
Age and Social History Impact Social Interactions between Bull Asian Elephants (Elephas maximus) at Denver Zoo

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population and Study Site
2.2. Data Collection
2.3. Data Analysis
3. Results
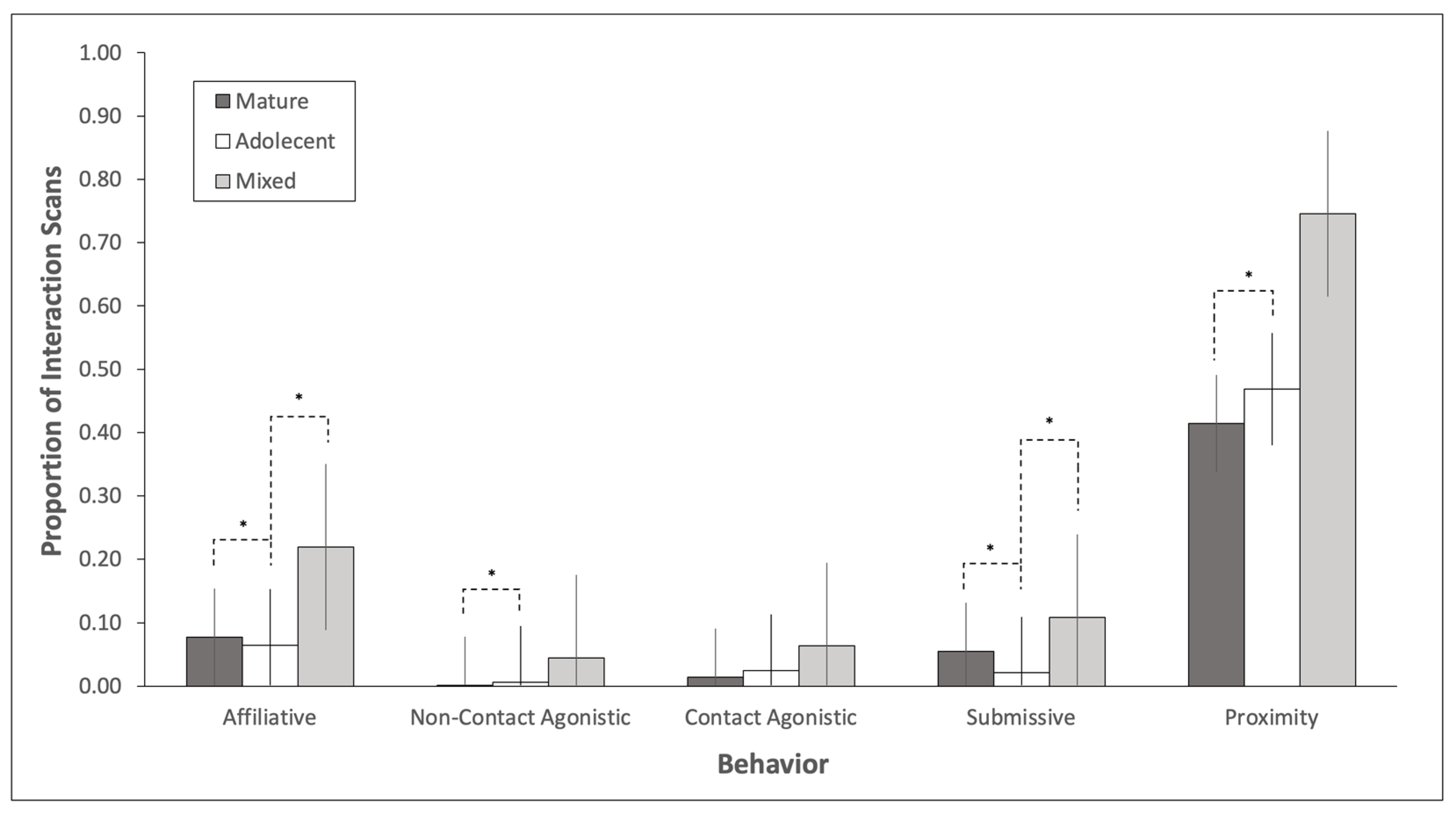
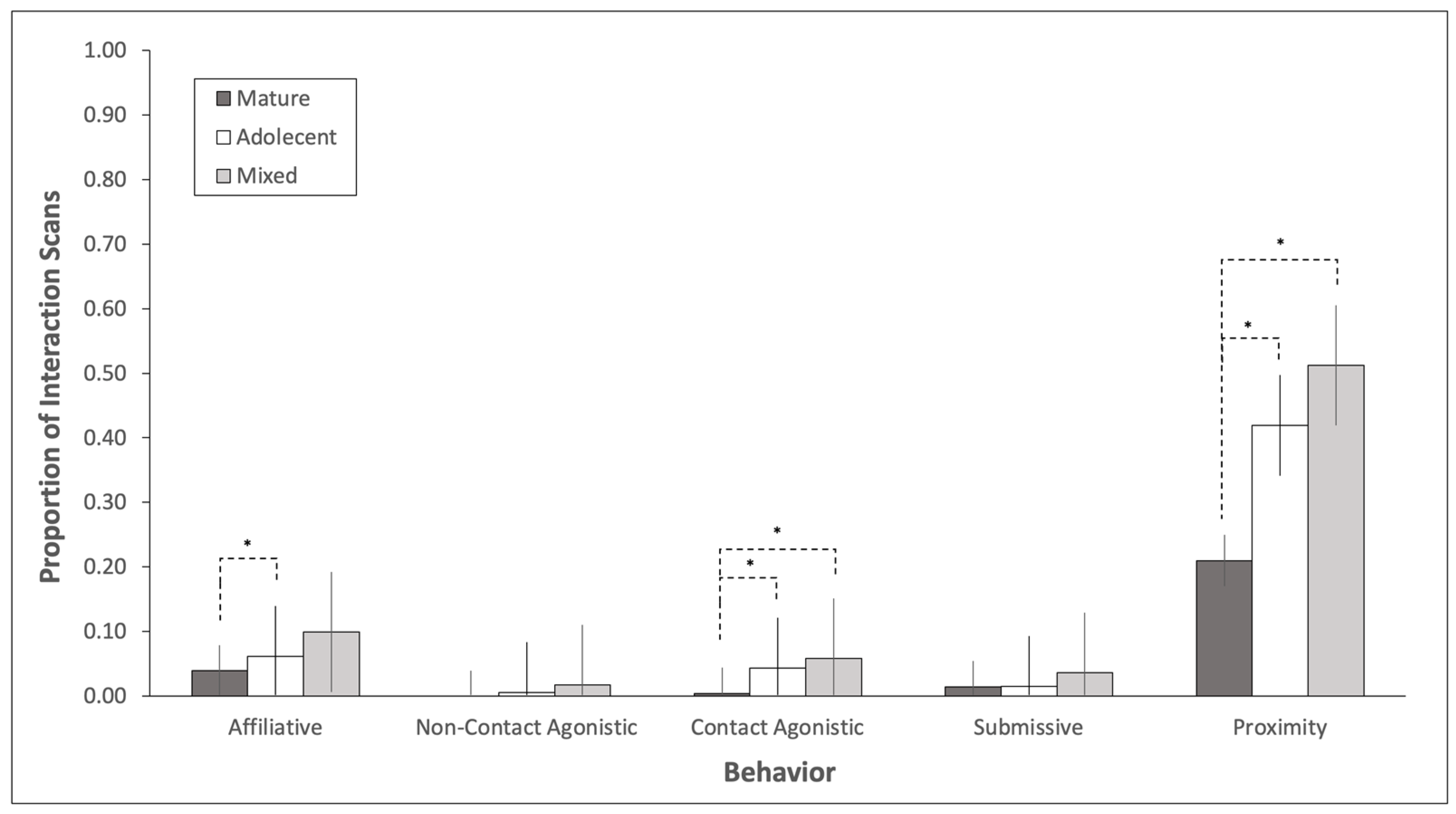
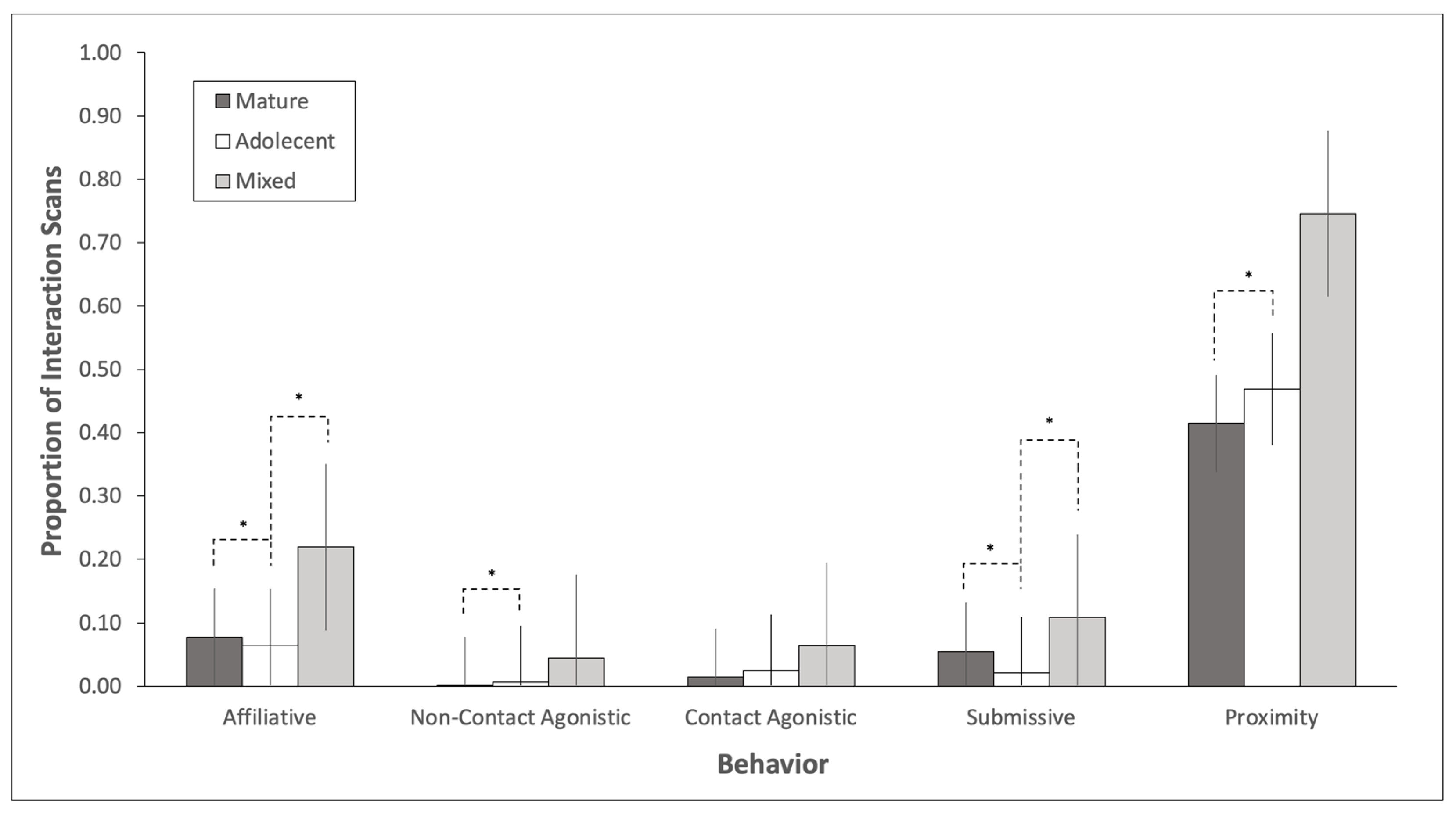
3.1. Adolescent Bull Behavior
3.2. Mature Bull Behavior
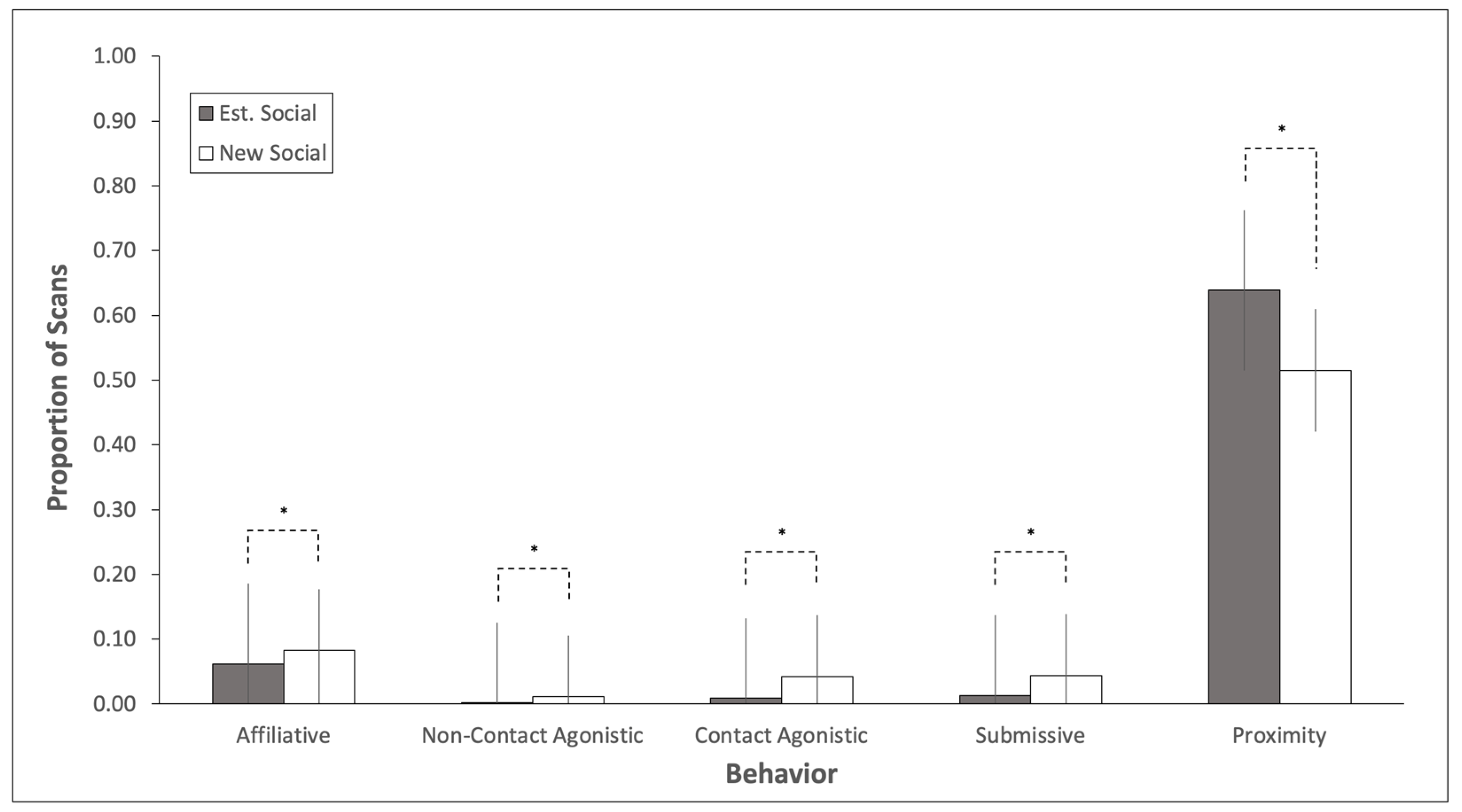
3.3. Social History Effects on Behavior
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Model Selection Tables for GLMER of Bull Elephant Behavior

{kind=link}
{kind=link}
{kind=link}
| Model | AIC | BIC | X2 | Parameters | p-Value |
|---|---|---|---|---|---|
| Affiliative | |||||
| SocialAgeLittle + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + NewSocial + InOutAccess*AccessArea + IntrosPeriod*NewSocial | 13,768 | 13,908 | - | 17 | - |
| SocialAgeLittle + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + NewSocial + InOutAccess*AccessArea | 13,766 | 13,898 | 1.020 | 16 | 0.313 |
| SocialAgeLittle + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + NewSocial | 13,763 | 13,878 | 1.432 | 14 | 0.698 |
| SocialAgeLittle + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod | 13,766 | 13,873 | 6.616 | 13 | 0.158 |
| SocialAgeLittle + TimeOfDay + Musth + InOutAccess + AccessArea | 13,767 | 13,866 | 9.693 | 12 | 0.084 |
| SocialAgeLittle + TimeOfDay + Musth + InOutAccess | 13,762 | 13,853 | 6.701 | 11 | 0.349 |
| SocialAgeLittle + TimeOfDay + Musth | 13,765 | 13,839 | 13.428 | 9 | 0.098 |
| Submissive | |||||
| SocialAgeLittles + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + NewSocial + InOutAccess*AccessArea + IntrosPeriod*NewSocial | 7908.1 | 8048.2 | - | 17 | - |
| SocialAgeLittles + TimeOfDay + InOutAccess + AccessArea + IntrosPeriod + NewSocial + InOutAccess*AccessArea + IntrosPeriod*NewSocial | 7907.0 | 8048.2 | 4.862 | 14 | 0.182 |
| SocialAgeLittles + TimeOfDay + InOutAccess + AccessArea + IntrosPeriod + NewSocial + InOutAccess*AccessArea | 7908.5 | 8015.6 | 8.365 | 13 | 0.079 |
| Non-contact Agonistic | |||||
| SocialAgeLittles + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + NewSocial + InOutAccess*AccessArea + IntrosPeriod*NewSocial | 2311.4 | 2451.4 | - | 17 | - |
| SocialAgeLittles + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + NewSocial + IntrosPeriod*NewSocial | 2309.2 | 2432.8 | 1.808 | 15 | 0.405 |
| SocialAgeLittles + TimeOfDay + Musth + InOutAccess + IntrosPeriod + NewSocial + IntrosPeriod*NewSocial | 2309.6 | 2424.9 | 4.198 | 14 | 0.241 |
| SocialAgeLittles + TimeOfDay + InOutAccess + IntrosPeriod + NewSocial + IntrosPeriod*NewSocial | 2306.1 | 2396.7 | 6.721 | 11 | 0.347 |
| SocialAgeLittles + TimeOfDay + IntrosPeriod + NewSocial + IntrosPeriod*NewSocial | 2303.0 | 2377.1 | 7.586 | 9 | 0.475 |
| Contact Agonistic | |||||
| SocialAgeLittles + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + NewSocial + InOutAccess*AccessArea + IntrosPeriod*NewSocial | 5368.2 | 5508.3 | - | 17 | - |
| SocialAgeLittles + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + NewSocial + IntrosPeriod*NewSocial | 5367.5 | 5491.1 | 3.274 | 15 | 0.195 |
| SocialAgeLittles + TimeOfDay + Musth + InOutAccess + IntrosPeriod + NewSocial + IntrosPeriod*NewSocial | 5365.7 | 5481.0 | 3.481 | 14 | 0.323 |
| Proximity | |||||
| SocialAgeLittles + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + NewSocial + InOutAccess*AccessArea + IntrosPeriod*NewSocial | 25,842 | 25,982 | - | 17 | - |
| SocialAgeLittles + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + NewSocial + IntrosPeriod*NewSocial | 25,840 | 25,964 | 1.772 | 15 | 0.412 |
| SocialAgeLittles + TimeOfDay + Musth + InOutAccess + IntrosPeriod + NewSocial + IntrosPeriod*NewSocial | 25,839 | 25,954 | 2.180 | 14 | 0.536 |
| Model | AIC | BIC | X2 | Parameters | p-Value |
|---|---|---|---|---|---|
| Affiliative | |||||
| SocialAgeOlder + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + NewSocial + InOutAccess*AccessArea + IntrosPeriod*NewSocial | 4392.6 | 4516.2 | - | 17 | - |
| SocialAgeOlder + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + NewSocial + InOutAccess*AccessArea | 4390.6 | 4506.9 | 0.011 | 16 | 0.916 |
| SocialAgeOlder + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + NewSocial | 4386.7 | 4488.4 | 0.038 | 14 | 0.998 |
| SocialAgeOlder + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod | 4384.8 | 4479.3 | 0.183 | 13 | 0.996 |
| SocialAgeOlder + TimeOfDay + Musth + InOutAccess + AccessArea | 4387.1 | 4474.3 | 4.430 | 12 | 0.489 |
| SocialAgeOlder + TimeOfDay + Musth + InOutAccess | 4389.5 | 4469.5 | 8.902 | 11 | 0.179 |
