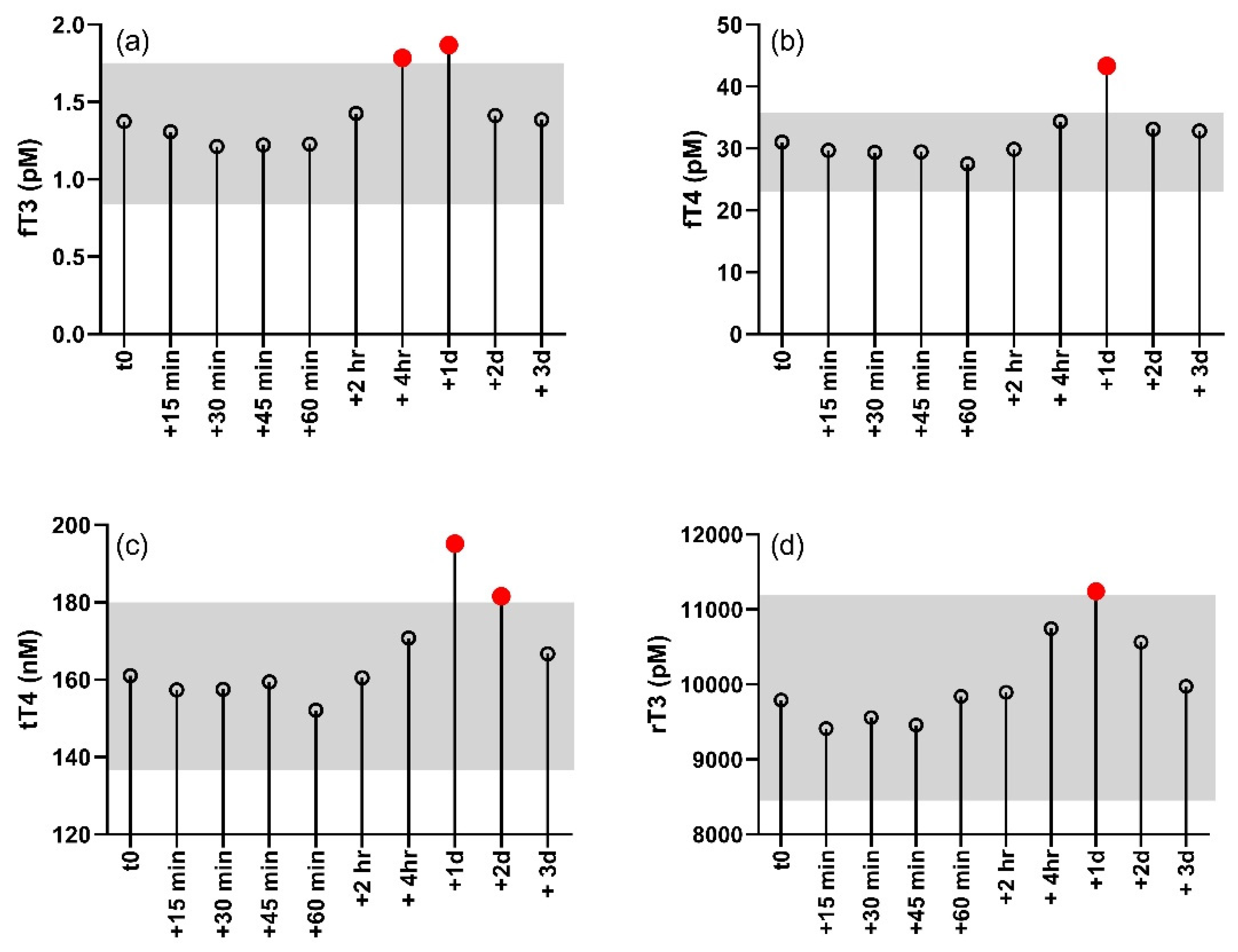
Changes in free and bound thyroid hormone in response to the TS are shown for each individual dolphin in
Figure 1. Individual levels of circulating thyroid hormones were variable between individuals, but all individuals showed an increase in fT3, fT4, tT4, and rT3 following the TS. Levels of fT3 were significantly higher than controls (−1 day) in blood samples collected four hours and one day after the TSH injection (F
10, 40 = 4.0,
p < 0.001; Dunnett’s
p < 0.05;
Figure 2). Similarly, rT3, fT4, and tT4 were significantly elevated one day after the injection, with tT4 remaining significantly elevated into the second day after the injection (rT3: F
10, 40 = 3.0,
p < 0.01; Dunnett’s
p < 0.05/fT4: F
10, 40 = 7.6,
p < 0.001; Dunnett’s
p < 0.05/tT4: F
10, 40 = 5.9,
p < 0.001; Dunnett’s
p < 0.05). All hormones showing a significant response to the TS returned to baseline by the third day after the TSH injection. Levels of tT3 did not significantly change in response to the TS, nor did ratios of the free and bound forms (fT3:fT4 and tT3:tT4) of the hormone.
