Simple Summary
Abandoned underground coal mines collect ground water that eventually finds its way to the surface. A common way to treat pollutants and sediments from this mine water is to create passive treatment ponds that remove contaminates before the water enters a surface stream. Tree Swallows and many other birds and wildlife live and breed near these treatment ponds, with potentially detrimental effects. We monitored 63 nests over two breeding seasons (2022–2023) to determine if nests located next to the ponds had better or worse reproductive outcomes than nests farther away. We found that nests close to the ponds (<70 m) had higher numbers of young that survived to the age of fledging than nests that were built farther away from the ponds (>820 m). While this suggests that the treatment ponds provided meaningful resources for Tree Swallows to reproduce, the numbers of offspring were unusually low compared to what was found in studies by other investigators at other sites. More research will be needed to determine why.
Abstract
Abandoned mine drainage treatment ponds could have contrasting effects on the reproductive success of birds living in the vicinity. The ponds and associated vegetation may, like any other body of freshwater, provide beneficial habitats for the insects that the birds use to feed their young; or instead, the ponds may act as an ecological trap, attracting the birds to a habitat that is poor in quality and negatively impacting their productivity. We monitored nests of an aerial insectivore, the Tree Swallow (Tachycineta bicolor), to determine whether the distance between the ponds and the nests affected various reproductive parameters including clutch size, hatch rate, number of nestlings, nestling size and mass, number of fledglings, fledging rate, and fledge date. Data were collected over two breeding seasons (2022 and 2023) from a swallow population in southwestern Pennsylvania, USA. We found that the nests closest to the treatment ponds had significantly more nestlings and fledglings, earlier fledge dates, and a better fledging rate when compared to nests that were more distant from the ponds. However, all these parameters were well below previously published values, which suggests that the mine drainage ponds provide good nesting habitats relative to what is available in the region but that they do not represent high-quality habitats for this species overall.
1. Introduction
Subsurface coal mines quickly fill with groundwater after mining operations cease. Such mine water can be contaminated from a process by which sulfur-containing rocks interact chemically with water to form sulfuric acid [1]. The acid leaches minerals like iron, manganese, and aluminum from rock layers and finds its way to the surface, where there can be detrimental effects on bacteria, plants, and animal life [2,3,4]. Treatment of mine drainage through the construction of passive settling ponds can improve water quality before it enters surface waterways [5]. A successful example of this system is found on the campus of Saint Vincent College in Latrobe, PA, USA, where a connected series of three abandoned mine drainage (AMD) ponds with multiple dikes were built between 1997 and 1998 [6].
There are thousands of AMD treatment facilities around the world, wherever coal has been mined, including Sweden, Slovakia, the Philippines, Australia, and the United Kingdom [5]. Many have passive settling ponds like the ones at Saint Vincent College. By design, the pond receiving mine inputs is far more polluted than the pond further downstream in the treatment system [2]. The polluted fraction of water can inhibit insect populations by depressing larval development stages [3,7]. For example, aquatic life can be poisoned by heavy metals, reproductively inhibited by fluctuating pH, and suffocated from the high amounts of metal particulates in the mine water [8]. In untreated waters, depressed insect populations can impact the vertebrate populations that rely upon them for food [9,10].
It is unclear whether AMD treatment ponds can be a valuable ecological asset for species that rely on aquatic insect life for food. Numerous scientific studies of bacteria and other simple organisms in AMD settling ponds have been published [4], but few have centered on birds. One study measured avian diversity at treatment ponds in comparison to natural wetlands and found that AMD ponds had reduced diversity of bird species [11]. Another study showed that AMD treatment ponds hosted a greater diversity of bird species at the ponds in comparison to natural wetlands nearby [12]. Neither of these studies investigated the reproductive success of the birds, such as the number of fledged offspring per nesting attempt. Measures of species diversity are not adequate to establish the value of a habitat. An AMD pond habitat could be an ecological trap, a common circumstance seen in disturbed areas in which animals settle in a particular habitat but do not have adequate levels of health and reproduction to replace themselves over their lifetimes [13,14,15].
This study measured bird productivity along a geographical gradient from the AMD ponds to more than 1 km away from the ponds. Bird productivity was defined by life history parameters like number of chicks per nest, egg laying date, and nestling size and weight. Prior studies have shown that birds that lay eggs earlier in the season have better reproductive success than those who delay [16,17]. Higher nestling weights near the time of fledging also correspond to better reproductive outcomes [18], and of course, if parents have more offspring, they will usually have better lifetime success than those who have few.
We chose to study the productivity of Tree Swallows (Tachycineta bicolor), as they readily nest in artificial wooden boxes that facilitate data collection. Further, food delivered to nestling Tree Swallows consists predominately of flying invertebrates that have aquatic larval stages, such as damselflies (order Odonata) and mayflies (order Ephemeroptera) [19]. This fact has led to the frequent use of Tree Swallows in studies of environmental contamination, since pollutants impacting water quality can have adverse effects on the organisms that live in the water, as well as those that depend on them in higher trophic levels [20]. Tree Swallows rarely travel more than 400 m from an active nest to find food [21], so individuals living near polluted waterways are likely to offer contaminated or otherwise impacted prey to their nestlings.
