
Nestling Diet of Two Sympatric Insectivorous Passerines in Different Habitats—A Metabarcoding Study
 , ,
, ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
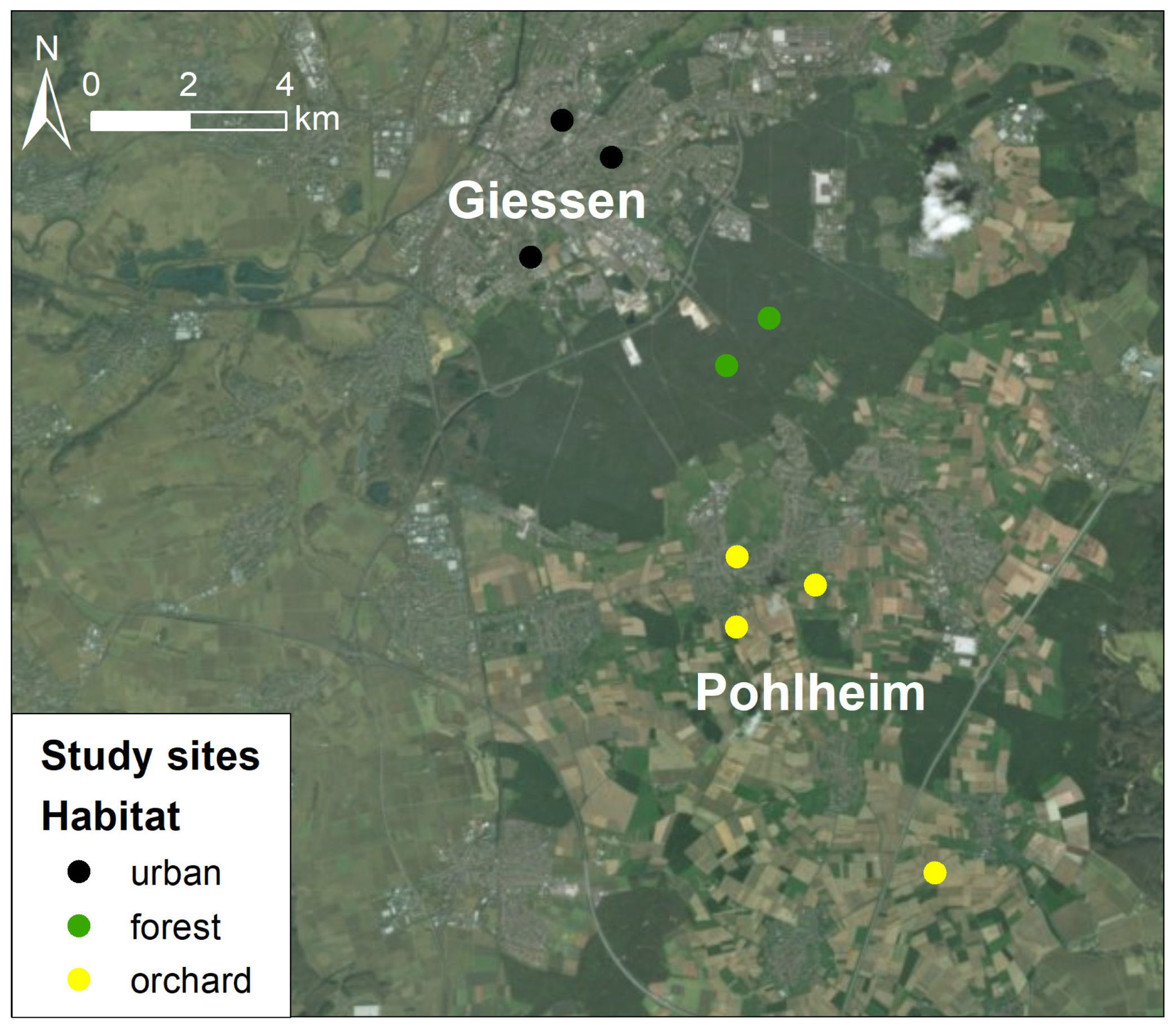
2.1. Study Sites and Nest Boxes
2.2. Faeces Collection and DNA Extraction from Faeces
2.3. Primer Performance and Selection
2.4. PCR Amplification of Prey DNA from Faeces
2.5. DNA Purification, Index PCR and Sequencing
2.6. Arthropod Samples from Trees
2.7. Data Processing—Bioinformatic Analyses of Sequences from Faecal Samples
2.8. Data Processing—Statistical Analyses and Biodiversity Indices
3. Results
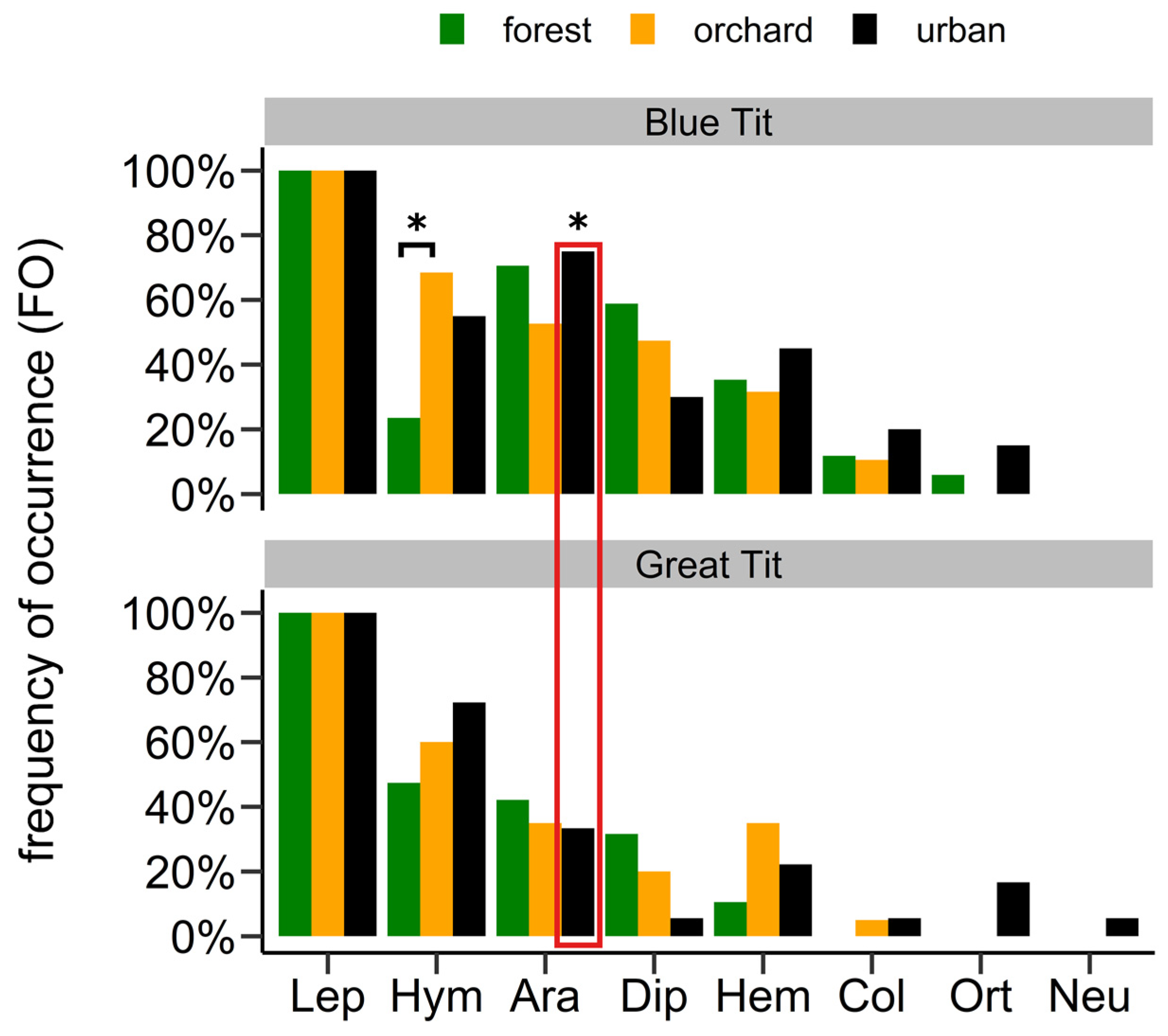
3.1. Diet Compositions of Nestlings
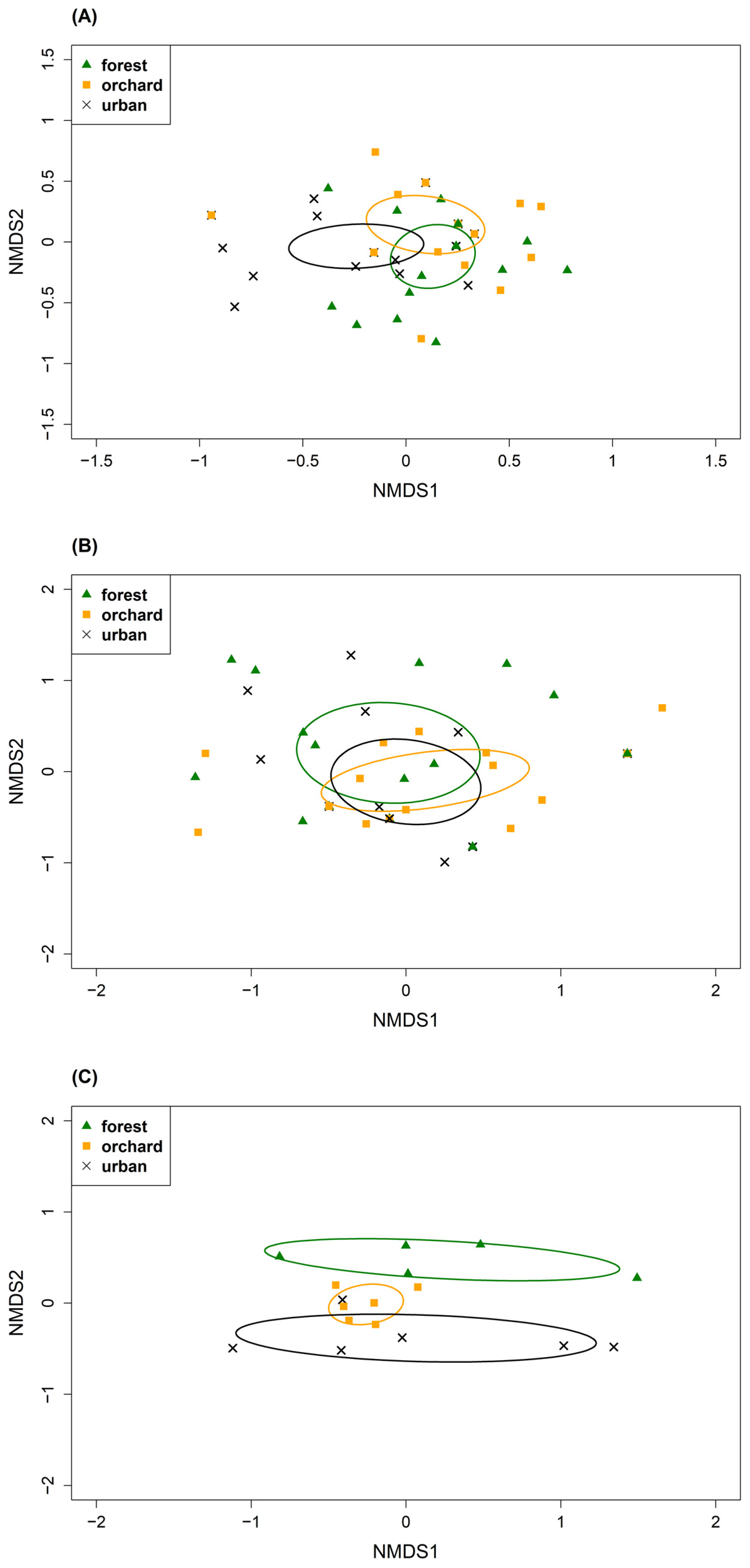
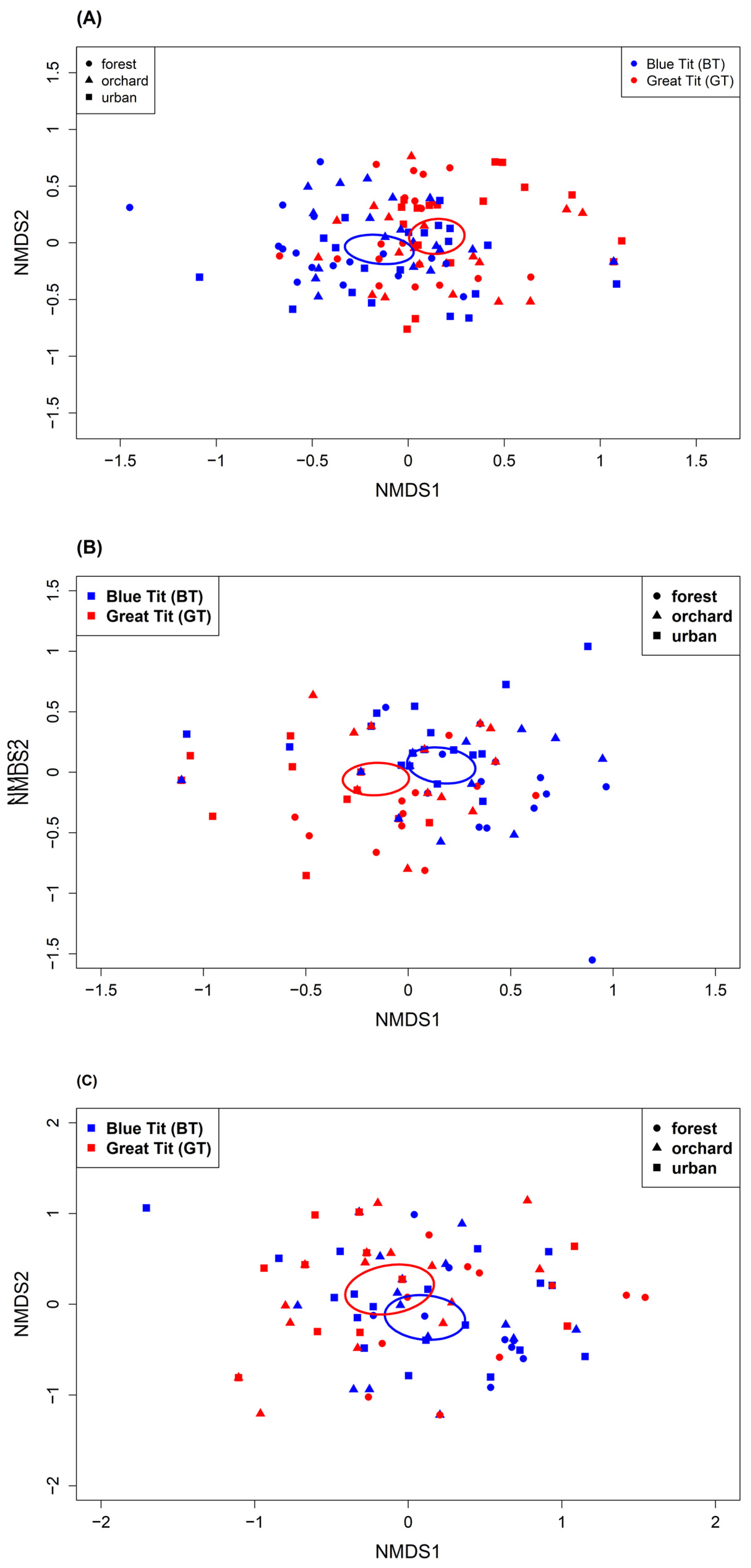
3.2. Differences in Diet Composition
3.3. Tree Samples—Availability of Arboreal Prey
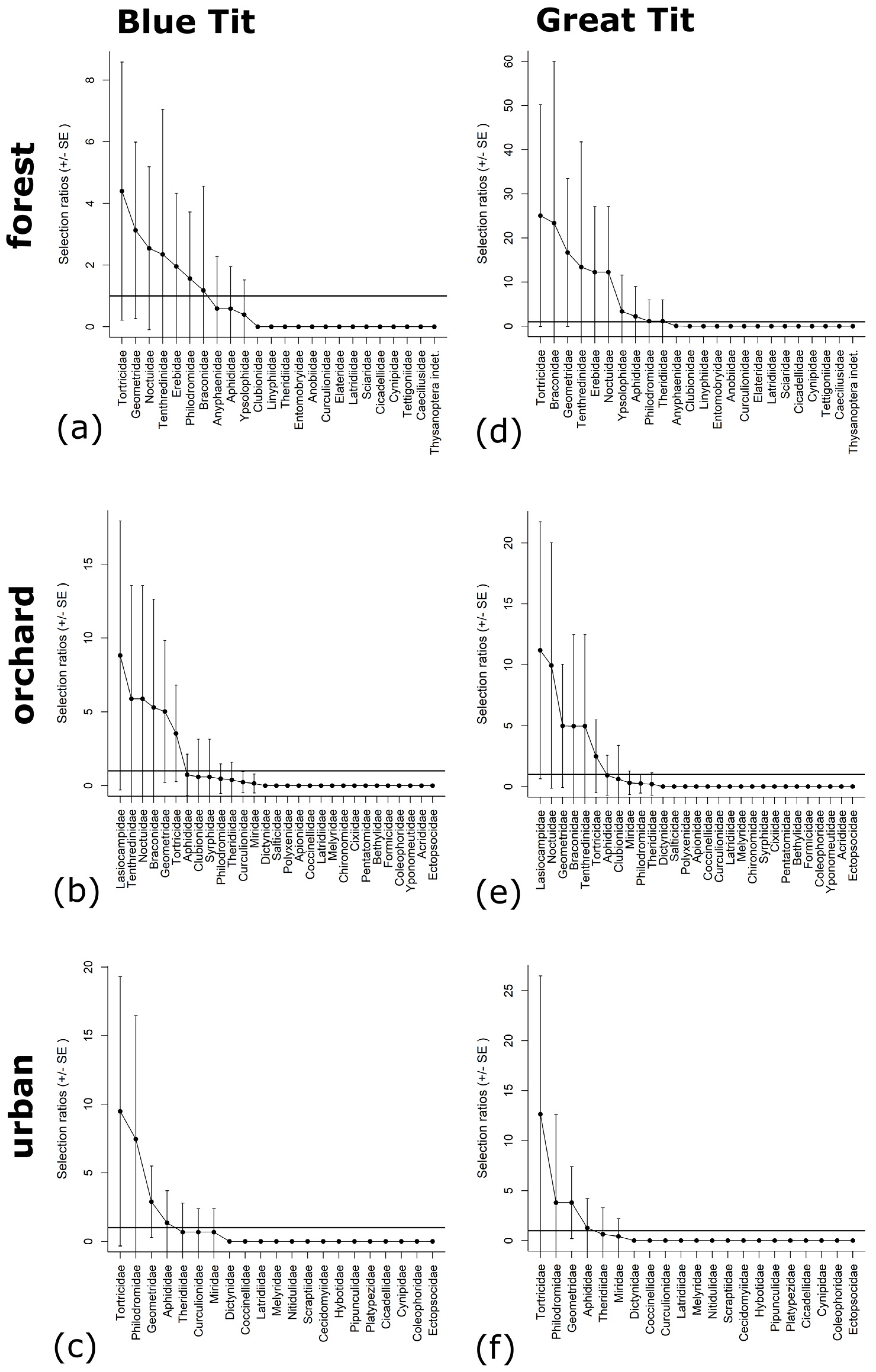
3.4. Parental Prey Selectivity
4. Discussion
4.1. Diet Composition
4.2. Differences in Diet Composition
4.3. Tree Samples—Availability of Arboreal Prey
4.4. Parental Prey Selectivity and Suitability as ‘Biodiversity Capsules’
4.5. Methodological Considerations
4.6. Study Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sala, O.E.; Chapin, F.S.; Armesto, J.J.; Berlow, E.; Bloomfield, J.; Dirzo, R.; Huber-Sanwald, E.; Huenneke, L.F.; Jackson, R.B.; Kinzig, A.; et al. Global Biodiversity Scenarios for the Year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef]
- Tscharntke, T.; Klein, A.M.; Kruess, A.; Steffan-Dewenter, I.; Thies, C. Landscape Perspectives on Agricultural Intensification and Biodiversity—Ecosystem Service Management. Ecol. Lett. 2005, 8, 857–874. [Google Scholar] [CrossRef]
- Foley, J.A.; DeFries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Chapin, F.S.; Coe, M.T.; Daily, G.C.; Gibbs, H.K.; et al. Global Consequences of Land Use. Science 2005, 309, 570–574. [Google Scholar] [CrossRef]
- Matson, P.A. Agricultural Intensification and Ecosystem Properties. Science 1997, 277, 504–509. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F.; Wyckhuys, K.A.G. Worldwide Decline of the Entomofauna: A Review of Its Drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Seibold, S.; Gossner, M.M.; Simons, N.K.; Blüthgen, N.; Müller, J.; Ambarlı, D.; Ammer, C.; Bauhus, J.; Fischer, M.; Habel, J.C.; et al. Arthropod Decline in Grasslands and Forests Is Associated with Landscape-Level Drivers. Nature 2019, 574, 671–674. [Google Scholar] [CrossRef]
- Hallmann, C.A.; Foppen, R.P.B.; van Turnhout, C.A.M.; de Kroon, H.; Jongejans, E. Declines in Insectivorous Birds Are Associated with High Neonicotinoid Concentrations. Nature 2014, 511, 341–343. [Google Scholar] [CrossRef]
