1. Introduction
Humans have shaped natural ecosystems for thousands of years [
1] affecting the distribution and richness of wildlife communities [
2,
3]. Livestock grazing is one of the most common human activities globally [
4] and is used in many animal husbandry systems. Transhumance is a traditional stock rearing system in Europe, dating back to ancient times [
5], where livestock exploits the upland pastures during the summer period and are moved to lowland pastures in the winter [
6,
7,
8]. In South Europe grazing by small ruminants shaped the grasslands of the pseudo-alpine zone for centuries [
6]. Nowadays, transhumance is considered a valuable practice for the management and conservation of these fragile ecosystems [
9], as it involves many ecologically important management techniques that preserve the better exploitation of rangelands [
10].
The impacts of grazing on wildlife have been widely studied (e.g., [
11,
12,
13]), but they are still not well-understood. Thus, they are not universal and affect wildlife in various ways [
12]. Grazing directly affects the basic wildlife habitat elements and specifically the vegetation cover, the food and the space, with both overwhelming and detrimental effects on wildlife populations [
12,
14]. Grazing is considered a useful land management and conservation tool [
13,
15], because it increases the structural diversity of vegetation and, consequently, the number of suitable microhabitats for a great spectrum of wildlife species that inhabit open landscapes [
16,
17]. However, both high-stocking rates and abandonment of grazing threaten grassland wildlife species [
13]. Overgrazing negatively affects the survival and growth of vegetation, which in turn leads to the reduction of suitable cover and food for wildlife and especially for avifauna [
12]. Overgrazing can decrease the protective shrub cover that might lead to a decrease of arthropods, the main food for many bird species, and to decreased nest cover, which leads to increased nest losses [
18]. Moreover, high densities of herbivores may decrease the nesting success of ground nesting birds through trampling [
13,
19,
20]. However, high stocking rates of sheep in Northern Europe showed positive effect on the density of alpine birds [
21]. On the other hand, grazing abandonment leads to shrub encroachment into grasslands, thus making unfavorable conditions for open habitat bird species [
13]. Nevertheless, the short-term abandonment can have positive effect on those species as it prevents the high encroachment of shrubs [
14].
Birds are widely used as indicators to monitor the effects of livestock grazing [
14,
22,
23] due to the wide range of niches they use, their high mobility, and their easy monitoring [
24,
25]. Grassland birds are highly associated with special vegetation characteristics derived from different management practices [
26], with some species being particularly susceptible to habitat changes resulting from grazing regime [
27,
28,
29]. Grazing intensity [
30] and the type of grazing animal [
29] affect bird species in different ways, through the shaping of vegetation structure and habitat quality [
3,
12,
18,
31]. Livestock, through its selective grazing, creates structurally heterogeneous grasslands [
13,
32]. Some researchers revealed that some bird species, in particular those feeding and nesting on the ground [
25], as well as shrub associated species [
18], were negatively affected by increasing grazing intensity [
33]. Other studies showed that increased grazing benefited ground nesting and foraging bird species, whereas it negatively affected shrub-associated species [
27]. These contradictory findings support the idea that grassland bird species require a mosaic of habitats because each species preferences varies widely from tall and dense to short and sparse vegetation [
12,
34,
35]. Also, bird species that share common ecological traits and life histories tend to respond in the same way to ecological changes [
36]. Consequently, conservationists may use grazing to diversify grasslands in favor of grassland bird species [
12,
35].
Meanwhile, during the last decades the socioeconomic changes in Europe affected the rural population activities with cascade effects on the landscape [
37,
38] and consequently, on the biodiversity [
39]. Human activities are responsible for a great loss of global biodiversity [
40,
41] severely affecting the alpine ecosystems [
42] and the alpine wildlife populations changed through habitat modifications [
43]. Nevertheless, a long-term study on Scandinavian mountains revealed that grazing by Reindeers (
Rangifer tarandus) held the grassland habitats stable over the years leading to a stable alpine bird community with fluctuations for some species [
44]. Transhumance is a deterministic force shaping natural alpine and pseudo-alpine grasslands for centuries but has disappeared in most European countries without extensive research of its ecological effects [
10]. However, in South Europe, transhumance still exists albeit with a declining trend [
45], leading to progressive woody vegetation encroachment [
46] in alpine grasslands with consequences on avian communities [
45].