| SocialAgeOlder + TimeOfDay + Musth | 4387.6 | 4453.0 | 10.961 | 9 | 0.204 |
| Submissive | |||||
| SocialAgeOlder + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + NewSocial + InOutAccess*AccessArea + IntrosPeriod*NewSocial | 1608.7 | 1732.2 | - | 17 | - |
| SocialAgeOlder + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + NewSocial + InOutAccess*AccessArea | 1607.2 | 1723.4 | 0.490 | 16 | 0.484 |
| SocialAgeOlder + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + NewSocial | 1604.9 | 1706.7 | 2.236 | 14 | 0.525 |
| SocialAgeOlder + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod | 1607.5 | 1702.0 | 6.851 | 13 | 0.144 |
| SocialAgeOlder + TimeOfDay + Musth + InOutAccess + AccessArea | 1606.3 | 1693.5 | 7.631 | 12 | 0.178 |
| SocialAgeOlder + TimeOfDay + Musth + AccessArea | 1606.1 | 1678.7 | 11.393 | 10 | 0.122 |
| Non-contact Agonistic | |||||
| SocialAgeOlder + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + NewSocial + InOutAccess*AccessArea + IntrosPeriod*NewSocial | 669.6 | 793.1 | - | 17 | - |
| SocialAgeOlder + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + NewSocial + IntrosPeriod*NewSocial | 668.7 | 777.7 | 3.157 | 15 | 0.206 |
| SocialAgeOlder + TimeOfDay + Musth + InOutAccess + IntrosPeriod + NewSocial + IntrosPeriod*NewSocial | 667.3 | 769.0 | 3.713 | 14 | 0.294 |
| SocialAgeOlder + TimeOfDay + Musth + IntrosPeriod + NewSocial + IntrosPeriod*NewSocial | 666.7 | 754.0 | 7.168 | 12 | 0.208 |
| SocialAgeOlder + TimeOfDay + IntrosPeriod + NewSocial + IntrosPeriod*NewSocial | 665.8 | 731.2 | 12.261 | 9 | 0.140 |
| SocialAgeOlder + TimeOfDay + IntrosPeriod + NewSocial | 665.9 | 724.1 | 14.343 | 8 | 0.111 |
| SocialAgeOlder + TimeOfDay + IntrosPeriod | 664.2 | 715.1 | 14.667 | 7 | 0.145 |
| Contact Agonistic | |||||
| SocialAgeOlder + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + NewSocial + InOutAccess*AccessArea + IntrosPeriod*NewSocial | 2951.3 | 3074.9 | - | 17 | - |
| SocialAgeOlder + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + NewSocial + InOutAccess*AccessArea | 2951.2 | 3067.5 | 1.903 | 16 | 0.168 |
| SocialAgeOlder + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + NewSocial | 2948.9 | 3050.6 | 3.519 | 14 | 0.318 |
| SocialAgeOlder + TimeOfDay + InOutAccess + AccessArea + IntrosPeriod + NewSocial | 2947.1 | 3027.1 | 7.787 | 11 | 0.254 |
| Proximity | |||||
| SocialAgeOlder + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + NewSocial + InOutAccess*AccessArea + IntrosPeriod*NewSocial | 10,112 | 10,236 | - | 17 | - |
| SocialAgeOlder + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + NewSocial + InOutAccess*AccessArea | 10,110 | 10,226 | 0.172 | 16 | 0.678 |
| SocialAgeOlder + TimeOfDay + Musth + InOutAccess + AccessArea + NewSocial + InOutAccess*AccessArea | 10,108 | 10,217 | 0.220 | 15 | 0.896 |
| Model | AIC | BIC | X2 | Parameters | p-Value |
| Affiliative | |||||
| NewSocial + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + InOutAccess*AccessArea + IntrosPeriod*NewSocial | 18,039 | 18,167 | - | 15 | - |
| NewSocial + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + InOutAccess*AccessArea | 18,037 | 18,157 | 0.725 | 14 | 0.395 |
| NewSocial + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod | 18,037 | 18,140 | 4.216 | 12 | 0.239 |
| NewSocial + TimeOfDay + Musth + InOutAccess + AccessArea | 18,036 | 18,036 | 5.523 | 11 | 0.238 |
| NewSocial + TimeOfDay + Musth + InOutAccess | 18,034 | 18,120 | 5.542 | 10 | 0.353 |
| NewSocial + TimeOfDay + InOutAccess | 18,032 | 18,167 | 9.517 | 8 | 0.301 |
| Submissive | |||||
| NewSocial + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + InOutAccess*AccessArea + IntrosPeriod*NewSocial | 9784.3 | 9912.6 | - | 15 | - |
| NewSocial + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + IntrosPeriod*NewSocial | 9784.2 | 9895.5 | 3.965 | 13 | 0.138 |
| Non-contact Agonistic | |||||
| NewSocial + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + InOutAccess*AccessArea + IntrosPeriod*NewSocial | 3038.2 | 3166.6 | - | 15 | - |
| NewSocial + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + IntrosPeriod*NewSocial | 3036.2 | 3147.5 | 2.029 | 13 | 0.363 |
| NewSocial + TimeOfDay + Musth + InOutAccess + IntrosPeriod + IntrosPeriod*NewSocial | 3038.2 | 3140.9 | 6.038 | 12 | 0.120 |
| NewSocial + TimeOfDay + Musth + IntrosPeriod + IntrosPeriod*NewSocial | 3036.7 | 3122.3 | 8.537 | 10 | 0.129 |
| Contact Agonistic | |||||
| NewSocial + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + InOutAccess*AccessArea + IntrosPeriod*NewSocial | 8390.7 | 8519.1 | - | 15 | - |
| Proximity | |||||
| NewSocial + TimeOfDay + Musth + InOutAccess + AccessArea + IntrosPeriod + InOutAccess*AccessArea + IntrosPeriod*NewSocial | 35,414 | 35,542 | - | 15 | - |