Our hypothesis was that birds locating their nests in boxes placed near Saint Vincent’s AMD ponds will show poorer productivity compared to birds that locate their nests farther away. We reasoned that the insect prey that parents use to produce eggs and feed their nestlings would be of a poor quality and quantity near the ponds due to the contamination and sediments found in the water. The reduced nutrient supply would be reflected in those birds’ reproductive outputs, including nestling size and mass [22,23].
2. Materials and Methods
2.1. Study Site
The study site was located near Saint Vincent College, a small residential institution with approximately 1400 students enrolled. The campus is in a suburban area about 53 km SE of the city of Pittsburgh, PA, USA. The nearest town to the college, Latrobe, has a population of 28,000 and is spread across 193 km2 at a density of 147 per km2. The site is in the Köppen–Geiger humid continental climate zone with hot summers and no dry season [24]. Temperatures at the nearest airport range from a mean daily low temperature of −5.9 °C in winter (January) to a mean daily high temperature of 28.3 °C in summer (July) [25]. The college campus is composed of tightly packed academic buildings and residence halls sited on top of a hill, with an adjacent cemetery and athletic fields positioned directly west and south, respectively (Figure 1). Going further in those directions, agricultural fields predominate, with small muddy hedgerows between them. The hedgerows do not contain permanent surface waters but are wet enough year-round to support cattails (Typha spp.), other wetland plants, and possibly the insect prey of Tree Swallows. Down the campus hill and to the north are the series of AMD ponds which drain into a stream carrying water affected by AMD entering from mines further west. An adjacent 4.9 ha recreational lake is relatively free from AMD and supports fish. Surprisingly, point-count surveys of birds at the lake that we conducted for a separate, concurrent study rarely recorded Tree Swallows (only 1 of 429 individual birds identified during point counts was a Tree Swallow) [26]. Land coverage within a 2.5 km radius of the site includes 38% forest; 33% human development or other non-agricultural cover; 21% croplands planted in corn; 6% soybean, wheat, or cover crops; and 1% bodies of water [27]. The forest canopy is composed of mixed oak and maple species, characteristic of the Eastern Temperate Forest ecoregion of North America [28]. The understory is dense with invasive shrubs, including several varieties of bush honeysuckle (Lonicera spp.).
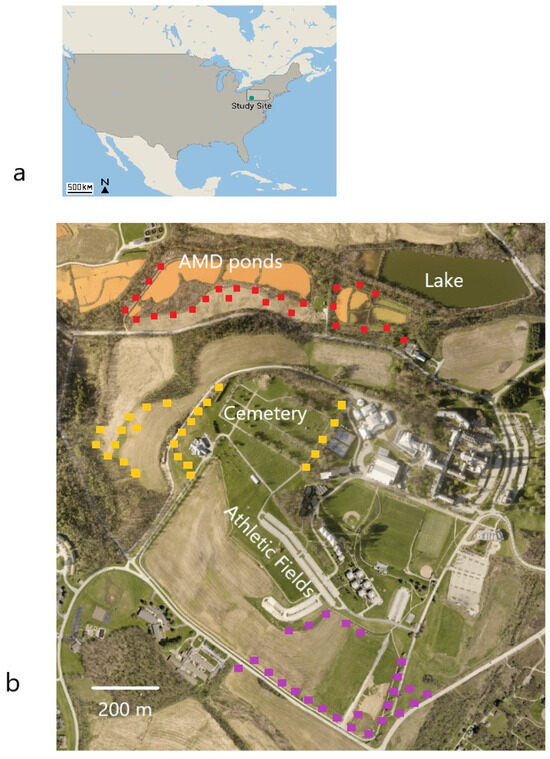
Figure 1.
(a) Location of the study site within the United States. (b) Aerial view of the study site. Tree Swallow nest boxes were placed near abandoned mine drainage (AMD) treatment ponds and at locations that were more distant from the ponds. Nest box locations are shown on the map as red squares (NEAR locations), yellow squares (MID locations), and purple squares (FAR locations). Box locations are approximated.
The AMD ponds are spread over 9 ha of land. Water tests indicate that the water entering the ponds during May–July in 2022 and 2023 averaged above the local authority’s maximum water quality criteria in total iron, manganese, and sulfate and below the water quality criteria for dissolved oxygen (DO) (Table 1).

Table 1.
Chemical properties of pond water found at study site and allowable concentrations according to Pennsylvania Water Quality Standards. Shaded cells indicate parameters outside of water quality standards.
2.2. Study Species
The Tree Swallow is a North American insectivorous Passerine species in the swallow and martin family (Hirundinidae). Different studies in different locations have shown some variation in the diet items consumed during the breeding season, but true flies (order Diptera), dragonflies and damselflies (order Odonata), mayflies (order Ephemeropta), and caddisflies (order Trichoptera) are the most frequent [17]. These insects are typically captured in midair from 0 to 12 m above the ground or water surface, but swallows may forage up to 100 m above these surfaces on occasion [21]. Tree Swallows are obligate migrants; they breed throughout Canada and the northern United States, and following reproduction, they travel to points south from the U.S. southeastern coast to Mexico and Central America. The global population size is estimated to be near 19 million and declining overall [29]. However, populations in Pennsylvania are stable [30]. The species is a secondary cavity nester that readily nests in artificial boxes, a trait that has facilitated many scientific studies [17]. It produces only a single brood per year, with incubation performed only by the female and other parental duties shared between male and female. Tree Swallows are socially monogamous but have one of the highest levels (>80%) of extra-pair paternity recorded among Passerines [17].