- Habel, J.C.; Samways, M.J.; Schmitt, T. Mitigating the Precipitous Decline of Terrestrial European Insects: Requirements for a New Strategy. Biodivers. Conserv. 2019, 28, 1343–1360. [Google Scholar] [CrossRef]
- Bauer, H.-G.; Bezzel, E.; Fiedler, W. Das Kompendium Der Vögel Mitteleuropas, Volume 2: Passeriformes—Sperlingsvögel, 2nd ed.; AULA-Verlag: Wiebelsheim, Germany, 2005; Volume 2, ISBN 9783891046487. [Google Scholar]
- Wahl, J.; Dröschmeister, R.; Gerlach, B.; Grüneberg, C.; Langgemach, T.; Trautmann, S.; Sudfeldt, C. Vögel in Deutschland—2014; DDA, BfN, LAG VSW: Münster, Germany, 2015; ISBN 9783981554359. [Google Scholar]
- Bowler, D.E.; Heldbjerg, H.; Fox, A.D.; de Jong, M.; Böhning-Gaese, K. Long-Term Declines of European Insectivorous Bird Populations and Potential Causes. Conserv. Biol. 2019, 33, 1120–1130. [Google Scholar] [CrossRef]
- Bańbura, J.; Bańbura, M. Blue Tits Cyanistes Caeruleus and Great Tits Parus Major As Urban Habitat Breeders. Int. Stud. Sparrows 2012, 36, 66–72. [Google Scholar] [CrossRef]
- Jokimäki, J.; Suhonen, J.; Jokimäki-Kaisanlahti, M.L.; Carbó-Ramírez, P. Effects of Urbanization on Breeding Birds in European Towns: Impacts of Species Traits. Urban Ecosyst. 2016, 19, 1565–1577. [Google Scholar] [CrossRef]
- Chace, J.F.; Walsh, J.J. Urban Effects on Native Avifauna: A Review. Landsc. Urban Plan. 2006, 74, 46–69. [Google Scholar] [CrossRef]
- Seress, G.; Liker, A. Habitat Urbanization and Its Effects on Birds. Acta Zool. Acad. Sci. Hungaricae 2015, 61, 373–408. [Google Scholar] [CrossRef]
- Cowie, R.J.; Hinsley, S.A. Feeding Ecology of Great Tits (Parus Major) and Blue Tits (Parus caeruleus), Breeding in Suburban Gardens. J. Anim. Ecol. 1988, 57, 611–626. [Google Scholar] [CrossRef]
- Glutz von Blotzheim, U.N.; Bauer, K.M. Handbuch Der Vögel Mitteleuropas. Band 13/1; Aula-Verlag: Wiesbaden, Germany, 1993; ISBN 3923527004. [Google Scholar]
- Newton, I. The Recent Declines of Farmland Bird Populations in Britain: An Appraisal of Causal Factors and Conservation Actions. Ibis 2004, 146, 579–600. [Google Scholar] [CrossRef]
- Minot, E.O. Effects of Interspecific Competition for Food in Breeding Blue and Great Tits. J. Anim. Ecol. 1981, 50, 375. [Google Scholar] [CrossRef]
- Dhondt, A.A.; Adriaensen, F. Experiments on Competition between Great and Blue Tit: Effects on Blue Tit Reproductive Success and Population Processes. Ostrich 1999, 70, 39–48. [Google Scholar] [CrossRef]
- Perrins, C.M. Tits and Their Caterpillar Food Supply. Ibis 1991, 133, 49–54. [Google Scholar] [CrossRef]
- Naef-Daenzer, L.; Naef-Daenzer, B.; Nager, R.G. Prey Selection and Foraging Performance of Breeding Great Tits Parus Major in Relation to Food Availability. J. Avian Biol. 2000, 31, 206–214. [Google Scholar] [CrossRef]
