4. Taxonomy
Sub-phylum ANTHOZOA Ehrenberg, 1831
Class OCTOCORALLIA Haeckel, 1866
Order MALACALCYONACEA McFadden, van Ofwegen and Quattrini, 2022
Family PARAMURICEIDAE Bayer, 1956
Genus NEOACIS Matsumoto and van Ofwegen, 2023
(replacement name for PARACIS Kükenthal, 1919)
urn:lsid:zoobank.org:act:0CB34176-16FA-4EC8-A2EF-84F8E0E92180
Acis (pars) Ridley, 1882: 126; Studer, 1887: 58; Wleft and Studer, 1889: LVI, 121; Hickson, 1905: 816; Thomson and Henderson, 1905: 297; Thomson and Henderson, 1906: 77; Kinoshita, 1909: 5–15; Thomson and Simpson, 1909: 232; Nutting 1910: 41; Thomson and Russell, 1909: 154; and Nutting, 1912: 80.
Stenella (pars) Thomson and Henderson, 1906: 36.
Paracis Kükenthal, 1919: 753, 837; 1924: 154; Aurivillius, 1931: 143; Bayer, 1956b: F206
Not Acis Duchassaing and Michelotti, 1860: 19; 1864: 14; Kölliker, 1865: 136; Pourtales, 1867-8: 132; Duchassaing, 1870: 15; and Kükenthal, 1919: 753, 836.
Not Acis; Kinoshita, 1909: 5–15.
Type species. Acis orientalis Ridley, 1882 (Mauritius).
Diagnosis. Colonies richly branched in one plane. Polyps situated all around the end branches, on thicker branches more to one side. Coenenchyme with surface layer of big spindles, with rounded tubercles on the outside. Below these big spindles lie smaller ones. Calyces cylindrical, with thorn scales. Polyps with collaret and points, tentacles with dragon wings. Sclerites red or colourless.