The relationship of transhumance with wildlife has received little attention [
45] and the consequences of transhumance abandonment and its different stocking rates on bird communities of the pseudo-alpine zone have not been studied yet at all. Therefore, it is important to understand the effects of transhumant livestock grazing on birds in the fragile pseudo-alpine grassland ecosystems. Within this context, our study aimed to examine how avian communities respond under different grazing regimes caused by transhumant livestock in pseudo-alpine zone. This study focused on natural grasslands occupying the pseudo-alpine zone with and without grazing transhumant herds during the summer period, whose structural vegetation characteristics evolved due to the presence and absence of livestock, respectively. The way that avian communities respond to habitat variables was analyzed in order to highlight the importance of transhumance in maintaining high quality habitats for the conservation of the threatened pseudo-alpine grassland avifauna.
4. Discussion
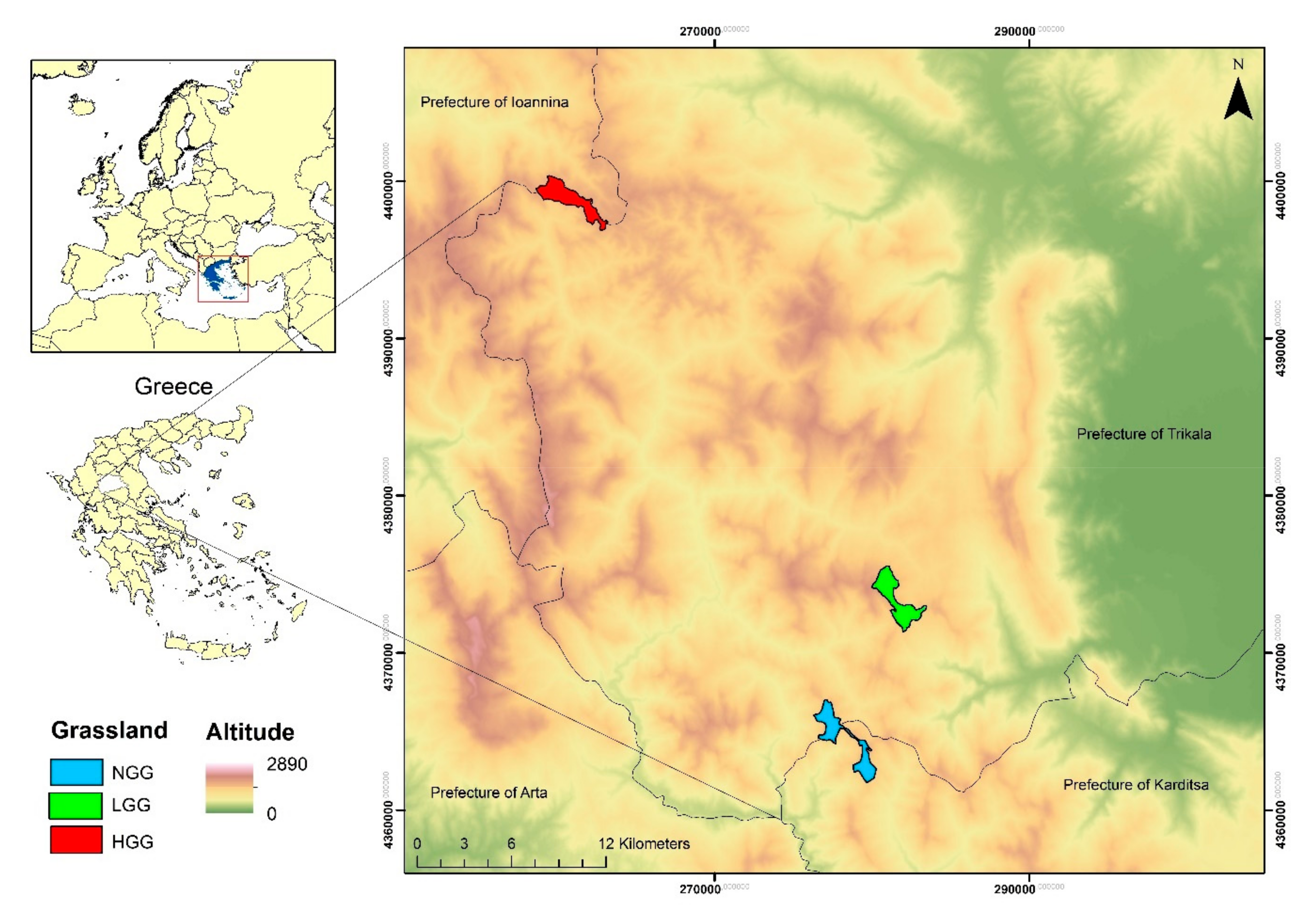
Avian communities were surveyed in representative pseudo-alpine grassland ecosystems of the South Pindos Mountains in central Greece. These grasslands were located above the tree line and selected according to the different levels of grazing transhumant livestock. Significant differences were found among the surveyed grasslands in regard to vegetation structure and the topography. Herbaceous vegetation height, heterogeneity of vegetation height and percent of rock cover were the key environmental variables that influenced bird species composition in the three grasslands. Our results indicate that there is a significant difference in the bird communities among the three studied grasslands. This is evident in the different bird species present in the three grasslands as well as in the differences of their diversity indicators (Smean, A, H′mean, S, WS, H′, Evar).