2.3. Nest Box Setup and Monitoring
Twenty-five wooden nest boxes were set up in early 2022 along and near (<70 m) the edge of AMD ponds, and another twenty-five were placed near the cemetery and adjacent fields at a distance from the ponds ranging from 260 to 490 m. Another 25 boxes were erected in time for the 2023 breeding season at an even farther distance from the AMD ponds, ranging from 820 to 1700 m. These latter boxes were positioned along a campus fitness path encircling a set of sports practice fields and an agricultural field. Distances were estimated to the nearest 10 m using the measurement tool on Google Maps [31].
All boxes were purchased from the Pennsylvania Game Commission’s Howard Nursery (Bellefonte, PA, USA) and featured an entrance hole of 3.6 cm in diameter. The boxes were hung 1.2 m meters above the ground on steel posts. Steel cone predator baffles were installed immediately below each box. The box orientation was set to face east, as that is the Tree Swallow’s documented preference [32]. Most boxes were placed in a line with an average of 30 m between each one (10 m minimum). Almost all boxes were used by nesting birds, about half by Tree Swallows. The other half were occupied by non-target species like House Wrens (Troglodytes aedon) and Eastern Bluebird (Sialia sialis). These other species, unlike the Tree Swallow, do not rely on insects with aquatic larval stages to feed their young [33,34], making them less suitable for our project.
Nest boxes were checked every 2–3 days from 15 March to 8 August each year. We noted the presence of adults, of nest material, completed nests, eggs, and nestlings by inserting a cell phone into a ventilation slot between the front and top panels of a box and taking a photo.
When nestlings were approximately 11 days old (range = 6–16), we opened the box, removed the nestlings, and placed them together in a plastic container for weighing with a digital scale. The average nestling weight was found by dividing the overall mass by the number of nestlings. We selected a single nestling for morphometric measurements to minimize the time spent at the nest. We were careful to avoid any nestling that appeared smaller or larger than the rest. The unflattened wing chord and several other parameters were measured using calipers to the nearest 1 mm. U.S. Fish and Wildlife numbered leg bands were applied to nestlings for use in future studies.
We tried to minimize disturbance by only handling nestlings once during their development, so that banding and measurements would be conducted on the same day. We targeted our visits to coincide with a nestling age between 7 and 12 days old. Leg bands do not often fit nestling Tree Swallows appropriately when they are younger than 7 days, and handling nestlings past day 12 risks premature fledging [22].
Knowing the dates of egg laying, hatching, and fledging was important for the calculation of nestling age, but usually, only two of these dates were known precisely, since we did not visit each nest every day. Nestling age was therefore an estimate, based on known dates when available, and on published species averages when not available. The incubation period for Tree Swallows is around 14 days (range = 11–20), and the average nestling period is around 20 days (range = 15–25) [17]. Tree Swallows have asynchronous hatching, with an average of 28 h between the first-hatched and last-hatched egg [35]. Our estimates of nestling age were based on the date when the first nestling hatched. It is possible that some nestlings considered here to be X days old may be X-1 days old due to hatching asynchrony, but given our effort to avoid measuring the smallest nestling in each brood, the number of mis-aged nestlings in our dataset should be close to zero.
Between 30% and 24% of nests in 2022 and 2023, respectively, became infested with hematophagous mites (Dermanyssus spp.), especially those that had not yet fledged by mid-June of each year. The degree of infestation was not quantified. We did not interfere with the mites, as we did not wish to bias our measurements of nestling health and parental reproductive success by treating some nests differently than others.
2.4. Data Analysis
More nests were monitored in 2023 than in 2022 (n = 45 and 20, respectively) due to the addition of more nest boxes in 2023. Morphometric data were collected on nestlings from only a subset of nests (n = 24 in 2023; n = 5 in 2022), because we were unable to consistently access nests within the short interval of time when nestlings were aged 7–12 days. This is especially true in 2022 when we were overly cautious with choosing when a nestling was the ideal size and age for handling.
Initial analyses began with examining reproductive parameters from nests in three distance categories relating to the AMD ponds: adjacent to the ponds (<70 m, n = 35), at an intermediate distance from the ponds (260–490 m, n = 10), and at a far distance from the ponds (820–1700 m, n = 18). These categories are hereafter referred to as NEAR, MID, and FAR nests, respectively.
Specific parameters calculated and compared across these three distance categories included the number of nests with at least one egg, clutch size, number of hatched nestlings, number of nests with at least one fledgling, earliest fledge date, number of fledglings, and three other measures of productivity (Table 2).
Table 2.
Calculations for Tree Swallow reproductive parameters using number of eggs, nestlings, and fledglings.
Each parameter was evaluated for the assumptions of normal distribution using kurtosis, skewness, and F-tests. All distributions were normal except for fledge date, which was consistent with a Poisson distribution. A repeated-measures multivariate analysis of variance (MANOVA) was run using JMP software (version 17.2.0) [36] to determine whether the dependent variables of clutch size, number of hatched nestlings, and number of fledglings could be predicted by the independent variables of year (2022, 2023) and distance to the AMD ponds (NEAR, MID, FAR). A general linear model (GLM) assuming the Poisson distribution for fledge date was used to determine if the fledge date was dependent on the year (2022, 2023) and distance to the AMD ponds (NEAR, MID, FAR).