Remarks. Grasshoff (1996) [
15] mentioned at least eight species occur in the Indo-Pacific without mentioning any names. WoRMS [
11] lists no less than 21 species.
The genus Neoacis (Paracis) and the genus Pseudothesea could be the same, but we are following Bayer’s separation of the two. Paracis ijimai and P. miyajimai (Kinoshita, 1909) are here re-described as species of a newly described genus which also includes P. squamata (Nutting, 1910). P. spinifera (Nutting, 1912) is here re-described as a type species of another newly described genus.
We re-examined the type of
Acis orientalis Ridley, 1882 and depict its sclerites as the original description of Ridley [
12] was rather ambiguous.
The sclerites of Neoacis (Paracis) resemble those of Discogorgia Kükenthal, 1919, but Paracis differs in having dragon wing sclerites in the tentacles and spindles in the inner coenenchyme. According to the re-examination of the type of Discogorgia campanulifera (type of the genus Discogorgia) using SEM, Discogorgia sclerites are different from Neoacis. Neoacis has short thornscale sclerites in the polyps, but Discogorgia does not.
Neoacis differs from Atlantic Scleracis, which also has big spindles, in having thornscales.
The Atlantic Acanthacis Deichmann, 1936 also resembles Neoacis in having big spindles, but it has bifurcated thornscales as well.
Some species of Euplexaura Verrill, 1869 can also have big spindles with rounded tubercles on the outer side, but those species are sparsely branched, have no calyces and, therefore, no thornscales, and the tentacles have no dragon wings.
Finally, some species of Thesea also can have spindles with rounded tubercles, but that genus also has no thornscales in the calyces.
Key to the 4 N-Pacific species and type of the genus Neoacis (Acis orientalis).
| 1. Dragon wing tentacle sclerites, polyps on three sides only, thornscales with blunt distal end; | 2 |
| -. Dragon wing tentacle sclerites, polyps on three sides/all sides of the colony, thornscales with pointed distal end. | 3 |
| 2. Thornscales with straight projection; | Neoacis pustulata |
| -. Thornscales with thickened middle. | Neoacis marianaensis sp. nov. |
| 3. Polyps on all sides of the colony, outer side of coenenchymal spindles with few rounded elevations with prickles; | Neoacis bayeri sp. nov. |
| -. Outer side of coenenchymal spindles completely covered by rounded elevations with prickles. | 4 |
