
“Draculamya” uraniae: A New Small-Sized Bivalve from the Mediterranean Sea (Galeommatida, Lasaeidae)


Abstract
:1. Introduction
2. Results
2.1. Systematics
- Class Bivalvia Linnaeus, 1758
- Subclass Autobranchia Groblen 1894
- Order Galeommatida Lemer, Bieler and Giribet, 2019
- Superfamily Galeommatoidea Gray, 1840
- Family Lasaeidae Gray, 1842
- Genus Draculamya Oliver and Lützen, 2011
- “Draculamya” uraniae new species (Figure 2)
2.2. Material Examined
2.2.1. “Draculamya” uraniae n. sp.
2.2.2. “Draculamya uraniae” n. sp. Additional Material
2.2.3. Other Material
2.3. Etymology
2.4. Description (Holotype Measurements in Parentheses)
2.5. Comparative Remarks
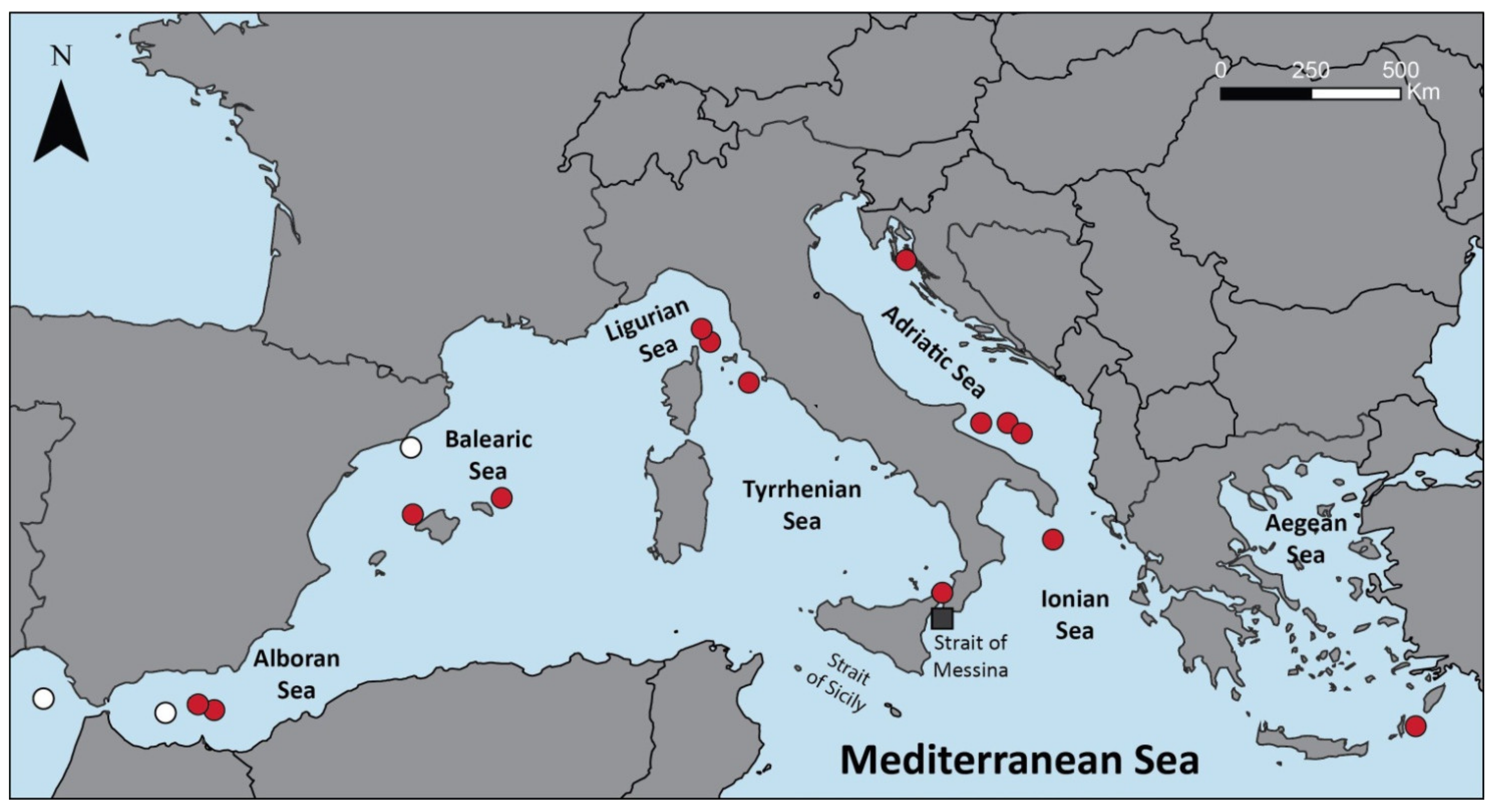
3. Distribution and Habitat
3.1. Geographical Distribution
3.2. Ecology
4. Discussion
4.1. Generic Placement
4.2. Draculamya vs. Glibertia
5. Material and Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations and Acronyms
References
- Li, J.; O’ Foighil, D.; Middelfart, P. The evolutionary ecology of biotic association in a megadiverse bivalve superfamily: Sponsorship required for permanent residency in sediment. PLoS ONE 2012, 7, e42121. [Google Scholar] [CrossRef]
- Chavan, A. Superfamily Leptonacea Gray, 1847. In Treatise on Invertebrate Zoology. Part N., Mollusca 6, Bivalvia 2; Moore, R.C., Ed.; University of Kansas and Geological Society of America: Lawrence, KS, USA, 1969; pp. 518–537. [Google Scholar]
- Boss, K.J. Mollusca. In Synopsis and Classification of Living Organisms; Parker, S.P., Ed.; McGraw Hill Book Company: New York City, NY, USA, 1982; Volume 1, pp. 945–1166. [Google Scholar]
- Purchon, R.D. Classification and evolution of the Bivalvia: An analytical study. Philos. Trans. R. Soc. B 1987, 316, 277–302. [Google Scholar] [CrossRef]
- Bieler, R.; Mikkelsen, P.M. Bivalvia—A look at the Branches. Zool. J. Linn. Soc. 2006, 148, 223–235. [Google Scholar] [CrossRef]