The differences in the vegetation structure of the grasslands would appear to mainly be a result of different levels of grazing transhumant livestock, which consequently affected the bird communities. Given that our findings are based on the interviews of the shepherds and the counting of their herds, the results from the analyses concerning the effects of grazing intensity should be treated with caution. Further experimental investigation is needed to estimate the grazing intensity by taking into account the possible impact of wild herbivores. In the lower altitude subalpine forests neighbor to our study grasslands, occur Roe Deers (
Capreolus capreolus), which are selective solitary browsers and probably use the grasslands as feeding grounds. Therefore, future studies should deal with the estimation of grazing intensity through the counting of dung [
59] of both livestock and wild herbivores in the pseudo-alpine grasslands. Also, differences in the interaction between soil nutrients and grazing may affect the vegetation structure of grasslands. It has been reported that heavy grazing reduces the herbaceous vegetation and soil nutrients because herbivores compress the soil, remove the vegetation biomass and as a result they reduce the amount of litter on the soil [
60]. Vegetation height was higher in NGG followed by LGG and HGG, while the heterogeneity of vegetation height was higher in LGG, followed by the NGG and the HGG.
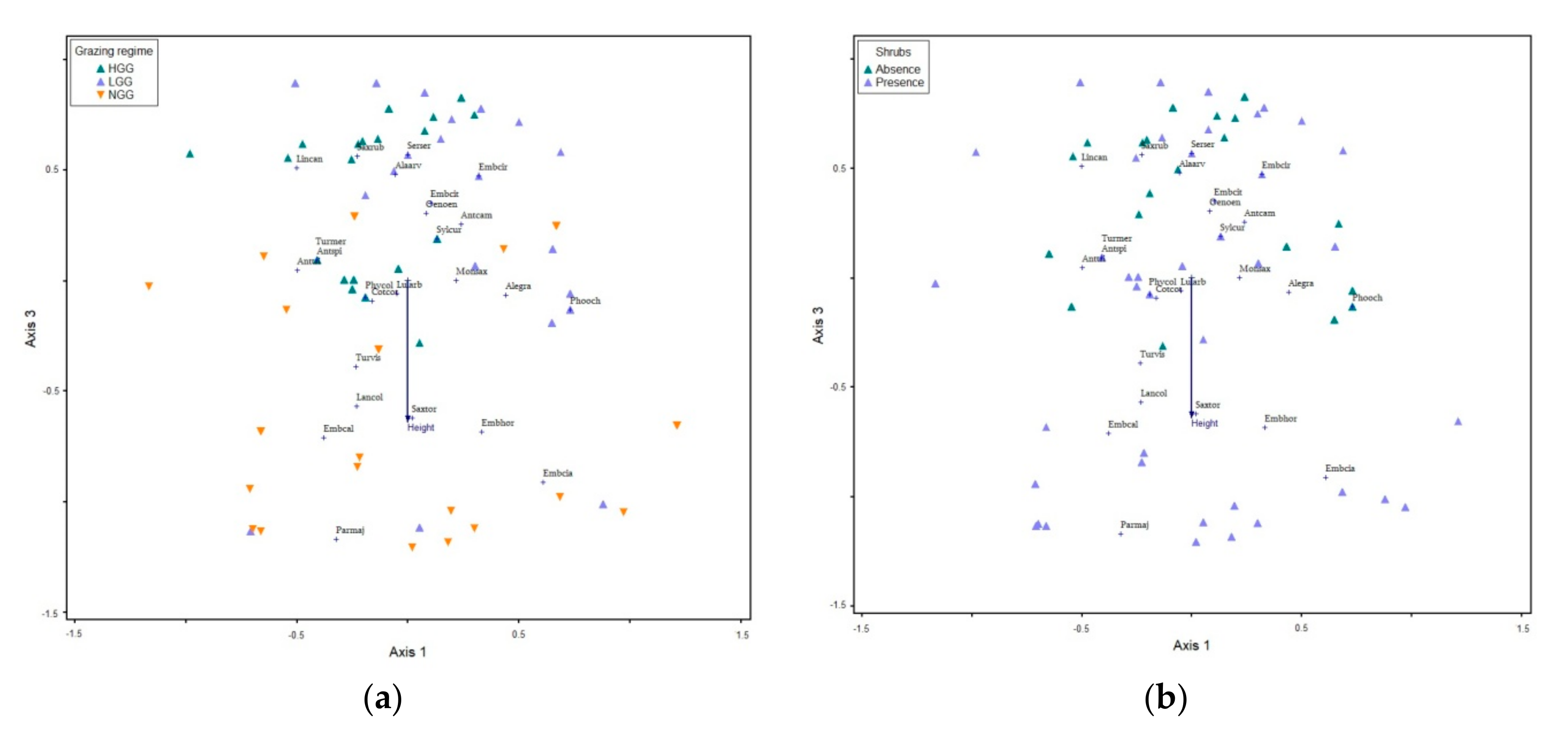
Interestingly, our results showed that bird species richness and bird diversity (alpha diversity) were affected only by the environmental and not by the topographic variables. Specific variations in the environmental characteristics, such as the difference in recorded vegetation height, vegetation height heterogeneity and rock cover were demonstrated to determine differences in bird species composition among the studied grasslands. Bird species richness and bird diversity corresponded positively to a decrease of vegetation height and vegetation height heterogeneity and to an increase of the percentage of rock cover. Vegetation height has been the main factor associated with avian diversity and composition in grasslands [
61,
62]. This difference in vegetation height demonstrates the importance of grassland mosaics with vegetation of different heights for grassland bird diversity. The analysis of gamma diversity in our study area showed that many grassland birds require a mosaic landscape with different habitat patches to satisfy their breeding demands and to improve their foraging conditions [
35]. Grazing increases the structural diversity and constructs variable microhabitats for a great proportion of species [
3,
17,
63]. The relatively low vegetation height derived from the seasonal grazing probably improved the nesting and foraging conditions for grassland species [
