
Seed Morphological Properties Related to Taxonomy in Silene L. Species

 ,
,  , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Seeds Used in This Work
2.2. Seed Images
2.3. General Description of Size and Shape
2.4. Seed Shape Quantification and Testing of the Models: J Index
2.5. Statistical Analysis
3. Results
3.1. General Morphological Analysis
3.1.1. Area
- Dorsal view
- Lateral view
3.1.2. Perimeter
- Dorsal view
- Lateral view
3.1.3. Circularity
- Dorsal view
- Lateral view
3.1.4. Roundness
- Dorsal view
- Lateral view
3.1.5. Aspect Ratio
- Dorsal view
- Lateral view
3.1.6. Solidity
- Dorsal view
- Lateral view
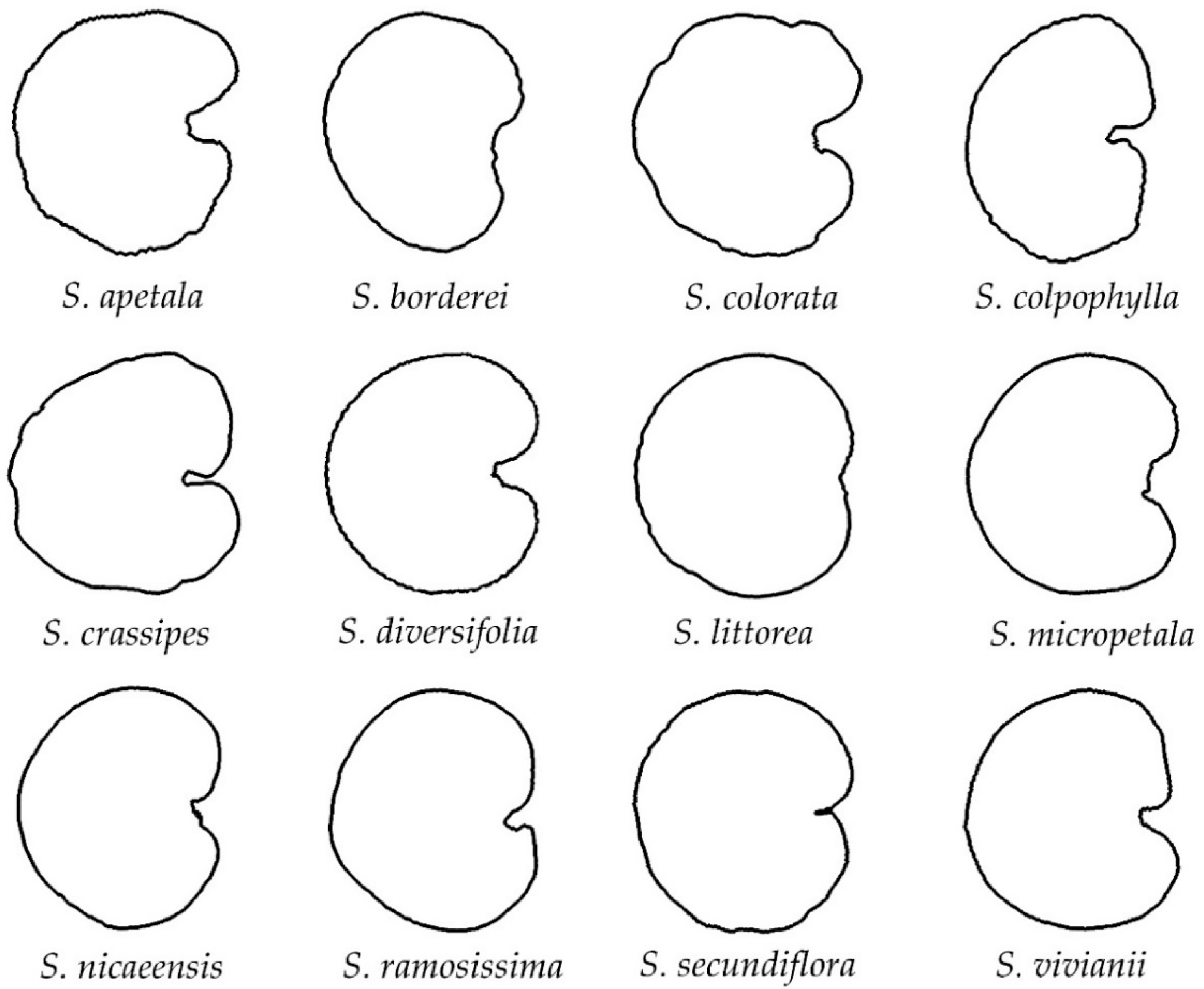
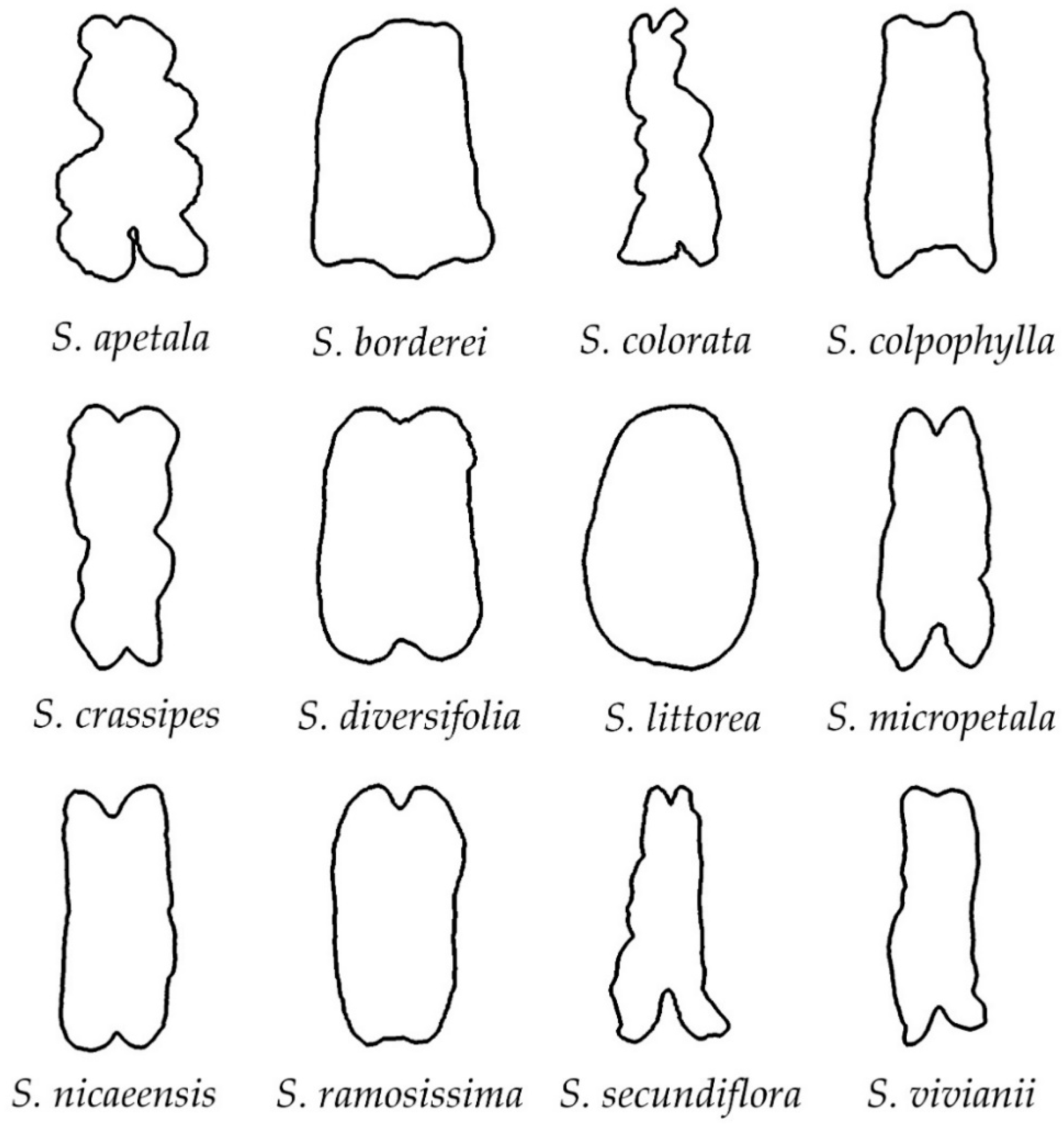
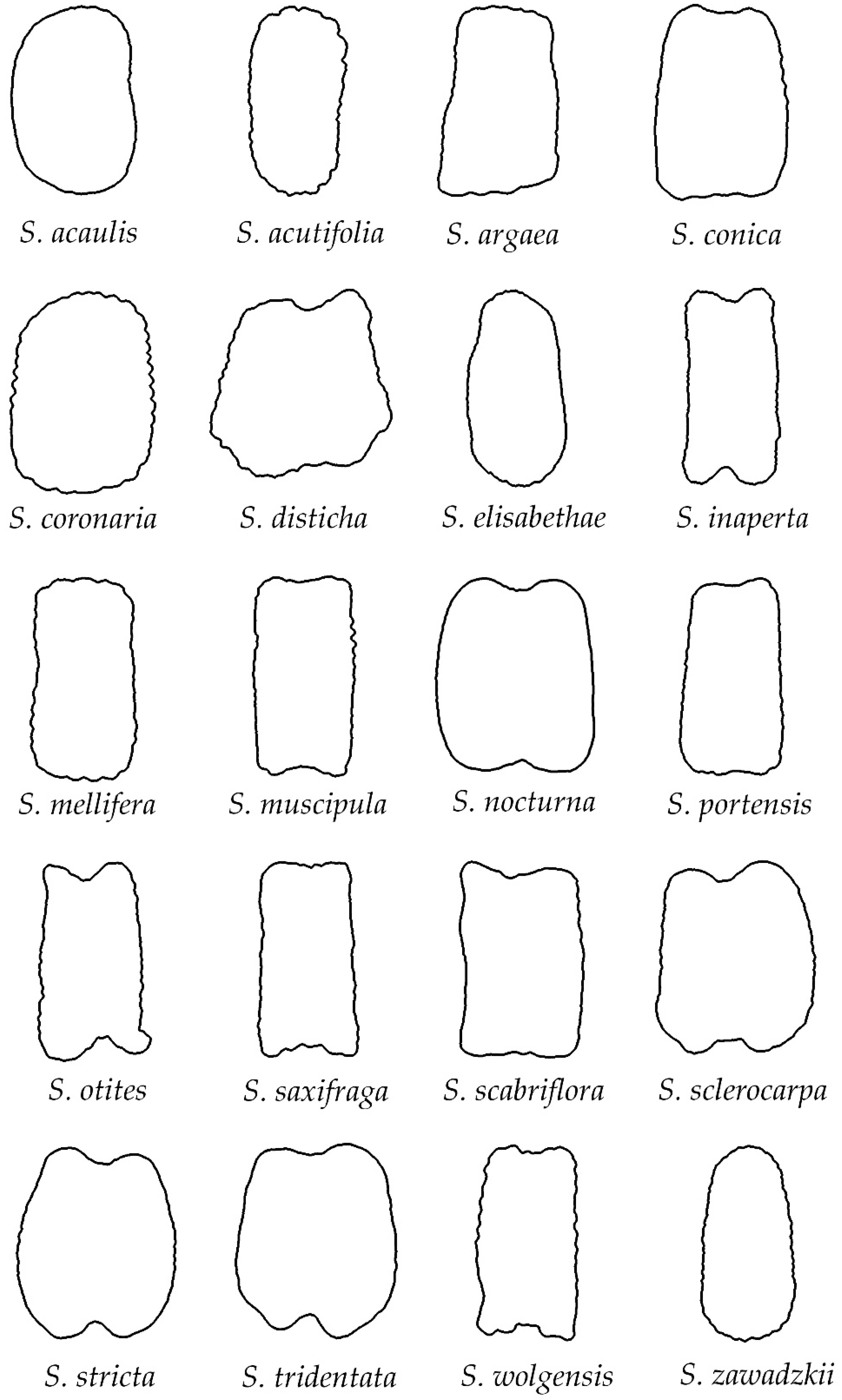
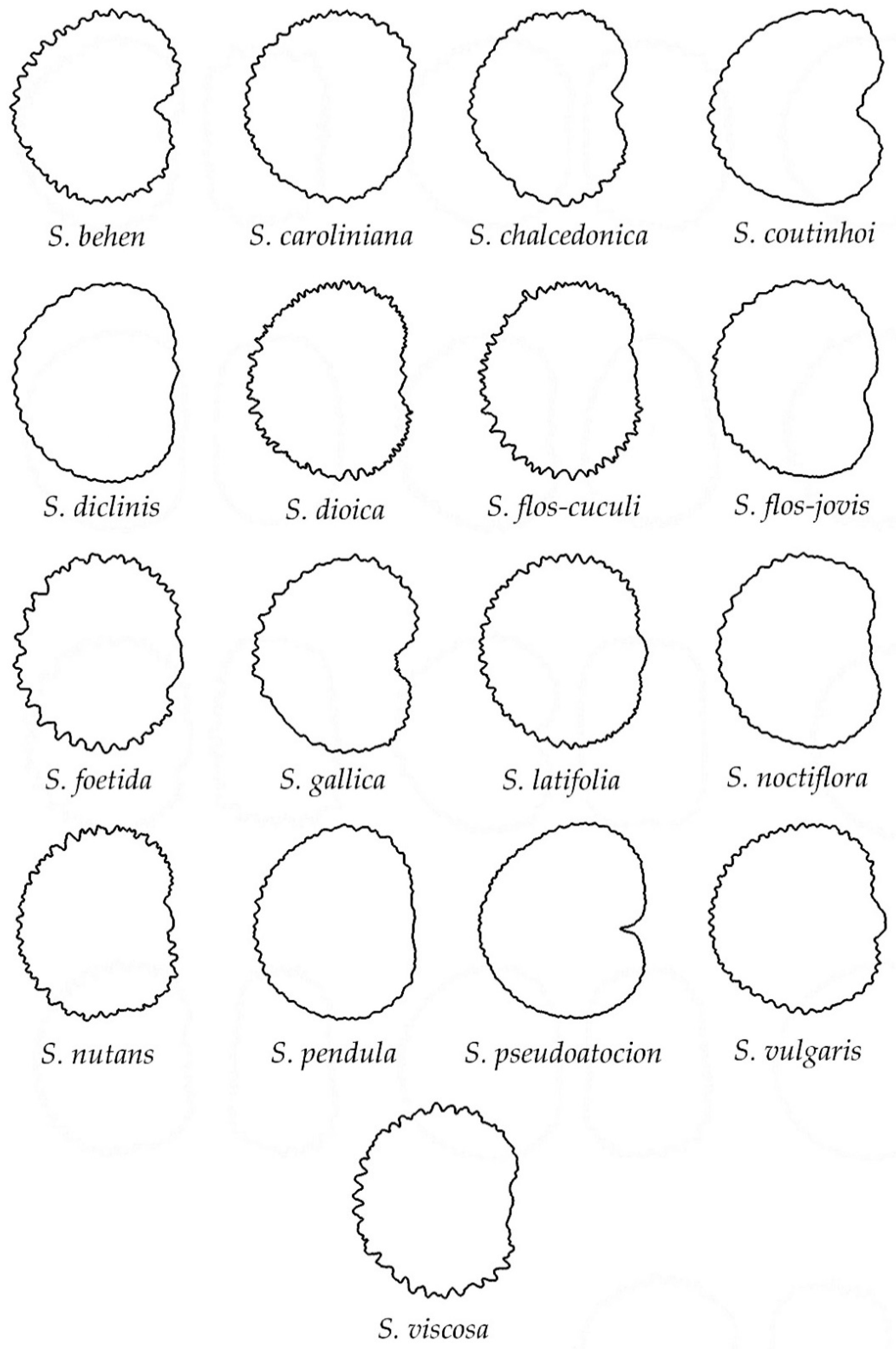
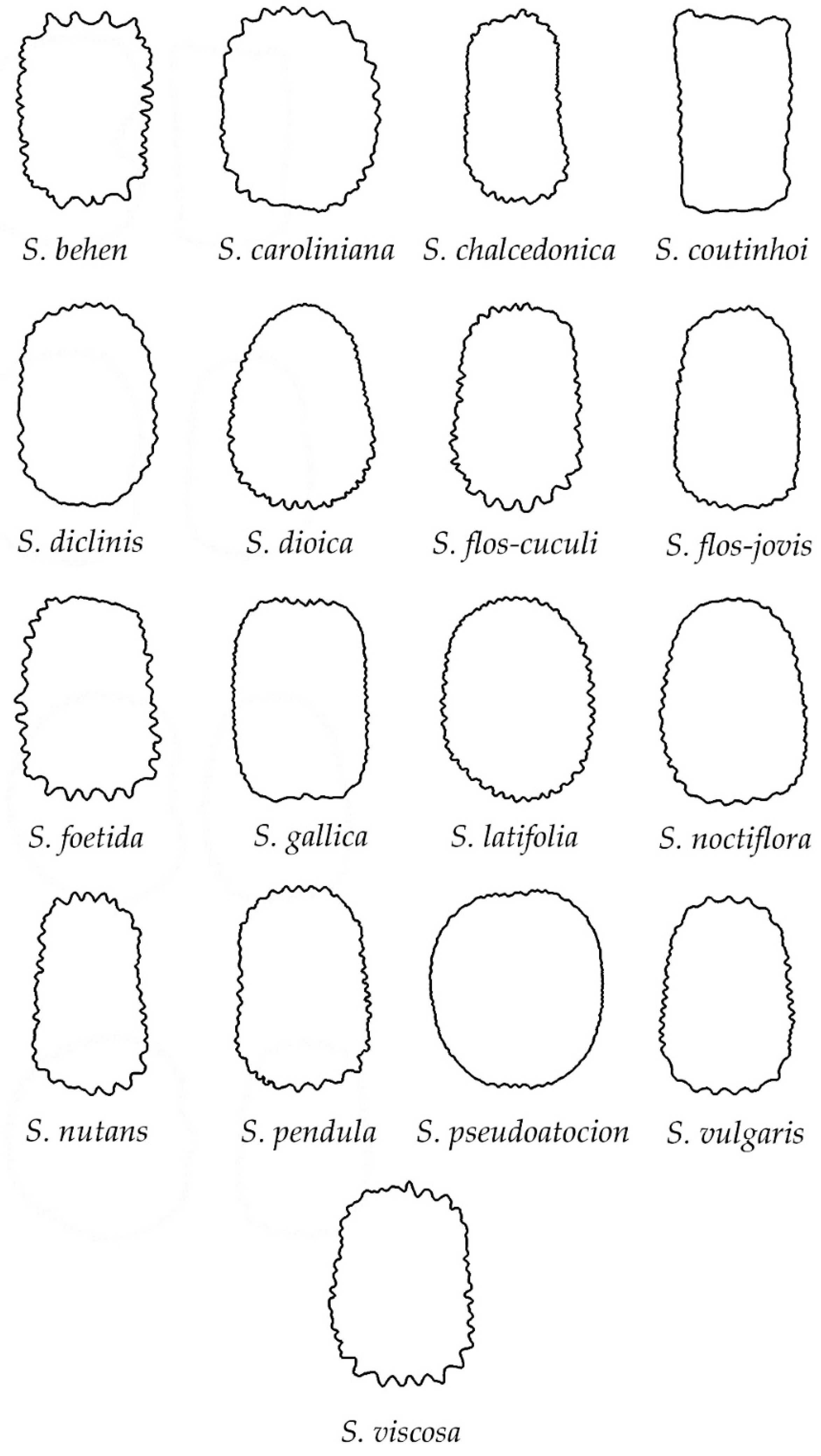
3.2. Classification of Silene Species in Groups Based on Their Seed Silhouette
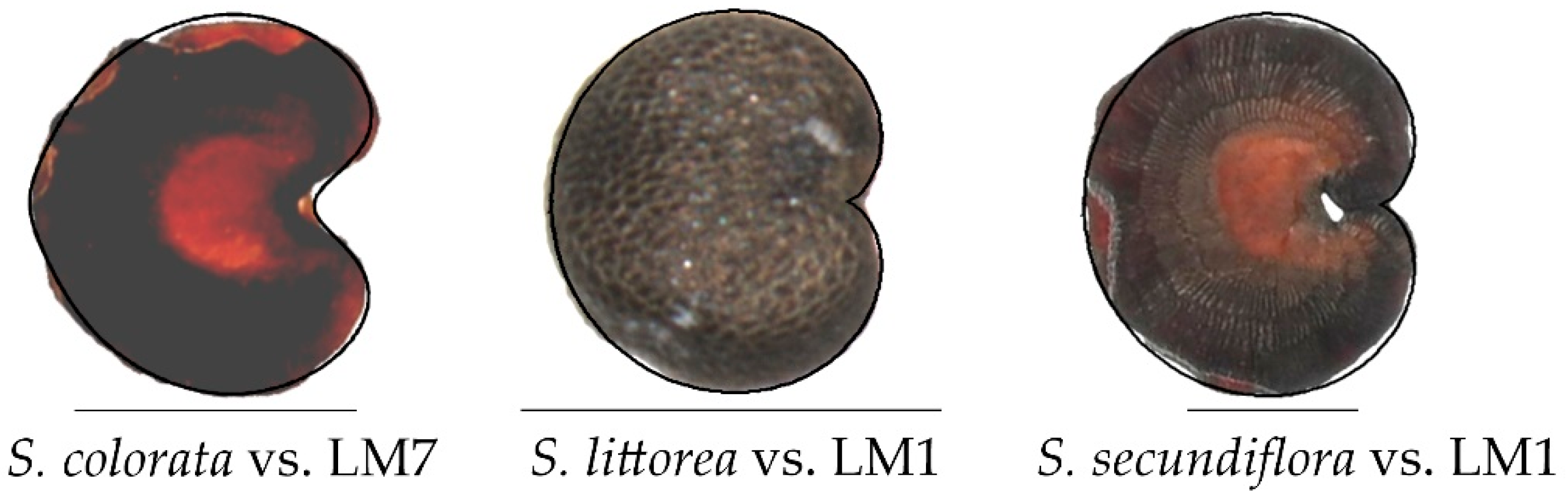
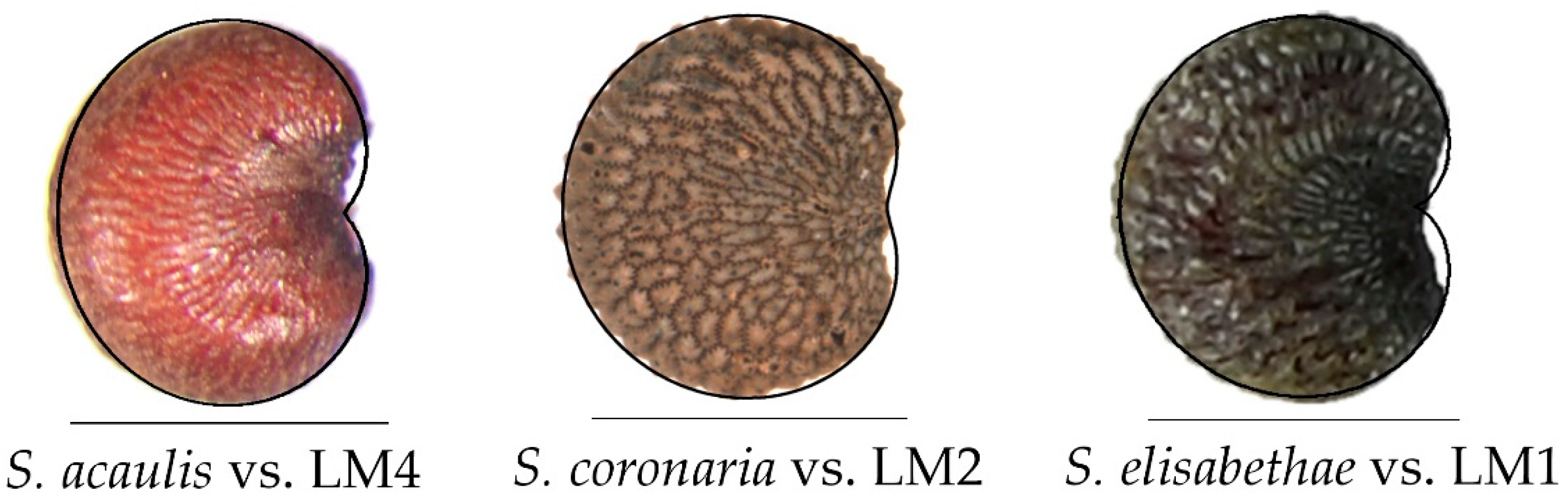
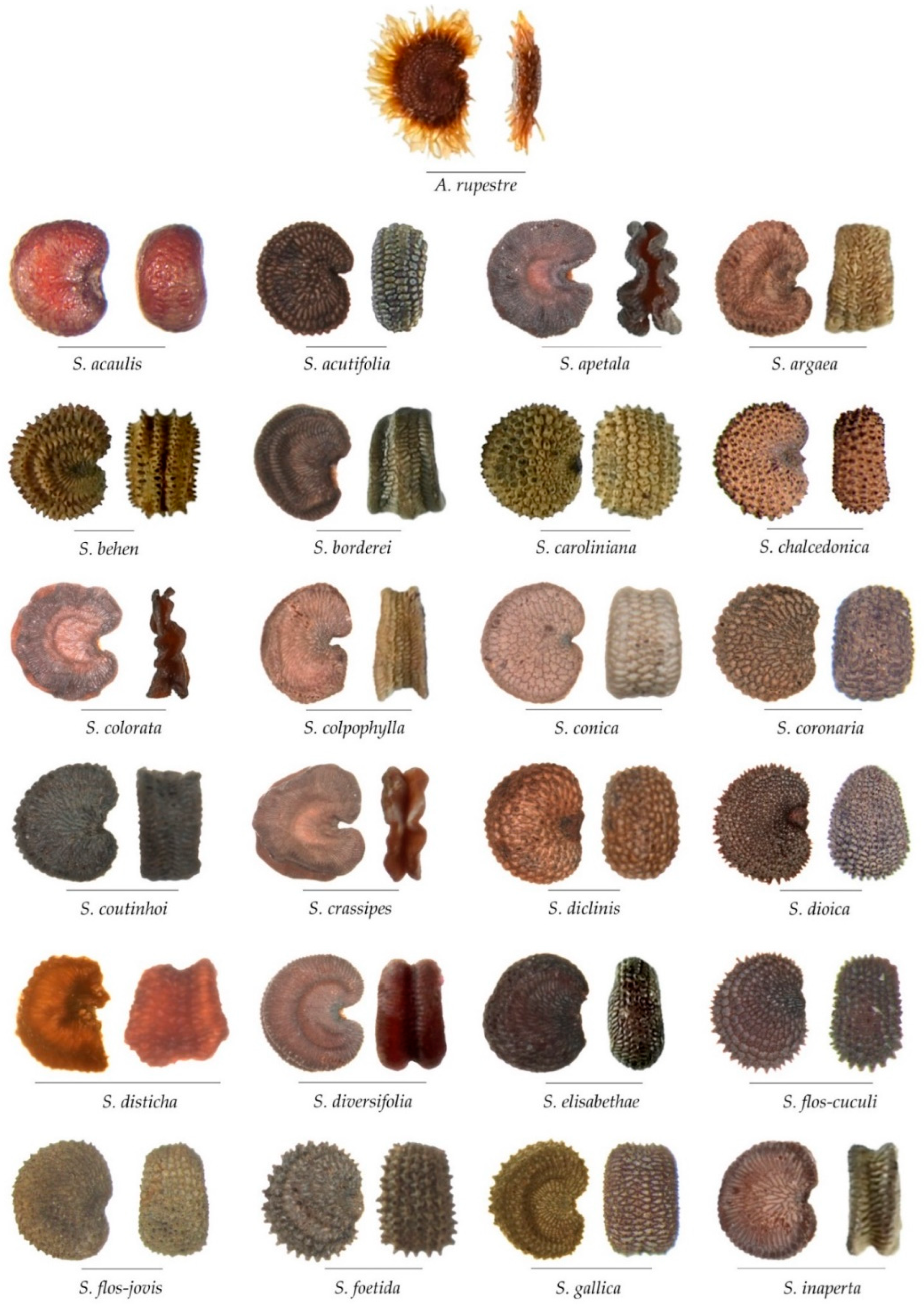
3.3. Morphological Comparison between the Three Groups and Related Genera
3.3.1. Lateral View
3.3.2. Dorsal View
3.4. Analysis of Shape by Geometric Models in Each of the Three Groups of Silene
3.4.1. Smooth Seeds
3.4.2. Rugose Seeds
3.4.3. Echinate Seeds
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A



Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Origin | Annual/Biannual/Perennial | Subgenus and Section |
|---|---|---|---|
| S. acaulis (L.) Jacq. | Botanical Garden of Babeș-Bolyai University. Cluj, Romania | Perennial | S. subg. Silene sect. Siphonomorpha Otth |
| S. acaulis (L.) Jacq | University of Oulu, Finland | Perennial | S. subg. Silene sect. Siphonomorpha Otth |
| S. acutifolia Link ex Rohrb. | Czech Republic | Perennial | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Acutifoliae Oxelman & F.Jafari |
| S. apetala Willd. | Arenal de Petrer, Alicante, Spain | Annual | S. subg. Silene sect. Silene |
| S. apetala Willd. | Caprala, Petrer, Alicante, Spain | Annual | S. subg. Silene sect. Silene |
| S. apetala Willd. | u. (IBP collection, Brno) | Annual | S. subg. Silene sect. Silene |
| S. argaea Fisch. & C.A.Mey. | u. (IBP collection, Brno) | Perennial | S. subg. Silene sect. Auriculatae (Boiss.) Schischk. |
| S. behen L. | u. (IBP collection, Brno) | Annual | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Behenantha Otth |
| S. borderei Jord. | u. (IBP collection, Brno) | Perennial | S. subg. Silene sect. Silene |
| S. caroliniana Walter | u. (IBP collection, Brno) | Perennial | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Physolychnis (Benth.) Bocquet |
| S. chalcedonica (L.) E.H.L.Krause | u. (IBP collection, Brno) | Perennial | S. subg. Lychnis (L.) Greuter sect. Lychnis (L.) Greuter |