- Pagani-Núñez, E.; Ruiz, I.; Quesada, J.; Negro, J.J.; Senar, J.C. The Diet of Great Tit Parus Major Nestlings in a Mediterranean Iberian Forest: The Important Role of Spiders. Anim. Biodivers. Conserv. 2011, 34, 355–361. [Google Scholar] [CrossRef]
- Mols, C.M.M.; Visser, M.E. Great Tits Can Reduce Caterpillar Damage in Apple Orchards. J. Appl. Ecol. 2002, 39, 888–899. [Google Scholar] [CrossRef]
- Powell, C.; Lill, A.; Johnstone, C.P. Body Condition and Chronic Stress in Urban and Rural Noisy Miners. Open Ornithol. J. 2013, 6, 25–31. [Google Scholar] [CrossRef]
- Krištín, A. Trophische Beziehungen Zwischen Singvögeln Und Wirbellosen Im Eichen-Buchenwald Zur Brutzeit. Der. Ornithol. Beobachter. 1992, 89, 157–169. [Google Scholar]
- Royama, T. Factors Governing the Hunting Behaviour and Selection of Food by the Great Tit (Parus major L.). J. Anim. Ecol. 1970, 39, 619–668. [Google Scholar] [CrossRef]
- Stenning, M. The Blue Tit; T & AD Poyser: London, UK, 2018. [Google Scholar]
- Betts, M.M. The Food of Titmice in Oak Woodland. J. Anim. Ecol. 1955, 24, 282. [Google Scholar] [CrossRef]
- Ceia, R.S.; Machado, R.A.; Ramos, J.A. Nestling Food of Three Hole-Nesting Passerine Species and Experimental Increase in Their Densities in Mediterranean Oak Woodlands. Eur. J. For. Res. 2016, 135, 839–847. [Google Scholar] [CrossRef]
- Török, J. Food Segregation in Three Hole-Nesting Bird Species during the Breeding Season. Ardea 1986, 74, 129–136. [Google Scholar]
- Van Balen, J.H. A Comparative Sudy of the Breeding Ecology of the Great Tit Parus Major in Different Habitats. Ardea 1973, 61, 1–93. [Google Scholar] [CrossRef]
- García-Navas, V.; Ferrer, E.S.; Sanz, J.J. Prey Selectivity and Parental Feeding Rates of Blue Tits Cyanistes Caeruleus in Relation to Nestling Age. Bird Study 2012, 59, 236–242. [Google Scholar] [CrossRef]
- Solonen, T. Breeding of the Great Tit and Blue Tit in Urban and Rural Habitats in Southern Finland. Ornis Fenn. 2001, 78, 49–60. [Google Scholar]
- Seress, G.; Hammer, T.; Bókony, V.; Vincze, E.; Preiszner, B.; Pipoly, I.; Sinkovics, C.; Evans, K.L.; Liker, A. Impact of Urbanization on Abundance and Phenology of Caterpillars and Consequences for Breeding in an Insectivorous Bird. Ecol. Appl. 2018, 28, 1143–1156. [Google Scholar] [CrossRef] [PubMed]
- Wawrzyniak, J.; Glądalski, M.; Kaliński, A.; Bańbura, M.; Markowski, M.; Skwarska, J.; Zieliński, P.; Bańbura, J. Differences in the Breeding Performance of Great Tits Parus Major between a Forest and an Urban Area: A Long Term Study on First Clutches. Eur. Zool. J. 2020, 87, 294–309. [Google Scholar] [CrossRef]
- Vaugoyeau, M.; Adriaensen, F.; Artemyev, A.; Bańbura, J.; Barba, E.; Biard, C.; Blondel, J.; Bouslama, Z.; Bouvier, J.C.; Camprodon, J.; et al. Interspecific Variation in the Relationship between Clutch Size, Laying Date and Intensity of Urbanization in Four Species of Hole-Nesting Birds. Ecol. Evol. 2016, 6, 5907–5920. [Google Scholar] [CrossRef] [PubMed]