16,
21]. Transhumance grazing holds vegetation height relatively low thus increasing food accessibility [
64,
65], locomotion on the ground and offering higher efficiency in predator detection [
66]. Moreover, presence of livestock dung may increase the available invertebrates [
67], which in turn may attract valuable numbers of birds. Also, the presence of rocks on those high-altitude grasslands attracts rock dueling species, which nest among the boulders and forage on the grazed areas thus increasing the total species diversity. The presence of shrubs may also have ambivalent effects on grassland birds. A study conducted in arctic riparian plains in Norway showed that intense grazing by Reindeers led to shrub loss which affected negatively both shrub and open associated bird species [
18]. This trend is probably associated with the decrease of nest and cover habitat, and insects, which are a primary food source for many species [
18,
68].
Avian communities are affected by the presence of transhumant livestock in the pseudo-alpine grassland ecosystems of South Pindos Mountains through the differences in vegetation structure. Bird communities among the two grazed grasslands (LGG and HGG) showed a lower separation than the communities between the two grazed (LGG and HGG) and the non-grazed (NGG) one. Transhumance is highlighted as an ecological management technique that shapes and maintains stable and dynamic the upland grasslands, while overcoming the plethora of ecological problems derived by intensive, sedentary livestock activities in high altitude ecosystems [
8,
10]. The seasonal exploitation of grasslands, the local livestock breeds that have adapted to the special conditions of the uplands and the shepherding exert low pressure on grasslands hence improves the nesting and foraging conditions for the threatened pseudo-alpine avifauna. A pan-European review and meta-analysis showed that the traditional practices and short-term abandonment of grazing are of high importance for the enhancement of richness and diversity [
14]. Compared to high intensity grazing, these low intensity systems resulted in higher richness and diversity. Furthermore, traditionally grazed grasslands were richer in arthropods than overgrazed; arthropods compose the diet for the majority of bird species inhabiting high altitude grasslands. However, the high intensity grazing was shown to benefit open habitat species due to the prevention of woody vegetation encroachment [
14]. Additionally, the short-term abandonment was shown to have similar positive effect on biodiversity. As a result, the pausing of grazing for longer periods may enhance the encroachment of woody vegetation and affect negatively the open habitat species [
14].
