1. Introduction
Electroencephalography (EEG) can be used to investigate the pain process in animals by assessing the electrical activity of the cerebral cortex in response to noxious stimulation [
1]. The raw EEG data recorded are the summation of the electrical activity occurring over time (i.e., the time domain). Fast Fourier transformation (FFT), an algorithm used to transform data from the time domain to the frequency domain, is then applied to the raw EEG. Useful variables can then be obtained and analysed. The three variables most used in pain and anaesthetic depth assessment are the median frequency (
F50), below which half the total power of the EEG is located; the spectral edge frequency (
F95), below which 95% of the total power is located; and the total power (
Ptot), the total signal power of the spectrum assessed. Certain changes in response to a noxious stimulus can then be examined for their potential use as indicators of pain perception [
2,
3].
Examining an animal’s response to pain in the laboratory can be done with the aid of general anaesthesia. In a conscious animal the application of noxious stimuli would result in the experience of pain. General anaesthesia, and therefore unconsciousness, significantly reduces the potential for stress for the animal. Another principal advantage is the capacity to use a non-analgesic control group, considered a key component of successful animal pain studies, without negatively impacting the animals’ welfare [
4]. Many of these animal studies have employed an anaesthetic protocol known as the Minimal Anaesthesia Model (MAM) [
3]. Here, the animal is anaesthetised to a point at which they are immobile and unconscious, and therefore not perceiving pain, but maintain cortical responsiveness to noxious stimuli.
The use of anaesthesia is well-recognised to affect brain activity as measured by EEG [
5]. Early studies in both humans and animals indicated a general depressant effect of anaesthesia on the EEG [
6,
7]. The nature and extent to which this depression occurs has been shown to depend on species, dose, time and agent [
8,
9,
10,
11].
Three of the commonly assessed frequency distribution variables of transformed EEG data,
F50,
F95 and
Ptot, change during nociception and with alterations of anaesthetic concentration in many mammalian species and humans [
3,
12,
13,
14]. There are two published descriptions of how these variables change in birds during anaesthesia, specifically in chickens during sevoflurane, isoflurane, methoxyflurane and halothane anaesthesia at different concentrations [
9,
10]. Both report the frequency variables changing in a similar manner as seen in mammals.
When considering the suitability of an anaesthetic agent for use in the MAM, the degree of cortical depression caused by that agent must be assessed. A reduction in cortical responsiveness is indicated by increasing levels of burst suppression in the EEG recording [
15]. Burst suppression is characterised by periods of isoelectric brain activity, equating to a functionally quiet cortex, interspersed with bursts of high-amplitude EEG [
16].
To allow comparisons between anaesthetic agents, a method of quantifying an equipotent dose is required. For the volatile or inhalant anaesthetic agents, such as halothane, this standardised dose is the minimum alveolar concentration (MAC) in animals with alveoli, and more correctly, the minimal anaesthetic concentration in those without, such as birds. Although halothane has been used extensively for mammals in the MAM, limited work has investigated the effects of anaesthesia on the EEG of birds [
9,
10].
Halothane anaesthesia has been demonstrated to produce negligible burst suppression and therefore minimal cortical depression in horses, rats and chickens at standardised anaesthesia concentrations [
8,
9,
10,
11]. Though it is becoming less readily available due to its known human toxicity profile and an undesirable safety profile in many species, halothane is still used in veterinary practice and remains on the World Health Organisations’ (WHO) list of essential medicines [
17]. Together these reasons have made halothane the agent of choice for the MAM, where it continues to be used in mammalian pain research [
13].
The level of cortical depression in chickens at different multiples of MAC was evaluated for several volatile anaesthetic agents [
9,
10]. Even the lowest concentration of isoflurane and sevoflurane anaesthesia evaluated produced marked burst suppression, indicating these modern agents alone are not suitable for use in the MAM. In contrast, halothane and methoxyflurane produced negligible burst suppression at all concentrations assessed. These results support the use of halothane and methoxyflurane in studies of avian pain where general anaesthesia is used.