- Bieler, R.; Carter, J.G.; Coan, E.V.; Bouchet, P.; Rocroi, J.-P. Nomenclator of Bivalve Families with a Classification of Bivalve families. Malacologia 2010, 52, 1–184. [Google Scholar] [CrossRef]
- Carter, J.G.; Altaba, C.R.; Anderson, L.C.; Araujo, R.; Biakov, A.S.; Bogan, A.E.; Campbell, D.C.; Campbell, M.; Chen, J.-H.; Cope, J.C.W.; et al. A synoptical classification of the Bivalvia (Mollusca). Paleontol. Contrib. 2011, 4, 1–47. [Google Scholar]
- Goto, R.; Kawakita, A.; Ishikawa, H.; Hamamura, Y.; Kato, M. Molecular phylogeny of the bivalve superfamily Galeommatoidea (Heterodonta, Veneroida) reveals dynamic evolution of symbiotic lifestyle and interphylum host switching. BMC Evol. Biol. 2012, 12, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bieler, R.; Mikkelsen, P.M.; Collins, T.M.; Glover, E.A.; González, V.L.; Graf, D.L.; Harper, E.M.; Healy, J.; Kawauchi, G.Y.; Sharma, P.P.; et al. Investigating the Bivalve Tree of Life—An exemplar–based approach combining molecular and novel morphological characters. Invertebr. Syst. 2014, 28, 32–115. [Google Scholar] [CrossRef] [Green Version]
- Lemer, S.; Bieler, R.; Giribet, G. Resolving the relationships of clams and cockles: Dense transcriptome sampling drastically improves the bivalve tree of life. Proc. R. Soc. B. 2019, 286, 20182684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gofas, S.; Le Renard, J.; Bouchet, P. Mollusca. In European Register of Marine Species: A Check–List of the Marine Species in Europe and a Bibliography of Guides to Their Identification; Collection Patrimoines Naturels, 50; Costello, M.J., Emblow, C., White, R.J., Eds.; Muséum National d’Histoire Naturelle: Paris, France, 2001; pp. 180–213. [Google Scholar]
- Galeommatida. European Register of Marine Species. Available online: http://www.marbef.org/data/aphia.php?p=taxdetails&id=1326015 (accessed on 15 April 2021).