- Fischbacher, M.; Naef-Daenzer, B.; Naef-Daenzer, L. Estimating Caterpillar Density on Trees by Collection of Frass Droppings. Ardea 1998, 86, 121–129. [Google Scholar]
- Mägi, M.; Mänd, R.; Tamm, H.; Sisask, E.; Kilgas, P.; Tilgar, V. Low Reproductive Success of Great Tits in the Preferred Habitat: A Role of Food Availability. Ecoscience 2009, 16, 145–157. [Google Scholar] [CrossRef]
- Arnold, K.E.; Ramsay, S.L.; Henderson, L.; Larcombe, S.D. Seasonal Variation in Diet Quality: Antioxidants, Invertebrates and Blue Tits Cyanistes Caeruleus. Biol. J. Linn. Soc. 2010, 99, 708–717. [Google Scholar] [CrossRef]
- Cholewa, M.; Wesołowski, T. Nestling Food of European Hole-Nesting Passerines: Do We Know Enough to Test the Adaptive Hypotheses on Breeding Seasons? Acta Ornithol. 2011, 46, 105–116. [Google Scholar] [CrossRef]
- Grzędzicka, E. Habitat and Diet Variability of Two Coexisting Tit Species in Central European Forests. Bird Study 2018, 65, 52–61. [Google Scholar] [CrossRef]
- Tanneberger, F.; Flinks, H.; Arbeiter, S.; Minets, M.; Hegemann, A. Diet Analysis of Wetland Passerine Nestlings Using Neck Collars or Faecal Sampling Produces Similar Results. Ardea 2017, 105, 145–152. [Google Scholar] [CrossRef]
- García-Navas, V.; Ferrer, E.S.; Sanz, J.J. Prey Choice, Provisioning Behaviour, and Effects of Early Nutrition on Nestling Phenotype of Titmice. Ecoscience 2013, 20, 9–18. [Google Scholar] [CrossRef]
- Caprio, E.; Rolando, A. Management Systems May Affect the Feeding Ecology of Great Tits Parus Major Nesting in Vineyards. Agric. Ecosyst. Environ. 2017, 243, 67–73. [Google Scholar] [CrossRef]
- Serrano-Davies, E.; Sanz, J.J. Habitat Structure Modulates Nestling Diet Composition and Fitness of Blue Tits Cyanistes Caeruleus in the Mediterranean Region. Bird Study 2017, 64, 295–305. [Google Scholar] [CrossRef]
- Symondson, W.O.C. Molecular Identification of Prey in Predator Diets. Mol. Ecol. 2002, 11, 627–641. [Google Scholar] [CrossRef] [PubMed]
- Zeale, M.R.K.; Butlin, R.K.; Barker, G.L.A.; Lees, D.C.; Jones, G. Taxon-Specific PCR for DNA Barcoding Arthropod Prey in Bat Faeces. Mol. Ecol. Resour. 2011, 11, 236–244. [Google Scholar] [CrossRef]
- Pompanon, F.; Deagle, B.E.; Symondson, W.O.C.; Brown, D.S.; Jarman, S.N.; Taberlet, P. Who Is Eating What: Diet Assessment Using next Generation Sequencing. Mol. Ecol. 2012, 21, 1931–1950. [Google Scholar] [CrossRef]
- Deagle, B.E.; Kirkwood, R.; Jarman, S.N. Analysis of Australian Fur Seal Diet by Pyrosequencing Prey DNA in Faeces. Mol. Ecol. 2009, 18, 2022–2038. [Google Scholar] [CrossRef]