Rock Doves (
Columbia livia) are a bird species commonly used in neuroscience research [
18,
19]. This work has produced advanced knowledge of Rock Dove physiology and neuroanatomy, making it a good avian model for application of the MAM. Such information is required for avian pain investigations using the MAM, as an EEG response to noxious stimulation in halothane-anaesthetised chickens has not been found [
20]. Ornithological studies may be required to provide analgesia to birds, and this study can provide founding data to inform this analgesic choice. As yet, no studies using Rock Doves in the MAM with any anaesthetic agent have been completed. To allow the Rock Dove to be used as the avian model in the MAM, the level of cortical depression of halothane anaesthesia must first be determined.
The aim of the current study was to assess the effect of different concentrations of halothane on the EEG responses of the Rock Dove. The aim of the current study was to assess the effects of different concentrations of halothane on the EEG responses of the Rock Dove. It is hypothesized that pigeons will have EEG responses to halothane, indicating minimal cortical depression, similar to that found in chickens [
9]. The following predictions were tested: (1) Rock Doves will have minimal burst suppression evident during the three tested halothane concentrations (1×, 1.5×, 2× MAC); (2) Rock Doves will show a linear dose–response to increasing halothane concentrations in both median frequency (
F50) and total power (
Ptot), with the
F50 decreasing and the
Ptot increasing as the halothane concentration increases; (3) Rock Doves will show a biphasic response in
F95, with a decrease with the first increased halothane concentration (1.5× MAC), then a return to near base levels at the highest concentration (2× MAC).
2. Materials and Methods
2.1. Animals
All procedures were approved by the Massey University Animal Ethics Committee (protocol 18/90). Eight adult Rock Doves (Columba livia) were sourced from a free-living population at Wellington Zoo, Wellington, New Zealand.
An a priori calculation to determine an appropriate sample size was completed before the study began using data from McIlhone et al. [
9,
10]. The power analysis indicated that six birds would provide an 84% probability of detecting a 5% change in burst suppression rates between MAC multiples tested. Given the unknown response of Rock Dove EEG to anaesthetic agents, two additional birds were used.
2.2. Housing and Husbandry
After arrival, all birds were examined by a veterinarian and deemed to be healthy. Birds were weighed and leg-banded with coloured plastic bands (8 mm clip ring, fifth day, CAN, New Zealand) for identification. Prophylactic parasiticides were administered due to regular contact with wild birds and unknown parasite burden. The drugs administered via crop tube were 0.2 mg kg−1 moxidectin (Cydectin Long Acting Endectocide for Cattle and Sheep 1 mg mL−1, Zoetis, New Zealand) and 25 mg kg−1 toltrazuril (Baycox Coccidiocide for Piglets 50 mg mL−1, Bayer, New Zealand).
The birds were housed in a temperature- (18–22 °C) and light-controlled room on a 12 h light–dark cycle. Four birds were kept in one cage (102L × 79W × 177H cm, model 210BB, Avi One, NSW, Australia) and four birds in another (83.5L × 53.5W × 180.5H cm, model 604T XL, Avi One, NSW, Australia). The cages were lined with newspaper and fitted with lengths of pinewood as elevated roosting areas. Visual inspection was completed twice a day during morning feeding and evening cleaning.
During the lit hours of 8:00 a.m. to 8:00 p.m., a seed-based diet was provided ad libitum (source: Dr Colin Walker, Melbourne Bird Vet, via pigeonracingpigeon.com, accessed on 24 June 2018). Water was always available ad libitum. To reduce the zoonotic transmission risk of
Chlamydophila psittaci, the water was medicated with doxycycline at 180 mg L
−1 (Doxyvet, The Australian Pigeon Company, NSW, Australia). Millet sprays, green vegetables, birdseed bells and water-bathing opportunities were rotated regularly for enrichment. Snuffle mats stocked with sorghum seed were provided [
21]. Physical manipulation of the items in each cage was kept to a minimum to prevent unnecessary disturbance of the birds. Cage furniture and containers were washed daily and disinfected weekly.
2.3. Anaesthesia
Following seven days of acclimation, data collection was begun. One bird was anaesthetised each day, in an order randomised by simple blind selection. On the day of anaesthesia, the bird was caught with a towel and removed from the cage before feeding time. The bird was transported in a covered animal carrier to the neuroscience laboratory several floors below the housing. The bird was then weighed (Navigator, Ohaus, NJ, USA) and the crop confirmed empty by palpation.