Appendix B. Model Output Tables
| Predictor | Level | Affiliative | Submissive | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Odds Ratio | ß | SE | z-Statistic | p | Odds Ratio | ß | SE | z-Statistic | p | ||
| SocialAge | Adolescent * | - | - | - | - | - | - | - | - | - | - |
| Mature | 1.479 | 0.392 | 0.152 | 2.582 | 0.010 | 3.078 | 1.124 | 0.155 | 7.252 | <0.001 | |
| Mixed | 2.226 | 0.800 | 0.224 | 3.574 | <0.001 | 2.460 | 0.900 | 0.260 | 3.461 | <0.001 | |
| TimeOfDay | Morning * | - | - | - | - | - | - | - | - | - | - |
| Afternoon | 0.735 | −0.308 | 0.192 | −1.606 | 0.108 | 0.398 | −0.922 | 0.218 | −4.234 | <0.001 | |
| Night | 0.112 | −2.187 | 0.164 | −13.309 | <0.001 | 0.100 | −2.307 | 0.218 | −9.125 | <0.001 | |
| InOutAccess | Both * | - | - | - | - | - | - | - | - | - | - |
| Inside | - | - | - | - | - | 1.679 | 0.518 | 1.157 | 0.448 | 0.654 | |
| Outside | - | - | - | - | - | 0.844 | −0.170 | 1.120 | −0.151 | 0.880 | |
| AccessArea | - | - | - | - | - | 0.999 | −0.001 | 0.064 | −0.009 | 0.993 | |
| Musth | None * | - | - | - | - | - | - | - | - | - | - |
| Indiv. 2 | 1.203 | 0.185 | 0.172 | 1.073 | 0.283 | - | - | - | - | - | |
| Indiv. 3 | 0.653 | −0.426 | 0.179 | −2.379 | 0.017 | - | - | - | - | - | |
| Indiv. 2 and 3 | 0.857 | −0.154 | 0.264 | −0.582 | 0.560 | - | - | - | - | - | |
| IntrosPeriod | 0 * | - | - | - | - | - | - | - | - | - | - |
| 1 | - | - | - | - | - | 1.490 | 0.399 | 0.186 | 2.142 | 0.032 | |
| NewSocial | 0 * | - | - | - | - | - | - | - | - | - | - |
| 1 | - | - | - | - | - | 4.460 | 1.495 | 0.173 | 8.636 | <0.001 | |
| InOutAccess*AccessArea | In | - | - | - | - | - | 0.711 | −0.342 | 0.169 | −2.017 | 0.044 |
| InOutAccess*AccessArea | Out | - | - | - | - | - | 0.981 | −0.019 | 0.065 | −0.290 | 0.772 |
| Predictor | Level | No-Contact Agonistic | Contact Agonistic | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Odds Ratio | ß | SE | z-Statistic | p | Odds Ratio | ß | SE | z-Statistic | p | ||
| SocialAge | Adolescent * | - | - | - | - | - | - | - | - | - | - |
| Mature | 0.175 | −1.741 | 0.416 | −4.187 | <0.001 | 0.619 | −0.479 | 0.292 | −1.639 | 0.101 | |
| Mixed | 1.341 | 0.294 | 0.307 | 0.957 | 0.338 | 1.029 | −0.028 | 0.420 | −0.067 | 0.947 | |
| TimeOfDay | Morning * | - | - | - | - | - | - | - | - | - | - |
| Afternoon | 0.496 | −0.702 | 0.325 | −2.160 | 0.031 | 0.531 | −0.634 | 0.352 | −1.802 | 0.071 | |
| Night | 0.029 | −3.538 | 0.472 | −7.498 | <0.001 | 0.100 | −2.307 | 0.411 | −5.608 | <0.001 | |
| InOutAccess | Both * | - | - | - | - | - | - | - | - | - | - |
| Inside | - | - | - | - | - | 1.692 | 0.526 | 0.396 | 1.328 | 0.184 | |
| Outside | - | - | - | - | - | 0.841 | −0.173 | 0.376 | −0.460 | 0.646 | |
| Musth | None * | - | - | - | - | - | - | - | - | - | - |
| Indiv. 2 | - | - | - | - | - | 0.663 | −0.412 | 0.351 | −1.174 | 0.241 | |
| Indiv. 3 | - | - | - | - | - | 0.286 | −1.253 | 0.398 | −3.146 | 0.002 | |
| Indiv. 2 and 3 | - | - | - | - | - | 0.839 | −0.176 | 0.499 | −0.352 | 0.725 | |
| IntrosPeriod | 0 * | - | - | - | - | - | - | - | - | - | - |
| 1 | 0.871 | −0.138 | 0.685 | −0.201 | 0.840 | 0.742 | −0.298 | 0.521 | −0.572 | 0.568 | |
| NewSocial | 0 * | - | - | - | - | - | - | - | - | - | - |
| 1 | 6.880 | 1.929 | 0.526 | 3.663 | <0.001 | 5.773 | 1.753 | 0.362 | 4.847 | <0.001 | |
| IntrosPeriod*NewSocial | 4.386 | 1.478 | 0.746 | 1.983 | 0.047 | 5.577 | 1.719 | 0.588 | 2.922 | 0.003 | |
| Predictor | Level | Proximity | ||||
| Odds Ratio | ß | SE | z-Statistic | p | ||
| SocialAge | Adolescent * | - | - | - | - | - |
| Mature | 0.421 | −0.865 | 0.219 | −3.944 | <0.001 | |
| Mixed | 1.606 | 0.474 | 0.358 | 1.322 | 0.186 | |
| TimeOfDay | Morning * | - | - | - | - | - |
| Afternoon | 0.758 | −0.278 | 0.313 | −0.888 | 0.374 | |
| Night | 0.132 | −2.028 | 0.319 | −6.350 | <0.001 | |
| InOutAccess | Both * | - | - | - | - | - |
| Inside | 0.909 | −0.095 | 0.284 | −0.335 | 0.737 | |
| Outside | 0.362 | −1.016 | 0.300 | −3.389 | <0.001 | |
| Musth | None * | - | - | - | - | - |
| Indiv. 2 | 0.512 | −0.669 | 0.282 | −2.375 | 0.018 | |
| Indiv. 3 | 0.288 | −1.244 | 0.297 | −4.185 | <0.001 | |
| Indiv. 2 and 3 | 0.638 | −0.449 | 0.416 | −1.078 | 0.281 | |
| IntrosPeriod | 0 * | - | - | - | - | - |
| 1 | 0.440 | −0.821 | 0.368 | −2.234 | 0.026 | |
| NewSocial | 0 | - | - | - | - | - |
| 1 | 1.788 | 0.581 | 0.239 | 2.433 | 0.015 | |
| IntrosPeriod*NewSocial | 3.273 | 1.186 | 0.488 | 2.428 | 0.015 | |
| Predictor | Level | Affiliative | Submissive | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Odds Ratio | ß | SE | z-Statistic | p | Odds Ratio | ß | SE | z-Statistic | p | ||