For morphometric analyses, we used JMP software [36] to create general linear models, testing for relationships among the dependent variables of nestling wing chord and body mass and the dependent variables of nestling age and distance group (NEAR, MID, or FAR). Interaction terms were included each time.
3. Results
3.1. Nest Parameters
All measured life history traits followed the same general pattern: whereas the NEAR group had the most favorable reproductive outcomes, the MID group had intermediate levels, and the FAR group had the least favorable outcomes (Table 3). In the MANOVA test comparing clutch size, number of hatched young, and number of fledglings to distance group, we found that this variable explained a significant amount of variance (F = 3.61, df = 55, p = 0.034). According to the model, over the three developmental stages of egg, nestling, and fledgling, the NEAR group had the greatest numerical success. The mean clutch size of the three distance groups was similar, varying by only 0.2 eggs (Table 3). However, all three groups showed a significant loss of offspring as time progressed from the nestling stage to the fledgling stage (F = 31.28, df = 54, p < 0.001). There appeared to be an increasing difference in this negative slope among the three groups in the later stages of reproduction (number of nestlings and fledglings) (Figure 2), but our test did not show this interaction to be significant (F = 1.66, df = 108, p = 0.166). There was also no significant effect of year (F = 0.43, df = 54, p = 0.517).
Table 3.
Productivity of Tree Swallow nests at three distances from abandoned mine drainage ponds, 2022–2023. NEAR nests were those less than 70 m from a pond edge (n = 35), MID nests were those from 260 to 490 m from a pond edge (n = 10), and FAR nests were those from 820 to 1700 m from a pond edge (n = 18).
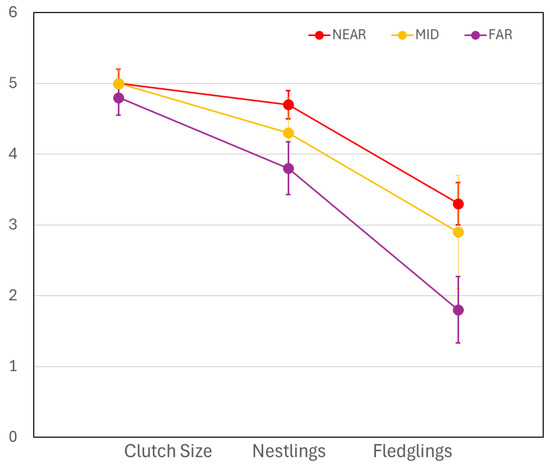
Figure 2.
Average numbers ± SE of Tree Swallow eggs, hatched young, and fledglings at nests NEAR to (red points), at MID distance from (yellow points), and FAR from (purple points) abandoned mine drainage ponds. Overall means of three distance groups were significantly different.
The mean hatch rate, mean fledge rate, and nest success rate all generally followed the same pattern as the numbers of eggs, nestlings, and fledglings. That is, rates were higher in the NEAR group and lower in the FAR group, with mostly intermediate values in the MID group (Table 3). The earliest fledge date in the NEAR group was 10 days earlier than that of the FAR group, and NEAR nests’ fledge dates averaged 16 days earlier (Table 3). Values for the MID group were intermediate. The GLM model showed that the distance group explained a significant amount of variance in fledge date (F = 18.39, df = 2, p < 0.001). No significant difference between years was found (F = 2.48, df = 1, p = 0.115).
The above statistics show that some nests in all groups failed to produce hatchlings or fledglings. Six failures were due to a nest being overtaken by a competing species, the Eastern Bluebird (Sialia sialis), and nests being destroyed by unknown predators. Ten nests failed due to the unexplained death of all nestlings, which happened in two nests of the NEAR group, two nests of the MID group, and six nests of the FAR group. In 6 of these 10 cases, the dead nestlings were found in a nest infested with hundreds of mites (Dermanyssus spp.). However, there were nests that produced fledged young that also had mites, and these were predominately in the NEAR group. Only 1 of 11 (9%) nests in the NEAR group that were infested with mites failed, whereas the failure rate following mite infestation was 100% in the MID group (2 of 2 nests failed) and 60% in the FAR group (3 of 5 nests failed).
3.2. Morphometrics
Nestling wing length and nestling age were strongly correlated, with the older nestlings showing longer wings (Figure 3; F = 21.14, df = 23, p < 0.001). There was no interaction between age and nest groups (F = 1.66, df = 23, p = 0.436). The distance groups showed no difference in wing length (Table 3; F = 3.16, df = 23, p = 0.206). The nestling body mass increased significantly with nestling age (Figure 4; F = 9.27, df = 23, p = 0.002). There was no difference in average nestling mass between distance groups (Table 3; F = 1.59, df = 23, p = 0.451), and the interaction term was not significant (F = 4.04, df = 23, p = 0.132).
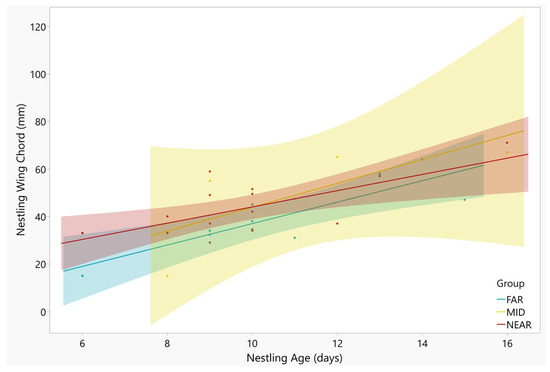
Figure 3.