A custom-made face mask with an eye shield was placed over the bird’s face and connected to a T-piece anaesthesia breathing system. Oxygen was supplied at a rate of 2 L min−1 for 60 s after which halothane (Halothane-Vet, Piramal Enterprises Limited, MH, India) was introduced incrementally. The vaporizer setting was increased by 1% (maximum 4%) every 60 s until spontaneous eyelid movement was abolished.
Tracheal intubation was performed with a 3.0 mm internal diameter polyvinyl chloride cuffed endotracheal tube (Jorvet, CO, USA) and connected to a small animal ventilator (SAV04, Vetronic Services, DEV, United Kingdom). The endotracheal tube cuffs were left uninflated. Following intubation, mechanical ventilation was immediately initiated. A 22-gauge 25 mm catheter (BD Insyte, Becton Dickinson Infusion Therapy, NJ, USA) was inserted into a medial metatarsal vein preferentially on the left side. Intravenous fluid therapy (5 mL kg−1 h−1, Hartmann’s solution) was delivered once the venous catheter was placed. A forced warm-air blower, infrared light and space blanket were used to minimise hypothermia.
All birds received 2.0 mg kg−1 IM meloxicam (Metacam Anti-Inflammatory Injectable for Dogs and Cats, Boehringer-Ingelheim, New Zealand) prior to anaesthesia being discontinued. Once fully recovered the birds were returned to their housing. As part of a further study, the birds were euthanised with intravenous pentobarbital when deeply anaesthetised with isoflurane at the end of the study.
2.4. Physiology and Anaesthesia Monitoring
A 5 Fr silicone feeding tube (Paediatric Feeding Tube, Mallinckrodt, SRY, United Kingdom) was used to sample respiratory gases by being passed through the side stream port of the endotracheal tube adaptor and situated in the distal tube. The feeding tube was connected to the sampling line of a multi-parameter monitor (Carescape B650 Anaesthetic Monitor; GE Healthcare, Finland) for continuous end-tidal agent and respiratory gas measurement at 200 mL min−1. This monitor was calibrated as per the manufacturer’s recommendations at regular intervals. An end-tidal halothane percent (FetHal) of 1.6–2.0% was used during anaesthetic stabilisation and the application of the monitoring equipment. An end-tidal carbon dioxide partial pressure (PetCO2) of 30–40 mmHg (4–5.3 kPa) was maintained via adjustments to the peak inspiratory pressure (PIP), expiratory time and fresh gas flow (FGF).
The multi-parameter monitor was used to measure the following cardiovascular and respiratory physiological parameters: pulse rate (from pulse oximetry), peripheral arterial oxygen haemoglobin saturation (SpO2), respiratory rate (fr) and respiratory gases (PetCO2, FetHal, inspired halothane percentage (FIHal), inspired oxygen percentage (FIO2)). Cloacal temperature (°C) was monitored every 5 to 15 min with a rectal thermometer. Physiological values, along with ventilation and anaesthesia settings, were recorded every five minutes throughout anaesthesia.
The Lead II ECG trace was acquired using 30-gauge 7 mm stainless steel electrodes (disposable subdermal needle electrode XS, Technomed, Limburg, The Netherlands) placed subcutaneously on the proximal cranial wing margins and a medial thigh. The electrodes were connected to a signal amplifier (DAM 50 differential amplifier; World Precision Instruments, FL, USA) via a custom-made breakout box (C. Johnson, Massey University, New Zealand). The data were digitized (Powerlab 7/35; AD Instruments, Australia) and recorded continuously (LabChart Pro v8.1.13; AD Instruments) on a personal desktop computer (iMac, Apple, Cupertino, CA, USA). A LabChart channel was programmed to detect ventricular depolarisation (QRS complexes) and display the heart rate (bpm) in real-time.
The following times were recorded: mask placement and preoxygenation, halothane start, intubation and stimulation periods (time of electrical stimulus application).