| SocialAge | Mature * | - | - | - | - | - | - | - | - | - | - |
| Adolescent | 2.173 | 0.776 | 0.372 | 2.084 | 0.037 | 1.352 | 0.301 | 0.593 | 0.508 | 0.611 | |
| Mixed | 1.185 | 0.170 | 0.471 | 0.360 | 0.719 | 3.029 | 1.108 | 0.853 | 1.300 | 0.194 | |
| TimeOfDay | Morning * | - | - | - | - | - | - | - | - | - | - |
| Afternoon | 0.976 | −0.024 | 0.287 | −0.085 | 0.933 | 0.375 | −0.981 | 0.521 | −1.881 | 0.060 | |
| Night | 0.185 | −1.688 | 0.229 | −7.360 | <0.001 | 0.094 | −2.365 | 0.463 | −5.108 | <0.001 | |
| AccessArea | - | - | - | - | - | 0.954 | −0. | 0.022 | −2.317 | 0.020 | |
| Musth | None * | - | - | - | - | - | - | - | - | - | - |
| Indiv. 2 | 0.431 | −0.842 | 0.276 | −3.050 | 0.002 | 0.421 | −0.866 | 0.503 | −1.721 | 0.085 | |
| Indiv. 3 | 0.570 | −0.562 | 0.228 | −2.468 | 0.014 | 0.182 | −1.704 | 0.477 | −3.575 | <0.001 | |
| Indiv. 2 and 3 | 0.503 | −0.686 | 0.491 | −1.399 | 0.162 | 1.399 | 0.336 | 0.746 | 0.451 | 0.652 | |
| Predictor | Level | Non-Contact Agonistic | Contact Agonistic | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Odds Ratio | ß | SE | z-Statistic | p | Odds Ratio | ß | SE | z-Statistic | p | ||
| SocialAge | Mature * | - | - | - | - | - | - | - | - | - | - |
| Adolescent | 10,449,869 | 16.162 | 9.206 | 1.756 | 0.079 | 5.140 | 1.637 | 0.696 | 2.352 | 0.019 | |
| Mixed | 12,362,769 | 16.330 | 9.214 | 1.772 | 0.076 | 9.854 | 2.288 | 0.911 | 2.510 | 0.012 | |
| TimeOfDay | Morning * | - | - | - | - | - | - | - | - | - | - |
| Afternoon | 1.307 | 0.267 | 0.451 | 0.593 | 0.553 | 0.457 | −0.784 | 0.422 | −1.856 | 0.063 | |
| Night | 0.230 | −1.481 | 0.450 | −3.289 | 0.001 | 0.222 | −1.506 | 0.458 | −3.290 | 0.001 | |
| AccessArea | - | - | - | - | - | 0.927 | −0.076 | 0.023 | −3.292 | <0.001 | |
| InOutAccess | Both * | - | - | - | - | - | - | - | - | - | - |
| Inside | - | - | - | - | - | 0.157 | −1.852 | 0.477 | −3.880 | <0.001 | |
| Outside | - | - | - | - | - | 0.709 | −0.345 | 0.417 | −0.826 | 0.409 | |
| IntrosPeriod | 0 * | - | - | - | - | - | - | - | - | - | - |
| 1 | 3.002 | 1.099 | 0.375 | 2.932 | 0.003 | 3.639 | 1.292 | 0.327 | 3.950 | <0.001 | |
| NewSocial | 0 * | - | - | - | - | - | - | - | - | - | - |
| 1 | - | - | - | - | - | 4.546 | 1.514 | 0.311 | 4.868 | <0.001 | |
| Predictor | Level | Proximity | ||||
|---|---|---|---|---|---|---|
| Odds Ratio | ß | SE | z-Statistic | p | ||
| SocialAge | Mature * | - | - | - | - | - |
| Adolescent | 6.096 | 1.808 | 0.535 | 3.381 | <0.001 | |
| Mixed | 7.012 | 1.948 | 0.815 | 2.390 | 0.017 | |
| TimeOfDay | Morning * | - | - | - | - | - |
| Afternoon | 0.705 | −0.349 | 0.432 | −0.808 | 0.419 | |
| Night | 0.203 | −1.595 | 0.434 | −3.676 | 0.001 | |
| InOutAccess | Both * | - | - | - | - | - |
| Inside | 74.020 | 4.304 | 1.419 | 3.034 | 0.002 | |
| Outside | 283.758 | 5.648 | 1.457 | 3.876 | <0.001 | |
| AccessArea | 1.301 | 0.263 | 0.078 | 3.388 | <0.001 | |
| Musth | None * | - | - | - | - | - |
| Indiv. 2 | 0.289 | −1.243 | 0.372 | −3.344 | <0.001 | |
| Indiv. 3 | 0.403 | −0.908 | 0.330 | −2.751 | 0.006 | |
| Indiv. 2 and 3 | 0.831 | −0.185 | 0.727 | −0.255 | 0.799 | |
| NewSocial | 0 | - | - | - | - | - |
| 1 | 3.862 | 1.351 | 0.271 | 4.983 | <0.001 | |
| InOutAccess*AccessArea | In | 0.864 | −0.146 | 0.221 | −0.661 | 0.509 |
| InOutAccess*AccessArea | Out | 0.720 | −0.328 | 0.080 | −4.114 | <0.001 |
| Predictor | Level | Affiliative | Submissive | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Odds Ratio | ß | SE | z-Statistic | p | Odds Ratio | ß | SE | z-Statistic | p | ||
| NewSocial | 0 * | - | - | - | - | - | - | - | - | - | - |
| 1 | 1.343 | 0.295 | 0.135 | 2.186 | 0.029 | 3.671 | 1.300 | 0.203 | 6.421 | <0.001 | |
| TimeOfDay | Morning * | - | - | - | - | - | - | - | - | - | - |
| Afternoon | 0.701 | −0.355 | 0.191 | −1.858 | 0.063 | 0.399 | −0.918 | 0.225 | −4.082 | <0.001 | |
| Night | 0.079 | −2.545 | 0.203 | −12.555 | <0.001 | 0.207 | −2.236 | 0.246 | −9.071 | <0.001 | |
| InOutAccess | Both * | - | - | - | - | - | - | - | - | - | - |
| Inside | 0.691 | −0.370 | 0.154 | −2.398 | 0.017 | 0.513 | −0.668 | 0.243 | −2.747 | 0.006 | |
| Outside | 0.588 | −0.531 | 0.196 | −2.711 | 0.007 | 0.749 | −0.289 | 0.229 | −1.260 | 0.208 | |
| Musth | None * | - | - | - | - | - | - | - | - | - | - |
| Indiv. 2 | - | - | - | - | - | 0.621 | −0.476 | 0.195 | −2.449 | 0.014 | |
| Indiv. 3 | - | - | - | - | - | 0.626 | −0.468 | 0.221 | −2.115 | 0.034 | |
| Indiv. 2 and 3 | - | - | - | - | - | 0.475 | −0.745 | 0.325 | −2.294 | 0.022 | |
| AccessArea | - | - | - | - | - | 0.974 | −0.027 | 0.010 | −2.697 | 0.007 | |
| IntrosPeriod | 0 * | - | - | - | - | - | - | - | - | - | - |
| 1 | - | - | - | - | - | 1.108 | 0.018 | 0.292 | 0.060 | 0.952 | |