| 4. Polyps on all sides of the colony, complex tubercles on point and tentacle sclerites; | Neoacis ofwegeni sp. nov. |
| -. Polyps on three sides of colony, point and tentacle sclerites with simple tubercles. | Neoacis orientalis. |
Neoacis orientalis (Ridley, 1882)
Acis orientalis Ridley, 1882: 126, pl. 5 Figures 1–6 (Mauritius).
Paracis orientalis; Kükenthal, 1924: 156; WoRMS, 2023.
NOT Paracis orientalis; Bayer, 1949: 200, Figures 1e and 5a–e, pl. 1 Figure 4 (Bikini) (USNM 44078).
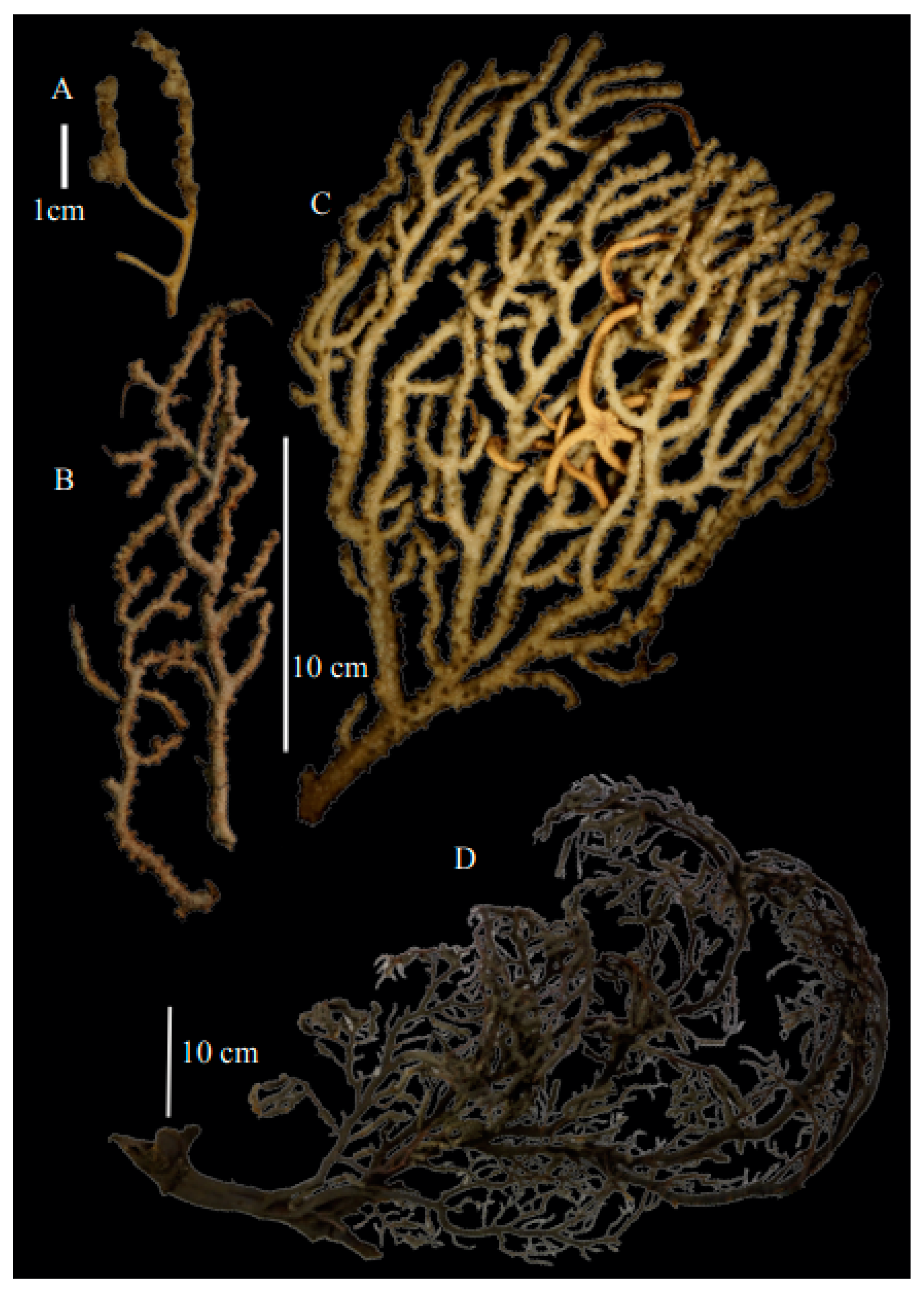
Material examined. Holotype BMNH1950.03.16.11, Mauritius, coll. Prof. S. G. Hickson, dry.
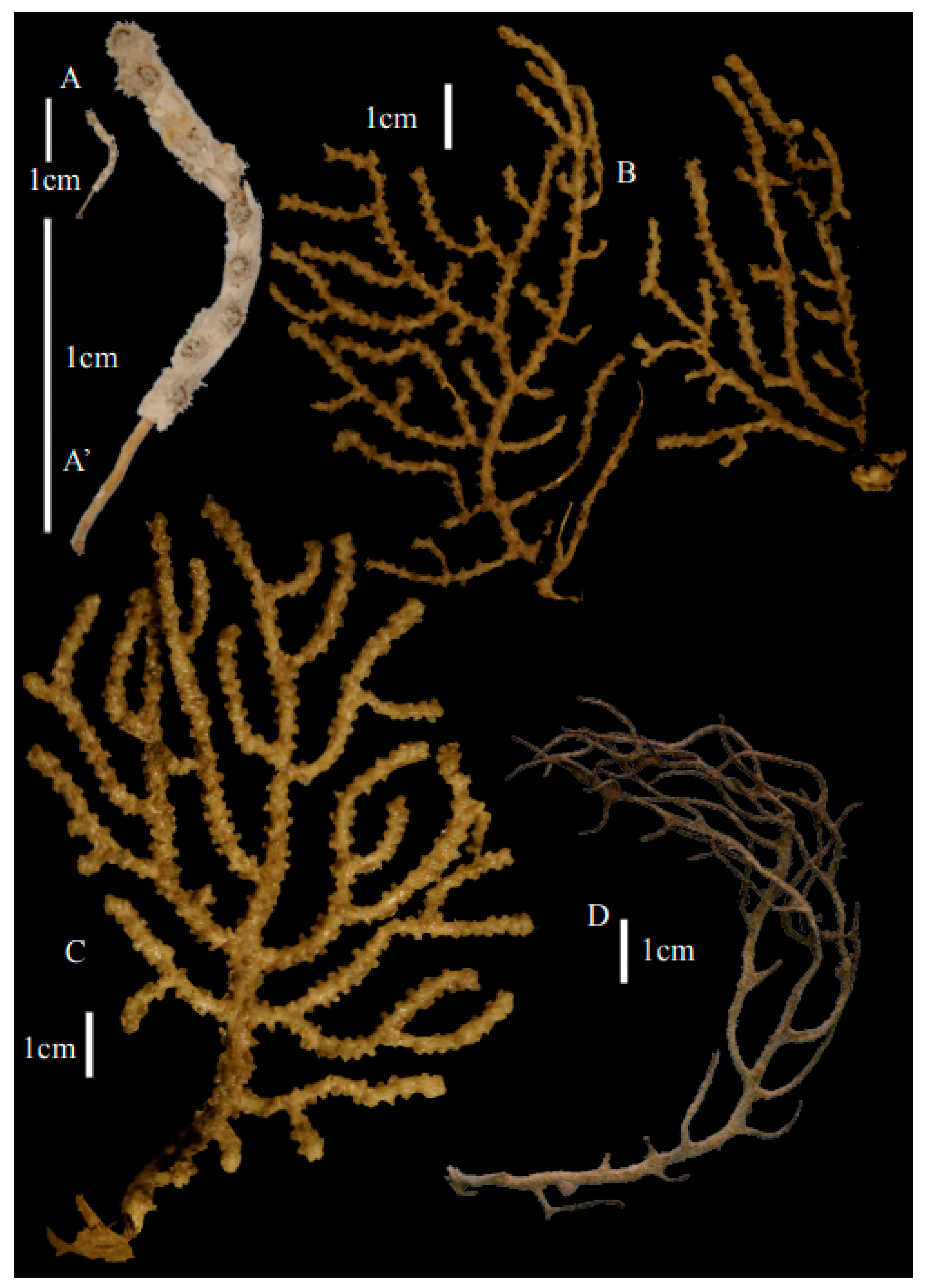
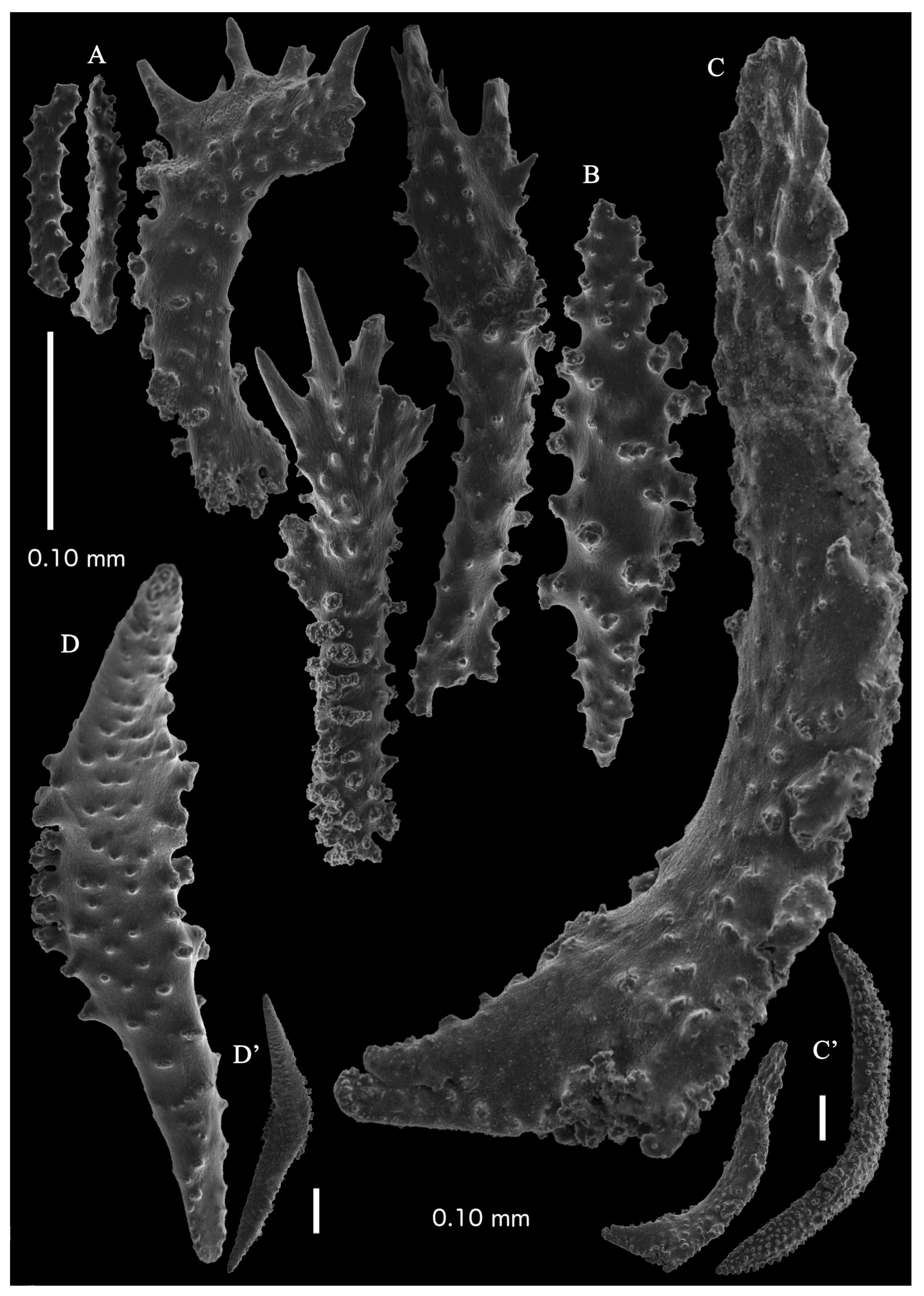
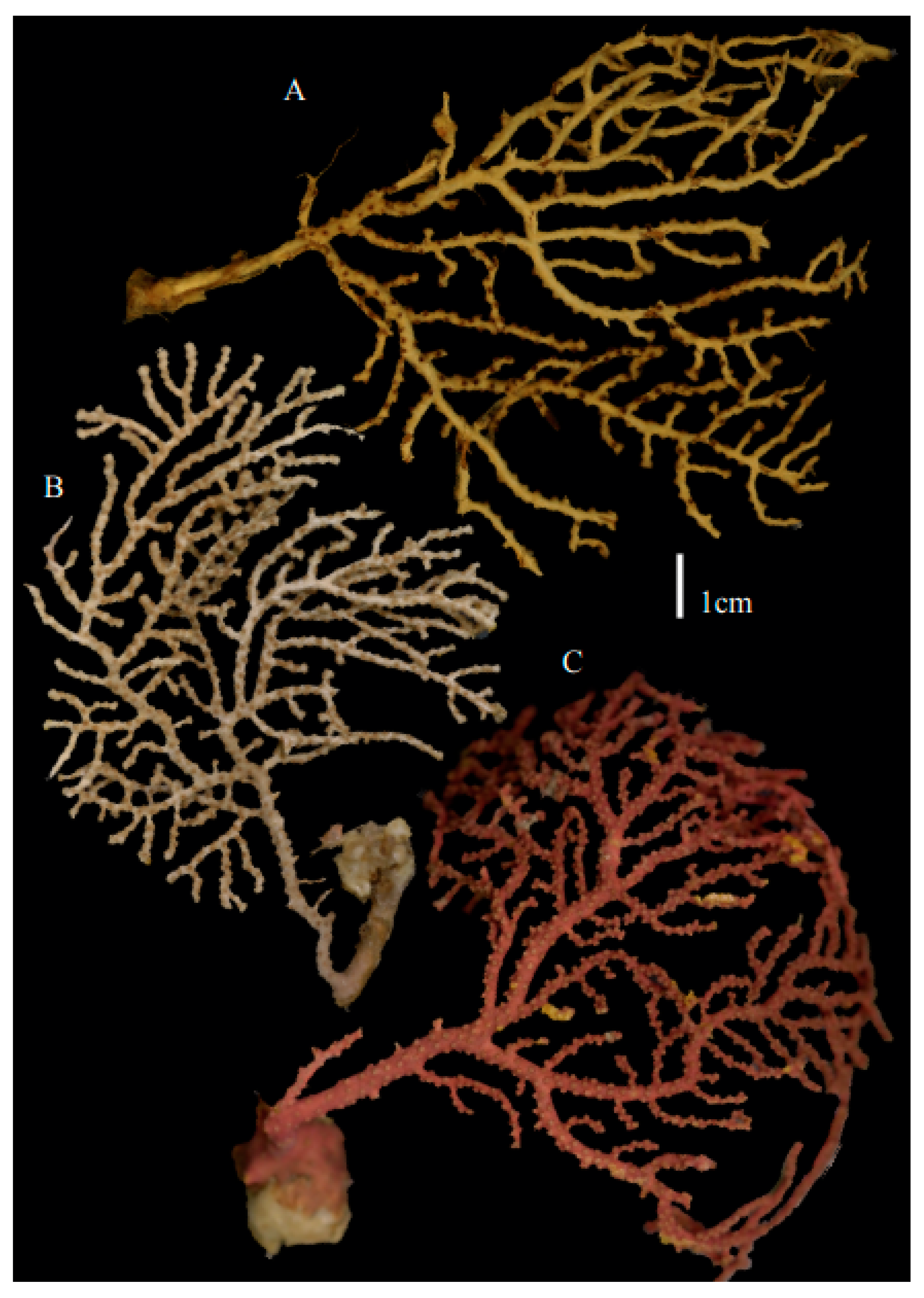
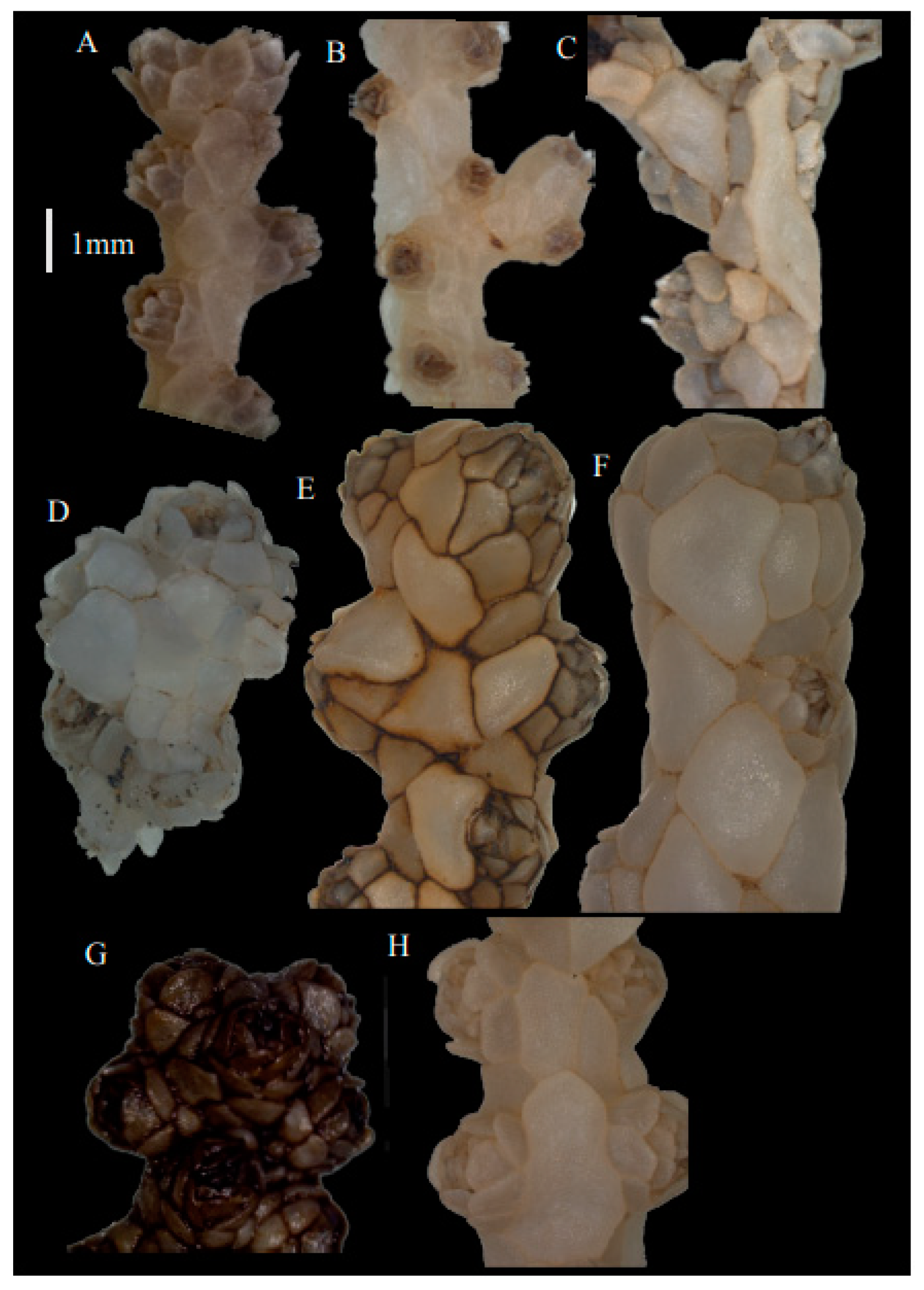
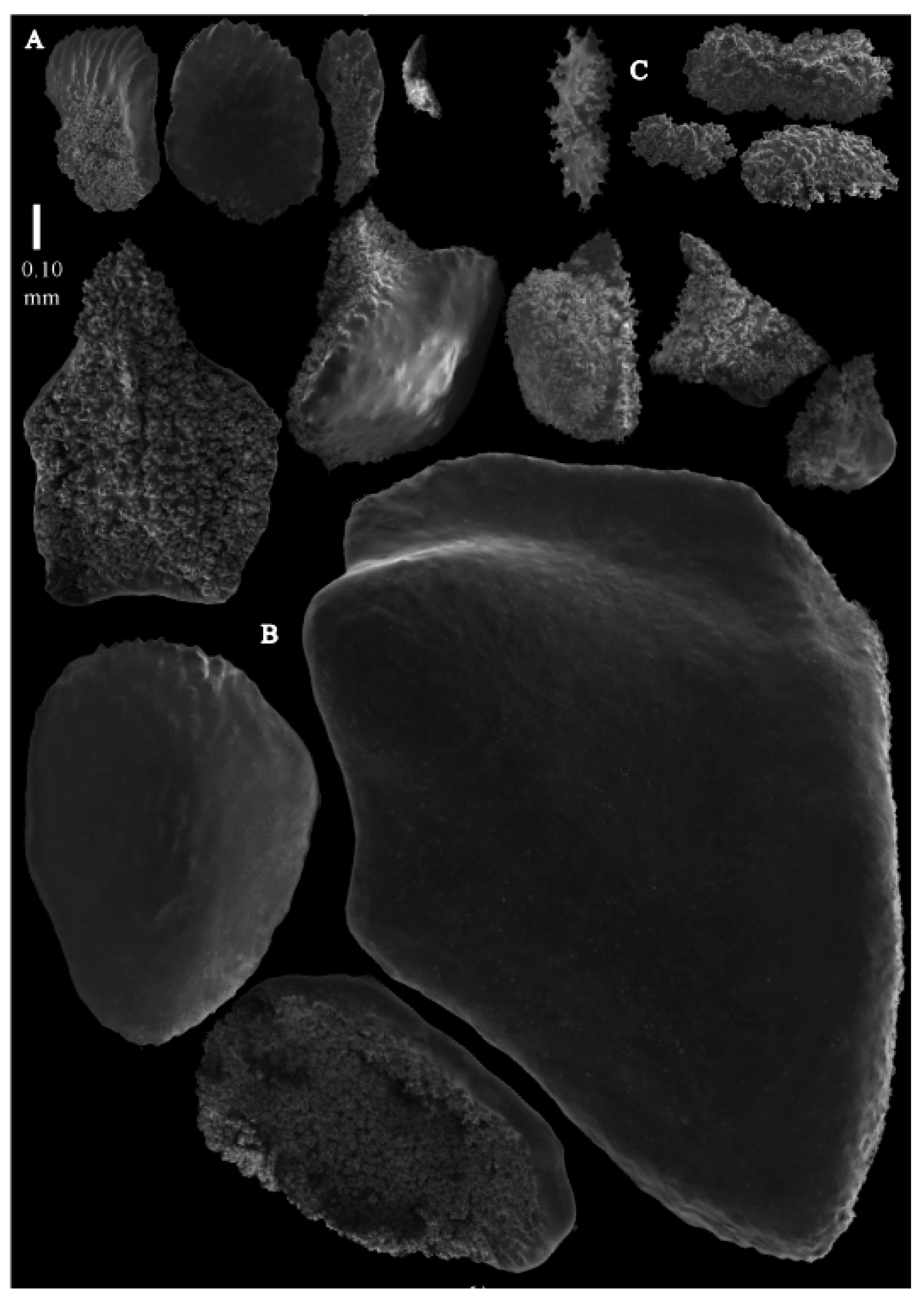
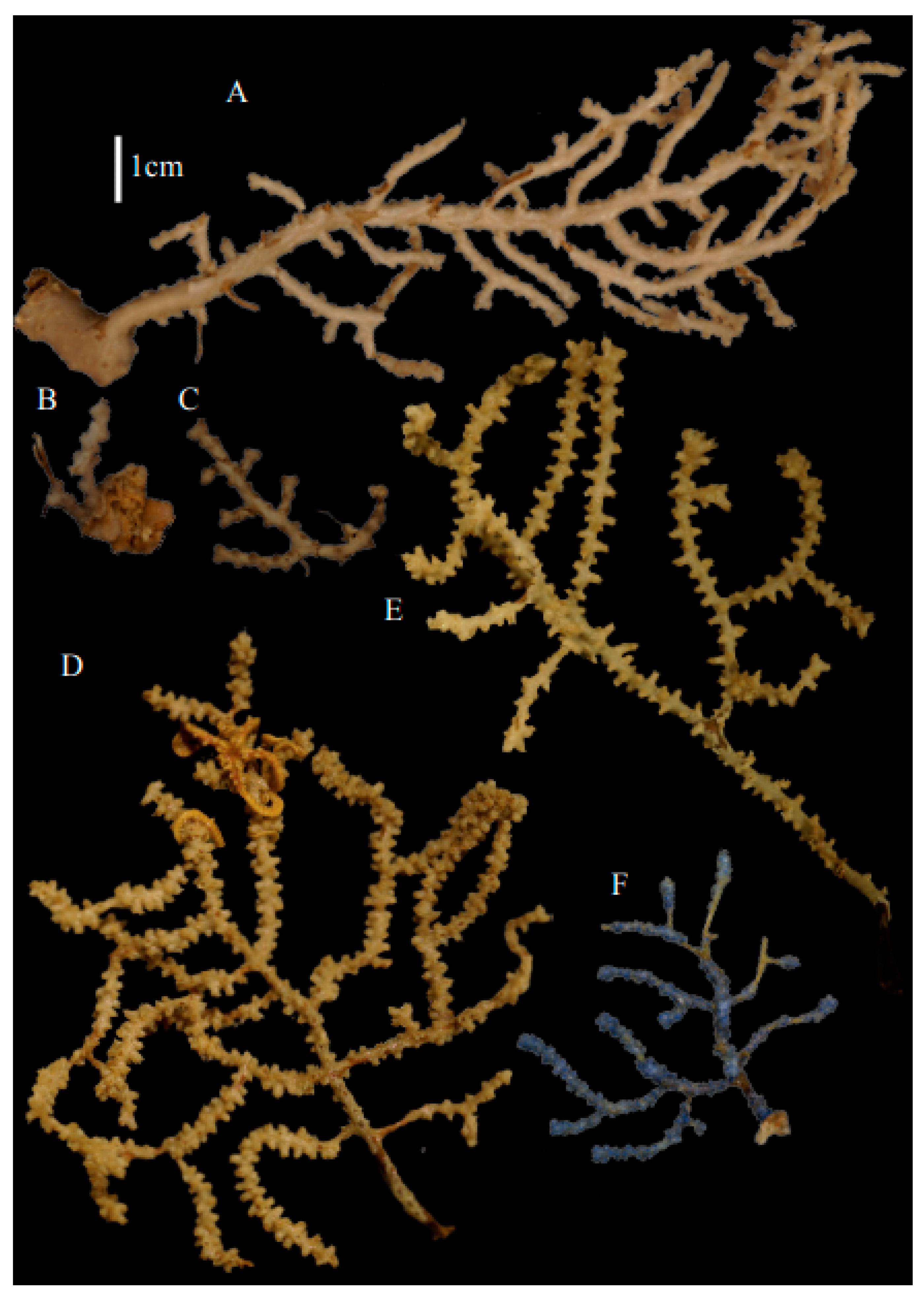
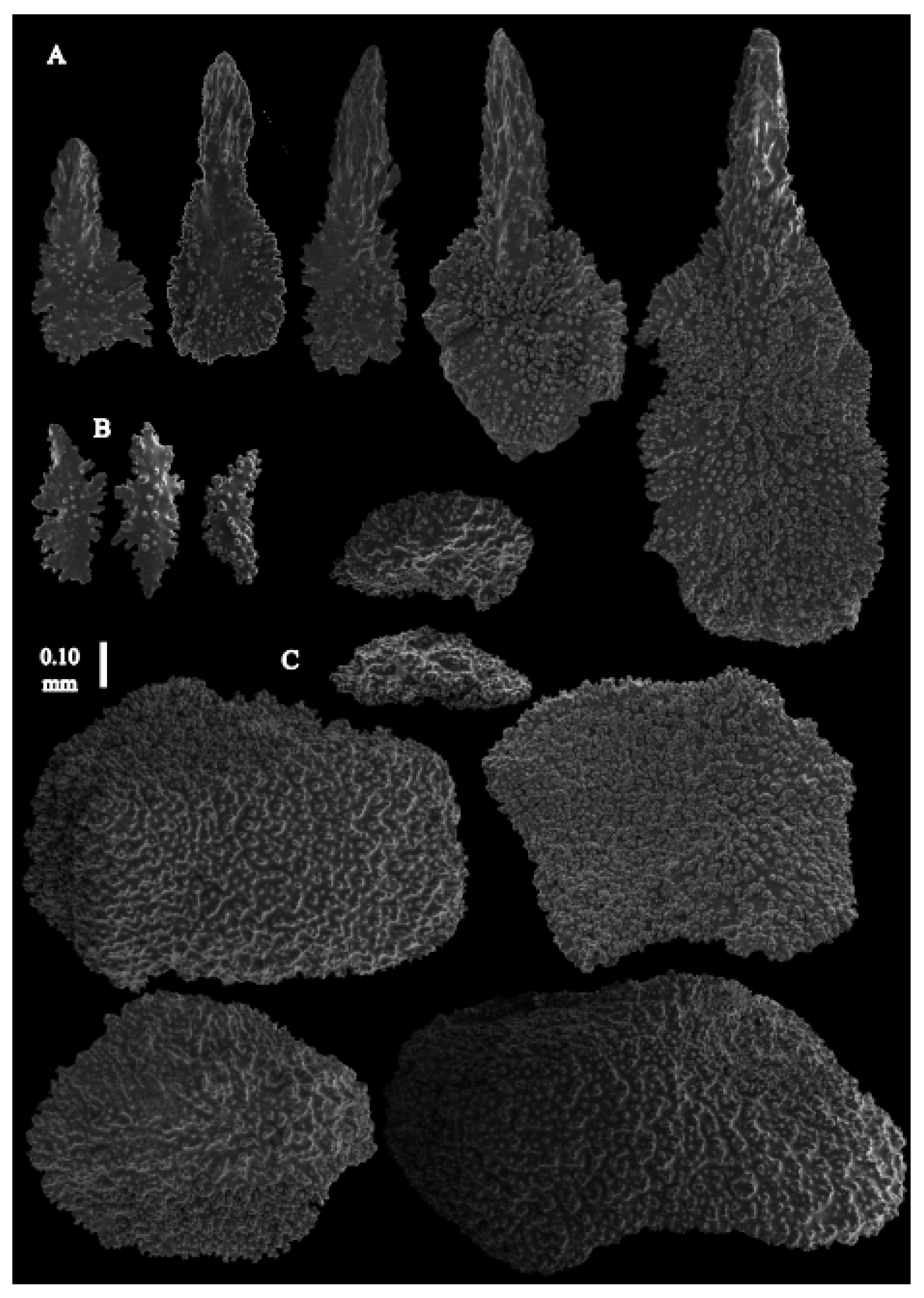
Re-description. Only a fragment of the branch (1.9 cm length) remained (
Figure 1A,A’), but the original specimen was part of a sparsely branched colony which was 9 cm high and 7 cm wide (Ridley pl. 5
Figure 1 [
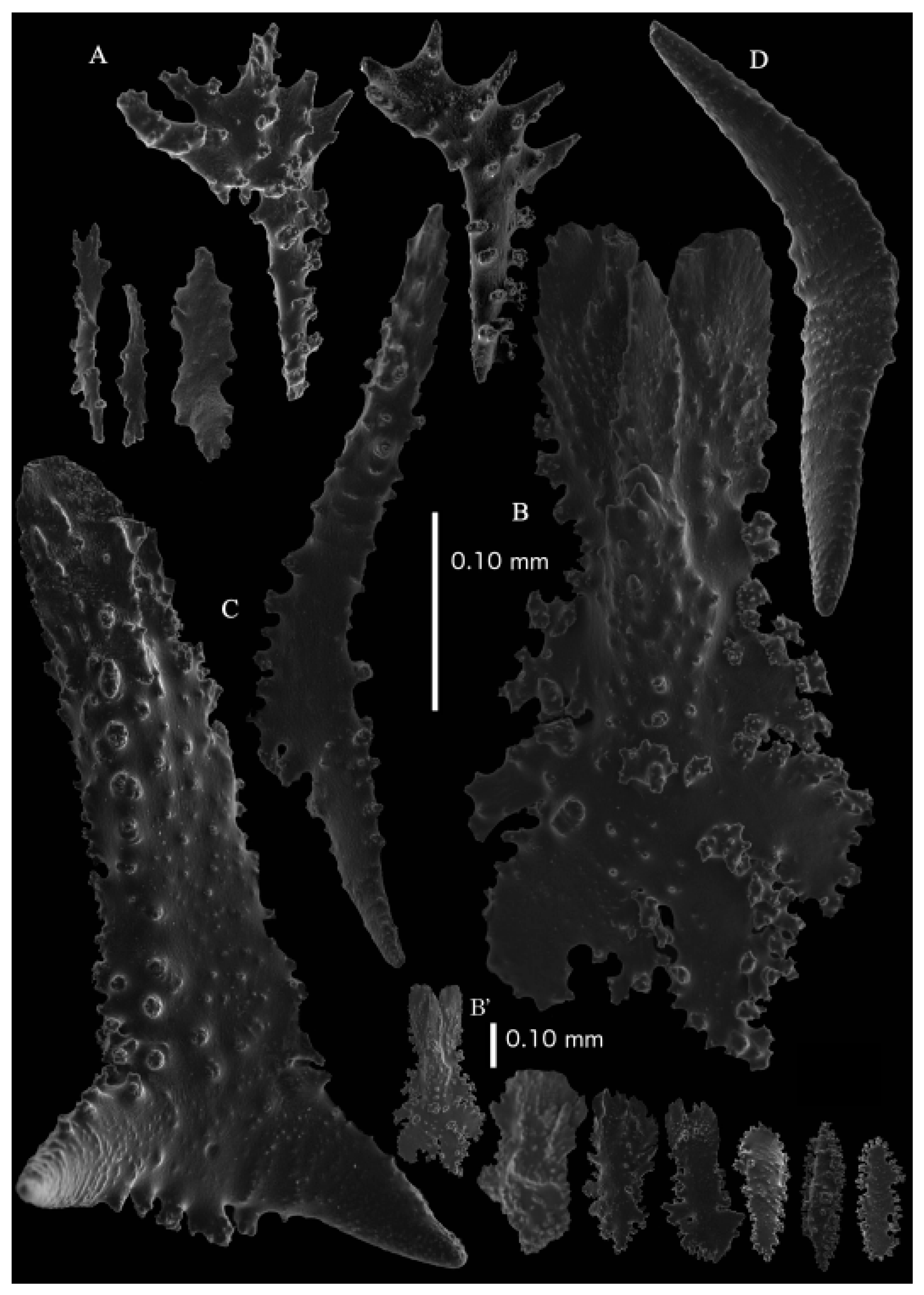
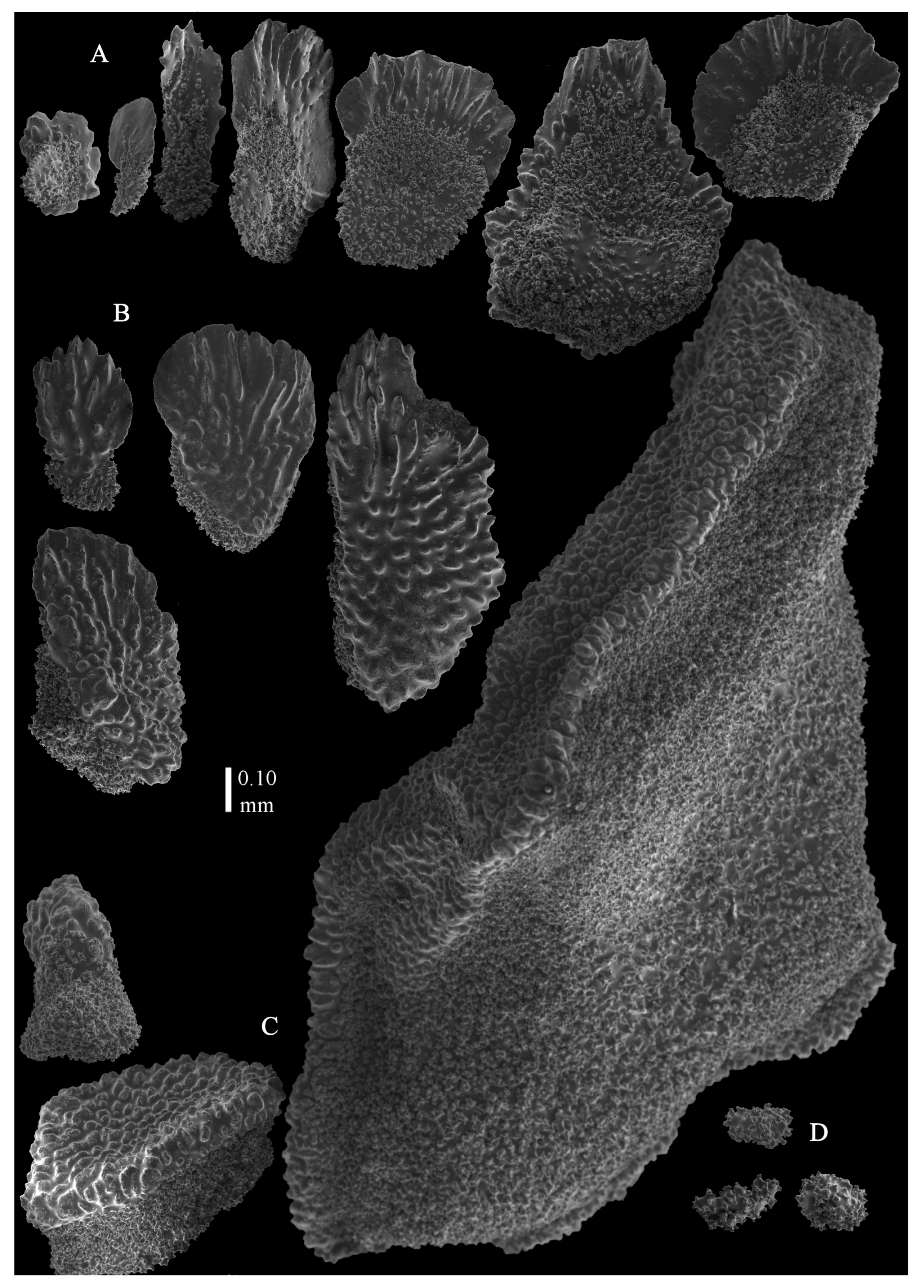
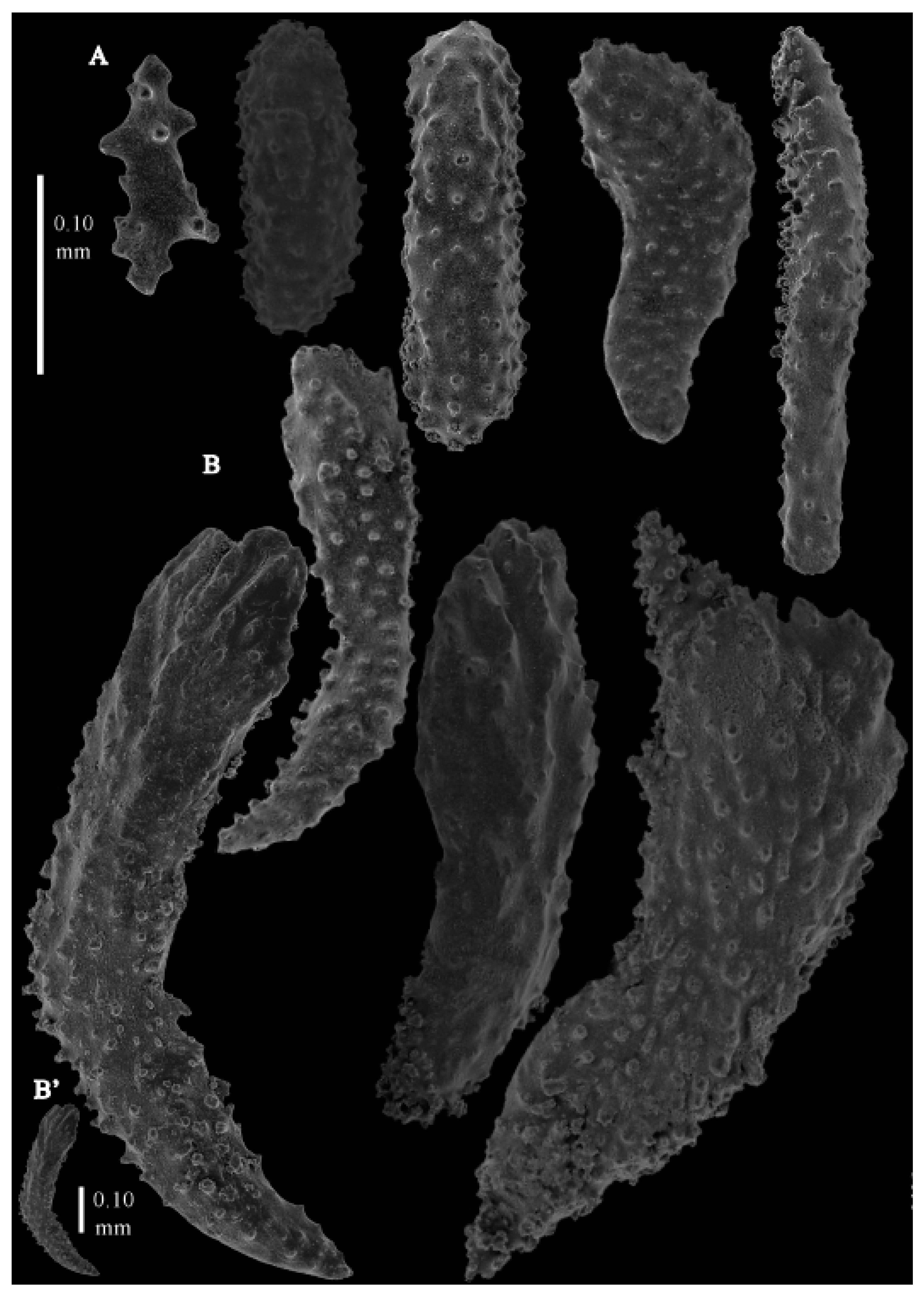
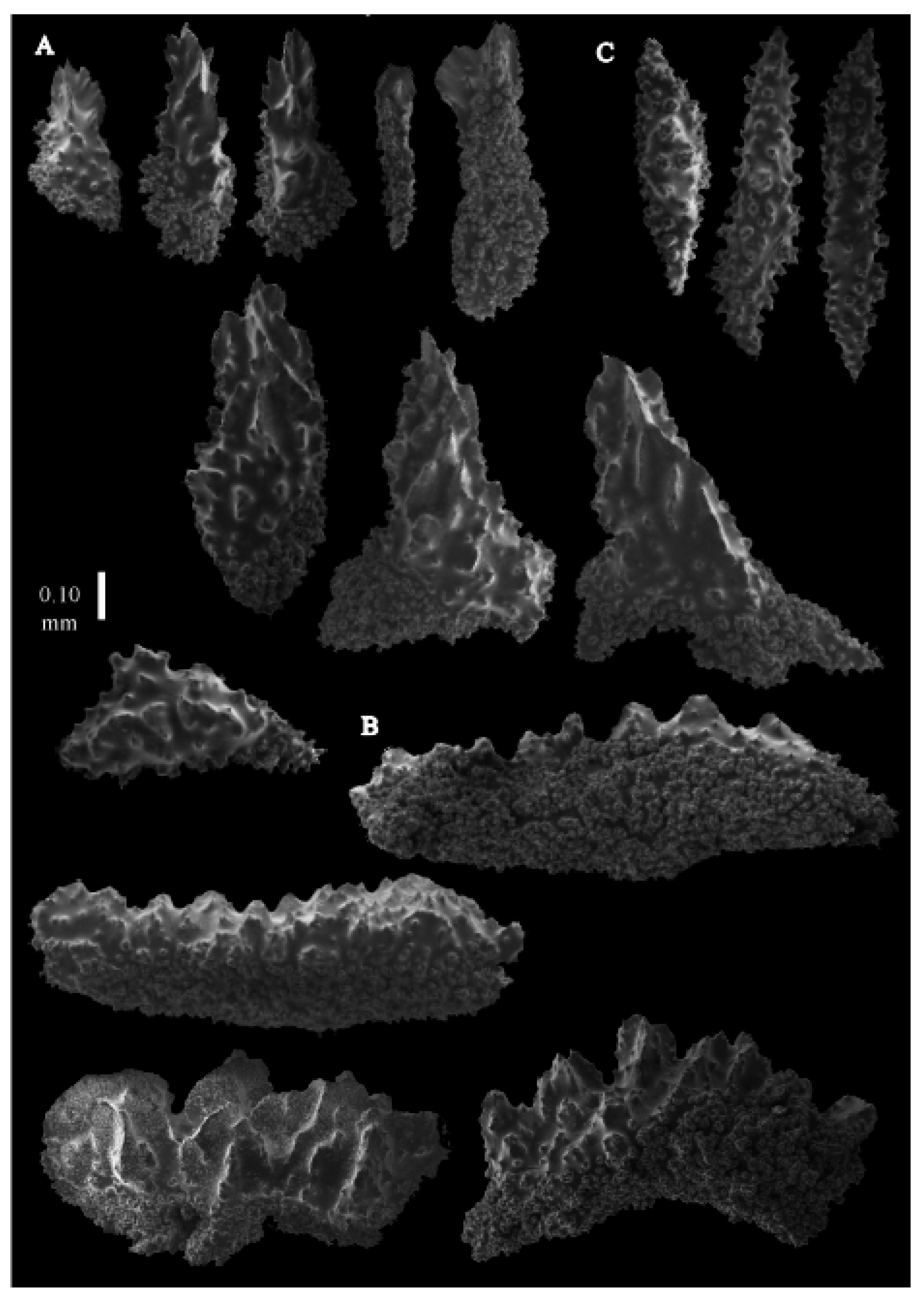
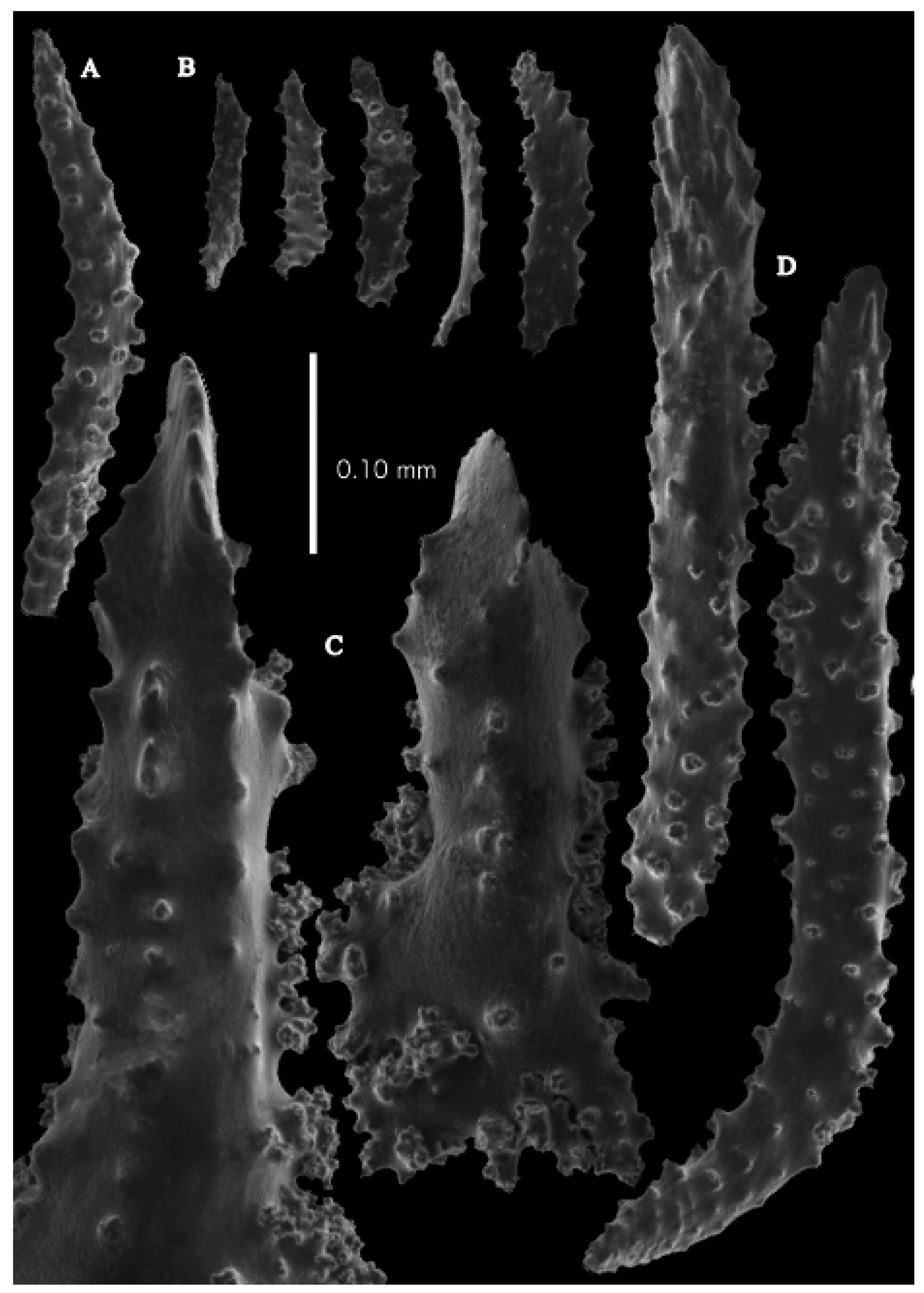
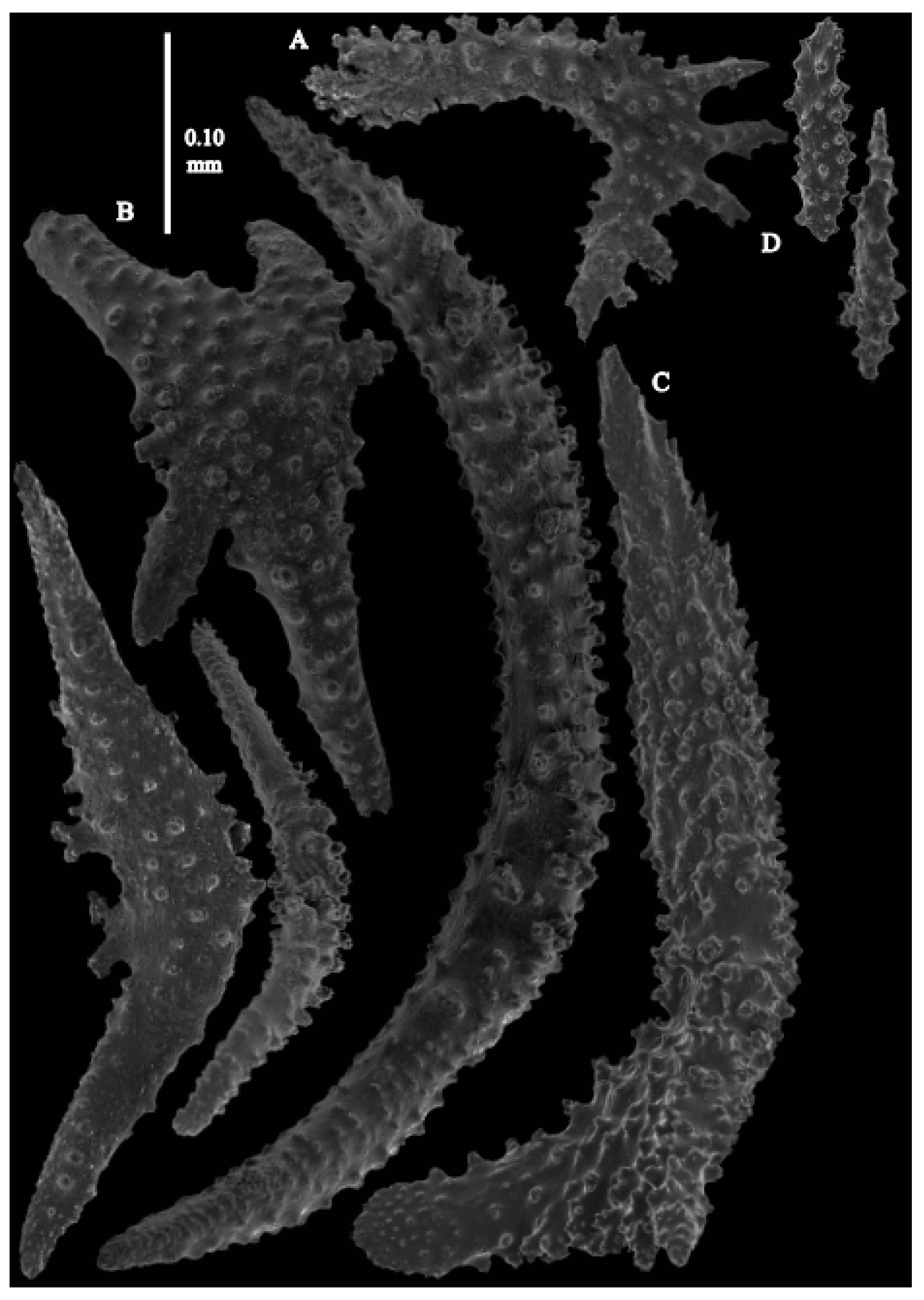