- Van Aartsen, J.J. Galeommatacea and Cyamiacea. La Conchiglia 1997, 279, 31–36, 61. [Google Scholar]
- Van Aartsen, J.J. Galeommatacea and Cyamiacea Part II. La Conchiglia 1997, 281, 27–53. [Google Scholar]
- Van Aartsen, J.J.; Carrozza, F. On “Lasea” pumila (S.V. Wood, 1851) and two new bivalves from European waters: Mancikellia divae n. sp. and Kelliopsis jozinae n. sp. (Bivalvia: Condylocardiidae and Montacutidae). La Conchiglia 1997, 285, 28–34. [Google Scholar]
- Giribet, G.; Peñas, A. A new Epilepton species from the Western Mediterranean. Iberus 1999, 16, 117–121. [Google Scholar]
- Gofas, S. Systematics of Planktomya, a bivalve genus with teleplanic larval dispersal. Bull. Mar. Sci. 2000, 67, 1013–1023. [Google Scholar]
- Holmes, A.M.; Gallichan, J.; Wood, H. Coracuta obliquata n. gen. (Chaster, 1897) (Bivalvia: Montacutidae)—First British record for 100 years. J. Conchol. 2006, 39, 151–158. [Google Scholar]
- Bogi, C.; van Aartsen, J.J. Posizione sistematica e distribuzione in Mar Mediterraneo di Montacuta semirubra Gaglini, 1992 e Montacuta cuneata Gaglini, 1992 (Bivalvia: Galeommatoidea). Boll. Malacol. 2007, 42, 112–114. [Google Scholar]
- Jespersen, Å.; Lützen, J.; Oliver, P.G. Morphology, biology and systematic position of Epilepton clarkiae (Clark, 1852) (Galeommatoidea: Montacutidae) a bivalve commensal with sipunculans. J. Conchol. 2007, 39, 391–402. [Google Scholar]
- Gofas, S.; Salas, C. A review of European “Mysella” species (Bivalvia: Montacutidae) with description of Kurtiella new genus. J. Molluscan Stud. 2008, 74, 119–135. [Google Scholar] [CrossRef]
- Hoeksema, D.F.; Simons, G.F. Bornia canariensis, a new marine species from the Canary Islands (Bivalvia, Heterodonta, Veneroida, Kelliidae). Basteria 2011, 75, 31–34. [Google Scholar]
- Gofas, S. A new species of Bornia (Bivalvia: Galeommatoidea) from southern Spain. Iberus 2012, 30, 41–48. [Google Scholar]
- Oliver, P.G. Taxonomy of some Galeommatoidea (Mollusca, Bivalvia) associated with deep–sea echinoids: A reassessment of the bivalve genera Axinodon Verrill & Bush, 1898 and Kelliola Dall, 1899 with descriptions of new genera Syssitomya gen. nov. and Ptilomyax gen. nov. Eur. J. Taxon. 2012, 12, 1–24. [Google Scholar] [CrossRef]
- Oliver, P.G.; Lützen, J. An anatomically bizarre, fluid–feeding, galeommatoidean bivalve: Draculamya porobranchiata gen. et sp. nov. (Mollusca: Bivalvia). J. Conchol. 2011, 40, 365–392. [Google Scholar]
- Sabelli, B.; Taviani, M. The making of the Mediterranean molluscan biodiversity. In The Mediterranean Sea; Goffredo, S., Dubinsky, Z., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 285–306. [Google Scholar] [CrossRef]
- MolluscaBase. Available online: http://www.molluscabase.org (accessed on 15 April 2021).
- Giribet, G.; Peñas, A. Fauna malacologica del litoral del Garraf. Iberus 1997, 15, 41–93. [Google Scholar]
- Peñas, A.; Rolán, E.; Luque, Á.A.; Templado, J.; Moreno, D.; Rubio, F.; Salas, C.; Sierra, A.; Gofas, S. Moluscos marinos de la isla de Alborán. Iberus 2006, 24, 25–151. [Google Scholar]
- Utrilla, O.; Gofas, S.; Urra, J.; Marina, P.; Mateo-Ramírez, A.; López-González, N.; González-García, E.; Salas, C.; Rueda, J.L. Molluscs from benthic habitats of Gazul mud volcano (Gulf of Cádiz). Sci. Mar. 2020, 84, 273–295. [Google Scholar] [CrossRef]
- Micali, P.; (Fano, Italy). Personal communication, 2021.
- Albano, P.G.; Azzarone, M.; Amati, B.; Bogi, C.; Sabelli, B.; Rilov, G. Low diversity or poorly explored? Mesophotic molluscs highlight undersampling in the Eastern Mediterranean. Biodivers. Conserv. 2020, 29, 4059–4072. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, L.; Freiwald, A. A unique and diverse amalgamated mollusk assemblage from the Coral Patch Seamount, eastern Atlantic. Misc. Malacolog. 2017, 7, 61–79. [Google Scholar]
- Hoffman, L.; (Senckenberg am Meer, Wilhelmshaven, Germany). Personal communication, 2021.