| IntrosPeriod*NewSocial | - | - | - | - | - | 1.931 | 0.658 | 0.349 | 1.885 | 0.059 | |
| Predictor | Level | Non-Contact Agonistic | Contact Agonistic | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Odds Ratio | ß | SE | z-Statistic | p | Odds Ratio | ß | SE | z-Statistic | p | ||
| NewSocial | 0 * | - | - | - | - | - | - | - | - | - | - |
| 1 | 3.516 | 1.257 | 0.370 | 3.397 | <0.001 | 6.647 | 1.894 | 0.285 | 6.647 | <0.001 | |
| TimeOfDay | Morning * | - | - | - | - | - | - | - | - | - | - |
| Afternoon | 0.552 | −0.594 | 0.297 | −2.002 | 0.045 | 0.443 | −0.814 | 0.295 | −2.762 | 0.006 | |
| Night | 0.053 | −2.931 | 0.318 | −9.212 | <0.001 | 0.086 | −2.455 | 0.327 | −7.505 | <0.001 | |
| InOutAccess | Both* | - | - | - | - | - | - | - | - | - | - |
| Inside | - | - | - | - | - | 1.679 | 0.518 | 1.154 | 0.449 | 0.653 | |
| Outside | - | - | - | - | - | 0.467 | −0.762 | 1.096 | −0.695 | 0.487 | |
| AccessArea | - | - | - | - | - | 0.957 | −0.044 | 0.060 | −0.737 | 0.461 | |
| Musth | None * | - | - | - | - | - | - | - | - | - | - |
| Indiv. 2 | 0.742 | −0.298 | 0.282 | −1.058 | 0.290 | 0.639 | −0.447 | 0.260 | −1.717 | 0.086 | |
| Indiv. 3 | 0.573 | −0.556 | 0.352 | −1.580 | 0.114 | 0.410 | −0.891 | 0.303 | −2.943 | 0.003 | |
| Indiv. 2 and 3 | 0.446 | −0.809 | 0.451 | −1.792 | 0.073 | 0.922 | −0.081 | 0.417 | −0.195 | 0.846 | |
| IntrosPeriod | 0 * | - | - | - | - | - | - | - | - | - | - |
| 1 | 0.906 | −0.098 | 0.513 | −0.192 | 0.848 | 0.961 | −0.039 | 0.403 | −0.098 | 0.922 | |
| InOutAccess*AccessArea | In | - | - | - | - | - | 0.657 | −0.419 | 0.215 | −1.949 | 0.051 |
| InOutAccess*AccessArea | Out | - | - | - | - | - | 1.026 | 0.025 | 0.061 | 0.414 | 0.679 |
| IntrosPeriod*NewSocial | 3.967 | 1.378 | 0.566 | 2.436 | 0.015 | 3.076 | 1.124 | 0.470 | 2.389 | 0.017 | |
| Predictor | Level | Proximity | ||||
|---|---|---|---|---|---|---|
| Odds Ratio | ß | SE | z-Statistic | p | ||
| NewSocial | 0 * | - | - | - | - | - |
| 1 | 2.097 | 0.741 | 0.239 | 3.094 | 0.002 | |
| TimeOfDay | Morning * | - | - | - | - | - |
| Afternoon | 0.631 | −0.461 | 0.310 | −1.486 | 0.137 | |
| Night | 0.122 | −2.106 | 0.307 | −6.869 | <0.001 | |
| InOutAccess | Both * | - | - | - | - | - |
| Inside | 2.252 | 0.812 | 0.915 | 0.888 | 0.375 | |
| Outside | 3.291 | 1.191 | 0.899 | 1.325 | 0.185 | |
| AccessArea | ||||||
| Musth | None * | - | - | - | - | - |
| Indiv. 2 | 0.854 | −0.158 | 0.262 | −0.601 | 0.548 | |
| Indiv. 3 | 0.592 | −0.525 | 0.286 | −1.836 | 0.114 | |
| Indiv. 2 and 3 | 1.024 | −0.024 | 0.413 | 0.057 | 0.955 | |
| IntrosPeriod | 0* | - | - | - | - | - |
| 1 | 0.599 | −0.512 | 0.355 | −1.443 | 0.149 | |
| InOutAccess*AccessArea | In | 1.056 | 0.055 | 0.147 | 0.372 | 0.710 |
| InOutAccess*AccessArea | Out | 0.873 | −0.136 | 0.051 | −2.665 | 0.008 |
| IntrosPeriod*NewSocial | 2.265 | 0.818 | 0.476 | 1.717 | 0.086 | |
References
- Funston, P.J.; Mills, M.G.L.; Richardson, P.R.K.; Jaarsveld, A.S. Reduced Dispersal and Opportunistic Territory Acquisition in Male Lions (Panthera leo). J. Zool. 2003, 259, 131–142. [Google Scholar] [CrossRef]
- Van Hooff, J.A.R.A.M.; Van Schaik, C.P. Male Bonds: Afilliative Relationships Among Nonhuman Primate Males. Behaviour 1994, 130, 309–337. [Google Scholar] [CrossRef]
- Bygott, D.J.; Bertram, B.C.R.; Hanby, J.P. Male Lions in Large Coalitions Gain Reproductive Advantages. Nature 1979, 282, 839–841. [Google Scholar] [CrossRef]
- Chiyo, P.I.; Archie, E.A.; Hollister-Smith, J.A.; Lee, P.C.; Poole, J.H.; Moss, C.J.; Alberts, S.C. Association Patterns of African Elephants in All-Male Groups: The Role of Age and Genetic Relatedness. Anim. Behav. 2011, 81, 1093–1099. [Google Scholar] [CrossRef]
- Connor, R.C.; Smolker, R.A.; Richards, A.F. Two Levels of Alliance Formation among Male Bottlenose Dolphins (Tursiops Sp.). Proc. Natl. Acad. Sci. USA 1992, 89, 987–990. [Google Scholar] [CrossRef] [PubMed]
- MacFarlane, A.M.; Coulson, G. Boys Will Be Boys: Social Affinity among Males Drives Social Segregation in Western Grey Kangaroos. J. Zool. 2009, 277, 37–44. [Google Scholar] [CrossRef]
- Parsons, K.; Durban, J.; Claridge, D.; Balcomb, K.; Noble, L.; Thompson, P. Kinship as a Basis for Alliance Formation among Male Bottlenose Dolphins (Tursiops truncatus) in the Bahamas. Anim. Behav. 2003, 66, 185–194. [Google Scholar] [CrossRef]
- Kie, J.G.; Bowyer, R.T. Sexual Segregation in White-Tailed Deer: Density-Dependent Changes in Use of Space, Habitat Selection, and Dietary Niche. J. Mammal. 1999, 80, 1004–1020. [Google Scholar] [CrossRef]
- Miquelle, D.G.; Peek, J.M.; Van Ballenberghe, V. Sexual Segregation in Alaskan Moose. Wildl. Monogr. 1992, 122, 3–57. [Google Scholar]