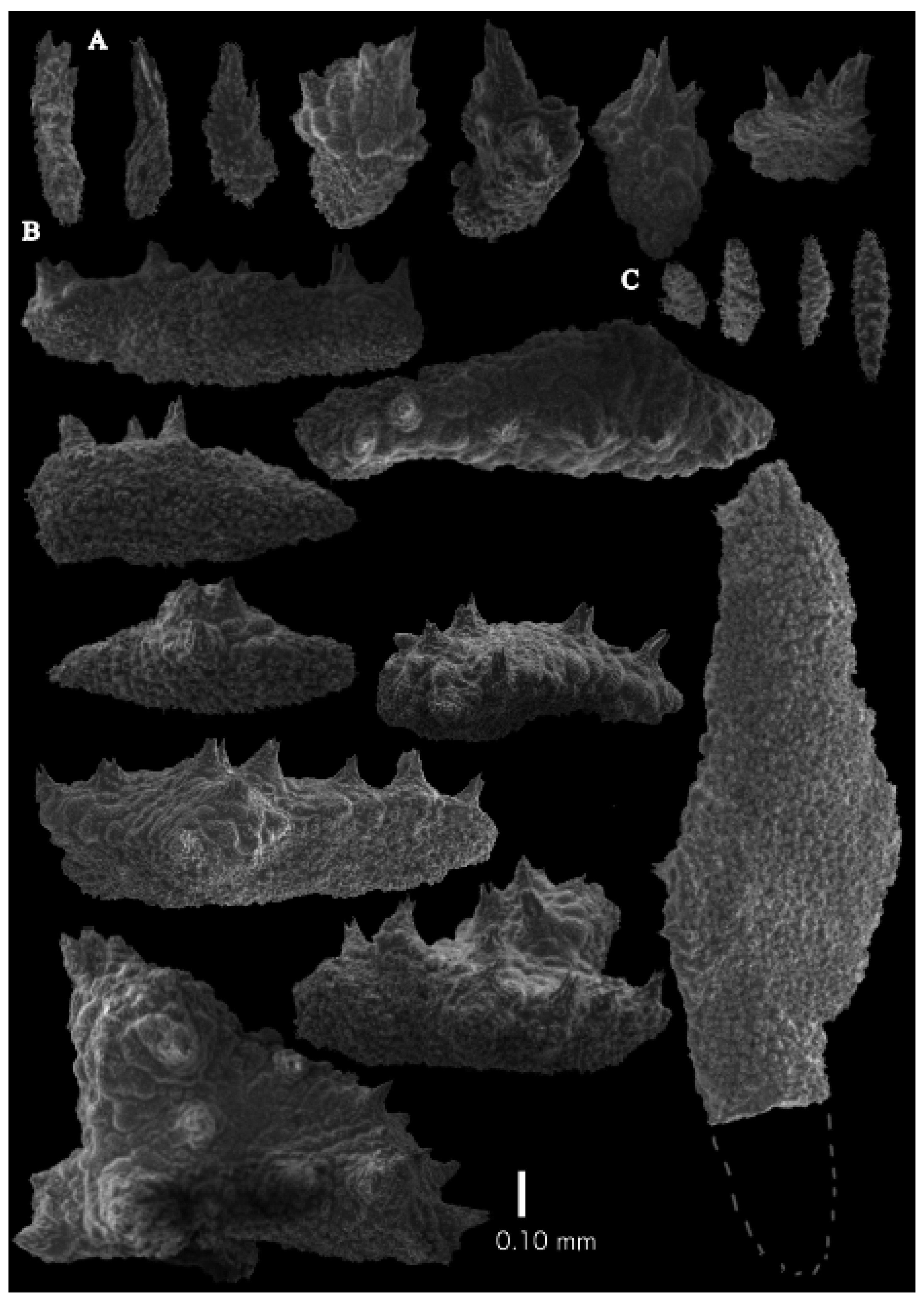
12]). The branches were about 1–1.4 mm wide. Calyces conical, 1–1.7 mm apart, present on three sides of the colony, leaving one side free of polyps. Polyps with collaret and points (
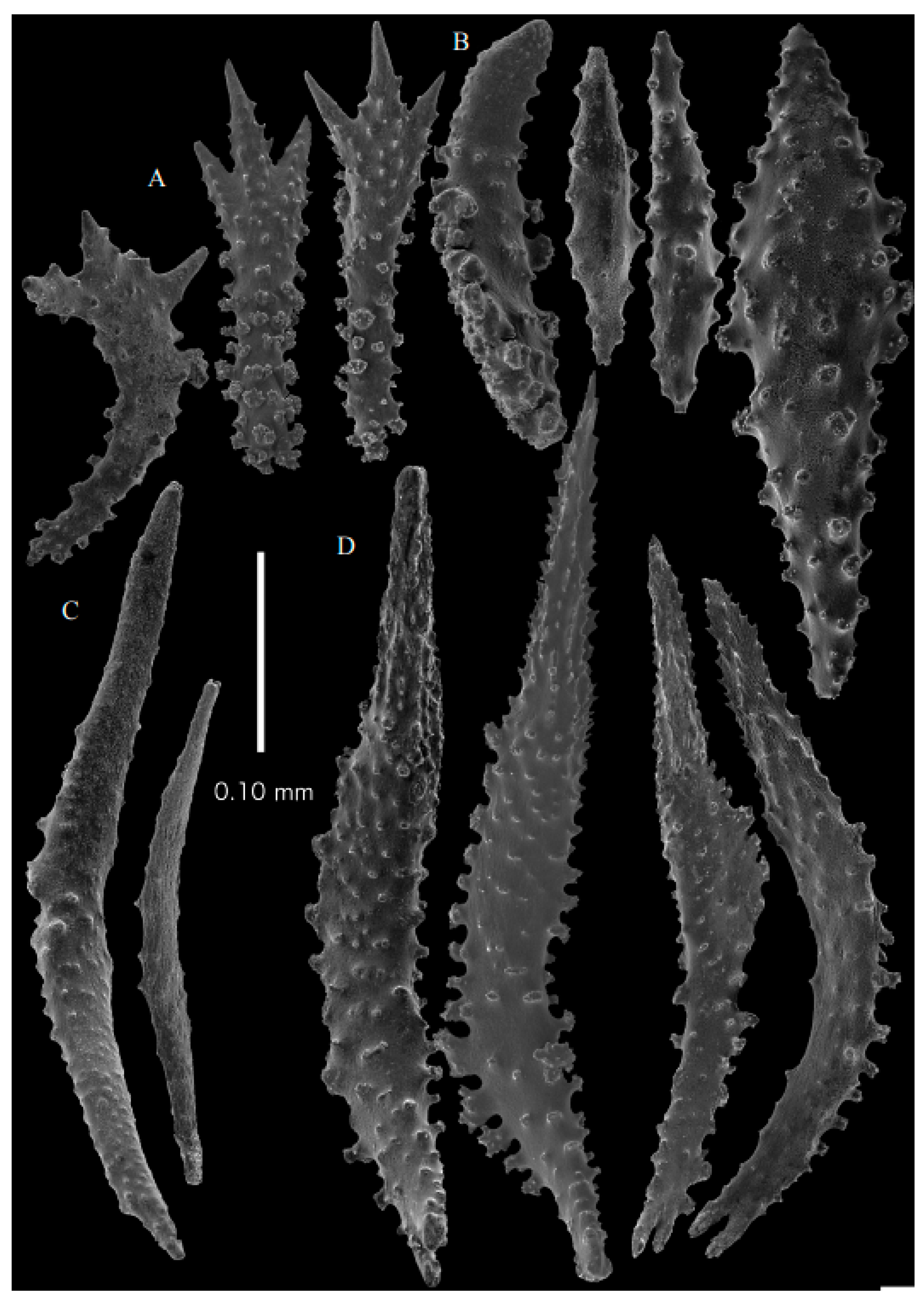
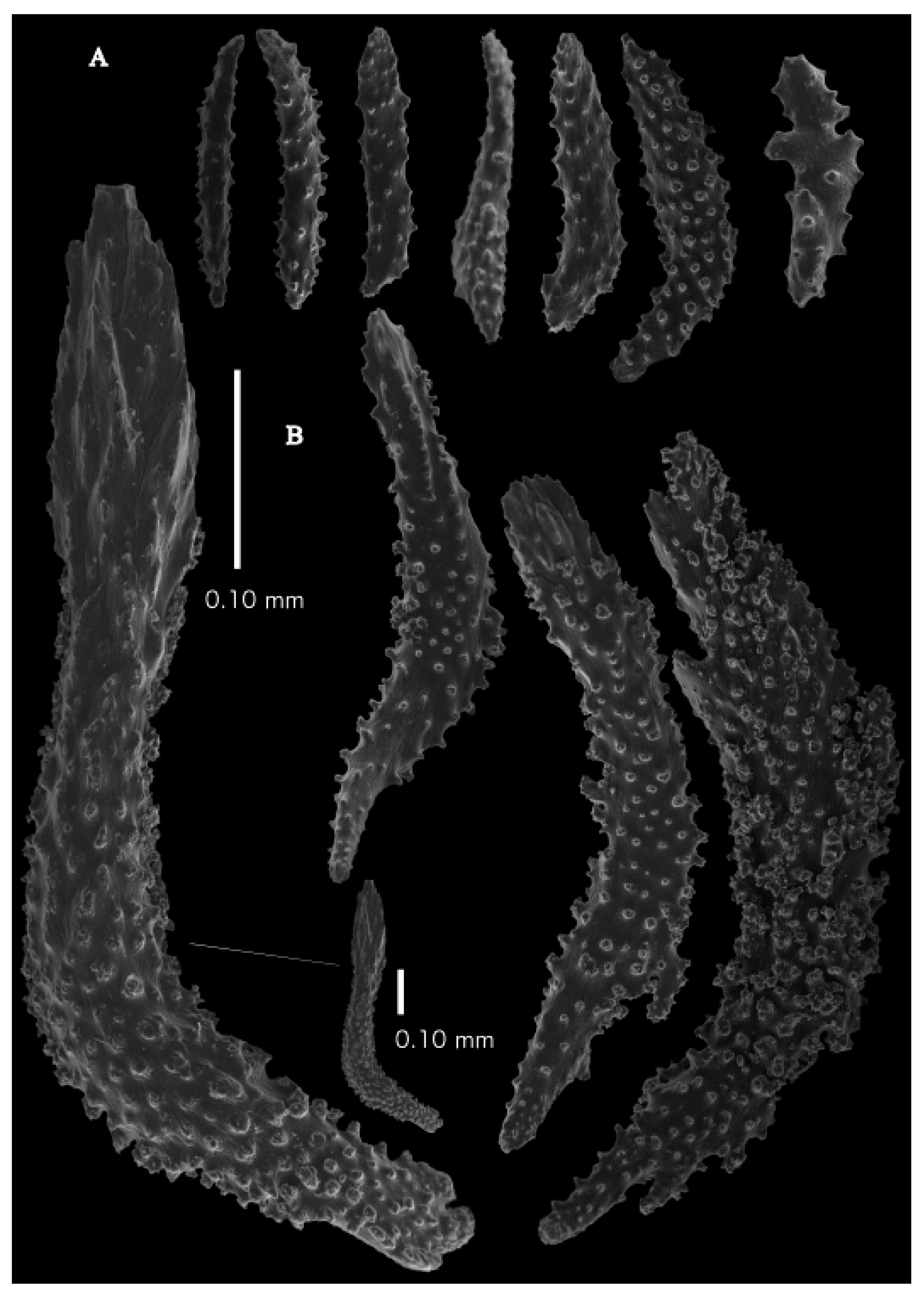
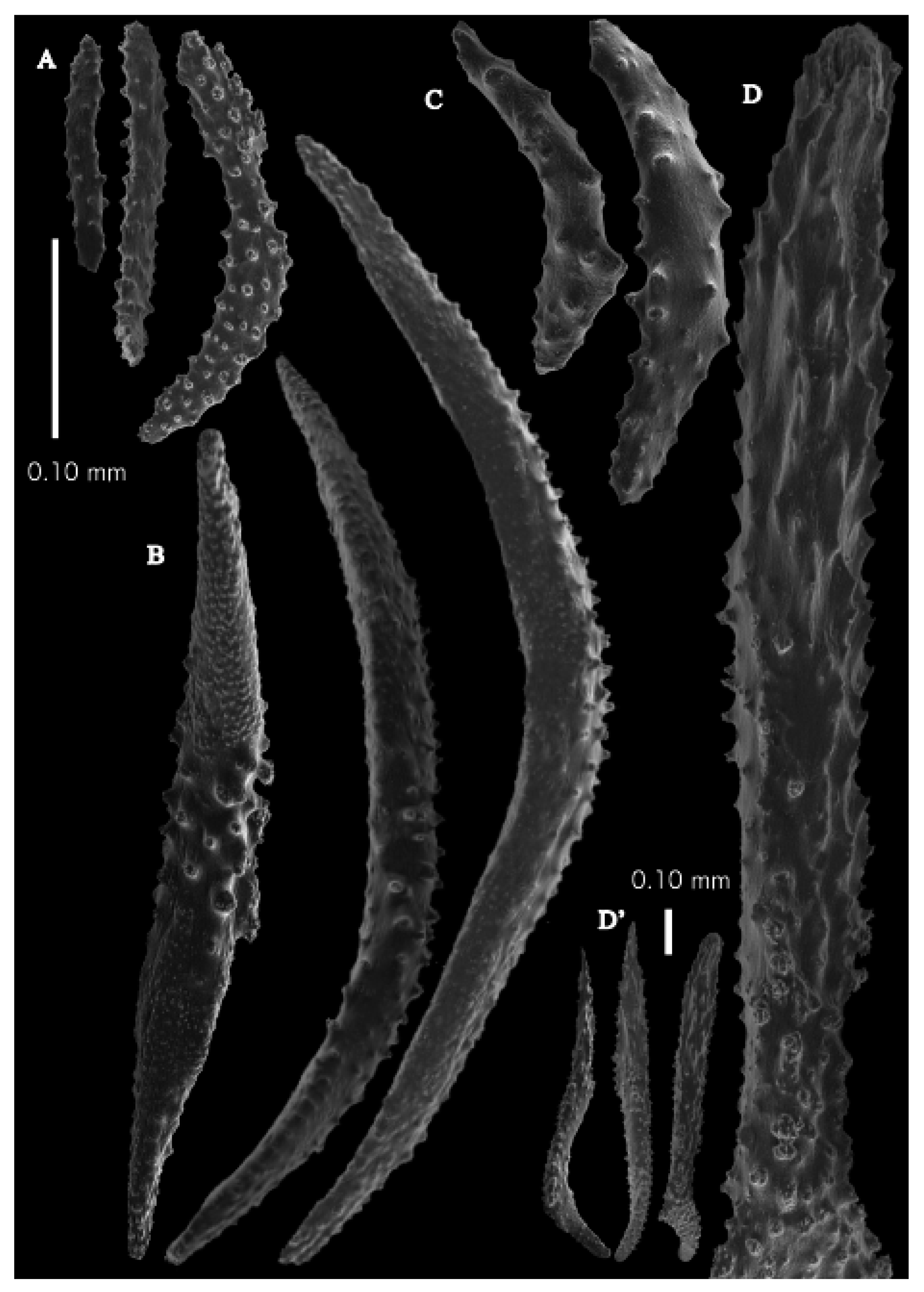
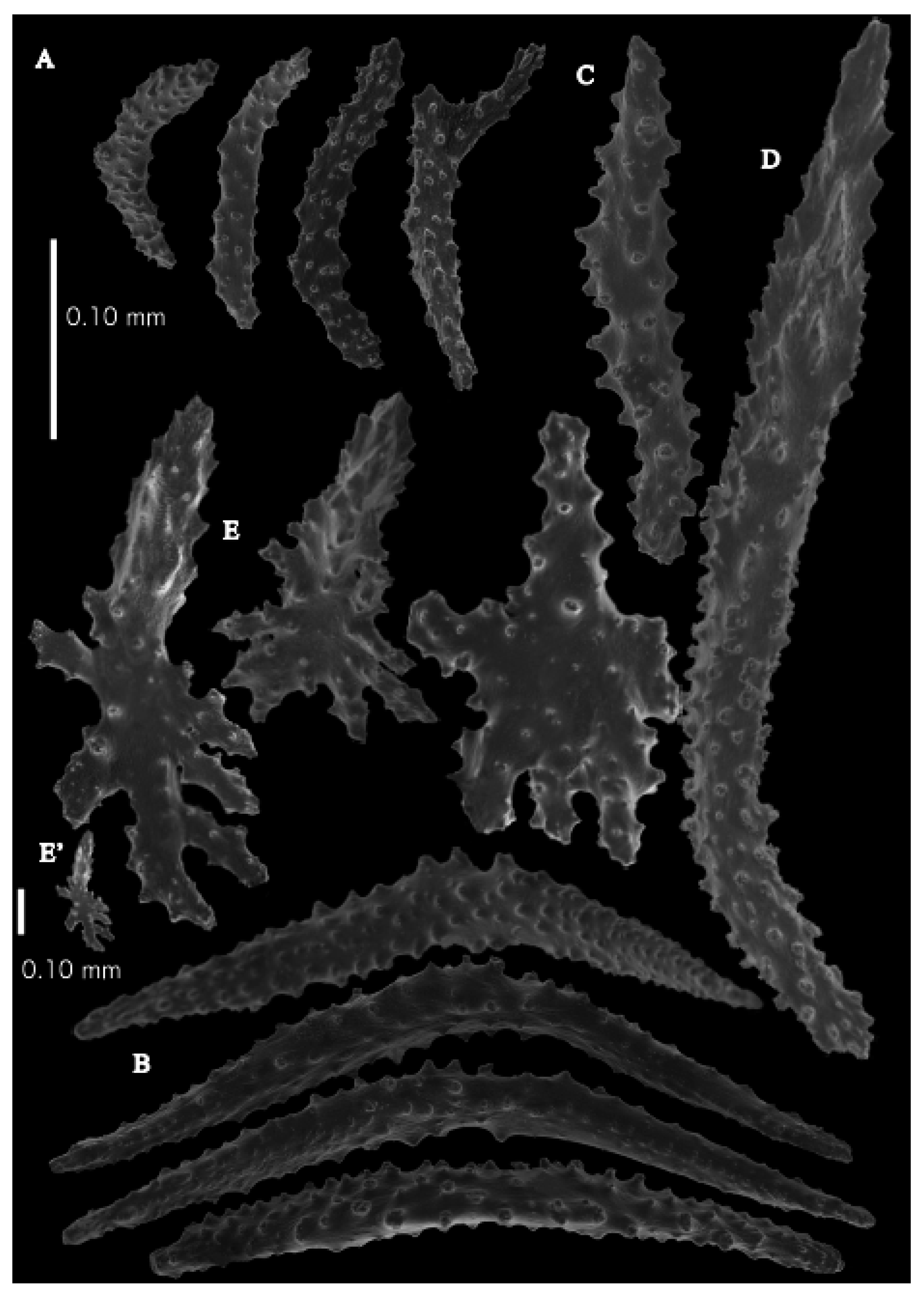
Figure 2A). Point sclerites are bent spindles (
Figure 3D), up to 0.5 mm long, with a lower part with small side branches and simple tubercles, and a spiny upper part. Collaret with almost smooth bent spindles (
Figure 3C), up to 0.4 mm long, with only some simple tubercles in the middle. Tentacles with dragon wing scales, sometimes torch-shaped (
Figure 3A), up to 0.2 mm long, with simple tubercles. Calyces with thornscales (
Figure 4A) 0.2–0.6 mm long. Lower part of these thornscales branched, with simple and complex tubercles and rounded elevations with prickles, upper part with simple tubercles and spines. Coenenchyme with big spindles, up to 1.5 mm long (
Figure 4B,B’). The outer side of these sclerites with rounded elevations with prickles, the inner side with simple and complex tubercles. Interior with smaller spindles, up to 0.4 mm long, with simple tubercles (
Figure 3B).