- O’Rourke, D.R.; Mangan, M.T.; Mangan, K.E.; Bokulich, N.A.; MacManes, M.D.; Foster, J.T. Lord of the Diptera (and Moths and a Spider): Molecular Diet Analyses and Foraging Ecology of Indiana Bats in Illinois. Front. Ecol. Evol. 2021, 9, 623655. [Google Scholar] [CrossRef]
- Morley, S.A.; Foley, M.M.; Duda, J.J.; Beirne, M.M.; Paradis, R.L.; Johnson, R.C.; McHenry, M.L.; Elofson, M.; Sampson, E.M.; McCoy, R.E.; et al. Shifting Food Web Structure during Dam Removal—Disturbance and Recovery during a Major Restoration Action. PLoS ONE 2020, 15, e0239198. [Google Scholar] [CrossRef]
- Trevelline, B.K.; Latta, S.C.; Marshall, L.C.; Nuttle, T.; Porter, B.A. Molecular Analysis of Nestling Diet in a Long-Distance Neotropical Migrant, the Louisiana Waterthrush (Parkesia motacilla). Auk 2016, 133, 415–428. [Google Scholar] [CrossRef]
- Dunn, J.C.; Stockdale, J.E.; Moorhouse-Gann, R.J.; McCubbin, A.; Hipperson, H.; Morris, A.J.; Grice, P.V.; Symondson, W.O.C. The Decline of the Turtle Dove: Dietary Associations with Body Condition and Competition with Other Columbids Analysed Using High-Throughput Sequencing. Mol. Ecol. 2018, 27, 3386–3407. [Google Scholar] [CrossRef]
- Kleinschmidt, B.; Burger, C.; Dorsch, M.; Nehls, G.; Heinänen, S.; Morkūnas, J.; Žydelis, R.; Moorhouse-Gann, R.J.; Hipperson, H.; Symondson, W.O.C.; et al. The Diet of Red-Throated Divers (Gavia stellata) Overwintering in the German Bight (North Sea) Analysed Using Molecular Diagnostics. Mar. Biol. 2019, 166, 77. [Google Scholar] [CrossRef]
- Rytkönen, S.; Vesterinen, E.J.; Westerduin, C.; Leviäkangas, T.; Vatka, E.; Mutanen, M.; Välimäki, P.; Hukkanen, M.; Suokas, M.; Orell, M. From Feces to Data: A Metabarcoding Method for Analyzing Consumed and Available Prey in a Bird-Insect Food Web. Ecol. Evol. 2018, 9, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Jarrett, C.; Powell, L.L.; McDevitt, H.; Helm, B.; Welch, A.J. Bitter Fruits of Hard Labour: Diet Metabarcoding and Telemetry Reveal That Urban Songbirds Travel Further for Lower-Quality Food. Oecologia 2020, 193, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Shutt, J.D.; Nicholls, J.A.; Trivedi, U.H.; Burgess, M.D.; Stone, G.N.; Hadfield, J.D.; Phillimore, A.B. Gradients in Richness and Turnover of a Forest Passerine’s Diet Prior to Breeding: A Mixed Model Approach Applied to Faecal Metabarcoding Data. Mol. Ecol. 2020, 29, 1199–1213. [Google Scholar] [CrossRef]
- Boyer, S.; Cruickshank, R.H.; Wratten, S.D. Faeces of Generalist Predators as “Biodiversity Capsules”: A New Tool for Biodiversity Assessment in Remote and Inaccessible Habitats. Food Webs 2015, 3, 1–6. [Google Scholar] [CrossRef]
- Hessisches Landesamt für Naturschutz, Umwelt und Geologie (HLNUG), Klimaportal Hessen. Available online: https://klimaportal.hlnug.de/witterungsbericht (accessed on 1 October 2021).