Wing lengths of nestling Tree Swallows and age at time of measurement. Only one nestling was measured per nest. The positive relationship between the two variables was significant (p < 0.001), with no difference between slopes of nests in the NEAR (red points), MID (yellow points), and FAR (blue points) groups (p = 0.436). Regression lines and 95% confidence intervals are shown for each group. The R2 values for the regression lines are as follows: NEAR = 0.45, MID = 0.55, FAR = 0.73).
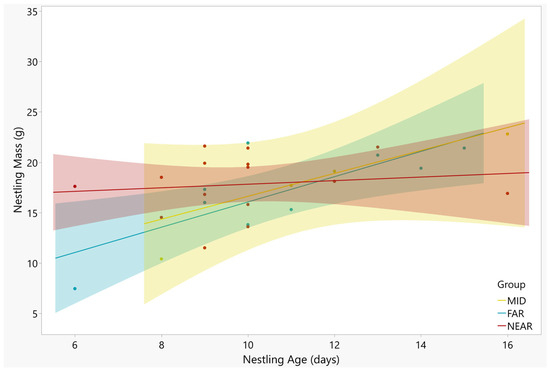
Figure 4.
The mass of nestling Tree Swallows and age at time of measurement. Only one nestling was measured per nest. The positive relationship between the two variables was significant (p = 0.002), with no difference found between nests in the NEAR (red points), MID (yellow points), and FAR (blue points) groups (p = 0.451). Regression lines and 95% confidence intervals are shown for each group. The R2 values for the regression lines are as follows: NEAR = 0.02, MID = 0.59, FAR = 0.59).
4. Discussion
We predicted that Tree Swallows nesting near the AMD ponds would show reduced measures of nest productivity compared to those that nested farther away from the ponds. The data firmly refute our prediction, since many of the reproductive parameters were higher among the NEAR nests compared to the MID and FAR nests. The differences between nest groups became greater as the stages of the nesting cycle progressed. This pattern suggests that whatever factors led to the reduced nest success in the MID and FAR groups were experienced during the development of hatched young and not at earlier stages.
4.1. Values of Reproductive Parameters in Comparison to Other Studies
With generally better reproductive outcomes in the NEAR group compared to the MID and FAR groups and no significant difference in nestling size or mass, it is tempting to conclude the AMD ponds had a favorable effect on Tree Swallow’s reproductive success. However, the absolute values for fledging rate and number of fledglings were lower than most published studies at non-polluted sites. In our study, the NEAR nests had a fledgling rate of 71%, and the mean number of fledglings was 3.3 ± 0.3. Elsewhere, a long-term study in the neighboring U.S. state of New York with a sample of 5081 nests had a fledgling success rate of 88%, and the average number of fledglings was 4.3 [17]. Other studies reported a success rate from 88 to 95%, with the average number of fledglings ranging from 4.0 to 4.9 [17,37,38,39,40,41,42,43]. The only studies with average values lower than what was found here are those that took place during periods with unusual temperature regimes or persistent rainfall [44,45] or those with nests exposed to residual mercury pollution in a nearby river [43]. In the latter case, the fledgling rate was 80%, and the average number of fledglings was 3.6, figures that are still higher than the NEAR nests of this study.
Numerous studies of Tree Swallows indicate that nesting success is negatively affected by unseasonably cold and rainy weather during the nestling stage [38,44,46]. This was not a factor during our study; temperatures were <±0.8 °C from climatic averages in May and June of 2022 and 2023, and the precipitation was below average during the same months, especially in 2023 [47]. In fact, the rainfall in May 2023 was only 45% of normal [26].
Aside from these population-level parameters, studies show that there are subsets of individuals that have consistently lower values in fledgling success rate and the number of fledglings. These include Tree Swallows that nest in landscapes with high percentages of agricultural monoculture lacking habitat heterogeneity within 5 km of a nest [42]. Our study site did contain corn (Zea mays), soybean (Glycine max), and wheat (Triticum aestivum) fields, but there were many hedgerows, riparian zones, and forest patches that preclude this explanation for the below-normal fledgling numbers. Late-nesting Tree Swallows also have lower fledging rates [16]. This likely explains the lower values we found in our FAR nests, but not the NEAR nests, since those had an earlier average lay date.
In sum, the data presented here for the NEAR nests are lower than expected, given previously published studies. We have rejected explanations relating to weather and habitat structure. It remains possible that polluted waters of the AMD ponds impact the nestling food supply and, ultimately, nestling survival.
4.2. Study Limitations and Suggestions for Future Research
Data on food availability and quality are a missing component of this study. It is not clear whether these variables vary with the distance to the AMD ponds. A related factor is how much nestlings in the NEAR group might have been compromised by metals in their diet due to bioaccumulation [48].
We measured nestlings at each nest only once, so the growth trajectory of individual nests cannot be examined. Significant nestling growth occurs during the first week after hatching [37], but we did not take body mass or wing length data on nestlings less than 6 days of age. With more data on younger nestlings, we might have found a difference in treatment groups or an interaction between them. These would be important data to have, since growth rates can change depending on food resources [23,49]. Such data would help determine whether the development of nestlings in our study differed from nestlings in other studies. Differences could help us understand why the reproductive outputs of NEAR nests were comparatively low. There may also have been internal physiological differences besides mass or structural size that may explain the lower values.