Colour. Colony white, anthocodial sclerites colourless, coenenchymal sclerites white.
Distribution. Only recorded from type locality.
Remarks. Ridley’s
Figure 5 and
Figure 6 [
12] made Bayer [
10] assume the coenenchymal sclerites had spines; consequently, he identified a specimen from Bikini as this species. This mistake was never corrected, leading to an erroneous part in Bayer’s key to the genera:
In his footnote 15 in the key, Bayer [
13] stated, “Although I have previously synonymised this genus (
Pseudothesea) with
Placogorgia (Bayer, 1959), it seems to be generically distinct, at least from the type-species of that genus. It may, however, be impossible to distinguish it from
Paracis, from which it was distinguished by Kükenthal [
3] on the basis of having sclerites of very diverse form, including strongly spinose, unilateral thornscales, but these occur in both
Pseudothesea and
Paracis” [
Neoacis in this manuscript]. Later on (1996), Grasshoff [
15] did examine the type of
P.
orientalis to describe his
P.
caecilia from New Caledonia but did not provide any images of the coenenchymal sclerites of
P.
orientalis. He mentioned calycular thornscales and tentacular dragon wing sclerites but did not say anything about the coenenchymal sclerites. Here, we finally present the coenenchymal sclerites of
Neoacis orientalis (type of the genus
Paracis), some of which resemble Ridley’s
Figure 5 and
Figure 6 [
12], but it is obvious spines are not present but only rounded elevations covered with prickles.
Neoacis bayeri sp. nov
urn:lsid:zoobank.org:act:D0B57ED9-1BDC-435C-91C3-C6685A085BB3
Material examined. Holotype
USNM 44078, previously identified by Bayer [
10] as
Paracis orientalis. Holotype Off Enyu Pass, Bikini Atoll, Ralik Chain, Marshall Islands, North Pacific Ocean, 11°4731′ N, 165°526′ E, 213–221 m, Bikini Scientific Resurvey, 1947, st. 30, coll. Russell, R. D., 22 August 1947.
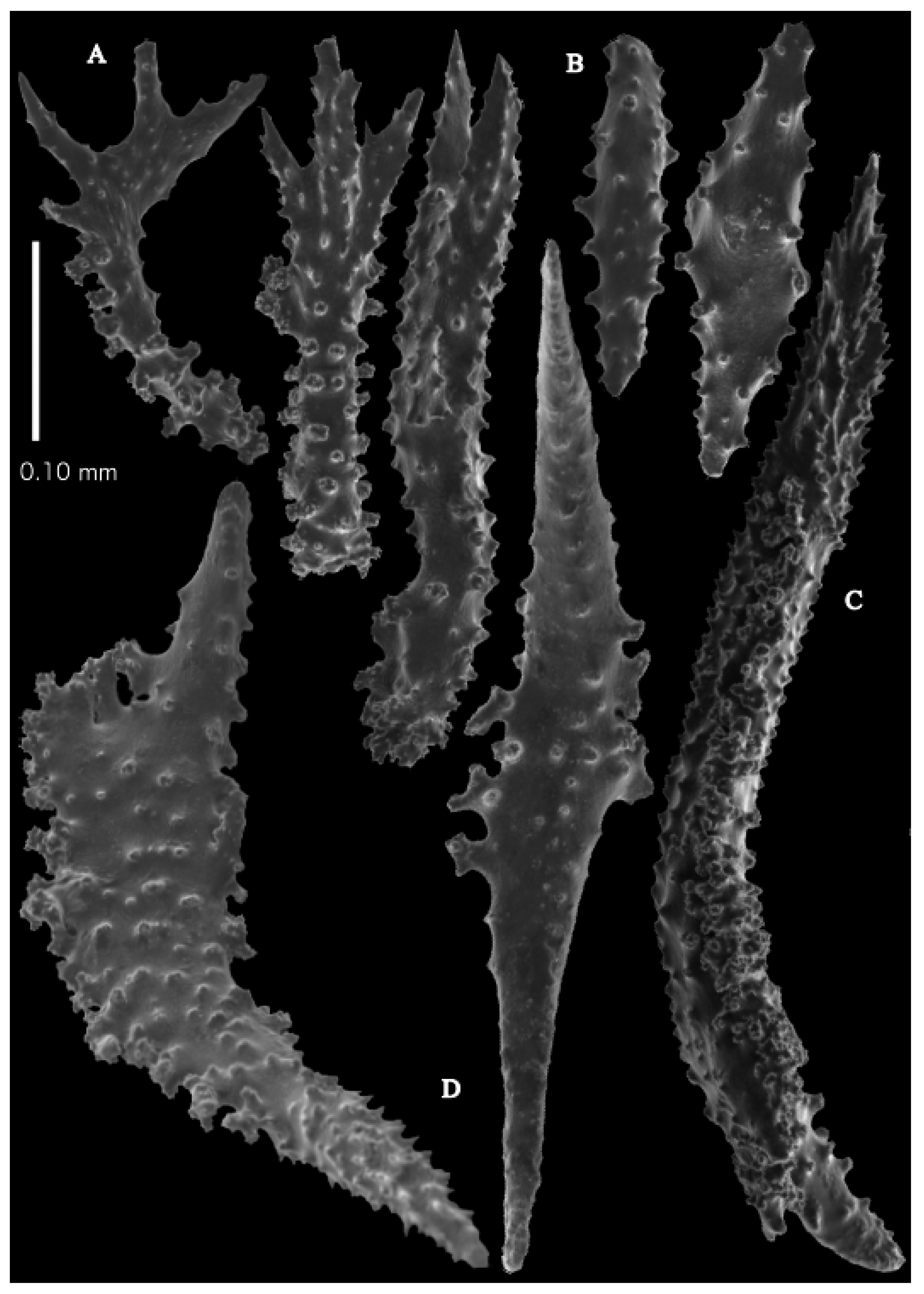
Description. Holotype 8.5 cm high and 7.7 cm wide (
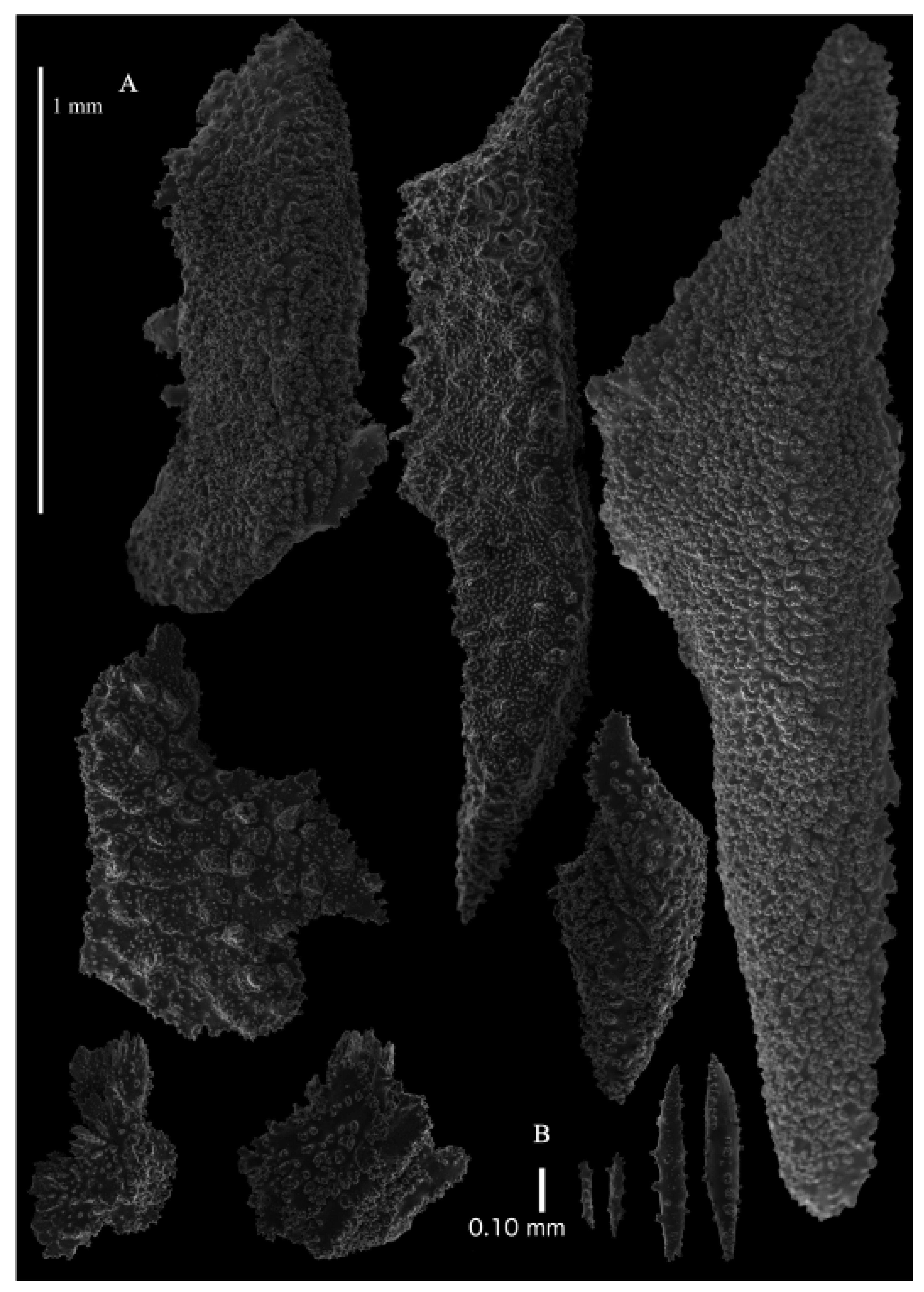
Figure 2B). Two stems arose from the spreading, encrusting base covering a branch of coral, but the two stems are separated now. Lateral branches arise at intervals of 10–15 mm along the main stem. Branches about 1.5–2 mm wide. Longest branch length 3.6 cm. Calyces conical, 1–3 mm apart, present all around the branches, partly two sides. Polyps with collaret and points (
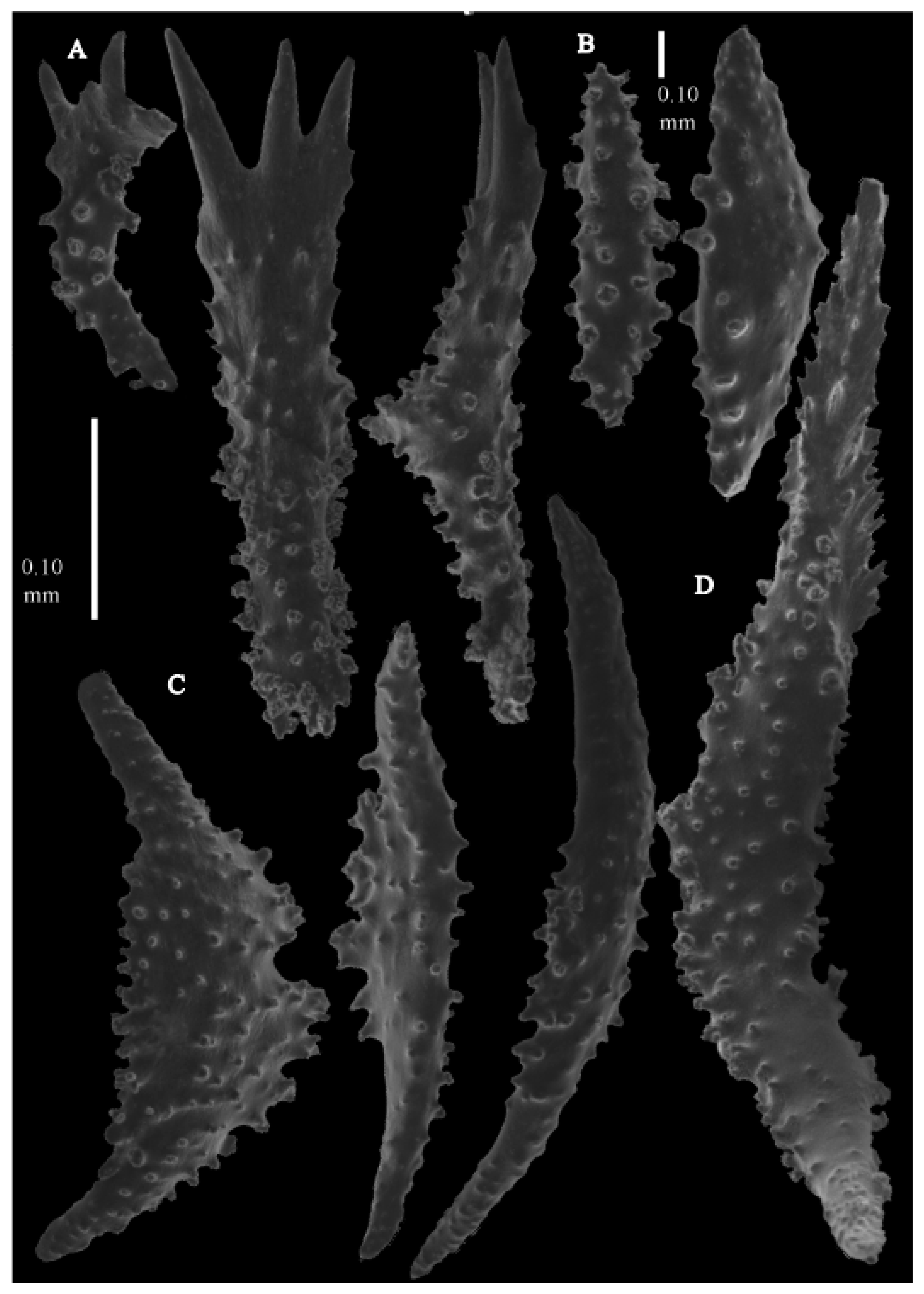
Figure 3B). Point sclerites are bent spindles (
Figure 5D), up to 0.7 mm long, with simple tubercles and spiny upper part. Collaret with bent spindles (
Figure 5C and
Figure 6C), up to 0.6 mm long, with simple tubercles, both ends less tuberculate. Tentacles with dragon wing scales (
Figure 5A), up to 0.35 mm long, with simple tubercles. Calyces with thornscales (
Figure 6A), 0.45–0.65 mm long. Lower part of these thornscales branched, with simple, complex, and rounded tubercles, upper part with tuberculate spines. Coenenchyme with big spindles, up to 1.5 mm long (
Figure 6B). The outer side of these sclerites with rounded tubercles, often placed on low protuberances, the inner side with simple and complex tubercles. Interior with smaller spindles, up to 0.4 mm long, with simple tubercles (
Figure 5B and
Figure 6D).