- Jusino, M.A.; Banik, M.T.; Palmer, J.M.; Wray, A.K.; Xiao, L.; Pelton, E.; Barber, J.R.; Kawahara, A.Y.; Gratton, C.; Peery, M.Z.; et al. An Improved Method for Utilizing High-Throughput Amplicon Sequencing to Determine the Diets of Insectivorous Animals. Mol. Ecol. Resour. 2019, 19, 176–190. [Google Scholar] [CrossRef]
- Emerson, B.C.; Casquet, J.; López, H.; Cardoso, P.; Borges, P.A.V.; Mollaret, N.; Oromí, P.; Strasberg, D.; Thébaud, C. A Combined Field Survey and Molecular Identification Protocol for Comparing Forest Arthropod Biodiversity across Spatial Scales. Mol. Ecol. Resour. 2017, 17, 694–707. [Google Scholar] [CrossRef]
- Martínez, J.; Martínez-de la Puente, J.; Herrero, J.; Del Cerro, S.; Lobato, E.; Rivero-de Aguilar, J.; Vásquez, R.A.; Merino, S. A Restriction Site to Differentiate Plasmodium and Haemoproteus Infections in Birds: On the Inefficiency of General Primers for Detection of Mixed Infections. Parasitology 2009, 136, 713–722. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA Primers for Amplification of Mitochondrial Cytochrome c Oxidase Subunit I from Diverse Metazoan Invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Masello, J.F.; Barbosa, A.; Kato, A.; Mattern, T.; Medeiros, R.; Stockdale, J.E.; Kümmel, M.N.; Bustamante, P.; Belliure, J.; Benzal, J.; et al. How Animals Distribute Themselves in Space: Energy Landscapes of Antarctic Avian Predators. Mov. Ecol. 2021, 9, 24. [Google Scholar] [CrossRef] [PubMed]
- Department of Bioinformatics & Systems Biology (Justus Liebig University, Giessen), Galaxy Giessen. Available online: https://www.computational.bio.uni-giessen.de/galaxy (accessed on 1 October 2020).
- Afgan, E.; Baker, D.; Batut, B.; van den Beek, M.; Bouvier, D.; Cech, M.; Chilton, J.; Clements, D.; Coraor, N.; Grüning, B.A.; et al. The Galaxy Platform for Accessible, Reproducible and Collaborative Biomedical Analyses: 2018 Update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef] [PubMed]
- Babraham Institute Cambridge, Braham Bioinformatics. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 1 October 2020).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Magoč, T.; Salzberg, S.L. FLASH: Fast Length Adjustment of Short Reads to Improve Genome Assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing Mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and Clustering Orders of Magnitude Faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Wright, E.S.; Vetsigian, K.H. Quality Filtering of Illumina Index Reads Mitigates Sample Cross-Talk. BMC Genom. 2016, 17, 876. [Google Scholar] [CrossRef]
- Quéméré, E.; Hibert, F.; Miquel, C.; Lhuillier, E.; Rasolondraibe, E.; Champeau, J.; Rabarivola, C.; Nusbaumer, L.; Chatelain, C.; Gautier, L.; et al. A DNA Metabarcoding Study of a Primate Dietary Diversity and Plasticity across Its Entire Fragmented Range. PLoS ONE 2013, 8, e58971. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2020. [Google Scholar]
- Shannon, C.E. A Mathematical Theory of Communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Jaccard, P. The Distribution of the Flora in the Alpine Zone. New Phytol. 1912, 11, 37–50. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package, R Package Version 2.5-6 2019. Available online: https://cran.r-project.org/web/packages/vegan/vegan.pdf (accessed on 19 December 2023).
- Faith, D.P.; Minchin, P.R.; Belbin, L. Compositional Dissimilarity as a Robust Measure of Ecological Distance. Vegetatio 1987, 69, 57–68. [Google Scholar] [CrossRef]
- Minchin, P.R. An Evaluation of the Relative Robustness of Techniques for Ecological Ordination. Vegetatio 1987, 69, 89–107. [Google Scholar] [CrossRef]
- Manly, B.; McDonald, L.; Thomas, D.; McDonald, T.; Erickson, W. Resource Selection by Animals: Statistical Design and Analysis for Field Studies, 2nd ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002; ISBN 1-4020-0677-2. [Google Scholar]
- Calenge, C. The Package “Adehabitat” for the R Software: A Tool for the Analysis of Space and Habitat Use by Animals. Ecol. Modell. 2006, 197, 516–519. [Google Scholar] [CrossRef]
- Banbura, J.; Lambrechts, M.M.; Blondel, J.; Perret, P.; Cartan-Son, M. Food Handling Time of Blue Tit Chicks: Constraints and Adaptation to Different Prey Types. J. Avian Biol. 1999, 30, 263–270. [Google Scholar] [CrossRef]
- Tremblay, I.; Thomas, D.; Blondel, J.; Perret, P.; Lambrechts, M.M. The Effect of Habitat Quality on Foraging Patterns, Provisioning Rate and Nestling Growth in Corsican Blue Tits Parus Caeruleus. Ibis 2005, 147, 17–24. [Google Scholar] [CrossRef]
- Wilkin, T.A.; King, L.E.; Sheldon, B.C. Habitat Quality, Nestling Diet, and Provisioning Behaviour in Great Tits Parus Major. J. Avian Biol. 2009, 40, 135–145. [Google Scholar] [CrossRef]
- Eeva, T.; Helle, S.; Salminen, J.-P.; Hakkarainen, H. Carotenoid Composition of Invertebrates Consumed by Two Insectivorous Bird Species. J. Chem. Ecol. 2010, 36, 608–613. [Google Scholar] [CrossRef]
- García-Navas, V.; Sanz, J.J. The Importance of a Main Dish: Nestling Diet and Foraging Behaviour in Mediterranean Blue Tits in Relation to Prey Phenology. Oecologia 2011, 165, 639–649. [Google Scholar] [CrossRef]
- Wesołowski, T.; Rowiński, P. Do Blue Tits Cyanistes Caeruleus Synchronize Reproduction with Caterpillar Peaks in a Primeval Forest? Bird Study 2014, 61, 231–245. [Google Scholar] [CrossRef]
- Cramp, S.; Perrins, C.M. Handbook of the Birds of Europe, the Middle East and North Africa. The Birds of the Western Palearctic. Vol. VII, Flycatchers to Shrikes; Oxford University Press: Oxford, UK, 1993. [Google Scholar]