- Riss, D.; Goodall, J. The Recent Rise to the Alpha-Rank in a Population of Free-Living Chimpanzees. IJFP 1977, 27, 134–151. [Google Scholar] [CrossRef]
- Allen, C.R.B.; Brent, L.J.N.; Motsentwa, T.; Weiss, M.N.; Croft, D.P. Importance of Old Bulls: Leaders and Followers in Collective Movements of All-Male Groups in African Savannah Elephants (Loxodonta africana). Sci. Rep. 2020, 10, 13996. [Google Scholar] [CrossRef] [PubMed]
- Vidya, T.N.C.; Fernando, P.; Melnick, D.J.; Sukumar, R. Population Differentiation within and among Asian Elephant (Elephas maximus) Populations in Southern India. Heredity 2005, 94, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Archie, E.A.; Hollister-Smith, J.A.; Poole, J.H.; Lee, P.C.; Moss, C.J.; Maldonado, J.E.; Fleischer, R.C.; Alberts, S.C. Behavioural Inbreeding Avoidance in Wild African Elephants. Mol. Ecol. 2007, 16, 4138–4148. [Google Scholar] [CrossRef] [PubMed]
- Vidya, T.N.C.; Sukumar, R. Social and Reproductive Behaviour in Elephants. Curr. Sci. 2005, 89, 1200–1207. [Google Scholar]
- Chiyo, P.I.; Moss, C.J.; Alberts, S.C. The Influence of Life History Milestones and Association Networks on Crop-Raiding Behavior in Male African Elephants. PLoS ONE 2012, 7, e31382. [Google Scholar] [CrossRef]
- LaDue, C.A.; Vandercone, R.P.G.; Kiso, W.K.; Freeman, E.W. Social Behavior and Group Formation in Male Asian Elephants (Elephas maximus): The Effects of Age and Musth in Wild and Zoo-Housed Animals. Animals 2022, 12, 1215. [Google Scholar] [CrossRef]
- Lee, P.C.; Poole, J.H.; Njiraini, N.; Sayialel, C.N.; Moss, C. Male Social Dynamics-Independence and Beyond. In The Amboseli Elephants: A Long-Term Perspective on a Long-Lived Mammal; University of Chicago Press: Chicago, IL, USA, 2011; pp. 260–271. [Google Scholar]
- Srinivasaiah, N.; Kumar, V.; Vaidyanathan, S.; Sukumar, R.; Sinha, A. All-Male Groups in Asian Elephants: A Novel, Adaptive Social Strategy in Increasingly Anthropogenic Landscapes of Southern India. Sci. Rep. 2019, 9, 8678. [Google Scholar] [CrossRef]
- Chiyo, P.I.; Wilson, J.W.; Archie, E.A.; Lee, P.C.; Moss, C.J.; Alberts, S.C. The Influence of Forage, Protected Areas, and Mating Prospects on Grouping Patterns of Male Elephants. Behav. Ecol. 2014, 25, 1494–1504. [Google Scholar] [CrossRef]
- Sukumar, R. Ecology of the Asian Elephant in Southern India. II. Feeding Habits and Crop Raiding Patterns. J. Trop. Ecol. 1990, 6, 33–53. [Google Scholar] [CrossRef]
- Chiyo, P.I.; Cochrane, E.P. Population Structure and Behaviour of Crop-Raiding Elephants in Kibale National Park, Uganda. Afr. J. Ecol. 2005, 43, 233–241. [Google Scholar] [CrossRef]
- Murphy, D.; Mumby, H.S.; Henley, M.D. Age Differences in the Temporal Stability of a Male African Elephant (Loxodonta africana) Social Network. Behav. Ecol. 2019, 31, 21–31. [Google Scholar] [CrossRef]
- Hartley, M.; Wood, A.; Yon, L. Facilitating the Social Behaviour of Bull Elephants in Zoos. Int. Zoo Yearb. 2019, 53, 62–77. [Google Scholar] [CrossRef]
- Ananth, D. Juvenile Musth. Zoos’ Print J. Rev. 2000, XV, 259–262. [Google Scholar] [CrossRef]
- Keerthipriya, P.; Nandini, S.; Vidya, T.N.C. Effects of Male Age and Female Presence on Male Associations in a Large, Polygynous Mammal in Southern India: The Asian Elephant. Front. Ecol. Evol. 2021, 9, 616666. [Google Scholar] [CrossRef]
- Dobson, S. Competition for Mates and Predominant Juvenile Male Dispersal in Mammals. Anim. Behav. 1982, 30, 1183–1192. [Google Scholar] [CrossRef]
- Keerthipriya, P.; Nandini, S.; Gautam, H.; Revathe, T.; Vidya, T.N.C. Musth and Its Effects on Male–Male and Male–Female Associations in Asian Elephants. J. Mammal. 2019, 101, 259–270. [Google Scholar] [CrossRef]
- Evans, K.E.; Harris, S. Adolescence in Male African Elephants, Loxodonta africana, and the Importance of Sociality. Anim. Behav. 2008, 76, 779–787. [Google Scholar] [CrossRef]
- Goldenberg, S.Z.; de Silva, S.; Rasmussen, H.B.; Douglas-Hamilton, I.; Wittemyer, G. Controlling for Behavioural State Reveals Social Dynamics among Male African Elephants, Loxodonta africana. Anim. Behav. 2014, 95, 111–119. [Google Scholar] [CrossRef]
- Lee, P.C.; Moss, C.J. African Elephant Play, Competence and Social Complexity. Anim. Behav. Cogn. 2014, 2, 144. [Google Scholar] [CrossRef]
- Sukumar, R. A Brief Review of the Status, Distribution and Biology of Wild Asian Elephants Elephas maximus. Int. Zoo Yearb. 2006, 40, 1–8. [Google Scholar] [CrossRef]
- Joshi, R. Tusker’s Social Bonds in Rajaji. Hystrix 2015, 26, 41–45. [Google Scholar] [CrossRef]