| S. colorata Poir. | Arenal de Petrer, Alicante, Spain | Annual | S. subg. Silene sect. Silene |
| S. colorata Poir. | Villena, Alicante, Spain | Annual | S. subg. Silene sect. Silene |
| S. colorata Poir. | u. (IBP collection, Brno) | Annual | S. subg. Silene sect. Silene |
| S. colpophylla Wrigley | France | Perennial | S. subg. Silene sect. Siphonomorpha Otth |
| S. colpophylla Wrigley | u. (IBP collection, Brno) | Perennial | S. subg. Silene sect. Siphonomorpha Otth |
| S. conica L. | Villena Alicante, Spain | Annual | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Conoimorpha Otth |
| S. conica L. | Albarracín, Teruel, Spain | Annual | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Conoimorpha Otth |
| S. conica L. | Berlin | Annual | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Conoimorpha Otth |
| S. conica L. | Berlin | Annual | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Conoimorpha Otth |
| S. coronaria (L.) Clairv. | Rostock Botanical garden | Perennial | S. subg. Lychnis (L.) Greuter sect. Agrostemma (DC.) Greuter |
| S. coronaria (L.) Clairv. | University of Berlin | Perennial | S. subg. Lychnis (L.) Greuter sect. Agrostemma (DC.) Greuter |
| S. coutinhoi Rothm. & P.Silva | Larouco, Ourense, Spain | Perennial | S. subg. Silene sect. Siphonomorpha Otth |
| S. crassipes Fenzl | Hania, Kriti, Greece | Annual | S. subg. Silene sect. Lasiocalycinae (Boiss.) Chowdhuri |
| S. diclinis (Lag.) M.Laínz | u. (IBP collection, Brno) | Perennial | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Melandrium (Röhl.) Rabeler |
| S. diclinis (Lag.) M.Laínz | u. (IBP collection, Brno) | Perennial | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Melandrium (Röhl.) Rabeler |
| S. diclinis (Lag.) M.Laínz | u. (IBP collection, Brno) | Perennial | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Melandrium (Röhl.) Rabeler |
| S. diclinis (Lag.) M.Laínz | Pla de Mora | Perennial | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Melandrium (Röhl.) Rabeler |
| S. dioica (L.) Clairv. | Berlin | Perennial | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Melandrium (Röhl.) Rabeler |
| S. dioica (L.) Clairv. | Rostock Botanical garden | Perennial | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Melandrium (Röhl.) Rabeler |
| S. dioica (L.) Clairv. | Orto Botanico Friulano, Udine Italy | Perennial | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Melandrium (Röhl.) Rabeler |
| S. disticha Willd. | Alcari, Estremadura, Portugal | Annual | S. subg. Silene sect. Silene |
| S. diversifolia Otth | Elda, Alicante, Spain | Annual | S. subg. Silene sect. Silene |
| S. elisabethae Jan | u. (IBP collection, Brno) | Perennial | S. subg. Behenantha (Otth) Torr. & A.Gray unplaced |
| S. flos-cuculi (L.) Greuter & Burdet | Berlin | Perennial | S. subg. Lychnis (L.) Greuter sect. Coccyganthe (Rchb.) Greuter |