The majority of birds inhibiting the studied pseudo-alpine grasslands are ground-dwelling species (
Table A1). The low vegetation height of the HGG was reflected by the greater number of indicator bird species (6) compared to the LGG (1) and NGG (2). The presence of two indicator species in NGG which are under conservation status also highlights the importance of no grazing areas. The Eurasian Skylark, the Yellowhammer, the Whinchat, the Woodlark, the Linnet, and the Northern Wheatear which are SPEC, showed great preferences for highly grazed grasslands. The Eurasian Skylark, a typical ground-associated grassland bird, showed a positive response to transhumance. Eurasian Skylarks avoid tall and dense vegetation [
49,
69] due to the difficulties they face in foraging and movement. Higher grazing pressure leads to highly exploited areas with low vegetation height and vertical structure, but favoring feeding opportunities in many birds by making food easy to access. Furthermore, the Yellowhammer and the Whinchat, two upland species highly associated to open areas and especially grasslands in Greece [
70], were positively affected by grazing due to their requirements for low herbaceous vegetation height. In our study area both species were absent from NGG and their higher concentrations were observed on HGG, indicating a positive effect of grazing on their numbers. A similar response was observed in the Woodlark, a species that prefers grazed pastures in order to benefit by low vegetation for breeding and foraging. Moreover, rock presence in grasslands is responsible for the occurrence of rock-dwelling species, hence it increases the number of species present. The Northern Wheatear is a typical species of stony high-altitude grasslands. It needs areas of low vegetation in order to feed [
71], and fallen rocks, boulders, and screes as nesting locations [
72], hence the combination of grazed sites with rocky areas favored the occurrence of the species in high grazing intensity pastures. On the other hand, only one bird species of high conservation value, the Tawny Pipit, was found to benefit from low grazing pressure in our study. The Tawny Pipit highly concentrated in LGG, resembling the habitat preferences of the species for high altitude stony pastures. The presence of a low number of small ruminants creates a heterogeneous vegetation mosaic with areas of low and tall herbaceous vegetation, which offers suitable foraging and nesting sites for the species respectively [
73]. Bird community in NGG showed a great separation from those in both grazed grasslands with two species, the Rock and the Corn Bunting, highly confined to those areas. The Rock Bunting was observed only in NGG and was probably favored by the presence of shrubs and the rock cover, which are important territory components of the species [
74]. Moreover, the Corn Bunting seemed to benefit from the presence of shrubs [
75,
76] in grasslands with absence of grazing.
Grassland bird species show different susceptibility to grazing disturbance [
35,
44,
77]. Our results demonstrate a separation of species between areas, bird species recorded in the NGG that depend on tall grass and shrubs were replaced by species associated with short grass in grazed grasslands. Specific management practices can satisfy the nesting and foraging requirements of a group of species, but cannot benefit others [
35,
78]. Areas with low or no grazing are necessary to ensure viable populations of tall grass [
14,
35,
79] and shrub associated species [
14]. On the other hand, in grazed grasslands, a short grass assemblage of birds is benefited [
35,
77]. Our results demonstrate that presence of both grazed and non-grazed grassland areas is essential for the conservation of the pseudo-alpine grassland birds as also observer by Isacch and Cardoni [
79]. Moreover, some researchers have proposed that some areas should not be managed, for obtaining the maximum diversity of grassland birds [
35,
78]. Thus, this habitat heterogeneity is necessary to ensure species coexistence and maintain the highest diversity of grassland bird communities [
18,
35,
62,
80,
81].
However, we are aware that our research may have some limitations. The first is that it is a single-year study and offers no long-term results on the effects of pastoral activities on bird communities. The second is the small number of survey points as a consequence of the small area and the harsh terrain of the grasslands under study, and the third is the absence of replicate sites of the NGG, LGG, and HGG.

 ,
, 


{kind=link}
{kind=link}