- Jainudeen, M.R.; Katongole, C.B.; Short, R.V. Musth and Sexual Activity in the Male Asiatic Elephant, Elephas maximus. J. Reprod. Fertil. 1971, 29, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Poole, J.H. Announcing Intent: The Aggressive State of Musth in African Elephants. Anim. Behav. 1989, 37, 153–155. [Google Scholar] [CrossRef]
- LaDue, C.A.; Schulte, B.A.; Kiso, W.K.; Freeman, E.W. Musth and Sexual Selection in Elephants: A Review of Signalling Properties and Potential Fitness Consequences. Behaviour 2021, 159, 207–242. [Google Scholar] [CrossRef]
- Lincoln, G.A.; Ratnasooriya, W.D. Testosterone Secretion, Musth Behaviour and Social Dominance in Captive Male Asian Elephants Living near the Equator. J. Reprod. Fertil. 1996, 108, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Slotow, R.; Van Dyk, G.; Poole, J.; Page, B.; Klocke, A. Older Bull Elephants Control Young Males. Nature 2000, 408, 425–426. [Google Scholar] [CrossRef]
- Mitani, J.C.; Merriwether, D.A.; Zhang, C. Male Affiliation, Cooperation and Kinship in Wild Chimpanzees. Anim. Behav. 2000, 59, 885–893. [Google Scholar] [CrossRef]
- Lahdenperä, M.; Mar, K.U.; Lummaa, V. Nearby Grandmother Enhances Calf Survival and Reproduction in Asian Elephants. Sci. Rep. 2016, 6, 27213. [Google Scholar] [CrossRef]
- McComb, K.; Moss, C.; Durant, S.M.; Baker, L.; Sayialel, S. Matriarchs as Repositories of Social Knowledge in African Elephants. Science 2001, 292, 491–494. [Google Scholar] [CrossRef]
- Ward, E.J.; Parsons, K.; Holmes, E.E.; Balcomb, K.C.; Ford, J.K. The Role of Menopause and Reproductive Senescence in a Long-Lived Social Mammal. Front. Zool. 2009, 6, 4. [Google Scholar] [CrossRef]
- Seltmann, M.W.; Helle, S.; Adams, M.J.; Mar, K.U.; Lahdenperä, M. Evaluating the Personality Structure of Semi-Captive Asian Elephants Living in Their Natural Habitat. R. Soc. Open Sci. 2018, 5, 172026. [Google Scholar] [CrossRef] [PubMed]
- Freeman, E.W.; Schulte, B.A.; Brown, J.L. Using Behavioral Observations and Keeper Questionnaires to Assess Social Relationships among Captive Female African Elephants. Zoo Biol. 2010, 29, 140–153. [Google Scholar] [CrossRef] [PubMed]
- Hambrecht, S.; Reichler, S. Group Dynamics of Young Asian Elephant Bulls (Elephas maximus Linnaeus, 1758) in Heidelberg Zoo—Integration of a Newcoming in an Established Herd. Zool. Gart. 2013, 82, 267–292. [Google Scholar] [CrossRef]
- Finch, K.; Sach, F.; Fitzpatrick, M.; Masters, N.; Rowden, L.J. Longitudinal Improvements in Zoo-Housed Elephant Welfare: A Case Study at ZSL Whipsnade Zoo. Animals 2020, 10, 2029. [Google Scholar] [CrossRef]
- Mason, G.J.; Veasey, J.S. How Should the Psychological Well-Being of Zoo Elephants Be Objectively Investigated? Zoo Biol. 2010, 29, 237–255. [Google Scholar] [CrossRef]
- Yon, L.; Williams, E.; Harvey, N.D.; Asher, L. Development of a Behavioural Welfare Assessment Tool for Routine Use with Captive Elephants. PLoS ONE 2019, 14, e0210783. [Google Scholar] [CrossRef]
- Meehan, C.L.; Hogan, J.N.; Bonaparte-Saller, M.K.; Mench, J.A. Housing and Social Environments of African (Loxodonta africana) and Asian (Elephas maximus) Elephants in North American Zoos. PLoS ONE 2016, 11, e0146703. [Google Scholar] [CrossRef]
- Prado-Oviedo, N.A.; Bonaparte-Saller, M.K.; Malloy, E.J.; Meehan, C.L.; Mench, J.A.; Carlstead, K.; Brown, J.L. Evaluation of Demographics and Social Life Events of Asian (Elephas maximus) and African Elephants (Loxodonta africana) in North American Zoos. PLoS ONE 2016, 11, e0154750. [Google Scholar] [CrossRef]
- Schreier, A.; Readyhough, T.S.; Moresco, A.; Davis, M.; Joseph, S. Social Dynamics of a Newly Integrated Bachelor Group of Asian Elephants (Elephas maximus): Welfare Implications. J. Appl. Anim. Welf. Sci. 2021. [Google Scholar] [CrossRef]
- Thevarajah, S.J.; Readyhough, T.S.; Davis, M.; Moresco, A.; Joseph, S.; Schreier, A.L. Nighttime Behavior and the Length of Social Relationships in Male Asian Elephants. J. Appl. Anim. Welf. Sci. 2021. [Google Scholar] [CrossRef]
- Readyhough, T.S.; Joseph, S.; Davis, M.; Moresco, A.; Schreier, A.L. Impacts of Socialization on Bull Asian Elephant (Elephas maximus) Stereotypical Behavior. J. Zool. Bot. Gard. 2022, 3, 113–130. [Google Scholar] [CrossRef]
- Altmann, J. Observational Study of Behavior: Sampling Methods. Behaviour 1974, 49, 227–267. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. JOSS 2019, 4, 1686. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2015, 67, i01. [Google Scholar] [CrossRef]
- R: A Language and Environment for Statistical Computing; version 4. 2.2; R Core Team: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 13 December 2022).
- Posit Team. RStudio: Integrated Development Environment for R; Posit Software; PBC: Boston, MA, USA, 2022; Available online: http://www.posit.co/ (accessed on 13 December 2022).