| S. flos-jovis (L.) Greuter & Burdet | Berlin | Perennial | S. subg. Lychnis (L.) Greuter sect. Agrostemma (DC.) Greuter |
| S. flos-jovis (L.) Greuter & Burdet | Gogela St Gallen | Perennial | S. subg. Lychnis (L.) Greuter sect. Agrostemma (DC.) Greuter |
| S. foetida Link ex Spreng. | Muiños, Ourense, Spain | Perennial | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Acutifoliae Oxelman & F.Jafari |
| S. gallica L. | Corse, France | Annual | S. subg. Silene sect. Silene |
| S. gallica L. | u. (IBP collection, Brno) | Annual | S. subg. Silene sect. Silene |
| S. gallica L. | St. Gallen, Switzerland | Annual | S. subg. Silene sect. Silene |
| S. inaperta L. | Elda, Alicante, Spain | Annual | S. subg. Silene sect. Muscipula |
| S. inaperta L. | Petrer, Alicante, Spain | Annual | S. subg. Silene sect. Muscipula (Tzvelev) Oxelman, F.Jafari & Gholipour |
| S. inaperta L. | u. (IBP collection, Brno) | Annual | S. subg. Silene sect. Muscipula (Tzvelev) Oxelman, F.Jafari & Gholipour |
| S. latifolia Poir. | Pego Alicante, Spain | Perennial | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Melandrium (Röhl.) Rabeler |
| S. latifolia Poir. | Brno, neighborhood of Bystrc, South Moravia. CZ | Perennial | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Melandrium (Röhl.) Rabeler |
| S. latifolia Poir. | Brno, neighborhood of Bystrc, South Moravia. CZ | Perennial | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Melandrium (Röhl.) Rabeler |
| S. latifolia Poir. | Larzac_Fr_2019 | Perennial | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Melandrium (Röhl.) Rabeler |
| S. latifolia Poir. | Locality of Liblice, district of Mělník, Central Bohemia. CZ | Perennial | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Melandrium (Röhl.) Rabeler |
| S. latifolia Poir. | Internal laboratoy material BFU. Selfcross (17 generations) | Perennial | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Melandrium (Röhl.) Rabeler |
| S. littorea Brot. | Melides, Baixo Alentejo, Portugal | Annual | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Psammophilae (Talavera) Greuter |
| S. mellifera Boiss. & Reut. | Calatayud, Zaragoza, Spain | Perennial | S. subg. Silene sect. Siphonomorpha Otth |
| S. micropetala Lag. | Nador, Morocco | Annual | S. subg. Silene sect. Silene |
| S. muscipula L. | Granada, Spain | Annual | S. subg. Silene sect. Muscipula (Tzvelev) Oxelman, F.Jafari & Gholipour |
| S. nicaeensis All. | Benicarló, Castellón, Spain | Biannual/Perennial | S. subg. Silene sect. Silene |
| S. nicaeensis All. | Algarve, Portugal | Biannual/Perennial | S. subg. Silene sect. Silene |
| S. noctiflora L. | Kuřim (CzR) | Annual | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Elisanthe (Fenzl ex Endl.) Ledeb. |