- Kovačević, Ž. Die Nahrungswahl Und Das Auftreten Der Pflanzenschädlinge. Anz. Für Schädlingskd. 1956, 29, 97–101. [Google Scholar] [CrossRef]
- Deschka, G.; Wimmer, J. Die Schmetterlingsfauna Der Kreuzmauer (Insecta: Lepidoptera). Beiträge Zur Naturkd. Oberösterreichs 2000, 9, 65–186. [Google Scholar]
- Reichholf, J.H. Der Ringelspinner Malacosoma Neustria: Früher Gefürchteter Schädling Und Nun Eine Seltenheit. Berichte Des Naturwissenschaftlichen Ver. Für Schwab. e.V. 2009, 113, 58–63. [Google Scholar]
- Kübler, S. Nahrungsökologie Stadtlebender Vogelarten Entlang Eines Urbangradienten; Humboldt University of Berlin: Berlin, Germany, 2006; Volume 44. [Google Scholar]
- Marciniak, B.; Nadolski, J.; Nowakowska, M.; Loga, B.; Bańbura, J. Habitat and Annual Variation in Arthropod Abundance Affects Blue Tit Cyanistes Caeruleus Reproduction. Acta Ornithol. 2007, 42, 53–62. [Google Scholar] [CrossRef]
- Ilyina, T.A.; Kerimov, A.B.; Zagubizhenko, M.V.; Maksimov, G.V. Seasonal Dynamics of Leaf-Eating Insects Biomass and Its Influence on Carotenoid Content in Feathers of Great Tit Nestlings. Russ. J. Ecol. 2013, 44, 507–514. [Google Scholar] [CrossRef]
- Sillanpää, S.; Salminen, J.-P.P.; Lehikoinen, E.; Toivonen, E.; Eeva, T. Carotenoids in a Food Chain along a Pollution Gradient. Sci. Total Environ. 2008, 406, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.O. Wasps. In Encyclopedia of Insects; Resh, V.H., Cardé, R.T., Eds.; Elsevier Science & Technology: Amsterdam, The Netherlands, 2009; pp. 1049–1051. [Google Scholar]
- Verkuil, Y.I.; Nicolaus, M.; Ubels, R.; Dietz, M.M.; Samplonius, J.M.; Galema, A.; Kiekebos, K.; De Knijff, P.; Both, C. DNA Metabarcoding Successfully Quantifies Relative Abundances of Arthropod Taxa in Songbird Diets: A Validation Study Using Camera-Recorded Diets. bioRxiv 2020. [Google Scholar] [CrossRef]
- Ramsay, S.L.; Houston, D.C. Amino Acid Composition of Some Woodland Arthropods and Its Implications for Breeding Tits and Other Passerines. Ibis 2003, 145, 227–232. [Google Scholar] [CrossRef]
- Mänd, R.; Tilgar, V.; Lõhmus, A.; Leivits, A. Providing Nest Boxes for Hole-Nesting Birds—Does Habitat Matter? Biodivers. Conserv. 2005, 14, 1823–1840. [Google Scholar] [CrossRef]
- Pollock, C.J.; Capilla-Lasheras, P.; McGill, R.A.R.; Helm, B.; Dominoni, D.M. Integrated Behavioural and Stable Isotope Data Reveal Altered Diet Linked to Low Breeding Success in Urban-Dwelling Blue Tits (Cyanistes caeruleus). Sci. Rep. 2017, 7, 5014. [Google Scholar] [CrossRef]
- Nour, N.; Currie, D.; Matthysen, E.; Van Damme, R.; Dhondt, A.A. Effects of Habitat Fragmentation on Provisioning Rates, Diet and Breeding Success in Two Species of Tit (Great Tit and Blue Tit). Oecologia 1998, 114, 522–530. [Google Scholar] [CrossRef]
- Török, J.; Tóth, L. Asymmetric Competition between Two Tit Species: A Reciprocal Removal Experiment. J. Anim. Ecol. 1999, 68, 338–345. [Google Scholar] [CrossRef]
- Navalpotro, H.; Pagani–Núñez, E.; Hernández–Gómez, S.; Senar, J.C. Comparing Prey Composition and Prey Size Delivered to Nestlings by Great Tits, Parus Major, and Blue Tits, Cyanistes Caeruleus, in a Mediterranean Sclerophyllous Mixed Forest. Anim. Biodivers. Conserv. 2016, 39, 129–139. [Google Scholar] [CrossRef]
- Gibb, J.A.; Betts, M.M. Food and Food Supply of Nestling Tits (Paridae) in Breckland Pine. J. Anim. Ecol. 1963, 32, 489–533. [Google Scholar] [CrossRef]
- Glądalski, M.; Bańbura, M.; Kaliński, A.; Markowski, M.; Skwarska, J.; Wawrzyniak, J.; Zieliński, P.; Cyżewska, I.; Bańbura, J. Differences in the Breeding Success of Blue Tits Cyanistes Caeruleus between a Forest and an Urban Area: A Long-Term Study. Acta Ornithol. 2017, 52, 59–68. [Google Scholar] [CrossRef]
- Clem, C.S.; Held, D.W. Species Richness of Eruciform Larvae Associated with Native and Alien Plants in the Southeastern United States. J. Insect Conserv. 2015, 19, 987–997. [Google Scholar] [CrossRef]
- Simon, S.; Bouvier, J.C.; Debras, J.F.; Sauphanor, B. Biodiversity and Pest Management in Orchard Systems. A Review. Agron. Sustain. Dev. 2010, 30, 139–152. [Google Scholar] [CrossRef]
- Müller, W.; Hess, R.; Nievergelt, B. Die Obstgärten Und Ihre Vogelwelt Im Kanton Zürich. Der Ornitol. Beob. 1988, 85, 123–157. [Google Scholar]
- Altieri, M.A.; Schmidt, L.L. The Dynamics of Colonizing Arthropod Communities at the Interface of Abandoned, Organic and Commercial Apple Orchards and Adjacent Woodland Habitats. Agric. Ecosyst. Environ. 1986, 16, 29–43. [Google Scholar] [CrossRef]
- Mackenzie, J.A.; Hinsley, S.A.; Harrison, N.M. Parid Foraging Choices in Urban Habitat and Their Consequences for Fitness. Ibis 2014, 156, 591–605. [Google Scholar] [CrossRef]
- Vesterinen, E.J.; Ruokolainen, L.; Wahlberg, N.; Peña, C.; Roslin, T.; Laine, V.N.; Vasko, V.; Sääksjärvi, I.E.; Norrdahl, K.; Lilley, T.M. What You Need Is What You Eat? Prey Selection by the Bat Myotis Daubentonii. Mol. Ecol. 2016, 25, 1581–1594. [Google Scholar] [CrossRef]
- Jarman, S.N.; McInnes, J.C.; Faux, C.; Polanowski, A.M.; Marthick, J.; Deagle, B.E.; Southwell, C.; Emmerson, L. Adélie Penguin Population Diet Monitoring by Analysis of Food DNA in Scats. PLoS ONE 2013, 8, e82227. [Google Scholar] [CrossRef] [PubMed]
- Hestetun, J.T.; Bye-Ingebrigtsen, E.; Nilsson, R.H.; Glover, A.G.; Johansen, P.O.; Dahlgren, T.G. Significant Taxon Sampling Gaps in DNA Databases Limit the Operational Use of Marine Macrofauna Metabarcoding. Mar. Biodivers. 2020, 50, 70. [Google Scholar] [CrossRef]