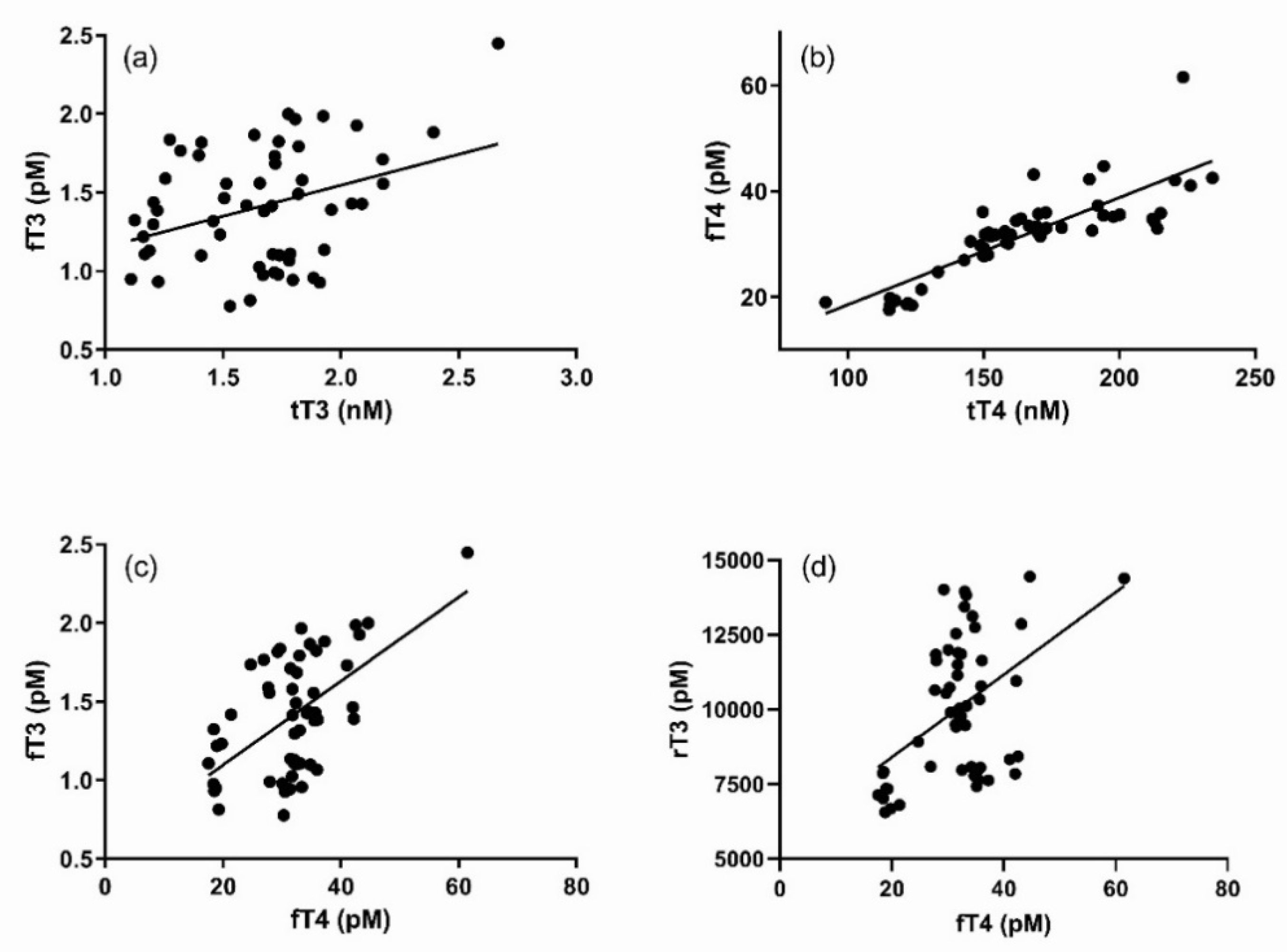
Results of regression analyses between thyroid hormones are shown in
Figure 3. There was a loose yet significant and direct relationship between fT3 and tT3 (
p < 0.01, r
2 = 0.12). A similar relationship existed between fT4 and tT4, but the amount of variance explained by the linear model was much higher (
p < 0.001, r
2 = 0.69) than that observed between fT3 and tT3. A significant, direct relationship was also observed between fT3 and fT4 (
p < 0.001, r
2 = 0.32). No relationship between rT3 and either tT3 or tT4 was observed, but there were loosely significant relationships between rT3 and fT3 (
p < 0.05, r
2 = 0.07), as well as between rT3 and fT4 (
p < 0.001, r
2 = 0.23).