| S. noctiflora L. | St Gallen, Switzerland | Annual | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Elisanthe (Fenzl ex Endl.) Ledeb. |
| S. noctiflora L. | St Gallen, Switzerland, 1986 | Annual | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Elisanthe (Fenzl ex Endl.) Ledeb. |
| S. nocturna L. | L’Abdet Alicante, Spain | Annual | S. subg. Silene sect. Silene |
| S. nocturna L. | Villena, Alicante, Spain | Annual | S. subg. Silene sect. Silene |
| S. nocturna L. | Peñíscola, Castellón, Spain | Annual | S. subg. Silene sect. Silene |
| S. nocturna L. | Forcall, Castellón, Spain | Annual | S. subg. Silene sect. Silene |
| S. nocturna L. | Czech Republic | Annual | S. subg. Silene sect. Silene |
| S. nutans L. | Besançon, France | Perennial | S. subg. Silene sect. Siphonomorpha Otth |
| S. nutans L. | Rostok, Germany. 2003. | Perennial | S. subg. Silene sect. Siphonomorpha Otth |
| S. otites (L.) Wibel | u. (IBP collection, Brno) | Biannual/Perennial | S. subg. Silene sect. Siphonomorpha Otth |
| S. otites (L.) Wibel | u. (IBP collection, Brno) | Biannual/Perennial | S. subg. Silene sect. Siphonomorpha Otth |
| S. otites (L.) Wibel | Bot.Al.Borza_Romania_2000 | Biannual/Perennial | S. subg. Silene sect. Siphonomorpha Otth |
| S. pendula L. | Czech Republic | Annual | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Behenantha Otth |
| S. portensis L. | Gea, Teruel, Spain | Annual | S. subg. Silene sect. Portenses F.Jafari & Oxelman |
| S. pseudoatocion Desf. | Torretes, Ibi, Alicante | Annual | S. subg. Silene sect. Silene |
| S. ramosissima Desf. | Oliva, Valencia, Spain | Annual | S. subg. Silene sect. Siphonomorpha Otth |
| S. saxifraga L. | u. (IBP collection, Brno) | Perennial | S. subg. Silene sect. Siphonomorpha Otth |
| S. scabriflora Brot. | Cavado, Minho, Portugal | Annual | S. subg. Silene sect. Silene |
| S. sclerocarpa Dufour | Villena, Alicante, Spain | Annual | S. subg. Silene sect. Silene |
| S. sclerocarpa Dufour | Peñíscola, Castellón, Spain | Annual | S. subg. Silene sect. Silene |
| S. sclerocarpa Dufour | Nador, Morocco | Annual | S. subg. Silene sect. Silene |
| S. secundiflora Otth | Pego Alicante, Spain | Annual | S. subg. Silene sect. Silene |
| S. stricta L. | Barbate, Cádiz, Spain | Annual | S. subg. Silene sect. Muscipula (Tzvelev) Oxelman, F.Jafari & Gholipour |
| S. tridentata Desf. | Villena, Alicante, Spain | Annual | S. subg. Silene sect. Silene |
| S. tridentata Desf. | Peñíscola, Castellón, Spain | Annual | S. subg. Silene sect. Silene |
| S. viscosa (L.) Pers. | u. (IBP collection, Brno) | Biannual/Perennial | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Physolychnis (Benth.) Bocquet |
| S. viscosa (L.) Pers | u. (IBP collection, Brno) | Biannual/Perennial | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Physolychnis (Benth.) Bocquet |
| S. vivianii Steud. | Saidia, Morocco | Annual | S. subg. Silene sect. Silene |