- Kvist, S. Barcoding in the Dark?: A Critical View of the Sufficiency of Zoological DNA Barcoding Databases and a Plea for Broader Integration of Taxonomic Knowledge. Mol. Phylogenet. Evol. 2013, 69, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Wiemers, M.; Fiedler, K. Does the DNA Barcoding Gap Exist?—A Case Study in Blue Butterflies (Lepidoptera: Lycaenidae). Front. Zool. 2007, 4, 8. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Blue Tit | Great Tit | |||||
|---|---|---|---|---|---|---|
| Sample Processing Steps | Forest | Orchard | Urban | Forest | Orchard | Urban |
| (1) PCR products in NGS Library for sequencing | 20 | 19 | 20 | 20 | 20 | 20 |
| (2) successfully sequenced samples | 17 | 19 | 20 | 19 | 20 | 18 |
| (3) after manual filtering steps to avoid false positives | 17 (11) | 19 (12) | 20 (15) | 19 (10) | 20 (17) | 18 (15) |
| % FO | Blue Tit | Great Tit | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Prev All | All | Forest | Orchard | Urban | Forest | Orchard | Urban | ||
| Order | Family | n = 113 | n = 113 | n = 17 | n = 19 | n = 20 | n = 19 | n = 20 | n = 18 |
| Araneae | Anyphaenidae | 14 | 12.4 | 11.8 | 5.3 | 20.0 | 15.8 | 20.0 | |
| Araneae | Araneidae | 8 | 7.1 | 17.6 | 21.1 | 5.3 | |||
| Araneae | Clubionidae | 6 | 5.3 | 5.3 | 10.0 | 5.0 | 11.1 | ||
| Araneae | Philodromidae | 29 | 25.7 | 47.1 | 21.1 | 55.0 | 5.3 | 10.0 | 16.7 |
| Araneae | Theridiidae | 7 | 6.2 | 10.5 | 10.0 | 5.3 | 5.0 | 5.6 | |
| Araneae | Thomisidae | 27 | 23.9 | 47.1 | 26.3 | 20.0 | 26.3 | 15.0 | 11.1 |
| Coleoptera | Buprestidae | 3 | 2.7 | 5.0 | 5.0 | 5.6 | |||
| Coleoptera | Curculionidae | 5 | 4.4 | 10.5 | 15.0 | ||||
| Coleoptera | Elateridae | 1 | 0.9 | 5.6 | |||||
| Coleoptera | Tenebrionidae | 2 | 1.8 | 11.8 | |||||
| Diptera | Asilidae | 1 | 0.9 | 5.3 | |||||
| Diptera | Muscidae | 3 | 2.7 | 11.8 | 5.0 | ||||
| Diptera | Phoridae | 1 | 0.9 | 5.0 | |||||
| Diptera | Psychodidae | 1 | 0.9 | 5.0 | |||||
| Diptera | Syrphidae | 6 | 5.3 | 11.8 | 5.3 | 5.0 | 5.3 | 5.6 | |
| Diptera | Tachinidae | 32 | 28.3 | 58.8 | 47.4 | 25.0 | 21.1 | 15.0 | 5.6 |
| Hemiptera | Aphididae | 25 | 22.1 | 17.6 | 26.3 | 30.0 | 10.5 | 30.0 | 16.7 |
| Hemiptera | Miridae | 10 | 8.8 | 17.6 | 5.3 | 15.0 | 10.0 | 5.6 | |
| Hemiptera | Pentatomidae | 1 | 0.9 | 5.0 | |||||
| Hymenoptera | Apidae | 4 | 3.5 | 5.9 | 10.0 | 5.6 | |||
| Hymenoptera | Braconidae | 46 | 40.7 | 11.8 | 47.4 | 45.0 | 36.8 | 40.0 | 61.1 |
| Hymenoptera | Ichneumonidae | 1 | 0.9 | 5.3 | |||||
| Hymenoptera | Tenthredinidae | 42 | 37.2 | 23.5 | 52.6 | 35.0 | 21.1 | 40.0 | 50.0 |
| Lepidoptera | Chimabachidae | 3 | 2.7 | 17.6 | |||||
| Lepidoptera | Crambidae | 1 | 0.9 | 5.3 | |||||
| Lepidoptera | Drepanidae | 12 | 10.6 | 29.4 | 5.3 | 15.8 | 15.0 | ||
| Lepidoptera | Erebidae | 36 | 31.9 | 58.8 | 21.1 | 10.0 | 57.9 | 20.0 | 27.8 |
| Lepidoptera | Geometridae | 93 | 82.3 | 94.1 | 89.5 | 85.0 | 78.9 | 80.0 | 66.7 |
| Lepidoptera | Lasiocampidae | 97 | 85.8 | 58.8 | 78.9 | 90.0 | 94.7 | 90.0 | 100.0 |
| Lepidoptera | Lycaenidae | 14 | 12.4 | 23.5 | 5.3 | 25.0 | 5.3 | 10.0 | 5.6 |
| Lepidoptera | Noctuidae | 73 | 64.6 | 76.5 | 52.6 | 85.0 | 57.9 | 80.0 | 33.3 |
| Lepidoptera | Notodontidae | 19 | 16.8 | 17.6 | 5.3 | 25.0 | 21.1 | 15.0 | 16.7 |
| Lepidoptera | Pyralidae | 18 | 15.9 | 35.3 | 21.1 | 30.0 | 5.3 | 5.0 | |
| Lepidoptera | Tortricidae | 84 | 74.3 | 88.2 | 94.7 | 70.0 | 78.9 | 60.0 | 55.6 |
| Lepidoptera | Yponomeutidae | 1 | 0.9 | 5.0 | |||||
| Lepidoptera | Ypsolophidae | 30 | 26.5 | 11.8 | 15.8 | 35.0 | 15.8 | 35.0 | 44.4 |
| Neuroptera | Hemerobiidae | 1 | 0.9 | 5.6 | |||||
| Orthoptera | Acrididae | 7 | 6.2 | 5.9 | 15.0 | 16.7 | |||
| Species | Habitat | Shannon Index H’ | Evenness E | Jaccard Similarity Index CCJ | ||
|---|---|---|---|---|---|---|
| Orchard | Urban | |||||
| (a) Lepidoptera | Blue Tit | forest | 2.2 | 0.92 | 0.83 | 0.75 |
| orchard | 1.9 | 0.81 | 0.75 | |||
| urban | 2.1 | 0.89 | ||||
| Great Tit | forest | 2.0 | 0.86 | 1.00 | 0.80 | |
| orchard | 2.0 | 0.87 | 0.80 | |||
| urban | 1.9 | 0.90 | ||||
| (b) other arthopods | Blue Tit | forest | 2.4 | 0.91 | 0.56 | 0.58 |
| orchard | 2.3 | 0.88 | 0.67 | |||
| urban | 2.5 | 0.91 | ||||
| Great Tit | forest | 2.2 | 0.90 | 0.47 | 0.44 | |
| orchard | 2.7 | 0.89 | 0.61 | |||
| urban | 2.5 | 0.84 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Höhn, D.; Masello, J.F.; Kümmel, M.N.; Griep, S.; Goesmann, A.; Quillfeldt, P. Nestling Diet of Two Sympatric Insectivorous Passerines in Different Habitats—A Metabarcoding Study. Birds 2024, 5, 67-89. https://doi.org/10.3390/birds5010005
Höhn D, Masello JF, Kümmel MN, Griep S, Goesmann A, Quillfeldt P. Nestling Diet of Two Sympatric Insectivorous Passerines in Different Habitats—A Metabarcoding Study. Birds. 2024; 5(1):67-89. https://doi.org/10.3390/birds5010005
Chicago/Turabian StyleHöhn, Daniel, Juan F. Masello, Marc N. Kümmel, Sven Griep, Alexander Goesmann, and Petra Quillfeldt. 2024. "Nestling Diet of Two Sympatric Insectivorous Passerines in Different Habitats—A Metabarcoding Study" Birds 5, no. 1: 67-89. https://doi.org/10.3390/birds5010005
APA StyleHöhn, D., Masello, J. F., Kümmel, M. N., Griep, S., Goesmann, A., & Quillfeldt, P. (2024). Nestling Diet of Two Sympatric Insectivorous Passerines in Different Habitats—A Metabarcoding Study. Birds, 5(1), 67-89. https://doi.org/10.3390/birds5010005