This study only included data from two years, both of which featured numerous nests with mite infestations. The mites can cause nests to fail due to abandonment by parents or the premature fledging of young [47], but many studies fail to find an effect of mite infestation on nestling survival [50,51,52]. Parasites in general may cause slower nestling growth and have a synergistic effect with other factors that might depress nest survivorship [50,53]. It is therefore possible that the MID and FAR nests, which had more nestling mortality associated with mite infestations, could have had unusually low reproductive outputs during our short study but could have higher values in a year without an abundance of mites. Factors like this, that are unrelated to the AMD treatment ponds, could obscure or reverse the patterns associated with the treatment ponds. Additional years of data collection would help.
The best experimental setup to test effects of AMD would be for one set of nest boxes to be placed around AMD ponds and another set placed around control ponds that contained no AMD. This was not possible at our study site. The FAR nests were farther away, not just from the AMD ponds, but also from any other permanent body of water. The birds nesting near the ponds fared better than the others. However, the limitations of the experimental setup should be considered before refuting the general hypothesis that a polluted waterbody represents an ecological trap and reduces reproductive success.
More research is planned to determine whether other species of birds show the same pattern as the Tree Swallow with regard to how the three distance groups showed different levels of productivity, as well as why those levels were all low compared to studies elsewhere. We will also study how the prey biomass depends on the distance to the treatment ponds and whether nestlings have measurable levels of heavy metals in their tissues.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/birds5030030/s1: Table S1: Nesting success parameters of Tree Swallow nests, 2022–2023, at different distances from abandoned mine drainage treatment ponds; Table S2: Morphometric parameters of nestling Tree Swallows, 2022–2023, at different distances from abandoned mine drainage treatment ponds.
Author Contributions
Conceptualization, J.S.K.; methodology, J.S.K.; formal analysis, J.S.K.; investigation, J.E.L., A.R.D., J.S.K. and I.L.; data curation, J.E.L., J.S.K., I.L. and A.R.D.; writing—original draft preparation, J.S.K.; writing—review and editing, J.S.K.; visualization, J.S.K.; supervision, J.S.K.; project administration, J.S.K.; funding acquisition, J.S.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by a generous gift from M.J. Seipler, as well as the Saint Vincent College Department of Biological Sciences and a Saint Vincent College Faculty Research Grant to J.S.K.
Institutional Review Board Statement
Birds were banded, and nests monitored under U.S. Fish and Wildlife Service bird-banding permit #23382 issued to J.S.K., and the work was approved by the Saint Vincent College Animal Care and Use Committee under protocol #22-017.
Data Availability Statement
This study’s complete dataset is contained within Tables S1 and S2. Water quality data for the AMD ponds in this study are archived at https://www.datashed.org/project-monastery-run-wetland-2 (accessed on 7 August 2024).
Acknowledgments
We thank Saint Vincent College, Saint Vincent Archabbey, and Wimmer Corporation for access to their land. B. Bollinger provided water test data. A. Wade provided considerable help in data entry and the formatting of this manuscript.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Bussiere, B. Acid mine drainage from abandoned mine sites: Problematic acid reclamation approaches. In Proceedings of the International Symposium on Geo-Environmental Engineering, Hangzhou, China, 8–10 September 2009; pp. 111–125. [Google Scholar]
- Porter, C.M.; Nairn, R.W. Ecosystem functions within a mine drainage passive treatment system. Ecol. Eng. 2008, 32, 337–346. [Google Scholar] [CrossRef]
- Hogsden, K.L.; Harding, J.S. Consequences of acid mine drainage for the structure and function of benthic stream communities: A review. Freshw. Sci. 2012, 31, 108–120. [Google Scholar] [CrossRef]
- Méndez-García, C.; Peláez, A.I.; Mesa, V.; Sánchez, J.; Golyshina, O.V.; Ferrer, M. Microbial diversity and metabolic networks in acid mine drainage habitats. Front. Microbiol. 2015, 6, 475. [Google Scholar] [CrossRef]
- Skousen, J.; Zipper, C.E.; Rose, A.; Ziemkiewicz, P.F.; Nairn, R.; McDonald, L.M.; Kleinmann, R.L. Review of passive systems for acid mine drainage treatment. Mine Water Environ. 2017, 36, 133–153. [Google Scholar] [CrossRef]
- Monastery Run Improvement Project. Saint Vincent College, Latrobe, Pa. Available online: https://www.stvincent.edu/academics/experiential-learning/community-outreach/monastery-run-improvement-project.html (accessed on 3 March 2024).