| S. vulgaris (Moench) Garcke | Elda, Alicante, Spain | Perennial | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Behenantha Otth |
| S. vulgaris (Moench) Garcke | Forcall, Castellón, Spain | Perennial | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Behenantha Otth |
| S. vulgaris (Moench) Garcke | CzR | Perennial | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Behenantha Otth |
| S. wolgensis (Hornem.) Otth | Bashkortostan, Rusia. | Biannual/Perennial | S. subg. Silene sect. Siphonomorpha Otth |
| S. zawadzkii Herbich | Berlin, Germany | Perennial | S. subg. Behenantha (Otth) Torr. & A.Gray sect. Physolychnis (Benth.) Bocquet |
| A. rupestre (L.) Oxelman | u. (IBP collection, Brno) | Perennial | - |
| V. alpina (L.) G.Don | Linz | Perennial | - |
| V. vulgaris Röhl. | u. (IBP collection, Brno) | Perennial | - |
| V. vulgaris Röhl. | Klembów PL 1951r. OBUW | Perennial | - |
| V. vulgaris Röhl. | Berlin | Perennial | - |
References
- Boissier, E. Flora Orientalis; Georg H.: Basel, Switzerland; Geneva, Switzerland, 1867; Volume 1, pp. 567–656. [Google Scholar]
- Rohrbach, P. Monographic der Gattung Silene; Verlag von Engelmann: Leipzig, Germany, 1869; pp. 1–249. [Google Scholar]
- Jafari, F.; Zarre, S.; Gholipour, A.; Eggens, F.; Rabler, R.K.; Oxelman, B. A new taxonomic backbone for the infrageneric classification of the species-rich genus Silene (Caryophyllaceae). Taxon 2020, 69, 337–368. [Google Scholar] [CrossRef]
- Hoseini, E.; Assadi, M.; Edalatiyan, M.N.; Ghahremaninejad, F. Seed micromorphology and its implication in subgeneric classification of Silene (Caryophyllaceae, Sileneae). Flora 2017, 228, 31–38. [Google Scholar] [CrossRef]
- Dadandi, M.Y.; Yildiz, K. Seed morphology of some Silene L. (Caryophyllaceae) species collected from Turkey. Turk. J. Bot. 2015, 39, 280–297. [Google Scholar] [CrossRef]
- Fawzi, N.M.; Fawzy, A.M.; Mohamed, A.A.A. Seed Morphological Studies on Some Species of Silene L. (Caryophyllaceae). Int. J. Bot. 2010, 6, 287–292. [Google Scholar] [CrossRef][Green Version]
- Ghazanfar, S.A. Seed characters as diagnostic in the perennial sections of the genus Silene (Family Caryophyllaceae). Pak. J. Bot. 1983, 15, 7–12. [Google Scholar]
- Hosny, A.I.; Zareh, M.M. Taxonomic studies of Silenoideae (Caryophyllaceae) in Egypt. 2. Taxonomic significance of seed characters in genus Silene L. with special reference to the seed coat sculpture. Taeckholmia 1993, 14, 37–44. [Google Scholar]
- Greenberg, A.K.; Donoghue, M.J. Molecular systematics and character evolution in Caryophyllaceae. Taxon 2011, 60, 1637–1652. [Google Scholar] [CrossRef]
- Arman, M.; Gholipour, A. Seed morphology diversity in some Iranian endemic Silene (Caryophyllaceae) species and their taxonomic significance. Acta Biol. Szeged. 2013, 57, 31–37. [Google Scholar]