Colour. Colony brown-grey with darker calyces. Anthocodial sclerites colourless, coenenchymal sclerites white.
Distribution. Only known from the type locality.
Etymology. Named in honour of Dr. Ted Bayer, who first described this material as
Paracis orientalis in 1949 [
10].
Remarks. This species very much resembles Paracis orientalis but differs in having coenenchymal spindles with fewer rounded elevations with prickles. Colony differs in having the polyps on all sides of the branches instead of three sides in P. orientalis.
It is noteworthy that the material we here re-describe looks different from the description of Bayer [
10]. He only depicted sclerites with spines ([
10]
Figure 5C) and not a single spindle with rounded elevations covered with prickles ([
10]
Figure 6V). However, in his description he mentioned “coenenchyme covered with irregular elongate plates very closely fitted as in mosaic. These ornamented with low protuberances, quite spinose near the calyces.” These clearly are the spindles with rounded elevations covered with prickles we found.
Neoacis marianaensis sp. nov.
urn:lsid:zoobank.org:act:84C13C51-1973-4B0D-B154-BC539C66430A
Material examined. Holotype, USNM1439577, 54 Fathom Bank, leeward coast, Rota Island, Northern Mariana Islands, North Pacific Ocean, 14.2067′ N, 145.18′ E, depth 98–164 m, R/V Panglou Oro, st. 11, coll. Grigg, R. W.; Eldredge, L. G., 25 October 1973.
Description. Holotype colony (USNM1439577) 13.5 cm high and 10.6 cm wide (
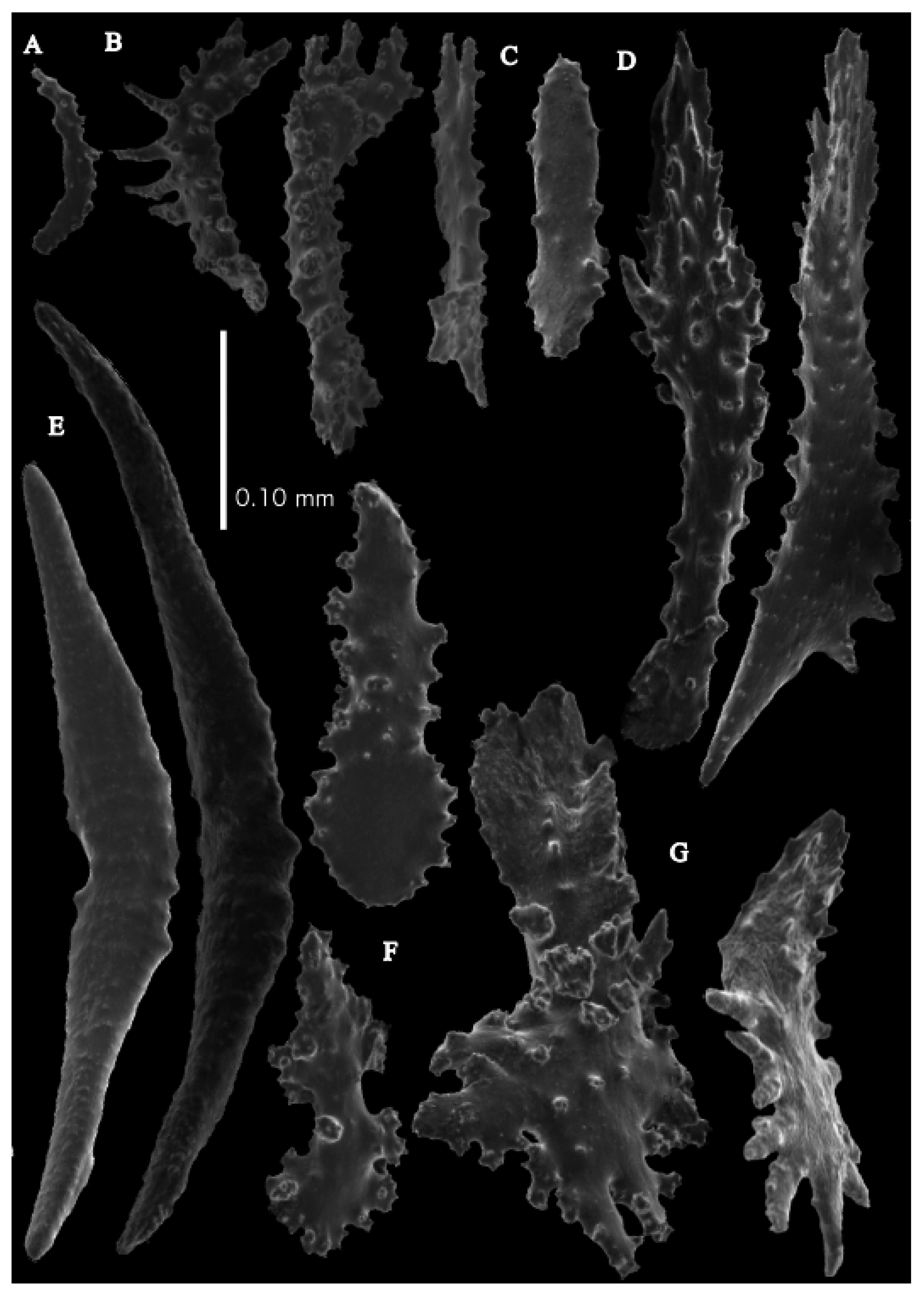
Figure 7A). Stem and branches flattened in the plane of branching. Branches about 0.93–3.42 mm wide. Calyces conical, 1.12–2.04 mm apart, present on three sides of the branches. Polyps with collaret and points (
Figure 2F). Point sclerites are bent spindles (
Figure 8D), up to 0.31 mm long, with simple tubercles and spiny upper part. Collaret with bent spindles (
Figure 8C), up to 0.27 mm long, with simple tubercles and both ends less tuberculate. Tentacles with dragon wing scales, but more torch-shaped (
Figure 8A), up to 0.17 mm long, with simple tubercles. Calyces with thornscales (
Figure 8E and
Figure 9A), up to 0.34 mm long. Lower part of these thornscales branched, with simple, complex, and rounded tubercles, upper part with one or two blunt spreading spines, and median part of these spines thickened. Coenenchyme with big spindles, up to 1.60 mm long (
Figure 8B). These sclerites with small spines on the outer surface and complex tubercles, often placed on low protuberances. Interior with smaller spindles, up to 0.26 mm long, with sparse simple tubercles (
Figure 8B and
Figure 9C).
Colour. Colony white with reddish-pink polyps. Anthocodial sclerites reddish-pink, coenenchymal sclerites white.
Distribution. The species is only known from the Northern Mariana Islands, North Pacific Ocean.
Etymology. The species is named after its type locality, Northern Mariana Islands.
Remarks. This species mostly looks like Japanese Neoacis pustulata to the naked eye and has large spindles in the coenenchyme with dragon wing sclerites in the tentacles, but differs from that species in having thornscales with projections thickened in the median part like Placogorgia coronata.
Neoacis ofwegeniMatsumoto, sp. nov.
urn:lsid:zoobank.org:act:D2316D74-E802-4346-BAEF-C7C7D9E82F24
Material examined. Holotype USNM49911 previously identified as Paracis sp. Alb- USFC/A3716, Ose Zaki, Suruga Bay, Honshu Island, Shizuoka Pref. Japan, Northwest Pacific, depth 119–229 m, Trawl–Tanner Beam, 8 ft, 11 May 1900, paratype SMBL-Cni1362 previously identified as Paracis squamata, off Kirime-zaki cape, depth 30–50 m, Sakai fish port, Minabe, Wakayama, Japan, 21 Mar. 1969, (probably collected by fisherman’s boat), dry.
Description. Holotype (USNM49911) colony 11.8 cm high and 8.9 cm wide (
Figure 1C). Lateral branches arise at intervals of 9.5–21.3 mm along the main stem. Branches about 2.38–3.77 mm wide. Calyces conical, 1.40–2.86 mm apart, present all around the branches. Polyps with collaret and points (
Figure 2C). Point sclerites are bent spindles (
Figure 10C,C’), up to 0.78 mm long, with complex tubercles and spiny upper part. Collaret with bent spindles (
Figure 10D,D’), up to 0.59 mm long, with simple tubercles, both ends less tuberculate. Tentacles with dragon wing scales (
Figure 10A), up to 0.34 mm long, with complex tubercles. Calyces with thornscales (
Figure 11A), 0.50–0.59 mm long. Lower part of these thornscales branched, with simple, complex, and rounded tubercles, upper part with tuberculate spines. Coenenchyme with big spindles, up to 1.67 mm long (
Figure 11C). The outer side of these sclerites with rounded tubercles, often placed on low protuberances, the inner side with simple and complex tubercles. Interior with smaller spindles, up to 0.76 mm long, with simple tubercles (
Figure 11B).
Colour. Colony brown-grey with darker calyces. Anthocodial sclerites colourless, coenenchymal sclerites white (
Figure 1C).
Variation. The paratype (SMBL-Cni1362) colony shape and branching looks a bit different from the holotype because the specimen has been stored under insufficient condition and is not enough to describe the detail of the polyps (
Figure 1D). Sclerites are very much like the holotype (
Figure 12 and
Figure 13).
Distribution. Suruga Bay, off Kii Peninsula (Pacific side of Japanese main island). The other known Japanese Neoacis species (N. pustulata) has a wider distribution than N. ofwegeni sp. nov. N. orientalis and N. bayerii sp. nov. have never been found in Japan.
Etymology. Named in honour of Dr. Leen P. van Ofwegen, who first recognised this species as new to science.
Remarks. This species differs from Neoacis orientalis and N. bayeri sp. nov (USNM44078) in having much more complex tubercles on point and tentacle sclerites. The coenenchymal spindles mostly resemble those of N. orientalis.
Neoacis pustulata(Wright and Studer, 1889)
Acis pustulata Wright and Studer, 1889: 122, pl. 24 Figure 1-1a, pl. 27 Figure 6 (Sagami Bay); Kinoshi-ta 1909: 5, pl. 1 Figure 2, pl. 2 Figures 10–12 (Uji island); Thomson and Simpson, 1909: 234, pl. 1 Figure 6, 7, pl. 9 Figure 5a, 5b.
Paracis pustulata Kükenthal 1924: 157, Figures 106, 107.
Material examined. Holotype, BMNH1889.5.27.140 Acis pustulata, Hyalonema-ground (Nishi-no yodomi), south of Japan, 35°11′ N 139°28′ E, H.M.S. Challenger st. 232, 345 fms, 12 May 1875. Other materials, UMUTZ-G108, Acis pustulata, Coral ground, Uji Is.: Satsuma (Kagoshima Pref.), depth 146–164 m (80–90 fathoms), det. and coll. Kumao Kinoshita, June, 1908, red; UMUTZ-G110, Acis pustulata, Coral ground, Uji Is.: Satsuma (Kagoshima Pref.), depth 146–164 m (80–90 fathoms), det. and coll. Kumao Kinoshita, late May, 1908, red; UMUTZ-G253 (=G110b), same data as UMUTZ-G110, figured colony, Kinoshita, 1909, white; UMUTZ-G201, Acis pustulata, Chikurase (Tsukurase?) Kagoshima Pref. depth 128 m (70 fathoms), Satsuma expedition, coll. M. Miyajima, 14 July 1899, Coral net, white; USNM50126 Acis pustulata, subsampled from UMUTZ-G253, white and UMUTZ-G110, red; previously unidentified material: ZMUC-ANT706, Northwest Pacific, 32°15′ N, 128°20′ E, depth 180 m (100 fms), coll. Kapt. Hans Christiansen, 17 April 1926, white; AKM418, Watari-se bank, off Izu Isls., Japan, Northwest Pacific, 34°02.8620′ N, 138°54.8090′ E–34°02.9190′ N, 138°54.6810′ E, depth 101.1–106.2 m, R/V Tansei-maru cruise, KT04-06, st. WS-2, coll. A.K. Matsumoto, 30 April 2004, white; AKM569, Off Takato, Tachibana-ura, Otsuki, Tosa, Kouchi pref., Japan, Northwest Pacific, 132°48.06′ E, 32°43.08′ N–132°47.68′ E, 32°43.12′ N, depth 84.75–83.1 m, F/B Kiryo-maru, st. 3, Coral-net, coll. A. K. Matsumoto, 7 October 2004, red; AKM801, Off Takarajima Is., Tokara Isls., Japan, East China Sea, 29°14.6410 N, 129°07.8392 E, depth 156 m, R/V Tansei-maru, KT07-2 cruise, st.DT5 (D8), coll. Yokose, 2 March 2007, white; AKM805, Off Takarajima Is., Tokara Isls., Japan, East China Sea, 29°14.6410 N, 129°07.8392 E, depth 156 m, R/V Tansei-maru cruise, KT07-2, st.DT5 (D8), coll. Yokose, 2 March 2007, white; AKM1390, West of Chichijima Is. Ogasawara Isls., Japan, 142°07.412′ E, 27°01.395′ N–142°07.467′ E, 27°01.360′ N, depth 139–144 m, R/V Tansei-maru cruise KT09-2, st.TW01-01, coll. A.K. Matsumoto, 19 March 2009; AKM1466, West of Chichijima Is. Ogasawara Isls. Japan, 142°07.466′ E–142°07.489′ E, 27°01.371′ N–27°01.344′ N, depth 135–136 m, R/V Tansei-maru cruise, KT09-02, st.TW01-02, 19 March 2009, coll. A.K. Matsumoto, white; AKM1499, West of Chichijima Is. Ogasawara Isls. Japan, 142°07.166′ E–142°07.251′ E, 27°02.942′ N–27°02.948′ N, depth 138–136 m, R/V Tansei-maru cruise, KT09-02, st. TW02-04, coll.A.K. Matsumoto, 19 March 2009, white.