- Lidman, J.; Jonsson, M.; Berglund, A.M.M. The effect of lead (Pb) and zinc (Zn) contamination on aquatic insect community composition and metamorphosis. Sci. Tot. Environ. 2020, 734, 139406. [Google Scholar] [CrossRef] [PubMed]
- Batty, L.C.; Atkin, L.; Manning, D.A.C. Assessment of the ecological potential of mine-water treatment wetlands using a baseline survey of macroinvertebrate communities. Environ. Pollut. 2005, 138, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Twining, C.W.; Shipley, J.R.; Winkler, D.W. Aquatic insects rich in omega-3 fatty acids drive breeding success in a widespread bird. Ecol. Letters 2018, 21, 1812–1820. [Google Scholar] [CrossRef] [PubMed]
- Wiseman, I.M.; Rutt, G.P.; Edwards, P.J. Constructed wetlands for minewater treatment: Environmental benefits and ecological recovery. Water Environ. J. 2004, 18, 133–138. [Google Scholar] [CrossRef]
- Lacki, M.J.; Hummer, J.W.; Webster, H.J. Avian diversity patterns at a constructed wetland: Use of ecological theory in the evaluation of a mine land reclamation technique. Int. J. Surf. Min. Reclam. Env. 1991, 5, 101–105. [Google Scholar] [CrossRef]
- Jaques, R.O.; Moorhouse-Parry, A.M.; Carline, R.; Mayes, W.M.; Hull, S.L. Biodiversity benefits of coal mine water remediation schemes for bird life. In Proceedings of the International Mine Water Association Congress “Mine Water Management for Future Generations”; Stanley, P., Wolkersdorfer, C., Wolkersdorfer, K., Eds.; International Mine Water Association: Cardiff, UK, 2021; pp. 206–211. [Google Scholar]
- Sievers, M.; Parris, K.M.; Swearer, S.E.; Hale, R. Stormwater wetlands can function as ecological traps for urban frogs. Ecol. Appl. 2018, 28, 1106–1115. [Google Scholar] [CrossRef]
- Titeux, N.; Aizpurua, O.; Hollander, F.A.; Sardà-Palomera, F.; Hermoso, V.; Paquet, J.-Y.; Mestdagh, X.; Settele, J.; Brotons, L.; Van Dyck, H. Ecological traps and species distribution models: A challenge for prioritizing areas of conservation importance. Ecography 2020, 43, 365–375. [Google Scholar] [CrossRef]
- Zhang, C.; Wen, L.; Wang, Y.; Liu, C.; Zhou, Y.; Lei, G. Can constructed wetlands be wildlife refuges? A review of their potential biodiversity conservation value. Sustainability 2020, 12, 1442. [Google Scholar] [CrossRef]
- Dawson, R.D. Timing of breeding and environmental factors as determinants of reproductive performance of tree swallows. Can. J. Zool. 2008, 86, 843–850. [Google Scholar] [CrossRef]
- Winkler, D.W.; Hallinger, K.K.; Ardia, D.R.; Robertson, R.J.; Stutchbury, B.J.; Cohen, R.R. Tree Swallow (Tachycineta bicolor), version 1.0. In Birds of the World; Poole, A.F., Ed.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar] [CrossRef]
- Tinbergen, J.M.; Boerlijst, M.C. Nestling weight and survival in individual Great Tits (Parus major). J. Anim. Ecol. 1990, 59, 1113–1127. [Google Scholar] [CrossRef]
- Mengelkoch, J.M.; Niemi, G.; Ronald, J.; Regal, R. Diet of the nestling Tree Swallow. Condor 2004, 106, 423–429. [Google Scholar] [CrossRef]
- McCarty, J.P. The number of visits to the nest by parents is an accurate measure of food delivered to nestlings in Tree Swallows. J. Field Ornithol. 2002, 73, 9–14. [Google Scholar] [CrossRef]
- McCarty, J.P.; Winkler, D.W. Foraging ecology and diet selectivity of Tree Swallows feeding nestlings. Condor 1999, 101, 246–254. [Google Scholar] [CrossRef]
- Ardia, D.R. Geographic variation in the trade-off between nestling growth rate and body condition in the Tree Swallow. Condor 2006, 108, 601–611. Available online: http://www.jstor.org/stable/4151081 (accessed on 7 August 2024). [CrossRef]
- Bize, P.; Metcalfe, N.B.; Roulin, A. Catch-up growth strategies differ between body structures: Interactions between age and structure-specific growth in wild nestling Alpine Swifts. Funct. Ecol. 2006, 20, 857–864. Available online: http://www.jstor.org/stable/3806594 (accessed on 7 August 2024). [CrossRef]
- Beck, H.; Zimmermann, N.; McVicar, T.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and future Köppen-Geiger climate classification maps at 1-km resolution. Sci. Data 2018, 5, 180214. [Google Scholar] [CrossRef]
- National Oceanic and Atmospheric Administration. NOAA Online Weather Data. Available online: https://www.weather.gov/wrh/Climate?wfo=pbz (accessed on 16 March 2023).
- Kellam, J.S. (Saint Vincent College, Latrobe, Pennsylvania, USA). Point count data collected between 12 June and 15 July in years 2022 and 2023. Unpublished work.
- AcreValue Parcel Map, Ag-Analytics Technology Company, Ithaca, NY, USA. Available online: https://www.acrevalue.com/map/?lat=40.294411&lng=-79.398106&zoom=13 (accessed on 4 March 2024).
- U.S. Environmental Protection Agency. Ecoregions of North America. Available online: https://www.epa.gov/eco-research/ecoregions-north-america (accessed on 16 March 2023).
- Rosenberg, K.V.; Dokter, A.M.; Blancher, P.J.; Sauer, J.R.; Smith, A.C.; Smith, P.A.; Stanton, J.C.; Panjabi, A.; Helft, L.; Parr, M.; et al. Decline of the North American avifauna. Science 2019, 366, 120–124. [Google Scholar] [CrossRef]
- Skinner, J. Tree Swallow Tachycineta bicolor. In Second Atlas of Breeding Birds in Pennsylvania; Wilson, A.M., Brauning, D.W., Mulvihill, R.S., Eds.; Pennsylvania University Press: University Park, PA, USA, 2012. [Google Scholar]
- Google Maps. Available online: www.google.com/maps (accessed on 4 March 2024).