- Gholipour, A.; Kuhdar, F. Seed morphology diversity in the certain Silene species (Caryophyllaceae) from Iran and its taxonomical significance. Taxon. Biosyst. 2014, 6, 107–118. [Google Scholar]
- Keshavarzi, M.; Mahdavinejad, M.; Sheidai, M.; Gholipour, A. Seed and pollen morphology of some Silene species (Caryophyllaceae) in Iran. Phytol. Balc. 2015, 21, 7–12. [Google Scholar]
- El Oqlah, A.A.; Karim, F.M. Morphological and anatomical studies of seed coat in Silene species (Caryophyllaceae) from Jordan. Arab Gulf J. Sci. Res. 1990, 8, 121–139. [Google Scholar]
- Hong, M.-P.; Han, M.-J.; Kim, K.-J. Systematic Significance of Seed Coat Morphology in Silene L. s. str. (Sileneae-Caryophyllaceae) from Korea. J. Plant Biol. 1999, 42, 146–150. [Google Scholar] [CrossRef]
- Perveen, A. Seed morphology of the genus Silene: Caryophyllaceae from Pakistan and Kashmir. Int. J. Biol. Biotechnol. 2009, 6, 219–227. [Google Scholar]
- Ocaña, M.E.; Juan, R.; Fernández, I.; Pastor, J. Estudio morfológico de semillas de Silene (Caryophyllaceae) del suroeste de España. Lagascalia 2011, 31, 21–45. [Google Scholar]
- Martín-Gómez, J.J.; Porceddu, M.; Bacchetta, G.; Cervantes, E. Seed Morphology in Species from the Silene mollissima Aggregate (Caryophyllaceae) by Comparison with Geometric Models. Plants 2022, 11, 901. [Google Scholar] [CrossRef] [PubMed]
- Brullo, C.; Brullo, S.; Giusso del Galdo, G.; Minissale, P.; Sciandrello, S. Silene peloritana (Caryophyllaceae) a new species from Sicily. Phytotaxa 2014, 172, 256–264. [Google Scholar] [CrossRef][Green Version]
- Yildiz, K.; Cirpici, A. Seed morphological studies of Silene L., from Turkey. Pak. J. Bot. 1998, 30, 173–188. [Google Scholar]
- Martín-Gómez, J.J.; Rewicz, A.; Rodríguez-Lorenzo, J.L.; Janoušek, B.; Cervantes, E. Seed Morphology in Silene Based on Geometric Models. Plants 2020, 9, 1787. [Google Scholar] [CrossRef]
- Juan, A.; Martín-Gómez, J.J.; Rodríguez-Lorenzo, J.L.; Janoušek, B.; Cervantes, E. New techniques for Seed Shape Description in Silene Species. Taxonomy 2022, 2, 1–19. [Google Scholar] [CrossRef]
- Rodríguez-Lorenzo, J.L.; Martín-Gómez, J.J.; Tocino, Á.; Juan, A.; Janoušek, B.; Cervantes, E. New Geometric Models for Shape Quantification of the Dorsal View in Seeds of Silene Species. Plants 2022, 11, 958. [Google Scholar] [CrossRef]
- Zhang, D.; Kamel, M.; Baciu, G. Integrated Image and Graphics Technologies; Kluwer Academic Publishers: Boston, MA, USA, 2006. [Google Scholar]
- Gray, A. Modern Differential Geometry of Curves and Surfaces with Mathematica; CRC Press: Boca Raton, FL, USA, 1998; pp. 163–165. [Google Scholar]
- Greuter, W. Silene (Caryophyllaceae) in Greece: A subgeneric and sectional classification. Taxon 1995, 44, 543–581. [Google Scholar] [CrossRef]