- Hamilton, W.D. The Genetical Evolution of Social Behaviour. II. J. Theor. Biol. 1964, 7, 17–52. [Google Scholar] [CrossRef]
- Meehan, C.L.; Mench, J.A.; Carlstead, K.; Hogan, J.N. Determining Connections between the Daily Lives of Zoo Elephants and Their Welfare: An Epidemiological Approach. PLoS ONE 2016, 11, e0158124. [Google Scholar] [CrossRef]
- Scott, N.L.; LaDue, C.A. The Behavioral Effects of Exhibit Size versus Complexity in African Elephants: A Potential Solution for Smaller Spaces. Zoo Biol. 2019, 38, 448–457. [Google Scholar] [CrossRef]
- Schmid, J.; Heistermannt, M.; GansloBer, U.; Hodges, J.K. Introduction of Foreign Female Asian Elephants (Elephas maximus) into an Existing Group: Behavioural Reactions and Changes in Cortisol Levels. Anim. Welf. 2001, 10, 357–372. [Google Scholar] [CrossRef]
- Blackett, T.A.; McKenna, C.; Kavanagh, L.; Morgan, D.R. The Welfare of Wild Animals in Zoological Institutions: Are We Meeting Our Duty of Care? Int. Zoo Yearb. 2017, 51, 187–202. [Google Scholar] [CrossRef]
- Rees, P.A. The Sizes of Elephant Groups in Zoos: Implications for Elephant Welfare. J. Appl. Anim. Welf. Sci. 2009, 12, 44–60. [Google Scholar] [CrossRef]
- Cameron, E.Z.; Ryan, S.J. Welfare at Multiple Scales: Importance of Zoo Elephant Population Welfare in a World of Declining Wild Populations. PLoS ONE 2016, 11, 22–25. [Google Scholar] [CrossRef] [PubMed]


| Behavior Category | Behavior | Definition | |
|---|---|---|---|
| Agonistic | Non-Contact | Approach head high | Actor moves toward recipient to within two body lengths with head above shoulders and ears out perpendicular |
| Charge | Rapid forward lunging or rapid gait by actor towards a stationary conspecific starting from more than two body lengths away | ||
| Chase | Actor rapidly pursues recipient, who is moving away from actor | ||
| Head shake | Actor holds head above shoulders and moves vigorously from side to side, up and down, or in circular motion | ||
| Supplant | Actor approaches to within two body lengths of conspecific without making contact, causing recipient to turn away or yield ground | ||
| Contact ** | Grasp tail | Actor places tail of conspecific into its own trunk while recipient attempts to move away from focal animal | |
| Kick | Actor strikes at recipient with rear limb | ||
| Mount | Actor rears up on hind legs and places forelegs on recipient | ||
| Push | Actor contacts conspecific with enough force to displace recipient | ||
| Spar | Two elephants mutually and simultaneously push one another backwards with force with heads and/or heads and trunks | ||
| Trunk over back | Actor places 2/3 or more of its trunk firmly over the back or head of a conspecific | ||
| Affiliative | Approach relaxed | Actor moves to within to within two body lengths of recipient with head low and ears lying flat against its head, not associated with any other behavior | |
| Body contact | Body contact unspecified in any other behavior (e.g., side-to-side rubbing or touching) | ||
| Play | Actor voluntarily spars, wrestles with, mounts, or chases recipient without obvious intent to do harm or display dominance; does not include when following agonistic interaction | ||
| Shares food/object | Actor either feeds or uses an object in concert with another elephant that is within one body length | ||
| Trunk tangle | Actor loosely entwines its trunk with that of recipient | ||
| Trunk to mouth | Actor places its trunk in another elephant’s mouth | ||
| Trunk touch/toward | Actor extends trunk toward recipient with or without touching; not associated with any other behavior | ||
| Submissive | Allow | Actor remains still and calmly permits physical contact by conspecific, including genital investigation | |
| Back into/toward | Actor takes two steps (minimum) backward towards another elephant to within one body length, with or without touching | ||
| Lower head or ears | Actor quickly drops head and/or ears in response to approach by another elephant | ||
| Run away | Actor flees from conspecific in response to its agonistic contact, display, or approach | ||
| Turn away/yield | Actor turns body away from or yields ground as a result of actions or encroachment by another elephant | ||
| Other | Bathe/swim | Actor lies, stands, or submerges in pool (includes spraying water on self); not associated with any other ethogram behavior | |
| Drink | Actor uses trunk to bring water to its mouth and drink | ||
| Dust/mud | Actor uses trunk to throw dirt, sand, shavings, or mud onto body while standing | ||
| Enrichment interaction | Actor interacts with provided non-food enrichment items | ||
| Feed | Actor ingests presented diet items; includes manipulating food items | ||
| Follow | Actor closely trails behind recipient, who is moving away from actor (at normal walking speed) | ||
| Genital investigation | Actor sniffs or touches genitals of another elephant with its trunk | ||
| Locomotion | Actor moves directionally along a horizontal surface (not while feeding); can include slow or fast walking or running | ||
| Rest | Stationary; lying down or standing with trunk resting loosely on the ground; eyes open or closed; not performing any other behavior | ||
| Stereotypy | Actor performs stereotypic behavior including head-bobbing or pacing | ||
| Wallow | Actor lies or rolls in mud or dirt | ||
| Other | Actor performs any behavior not on ethogram | ||
| Out of View | Out of view | Actor cannot be seen or cannot be distinguished from other elephants | |
| Variable | Description | Reference Level |
|---|---|---|
| Affiliative | Binary variable indicating if the focal animal was engaging in affiliative behavior (0 = no, 1 = yes) | NA—response variable |
| ContactAg | Binary variable indicating if the focal animal was engaging in contact agonistic behavior (0 = no, 1 = yes) | NA—response variable |
| NonContactAg | Binary variable indicating if the focal animal was engaging in non-contact agonistic behavior (0 = no, 1 = yes) | NA—response variable |
| Submissive | Binary variable indicating if the focal animal was engaging in submissive behavior (0 = no, 1 = yes) | NA—response variable |
| SocialAgeLittles | Categorical variable indicating the composition of the social group (Adolescents, Mixed, Mature) excluding the focal animal | Adolescents |
| SocialAgeOlder | Categorical variable indicating the composition of the social group (Adolescents, Mixed, Mature) excluding the focal animal | Mature |
| AccessArea | Continuous variable indicating the size of the area that the focal animal had access to (per 1000 ft2); 2.00–47.37 | 2000 ft2 |
| InOutAccess | Categorical variable indicating if focal animal had access inside (in), outside (out), or both (both) | Both |
| Musth | Categorical variable indicating which elephant (if any) was in musth during the time of the observation session | None |
| TimeOfDay | Categorical variable indicating if observations took place in the morning, afternoon, or night | Morning |
| IntrosPeriod | Binary variable indicating if the observations took place during the 5 mo introductory period (1) or after (0) | After Introductions |
| NewSocial | Binary variable indicating if the social group included novel individuals (1) or only established social partners (0) | Established |
| InOutAccess*AccessArea | Interaction term between InOutAccess (inside, outside, both) and AccessArea | Both:GroupSize |
| IntrosPeriod*NewSocial | Interaction term between IntrosPeriod (0 = after, 1 = during) and NewSocial (0 = established, 1 = new) | After:Established |
| SessionID | Categorical variable identifying the specific focal session that a scan occurred during | N/A—(used as a random effect) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Readyhough, T.S.; Davis, M.; Joseph, S.; Moresco, A.; Schreier, A.L. Age and Social History Impact Social Interactions between Bull Asian Elephants (Elephas maximus) at Denver Zoo. J. Zool. Bot. Gard. 2023, 4, 191-213. https://doi.org/10.3390/jzbg4010018
Readyhough TS, Davis M, Joseph S, Moresco A, Schreier AL. Age and Social History Impact Social Interactions between Bull Asian Elephants (Elephas maximus) at Denver Zoo. Journal of Zoological and Botanical Gardens. 2023; 4(1):191-213. https://doi.org/10.3390/jzbg4010018
Chicago/Turabian StyleReadyhough, Taylor S., Maura Davis, Sharon Joseph, Anneke Moresco, and Amy L. Schreier. 2023. "Age and Social History Impact Social Interactions between Bull Asian Elephants (Elephas maximus) at Denver Zoo" Journal of Zoological and Botanical Gardens 4, no. 1: 191-213. https://doi.org/10.3390/jzbg4010018
APA StyleReadyhough, T. S., Davis, M., Joseph, S., Moresco, A., & Schreier, A. L. (2023). Age and Social History Impact Social Interactions between Bull Asian Elephants (Elephas maximus) at Denver Zoo. Journal of Zoological and Botanical Gardens, 4(1), 191-213. https://doi.org/10.3390/jzbg4010018