- Baldwin, C.; Tornabene, L.; Robertson, D.R. Below the mesophotic. Sci. Rep. 2018, 8, 4920. [Google Scholar] [CrossRef] [PubMed]
- Cerrano, C.; Bastari, A.; Calcinai, B.; Di Camillo, C.; Pica, D.; Puce, S.; Valisano, L.; Torsani, F. Temperate mesophotic ecosystems: Gaps and perspectives of an emerging conservation challenge for the Mediterranean Sea. Eur. Zool. J. 2019, 86, 370–388. [Google Scholar] [CrossRef] [Green Version]
- Taviani, M. Associazioni a Molluschi pleistoceniche-attuali dragate nell’Adriatico meridionale. Boll. Zool. 1978, 45, 297–306. [Google Scholar] [CrossRef] [Green Version]
- Bonfitto, A.; Oliverio, M.; Sabelli, B.; Taviani, M.A. Quaternary deep-sea marine molluscan assemblage from East Sardinia (Western Tyrrhenian Sea). Boll. Malacol. 1994, 11, 305–309. [Google Scholar]
- Di Geronimo, I.; Bellagamba, M. Malacofaune dei dragaggi BS 77–1 e BS 77–2 (Sardegna nord orientale). Boll. Soc. Paleontol. Ital. 1985, 24, 111–129. [Google Scholar]
- Van der Meulen, J. Glibertia prosperi n. gen., n. sp. (Fam. Condylocardiidae), from Pliocene deposits. Basteria 1951, 15, 49–53. [Google Scholar]
- Bernard, F. Etudes comparatives sur la coquille des lamellibranches Condylocardia. J. Conchyliol. 1897, 44, 169–207. [Google Scholar]
- Middelfart, P. A revision of the Australian Condylocardiinae. Molluscan Res. 2002, 22, 23–85. [Google Scholar] [CrossRef]
- Middelfart, P. Revision of the Australian Cuninae sensu lato (Bivalvia: Carditoidea: Condylocardiidae). Zootaxa 2002, 112, 1–124. [Google Scholar] [CrossRef]
- Janssen, A.W.; van der Slik, L. De fossiele schelpen van de Nederlandse stranden en zeegaten, tweede serie, 5. Basteria 1972, 36, 171–180. [Google Scholar]
- Janssen, A.W. Systematische lijst van Nederlandse recente en fossiele mollusken. Meded. Werkgr. Tert. Kwart. Geol. 1975, 12, 115–170. [Google Scholar]
- Marquet, R. The molluscan fauna of the Kruisschans Member (Lilli Formation, Late Pliocene) in the Antwerp area (Belgium). Meded. Werkgr. Tert. Kwart. Geol. 1993, 30, 83–103. [Google Scholar]
- Marquet, R. Ecology and evolution of Pliocene bivalves from the Antwerp Basin. Bull. Inst. R. Sci. Nat. Belg. Sci. Terre 2004, 74, 205–212. [Google Scholar]
- Marquet, R. The Neogene Bivalvia (Heterodonta and Anomalodesmata) and Scaphopoda from Kallo and Doel (Oost Vlaanderen, Belgium). Palaeontos 2005, 6, 1–142. [Google Scholar]
- Wesselingh, F.P.; Moerdijk, P.W. (Eds.) Fossiele Schelpen van de Nederlandse Kust; NCB Naturalis: Leiden, The Netherlands, 2010; pp. 1–332. [Google Scholar]
- Moerdijk, P.W.; (Middelburg, The Netherlands). Personal communication, 2018.