- POWO. Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. 2022. Available online: https://www.plantsoftheworldonline.org/ (accessed on 15 June 2022).
- Rasband, W.S. ImageJ; U.S. National Institutes of Health: Bethesda, MD, USA, 2018. Available online: http://imagej.nih.gov/ij/ (accessed on 15 June 2022).
- Zdilla, M.J.; Hatfield, S.A.; McLean, K.A.; Cyrus, L.M.; Laslo, J.M.; Lambert, H.W. Circularity, solidity, axes of a best fit ellipse, aspect ratio, and roundness of the foramen ovale: A morphometric analysis with neurosurgical considerations. J. Craniofacial Surg. 2016, 27, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Cervantes, E.; Martín-Gómez, J.J.; Saadaoui, E. Updated Methods for Seed Shape Analysis. Scientifica 2016, 2016, 5691825. [Google Scholar] [CrossRef] [PubMed]
- Cervantes, E.; Martín-Gómez, J.J.; Espinosa-Roldán, F.E.; Muñoz-Organero, G.; Tocino, Á.; Cabello-Sáenz de Santamaría, F. Seed morphology in key Spanish grapevine cultivars. Agronomy 2021, 11, 734. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Campbell, G.; Skillings, J.H. Nonparametric Stepwise Multiple Comparison Procedures. J. Am. Stat. Assoc. 1985, 80, 998–1003. [Google Scholar] [CrossRef]
- Sokal, R.R.; Braumann, C.A. Significance Tests for Coefficients of Variation and Variability Profiles. Syst. Zool. 1980, 29, 50. [Google Scholar] [CrossRef]
- Rautenberg, A.; Hathaway, L.; Oxelman, B.; Prentice, H.C. Geographic and phylogenetic patterns in Silene section Melandrium. (Caryophyllaceae) as inferred from chloroplast and nuclear DNA sequences. Mol. Phylogenet. Evol. 2010, 57, 978–991. [Google Scholar] [CrossRef] [PubMed]
- Zluvova, J.; Lengerova, M.; Markova, M.; Hobza, R.; Nicolas, M.; Vyskot, B.; Charlesworth, D.; Negrutiu, I.; Janousek, B. The inter-specific hybrid Silene latifolia-S. viscosa reveals early events of sex chromosome evolution. Evol. Dev. 2005, 7, 327–336. [Google Scholar] [CrossRef]
- Talavera, S. Silene L. In Flora Iberica; Castroviejo, S., Aedo, C., Laínz, M., Muñoz Garmendia, F., Nieto Feliner, G., Paiva, J., Benedí, C., Eds.; Real Jardín Botánico, CSIC: Madrid, Spain, 1997; Volume 8, pp. 149–152. [Google Scholar]
- Morton, J.K. Silene Linnaeus. Flora of North America North of Mexico, Flora of North America Association. 2019. Available online: http://floranorthamerica.org/Silene (accessed on 28 January 2022).
- Chater, A.O.; Walters, S.M. Silene L. In Flora Europaea; Tutin, T.G., Heywood, V.H., Burges, N.A., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1964; Volume 1, pp. 158–181. [Google Scholar]
- Davis, P.H.; Mill, R.R.; Tan, K. (Eds.) Silene L. In Flora of Turkey and the East Aegean Islands; Edinburgh University Press: Edinburgh, UK, 1988; Volume 10, pp. 76–81. [Google Scholar]
- Danin, A.; Fragman-Sapir, O. Flora of Israel Online. 2016. Available online: https://flora.org.il/en/plants/ (accessed on 15 June 2022).