- Ardia, D.R.; Pérez, J.H.; Clotfelter, E.D. Nest box orientation affects internal temperature and nest site selection by Tree Swallows. J. Field Ornithol. 2006, 77, 339–344. [Google Scholar] [CrossRef]
- Pinkowski, B.C. Feeding of nestling and fledgling Eastern Bluebirds. Wilson Bull. 1978, 90, 84–98. [Google Scholar]
- Grabarczyk, E.E.; Gill, S.A.; Vonhof, M.J.; Alabady, M.S.; Wang, Z.; Schmidt, J.M. Diet composition and diversity does not explain fewer, smaller urban nestlings. PLoS ONE 2022, 17, e0264381. [Google Scholar] [CrossRef] [PubMed]
- Clotfelter, E.D.; Whittingham, L.A.; Dunn, P.O. Laying order, hatching asynchrony and nestling body mass in Tree Swallows Tachycineta bicolor. J. Avian Biol. 2000, 31, 329–334. Available online: http://www.jstor.org/stable/3677414 (accessed on 7 August 2024). [CrossRef]
- JMP Trial version 17.2.0; JMP Statistical Discovery, LLC: Cary, NC, USA, 2024; Available online: https://www.jmp.com/en_us/software/data-analysis-software.html (accessed on 13 March 2024).
- Quinney, T.E.; Hussell, D.J.T.; Ankney, C.D. Sources of variation in growth of Tree Swallows. Auk 1986, 103, 389–400. [Google Scholar] [CrossRef]
- Wheelwright, N.T.; Freeman-Gallant, C.R.; Mauck, R.A. Nestling Savannah Sparrows and Tree Swallows differ in their sensitivity to weather. Ornithology 2022, 139, ukac032. [Google Scholar] [CrossRef]
- Zach, R. Hatching asynchrony, egg size, growth, and fledging in Tree Swallows. Auk 1982, 99, 695–700. [Google Scholar]
- Doherty, P.F.; Grubb, T.C. Reproductive success of cavity-nesting birds breeding under high-voltage powerlines. Am. Midl. Nat. 1998, 140, 122–128. [Google Scholar] [CrossRef]
- Nooker, J.K.; Dunn, P.; Whittingham, L.A. Effects of food abundance, weather, and female condition on reproduction in tree swallows (Tachycineta bicolor). Auk 2005, 122, 1225–1238. [Google Scholar] [CrossRef]
- Ghilain, A.; Bélisle, M. Breeding success of tree swallows along a gradient of agricultural intensification. Ecol. Appl. 2008, 18, 1140–1154. [Google Scholar] [CrossRef] [PubMed]
- Hallinger, K.K.; Cristol, D.A. The role of weather in mediating the effect of mercury exposure on reproductive success in tree swallows. Ecotoxicology 2011, 20, 1368–1377. [Google Scholar] [CrossRef] [PubMed]
- Cox, A.R.; Robertson, R.J.; Rendell, W.B.; Bonier, F. Population decline in tree swallows (Tachycineta bicolor) linked to climate change and inclement weather on the breeding ground. Oecologia 2020, 192, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Griebel, I.A.; Dawson, R.D. Predictors of nestling survival during harsh weather events in an aerial insectivore, the Tree Swallow (Tachycineta bicolor). Can. J. Zool. 2019, 97, 81–90. [Google Scholar] [CrossRef]
- McArthur, S.L.; McKellar, A.E.; Flood, N.J.; Reudink, M.W. Local weather and regional climate influence breeding dynamics of Mountain Bluebirds (Sialia currucoides) and Tree Swallows (Tachycineta bicolor): A 35-year study. Can. J. Zool. 2017, 95, 271–277. [Google Scholar] [CrossRef]
- Moss, W.W.; Camin, J.H. Nest parasitism, productivity, and clutch size in purple martins. Science 1970, 168, 1000–1003. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Zhu, Y.; Yu, R.; Zheng, X. A Pilot Study on Bioaccumulation and Tissue Distribution of Mercury in Barn Swallow (Hirundo rustica). Toxics 2024, 12, 206. [Google Scholar] [CrossRef] [PubMed]
- McCarty, J.P. Variation in growth of nestling Tree Swallows across multiple temporal and spatial scales. Auk 2001, 118, 176–190. [Google Scholar] [CrossRef]
- Merino, S.; Potti, J. Mites and blowflies decrease growth and survival in nestling pied flycatchers. Oikos 1995, 73, 95–103. Available online: https://www.jstor.org/stable/3545730 (accessed on 7 August 2024). [CrossRef]
- Bauchau, V. Do parasitic mites decrease growth of nestling pied flycatchers Ficedula hypoleuca? Ardea-Wageningen 1997, 85, 243–248. [Google Scholar]
- Carleton, R.E. Ectoparasites affect hemoglobin and percentages of immature erythrocytes but not hematocrit in nestling Eastern Bluebirds. Wilson J. Ornithol. 2008, 120, 565–568. [Google Scholar] [CrossRef]
- Moller, A.P. Effects of parasitism by a haematophagous mite on reproduction in the barn swallow. Ecology 1990, 71, 2345–2357. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).