2.5. EEG Recording
A single-channel EEG configuration, based on that reported by McIlhone et al. [
9], was used. Stainless steel 30-gauge 7mm electrodes (disposable subdermal needle electrode XS, Technomed, Limburg, The Netherlands) were placed subcutaneously on the midline of the dorsal scalp (noninverting) and caudal to the right external auditory meatus (inverting) with the earth electrode on the dorsal neck. The electrodes were connected to a signal amplifier set with a gain of 1000× via a custom-made breakout box. The data were digitized at a rate of 1.0 kHz using DC offset, a low filter of 1 Hz and a high filter of 500 Hz with continuous recording and storage on a personal desktop computer. Recordings were taken for five minutes at each MAC multiple.
The published MAC of halothane for Rock Doves was used to calculate the tested F
etHal concentrations [
22]. The MAC multiples of 1×, 1.5× and 2× MAC therefore equated to F
etHal values of 1.6, 2.4 and 3.2%, respectively. The EEG was recorded at each of the three concentrations tested in ascending (a) then descending (d) order, giving five test MAC multiples of 1a, 1.5a, 2, 1.5d and 1d. To allow equilibration time between F
etHal and the target tissue concentration, the F
etHal was held stable at the test level for 10 min prior to the EEG recording. All anaesthesia and EEG recordings were completed in a Faraday cage.
2.6. Data Analysis
Data extraction and analysis were completed off-line after data collection was complete. Following manual inspection, noise artefacts, such as electromyography signals, were removed from the raw EEG signal. Fast Fourier transformation was completed for each five-minute recording using custom-written software (C. Johnson, Massey University, New Zealand) and the median frequency (F50), spectral edge frequency (F95) and the total power (Ptot) were established. For one-second bins, 300 points of data per MAC level per bird were smoothed using a moving 10-s average and analysed using a repeated-measures one-way ANOVA between the five MAC levels tested (consecutive order: 1×, 1.5×, 2×, 1.5× and 1× MAC). The conditions for the assumption of normality by the D’Agostino–Pearson method were met, and sphericity was not assumed. No transformation of variable data was required. When a significant effect of concentration was detected, multiple comparisons among MAC multiples were undertaken, corrected using Tukey post hoc analysis. All data analysis was completed using Prism 8 for macOS (version 8.4.2, GraphPad Software, San Diego, CA, USA). A p-value of <0.05 was considered significant.
Burst suppression was calculated from a two-minute section of the raw EEG recorded at each MAC multiple. The two-minute section was chosen as the segment of waveform with the least artefact. This provided the longest consecutive period for review, and therefore a more accurate reflection of the occurrence of any burst suppression. The amount of burst suppression was expressed as a percentage of the inactive EEG (the suppression) compared to active EEG (the burst) in the two-minute section. To be counted as inactive, the suppression periods had to occur continuously for at least 0.5 s. Similarly, to be included as active, the burst period had to occur continuously for at least 0.5 s. The active EEG was assigned at 30 µV or above. As previously defined by Gibson et al. [
23], the inactive EEG was defined as having 1/8th of the amplitude of the active EEG, giving an inactive EEG threshold of ≥3.75 µV. A repeated-measures one-way ANOVA was used to assess any differences in burst suppression percentage among halothane concentrations.