- Frajman, B.; Thollesson, M.; Oxelman, B. Taxonomic revision of Atocion and Viscaria (Sileneae, Caryophyllaceae). Bot. J. Linn. Soc. 2013, 173, 194–210. [Google Scholar] [CrossRef]










| Dorsal View | Lateral View | |
|---|---|---|
| Area | Mean value: 0.71 mm2 (SD = 0.38; CV = 53.7) Min. value: 0.24 mm2 (S. ramosissima) Max. value: 1.81 mm2 (S. pseudoatocion) | Mean value: 0.92 mm2 (SD = 0.50; CV = 53.9) Min. value: 0.25 mm2 (S. disticha) Max. value: 2.94 mm2 (S. secundiflora) |
| Perimeter | Mean value: 3.63 mm (SD = 0.99; CV = 27.3) Min. value: 2.08 mm (S. ramosissima) Max. value:6.49 mm (S. secundiflora) | Mean value: 3.90 mm (SD = 1.09; CV = 27.9) Min. value: 2.13 mm (S. disticha) Max. value:6.75 mm (S. secundiflora) |
| Circularity | Mean value: was 0.65 (SD = 0.11; CV = 17.1) Min. value: 0.39 (S. secundiflora) Max. value: 0.82 (S. littorea) | Mean value: 0.72 (SD = 0.08; CV = 10.9) Min. value: 0.45 (S. behen) Max. value: 0.85 (S. littorea) |
| Roundness | Mean value: 0.63 (SD = 0.14; CV = 21.7) Min. value: 0.30 (S. vivianii) Max. value: 0.89 (S. disticha) | Mean value: 0.82 (SD = 0.06; CV = 6.9) Min. value: 0.73 (S. mellifera) Max. value: 0.91 (S. secundiflora) |
| Aspect ratio | Mean value: 1.68 (SD = 0.44; CV = 26.1) Min. value: 1.12 (S. disticha) Max. value: 3.33 (S. vivianii) | Mean value: 1.22 (SD = 0.09; CV = 7.3) Min. value: 1.11 (S. secundiflora) Max. value: 1.40 (S. mellifera) |
| Solidity | Mean value: 0.939 (SD = 0.04; CV = 4.5) Min. value: 0.822 (S. secundiflora) Max. value: 0.987 (S. littorea) | Mean value: 0.957 (SD = 0.01; CV = 1.4) Min. value: 0.917 (S. behen) Max. value: 0.983 (S. littorea) |
| Morphological Type | Smooth | Rugose | Echinate |
|---|---|---|---|
| S. subg. Silene | S. apetala S. borderei S. colorata S. colpophylla S. crassipes S. diversifolia S. micropetala S. nicaeensis S. ramosissima S. secundiflora S. vivianii | S. acaulis S. argaea S. disticha S. inaperta S. mellifera S. muscipula S. nocturna S. otites S. portensis S. saxifraga S. scabriflora S. sclerocarpa S. stricta S. tridentata S. wolgensis | S. coutinhoi S. gallica S. nutans S. pseudoatocion |
| S. subg. Behenantha | S. littorea | S. acutifolia S. conica S. elisabethae S. zawadzkii | S. behen S. caroliniana S. diclinis S. dioica S. foetida S. latifolia S. noctiflora S. pendula S. viscosa S. vulgaris |
| S. subg. Lychnis | S. coronaria | S. chalcedonica S. flos-cuculi S. flos-jovis |
| Seed Type or Species | N | A | P | AR | C | R | S |
|---|---|---|---|---|---|---|---|
| Smooth | 361 | 1.08 d (52.1) | 4.14 c (23.0) | 1.18 a (7.2) | 0.75 d (6.4) | 0.85 d (6.7) | 0.958 c (1.2) |
| Rugose | 774 | 0.63 c (58.1) | 3.15 b (27.7) | 1.24 d (7.4) | 0.76 e (7.1) | 0.81 b (7.0) | 0.957 c (1.3) |
| Echinate | 770 | 1.14 e (37.8) | 4.56 d (18.7) | 1.23 c (6.9) | 0.67 c (12.3) | 0.82 c (6.3) | 0.955 c (1.5) |
| A. rupestre (S. alpestris) | 20 | 1.03 de (9.2) | 8.51 e (17.1) | 1.31 e (7.0) | 0.19 a (32.6) | 0.77 a (7.0) | 0.785 a (5.6) |
| V. alpina (S. suecica) | 20 | 0.33 b (10.6) | 2.31 a (5.9) | 1.23 cd (3.9) | 0.79 f (2.1) | 0.82 bc (4.0) | 0.970 d (0.8) |
| V. vulgaris (S. viscaria) | 60 | 0.26 a (14.0) | 2.25 a (8.0) | 1.19 b (5.6) | 0.65 b (7.1) | 0.84 d (5.8) | 0.947 b (1.0) |
| Seed Type or Species | N | A | P | AR | C | R | S |
|---|---|---|---|---|---|---|---|
| Smooth | 349 | 0.57 d (49.8) | 3.59 c (28.9) | 2.21 d (24.4) | 0.56 b (22.7) | 0.48 b (22.6) | 0.894 b (6.4) |
| Rugose | 742 | 0.52 c (43.2) | 3.00 b (21.1) | 1.62 b (23.6) | 0.70 d (11.4) | 0.65 d (22.5) | 0.945 d (3.4) |
| Echinate | 743 | 0.97 e (40.7) | 4.28 d (19.2) | 1.50 a (11.6) | 0.64 c (15.6) | 0.67 e (11.1) | 0.955 e (2.4) |
| A. rupestre (S. alpestris) | 15 | 0.51 cd (18.5) | 5.39 e (20.2) | 3.01 e (30.9) | 0.23 a (22.5) | 0.36 a (26.0) | 0.772 a (5.8) |
| V. alpina (S. suecica) | 20 | 0.23 b (16.1) | 2.11 a (9.7) | 2.01 d (8.0) | 0.65 c (5.9) | 0.50 b (7.9) | 0.963 e (1.2) |
| V. vulgaris (S. viscaria) | 60 | 0.19 a (15.9) | 2.02 a (7.6) | 1.80 c (7.3) | 0.58 b (8.7) | 0.56 c (7.3) | 0.931 c (1.9) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martín-Gómez, J.J.; Rodríguez-Lorenzo, J.L.; Juan, A.; Tocino, Á.; Janousek, B.; Cervantes, E. Seed Morphological Properties Related to Taxonomy in Silene L. Species. Taxonomy 2022, 2, 298-323. https://doi.org/10.3390/taxonomy2030024
Martín-Gómez JJ, Rodríguez-Lorenzo JL, Juan A, Tocino Á, Janousek B, Cervantes E. Seed Morphological Properties Related to Taxonomy in Silene L. Species. Taxonomy. 2022; 2(3):298-323. https://doi.org/10.3390/taxonomy2030024
Chicago/Turabian StyleMartín-Gómez, José Javier, José Luis Rodríguez-Lorenzo, Ana Juan, Ángel Tocino, Bohuslav Janousek, and Emilio Cervantes. 2022. "Seed Morphological Properties Related to Taxonomy in Silene L. Species" Taxonomy 2, no. 3: 298-323. https://doi.org/10.3390/taxonomy2030024
APA StyleMartín-Gómez, J. J., Rodríguez-Lorenzo, J. L., Juan, A., Tocino, Á., Janousek, B., & Cervantes, E. (2022). Seed Morphological Properties Related to Taxonomy in Silene L. Species. Taxonomy, 2(3), 298-323. https://doi.org/10.3390/taxonomy2030024