4. Discussion
Stimulation of the thyroid is a potential tool for evaluating thyroid dysfunction. Prior to the development of sensitive TSH assays, the use of TRH was used to differentiate secondary and tertiary hypothyroidism. The use of TSH can similarly be used to directly assess dysfunction in the thyroid gland. TRH stimulation has only been reported once in a toothed whale, the beluga, and produced no significant changes in circulating thyroid hormones [
11]. In contrast, the use of both bovine TSH and human recombinant TSH have been used to effectively stimulate the production and secretion of thyroid hormones in belugas and bottlenose dolphins [
1,
7,
8].
West and colleagues [
7] were the first to report on the use of human recombinant TSH to stimulate the thyroid of a bottlenose dolphin. A single animal was used in the study and the intramuscular administration of 0.9 mg of TSH produced a modest increase in fT3 (51%) and tT3 (29%), but no distinct pattern was observed for either fT4 or tT4. In the current study, the same human recombinant TSH was utilized for thyroid stimulation, but the dose was increased to 1.5 mg per subject in hopes of producing a more pronounced effect. The sample size was also increased to determine whether changes in thyroid hormones were repeatable across animals. Despite the increased dose of TSH, the mean maximum increase in fT3 was 44%, which was not substantially different from that previously measured [
7]. However, whereas no distinctive increase in either fT4 or tT4 was observed by West and colleagues [
7], an increase of 47% was observed in fT4 with a more modest mean maximum increase in tT4 of 23%. No significant change in tT3 was observed and rT3 increased by a modest 14%, although the change was significant.
All of the thyroid hormones that significantly increased in response to the TS showed peak concentrations the day after the TSH injection. However, a significant increase was observed in fT3 as early as four hours after the TSH injection and a significant increase in tT4 persisted for up to two days after the injection. The pattern is suggestive of a time lag in thyroid hormone dynamics that is potentially reflective of feedback regulation. Thyroid regulation is sensitive to T3:T4, so it is probably not surprising that the strongest direct relationships existed between fT3 and fT4, as well as fT4 and tT4. A post-hoc analysis of both fT3:fT4 and tT3:tT4 showed no significant change in the ratio of the hormones throughout the post-injection observation period. Nevertheless, the delays between the first periods identified as significantly higher than baseline in fT4 (+one day) and tT4 (+one day) relative to the first period of significant increase in fT3 possibly reflects the time required to change peripheral rates of T4 deiodination in response to a growing excess of fT3. The significant relationship between rT3 and fT4, and the corresponding timing in their significant changes, supports an upregulation of T4 deiodination to the rT3 form.
It is feasible that the testing paradigm employed in the current study affected thyroid hormone levels, as the dolphins involved would have mounted a stress response to the out-of-water portion of the procedure during which sequential blood samples were collected. A similar paradigm was previously used to study the stress response in the bottlenose dolphin and the production of corticosteroids increased rapidly after voluntary beaching and the setting of the needle for sequential blood sampling [
3]. However, neither free nor total T3 and T4 demonstrated any significant change in response to the stress test [
3], suggesting that samples collected in the current study under a similar paradigm were likely not affected by an acute stress response. Under prolonged elevation of serum cortisol, feedback mechanisms might be expected to suppress thyroid hormone production. At elevated levels, cortisol is known to suppress TSH production and inhibit the conversion of T4 to T3 in humans. Similar to humans, significant reductions in fT3 and fT4 have been observed in bottlenose dolphins under pharmacological elevation of circulating cortisol [
12], and tT3 has been observed to decline in response to elevated cortisol in some age classes of the northern elephant seal (
Mirounga angustirostris) when submitted to adrenocorticotropic hormone stimulations [
13].
The results of this study provide baseline information on the response of the thyroid to TSH stimulation that can be utilized for clinical investigation of dolphin thyroid dysfunction; however, the method could be improved. The TSH utilized in the current study and by West and colleagues [
7] was human-derived. Both TSH and its receptor are highly derived for specificity across vertebrate species, minimizing cross-reactivity with glycoprotein receptors, and with receptor specificity and bioactivity potentially peaking with the evolution of the mammals [
14]. Thus, it is possible that the effectiveness of the human recombinant TSH was limited in its ability to promote production and secretion of thyroid hormone. Currently, no bottlenose dolphin-derived TSH is available for use in thyroid stimulation tests, nor are there commercial antibodies available to facilitate regular assessment of dolphin TSH. Development of specific antibodies for bottlenose dolphin TSH could greatly improve assessment of thyroid dysfunction, as performed in other species e.g., [
15]. Similarly, development of a species-specific thyroid hormone secretagogue (TSH) would presumably enhance the response of the thyroid to stimulation and enable more biologically-relevant kinetic analyses of thyroid hormone release and clearance, e.g., see [
16]. The development of such tools is costly and would generally be limited to specific research efforts in exotic species, i.e., it would have limited clinical utility because of its limited availability. However, the bottlenose dolphin has high-profile commercial value due to its abundance in marine parks, zoos, and aquaria, and its value could be used to leverage the development of tools that better enable veterinary assessment of thyroid function in this species.







{kind=link}
{kind=link}
{kind=link}