4. Discussion
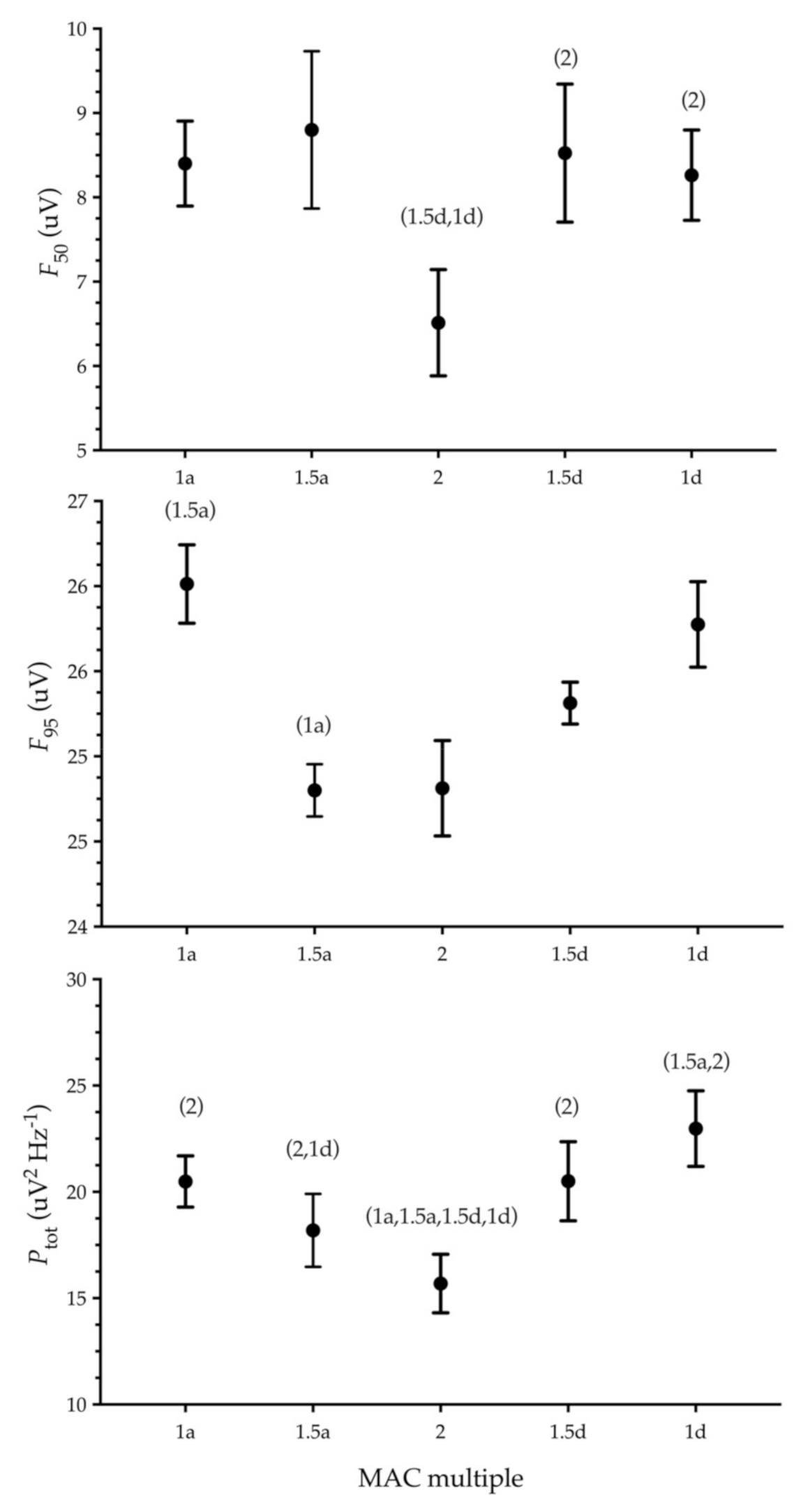
This study describes the effect of different halothane concentrations on the occurrence of burst suppression and changes in the spectral frequencies in the EEG of the Rock Dove. Negligible amounts of burst suppression were observed, and these were seen in only 3 of the 40 recorded EEG periods. Generally, the median and spectral edge frequencies and total power of the EEG reduced as the halothane concentration became greater, and they increased again as the anaesthetic concentration was lessened. Our hypotheses were not all supported given these results.
The burst suppression exhibited by Rock Doves over the three MAC multiples is similar to what has been observed with this agent in chickens, rats, dogs and horses [
9,
12,
24,
25,
26]. Burst suppression is a consequence of decreased cerebral metabolism and occurs when normal cortical neuronal activity intermittently ceases [
27]. In anaesthetised and comatose patients, this arrest of neocortical neuronal depolarisation creates the inactive EEG seen during the suppression periods [
28]. As anaesthesia deepens, the burst periods decrease until only suppressed and completely inactive, or “flatline”, EEG is seen [
29]. Burst suppression is therefore seen during periods of reduced cortical functionality and can be pathologic (e.g., hypoxic brain injury) or drug-induced, including general anaesthesia [
16]. The current findings demonstrate that electrical cortical activity remains in halothane-anaesthetised Rock Doves even at concentrations up to 2 MAC. This means that halothane may be a useful anaesthetic agent for Rock Doves when cortical responses are desired, for example in nociceptive studies [
3].