{kind=link}
{kind=link}
| Latitude | Longitude | Depth (m) | Location | Cruise | Station | Gear | Remarks |
|---|---|---|---|---|---|---|---|
| 36°31.2747′ N Start 36°31.2883′ N′ End | 02°55.5708′ W Start 02°54.5393′ W′ End | 475–408 | Seco de los Olivos | COBAS | 07 | Rock dredge | |
| 36°31.5124′ N Start 36°31.5017′ N′ End | 02°50.8713′ W Start 02°50.4744′ W′ End | 108–98 | Seco de los Olivos | COBAS | 08 | Rock dredge | paratyp. 4–5 |
| 36°30.9787′ N Start 36°30.9659′ N′ End | 2°49.7966′ W Start 02°49.3631′ W′ End | 336–330 | Seco de los Olivos | COBAS | 09 | Rock dredge | |
| 39°52.4709 N Start 39°51.6871′ N′ End | 02°37.6097 E Start 02°38.3251 E′ End | 175–105 | NW Mallorca | COBAS | 86 | Rapido dredge | paratyp. 6–7 |
| 39°51.8380′ N | 02°38.1814′ E | 103 | NW Mallorca | COBAS | 89 | Grab | paratyp. 8–9 |
| 40°00.6453′ N | 04°23.3727′ E | 342 | NE Menorca | COBAS | 102 | Grab | |
| 40°01.3378′ N | 04°22.4614 | 285 | NE Menorca | COBAS | 103 | Grab | paratyp. 15–18 |
| 40°02.9378′ N | 04°22.4172′ E | 380 | NE Menorca | COBAS | 106 | Grab | |
| 40°03.0347′ N Start 40°02.0694′ N′ End | 04°21.8537′ E Start 04°22.5835′ E′ End | 461–309 | NE Menorca | COBAS | 108 | Rock dredge | |
| 40°01.1474′ N Start 40°01.5598′ N′ End | 04°23.3716′ E Start 04°23.1062′ E′ End | 367–340 | NE Menorca | COBAS | 109 | Rock dredge | paratyp. 19–23 |
| 39°57.7525′ N Start 39°57.3255′ N′ End | 04°25.8497′ E Start 04°25.1147′ E′ End | 468–217 | NE Menorca | COBAS | 110 | Rock dredge | |
| 39° 34.9250′ N | 18° 23.3616′ E | 501 | Off S. Maria di Leuca | CORSARO | CR55 | Dredge | |
| 41° 17.6884′ N | 17° 8.3017′ E | 314 | Off Bari | SETE06 | SE06-9 | Grab | |
| 41° 22.6999′ N | 17° 6.7500′ E | 286 | Off Bari | SETE06 | SE13 | Rock dredge | paratyp. 10–11 |
| 41°39.836′ N | 16°53.342′ E | 154 | Off Bari | SASSI08 | SI08-57 | Grab | |
| 41°18.015′ N | 17°16.653′ E | 795 | Off Bari | SASSI08 | SI08-75 | Box corer | |
| 35°24.9100′ N | 27°13.4300′ E | 453 | S Karpathos | GECO | Geco18 | Rock dredge | paratyp. 12–13 |
| 35° 30.8230′ N Start 33° 31.2320′ N′ End | 14° 6.0020′ E Start 14° 5.7100′ E′ End | 530–467 | SW Malta | MARCOS | Marcos 46 | Trawl | |
| 37° 58.0000′ N | 15° 39.0000′ E | 70 | Off Lazzàro, Reggio Calabria | Scuba | holotype | ||
| 43° 29.3884′ N | 9° 49.4550′ E | 180 | Off NW Gorgona Island | Trawl | paratyp. 1 | ||
| 38° 16.0650′ N | 15° 39.4350′ E | 48 | Capo Peloro, Messina | Scuba | paratyp. 2 | ||
| 43° 16.1417′ N | 10° 7.18332′ E | 150 | Off NE Capraia Island | Trawl | paratyp. 3 | ||
| 43° 8.5616′ N | 9° 34.9684′ E | 384 | Off NW Capraia Island | Trawl | paratyp.14 | ||
| 43° 10.2600′ N | 9° 50.9683′ E | 180 | Off Capraia Island | Trawl | |||
| 42° 55.4033′ N | 9° 50.9816′ E | 315 | Off Capraia Island | Trawl | |||
| 42° 58.6800′ N | 9° 36.7350′ E | 350 | Off Capraia Island | Trawl | |||
| 43° 8.0134′ N | 9° 37.71498′ E | 400 | Off Capraia Island | Trawl | |||
| 42° 20.4817′ N | 10° 51.0784′ E | 95 | Giglio Island | Trawl | |||
| 38° 15.4650′ N | 15° 42.6750′ E | 54 | Scilla (Italy) | Scuba | |||
| 44° 46.8533′ N | 14° 39.3517′ E | 45 | Off Rab Island (Croatia) | Scuba |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romani, L.; Bartolini, S.; Oliver, P.G.; Taviani, M. “Draculamya” uraniae: A New Small-Sized Bivalve from the Mediterranean Sea (Galeommatida, Lasaeidae). Taxonomy 2021, 1, 142-151. https://doi.org/10.3390/taxonomy1020011
Romani L, Bartolini S, Oliver PG, Taviani M. “Draculamya” uraniae: A New Small-Sized Bivalve from the Mediterranean Sea (Galeommatida, Lasaeidae). Taxonomy. 2021; 1(2):142-151. https://doi.org/10.3390/taxonomy1020011
Chicago/Turabian StyleRomani, Luigi, Stefano Bartolini, P. Graham Oliver, and Marco Taviani. 2021. "“Draculamya” uraniae: A New Small-Sized Bivalve from the Mediterranean Sea (Galeommatida, Lasaeidae)" Taxonomy 1, no. 2: 142-151. https://doi.org/10.3390/taxonomy1020011