Spectral variables of transformed EEG data (
F50,
F95 and
Ptot) have been used in mammalian species to evaluate responses to noxious stimuli as a proxy for pain perception [
30,
31]. For MAM studies, it is important that the anaesthetic agent has minimal impact on these variables. Data recorded from humans have demonstrated that the changes in the frequency distribution of the EEG reflect the perceived intensity of a painful stimulus, a function of the cerebral cortex [
2]. Each frequency variable has shown a propensity for change in response to either the anaesthetic concentration or noxious stimuli. This reflects previous results, where overall, the EEG frequency distribution variables reduced with global cortical depression as anaesthesia concentration increased [
12].
The present results show a significantly decreased
F50 at 2× MAC compared to 1.5d and 1d MAC. A similar dose-dependent decrease was seen in chickens anaesthetised with halothane [
9]. The changes in
F95 of the Rock Doves in the current study are like those in chickens and mammals, with reduced levels of
F95 as the halothane concentration increases, though this was significant only between the 1a and 1.5a MAC multiples. This inverse relationship between the spectral edge (
F95) and anaesthetic concentration is also reported in other studies [
11,
14,
32]. The changes in the
F50 and
Ptot are more often associated with a reaction to noxious stimuli [
30,
32]. Generally,
F50 is seen to increase with pain, while the
Ptot decreases, as is often seen in mammalian studies [
3,
31].
The cause for the discrepancy between the ascending and descending sequences in the current study is not clear. For
F95, it may be postulated that over time there is a greater accumulation of halothane due to its physicochemical properties, specifically the relatively high degree of blood solubility [
33]. This increased solubility likely equates to a longer period required to equilibrate between plasma and the target location, but also means that as the anaesthetic progresses, there could be a degree of accumulation of halothane. Together, these properties might create the results seen, where only the early time points occurring at 1a and 1.5a MAC displayed a significant difference.
As the a priori calculation was performed for burst suppression, the small number of birds used resulted in a relatively wider spread of data for the frequency variables. The
F95 may have shown a significant change with a larger sample size, as seen with chickens [
9]. This would also likewise apply to the
F50, which inherently has a higher degree of variability due to its location within the frequency distribution.
The total power (
Ptot) changed significantly between 2× MAC and all other MAC multiples tested in the current study. A significant reduction in the
Ptot occurred with the highest halothane concentration, with the inverse seen as the halothane decreased to the lowest concentration. The general reduction in the total power of the EEG with increasing halothane concentration is in opposition to that seen in chickens during a similar evaluation [
9]. However, a similar response of decreasing total power was seen in the left somatosensory hemisphere in rats when assessed for EEG changes when the halothane concentration was increased [
11]. In that study, the changes in
Ptot were numerically small (<25% decrease from 1.25× to 1.75× MAC) and described as unlikely to be of biological significance. The magnitude of change was similar in the current study, and a similar conclusion might be drawn with an equally small change seen in
Ptot with a 20% decrease from 1× MAC to 2× MAC. In another study using anaesthesia infusion on a background of halothane anaesthesia in dogs, a decrease in
Ptot at the highest plasma concentration was seen [
12]. Here, the decreased
Ptot at the highest plasma concentration was ascribed to the general depression of cortical activity and reduced neuronal activation. This may be a partial explanation for the results seen in the current study. A further cause of the disparity from the results in chickens may be the MAC levels used. In the current study, the MAC reference was determined using an electrical stimulus, whereas the MAC value used in the chicken study was based on a mechanical stimulus [
34]. The resultant comparison of results could therefore not accurately reflect the true situation.
There are specific limitations in the present study that may influence the interpretation of data. The small number of birds, though calculated to show a sufficient response in the level of burst suppression with a priori testing, may have given non-significant differences due to more inter-individual variation than expected [
9]. An increased sample size may have decreased the impact of this variation. Finally, it must be re-iterated that the use of halothane in the MAM using Rock Doves is intended to further the capacity of pain research in the laboratory setting. Once established, however, this model will lay the foundation for the scientific-based clinical application of analgesics to birds from both private and wild ornithological sources.


 ,
, 
{kind=link}