Re-description. Holotype (BMNH1889.5.27.140) colony 12.6 cm high and 8.5 cm wide (
Figure 14A). Stem and branches flattened in the plane of branching. Branches about 1.5–2 mm wide. Calyces conical, 1–1.5 mm apart, present on three sides of the branches (
Figure 14A is the polyp-free side of the colony). Polyps with collaret and points (
Figure 2D). Point sclerites are bent spindles (
Figure 14C), up to 0.39 mm long, with simple tubercles and spiny upper part. Collaret with bent spindles (
Figure 14D), up to 0.32 mm long, with simple tubercles, both ends less tuberculate. Tentacles with dragon wing scales (
Figure 14A), up to 0.20 mm long, with simple tubercles. Calyces with thornscales (
Figure 14B,B’), up to 0.4 mm long. Lower part of these thornscales branched, with simple, complex, and rounded tubercles, upper part with one or two blunt spreading spines. Coenenchyme with big spindles, up to 3.0 mm long (
Figure 15A). These sclerites with small spines on the outer surface and complex tubercles, often placed on low protuberances. Interior with smaller spindles, up to 0.5 mm long, with sparse simple tubercles (
Figure 15B).
Colour. Holotype colony white with violet polyps. Anthocodial sclerites violet, coenenchymal sclerites white.
Variation. UMUTZ-G253
Acis pustulata, white colour, which was figured colony in Kinoshita (
Figure 2E,
Figure 14B,
Figure 17 and
Figure 18). UMUT-G110 is the purplish-red coloured colony (
Figure 14C) and was in the same jar as UMUTZ-G253.
Distribution. East China Sea and Pacific side of Japanese water from Okinawa, Ogasawara (Bonin) Islands.
Remarks. This species differs from the other three described here in having thornscales with blunt-ended spines and coenenchymal sclerites having small spines on the outer surface. Kinoshita [
1] mentioned only one white colony, which is UMUTZ-G253. Kinoshita wrote in the description that “Namely in one specimen the spicules are white, while in all the other specimens they are purplish red.”. UMUTZ-G201, the white specimen, is not mentioned by Kinoshita [
1]. Kinoshita mentions that the polyps occur only exceptionally on the posterior surface of the colony (
Figure 14B).
Genus IMBRICACIS gen. nov.
urn:lsid:zoobank.org:act:5994B4BE-4EC4-4FBD-9459-D16C72EFCD7F
Type species: Acis ijimai Kinoshita, 1909.
Diagnosis. Colonies branch in one plane. Polyps situated on three sides. Coenenchyme with large polygonal thick plates, with their outer surface smooth or with rounded tubercles, inner surface granular. Calycular sclerites similar but smaller. Points with bulky rods with upper end smooth or spiny and lower end tuberculate. Collaret not present.
Etymology. Combination of imbrico (Latin for tile) and Acis.
Remarks. One other species has been described from Japan, Acis miyajimai Kinoshita, 1909. At least three other species exist in the Indo-Pacific, Acis squamata Nutting, 1910, a species from Hawaii described as belonging to a new genus in the unpublished thesis of K. Muzik (USNM56858) and a species from the Indian Ocean (USNM89348) newly described here. The genus differs from other genera with big plates in lacking a collaret and not having thornscales in the calyx.
Key to the species (five species).
| 1. Outer side of the coenechymal plates is smooth; | 2 |
| -. Outer side of the coenechymal plates has rounded tubercles. | 3 |
| 2. Point sclerites strongly tuberculate with leafy distal end; | Imbricacis foliata sp. nov. |
| -. Point sclerites weakly tuberculate with spiny distal end. | Imbricacis miyajimai |
| 3. Point sclerites with distal end wide, not spiny; | Imbricacis hawaiiensis sp. nov. |
| -. Point sclerites with spiny distal end. | 4 |
| 4. Stout calycular rods present, some calycular sclerites with smooth distal end; | Imbricacis squamata |
| -. No calycular rods and no calycular sclerites with smooth distal end. | Imbricacis ijimai |
Imbricacis ijimai (Kinoshita, 1909)
Acis ijimai Kinoshita, 1909: 8, pl. 1 Figure 3, pl. 2 Figures 13–16 (Uji island); Kükenthal, 1924: 158.
Paracis ijimai Kükenthal, 1924: 158.
Not Paracis cf. ijimai Aurivillius, 1931: 146, Figure 28, pl. 3 Figure 6 (Okinose Bank, Misaki, Sagami Bay).
Not Paracis squamata Bayer 1949: 198, Figure 6a–e, pl. 1 Figure 1 (Bikini (USNM44077)); Muzik 1979: 62 (Philippine (USNM49909)).
Material examined. Holotype UMUTZ-G097 Acis ijimai Coral ground of Uji Is., Kagoshima Pref., Japan, East China Sea, depth 80–90 fathoms (146–164 m), Satsuma Expedition, June 1908, coll. Kumao Kinoshita; syntype USNM73492 Acis ijimai same data as UMUTZ-G097; other materials USNM44077 previously identified as Paracis squamata by Bayer 1949, Ralik Chain, Bikini Atoll, Off Enyu Pass, Marshall Islands, North Pacific Ocean, 11.4731′ N, 165.526′ E, depth 213–221 m, Bikini Scientific Resurvey, 1947, st. 30, coll. Russell, R. D., 22 August 1947; USNM49909 previously identified as Paracis squamata by F. M. Bayer and Muzik 1979, Alb-USFC/A5398A, Gigantangan Island, between Masbate and Leyte Islands, Samar Sea, Philippines, 11.5867 N, 124.23 E, depth 208 m, R/V Albatross Philippines Expedition, Trawl–Agassiz beam, 12 ft, with mud bag, 15 March 1909; USNM77282 Kermadec Islands, NewZealand, South Pacific Ocean, 30.63′ S, 178.52′ W, depth 219 m, R/V Viti, Leeward Cruise, st. C530, 19 September 1960, det. Bayer; USNM1439589-2 Tinian Channel, Tinian Island, Northern Mariana Islands, North Pacific Ocean, 14.917′ N, 145.615′ E, depth 116–146 m, R/V Panglou Oro, st.15, 27 October 1973, coll. Grigg, R. W.; Eldredge, L. G.; AKM162, Hirase, Okinawa, Japan, depth ca.130 m, R/V Tansei-maru cruise KT02-03, St. A-5, 1 m Dredge, 15 April 2002; AKM550 Off Takato, Tachibana-ura, Otsuki, Tosa, Japan, 132°48.06′ E, 32°43.08′ N–132°47.68′ E, 32°43.12′ N, Kiryo-maru, st.3, depth 84.75–83.1 m, 7 October 2004; AKM1214 Off Yokoate-jima Is. Tokara Isls., Okinawa, Japan, 129°04.09′ E, 28°54.90′ N, depth 395 m, R/V Tansei-maru cruise KT07-21, st. DY205, 30 August 2007, coll. Yokose; AKM1221, Off Takarajima Is. Okinawa, Japan, 129°10.43′ E, 29°05.29′ N, depth 334 m, R/V Tansei-maru cruise KT07-21, st. DT203-1, 31 August 2007, coll. Yokose; AKM1389, West of Chichijima Is. Ogasawara Isls., Japan, 142°07.412′ E, 27°01.395′ N–142°07.467′ E, 27°01.360′ N, depth 139–144 m, R/V Tansei-maru cruise KT09-2, st.TW01-01, 19 March 2009, coll. A.K. Matsumoto; AKM1475, West of Chichijima Is. Ogasawara Isls., Japan, 142°07.466′ E, 27°01.371′ N–142°07.489′ E, 27°01.344′ N, depth 135–136 m, R/V Tansei-maru cruise KT09-2, st.TW01-02, 19 March 2009, coll. A.K. Matsumoto.
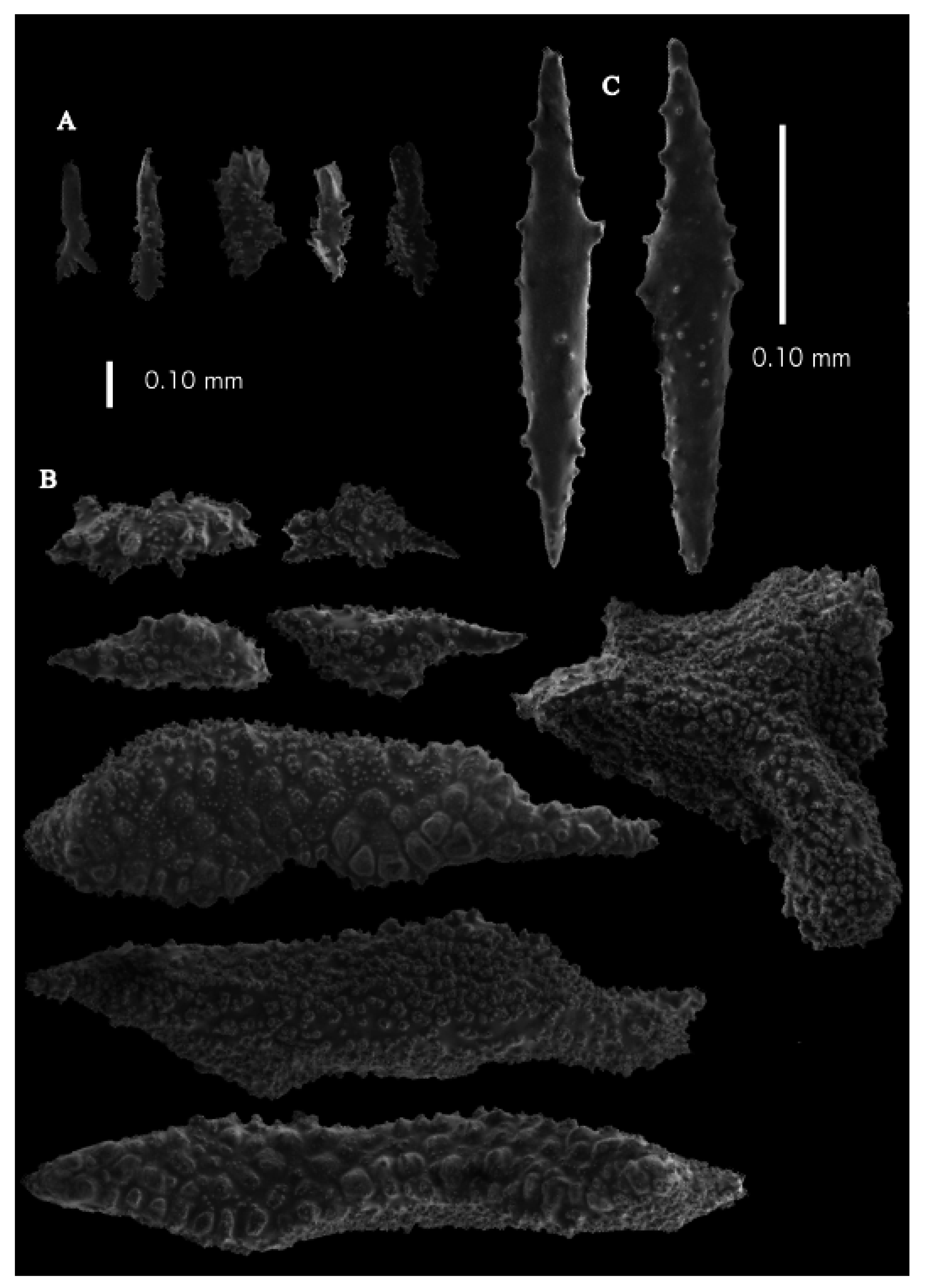
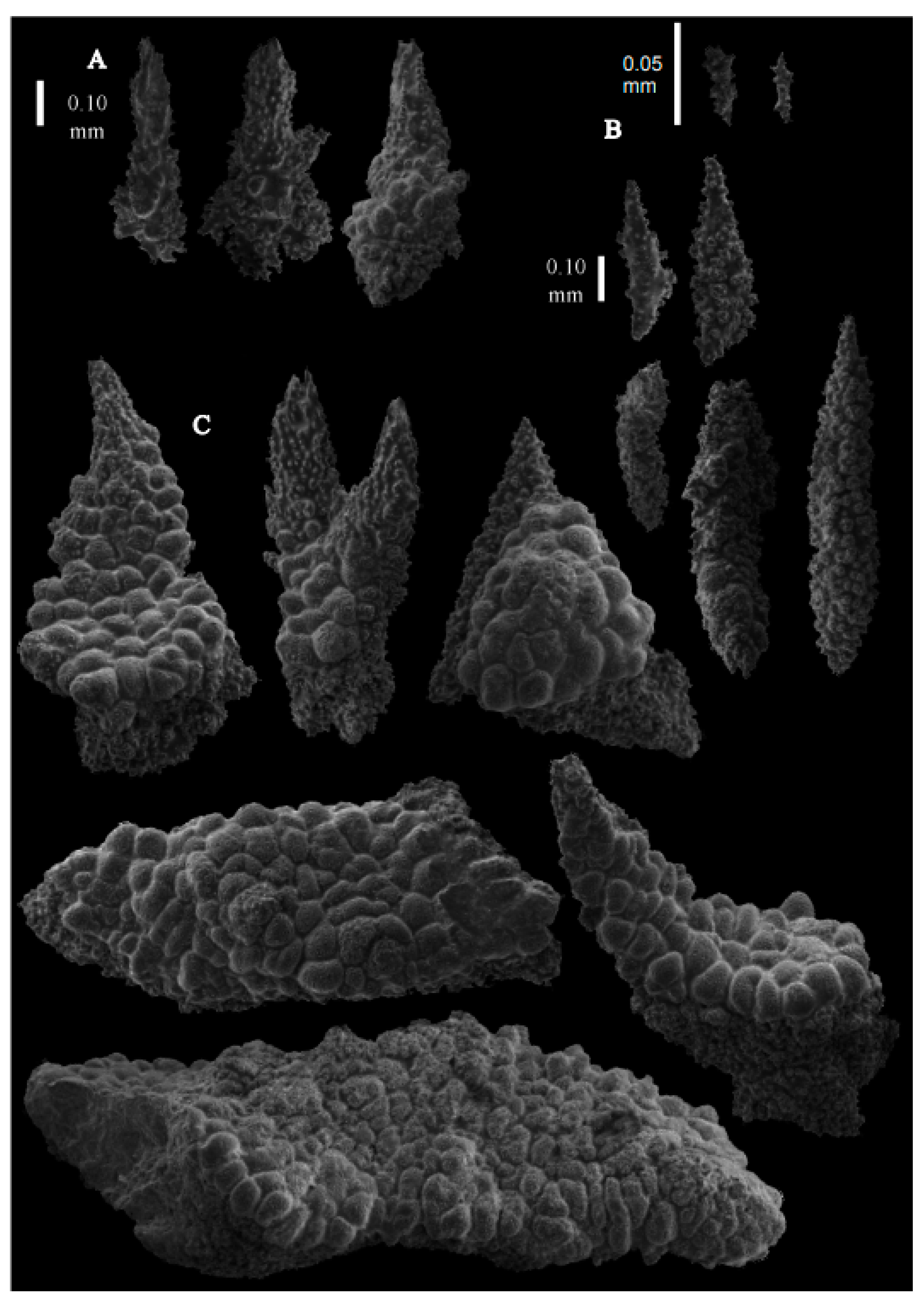
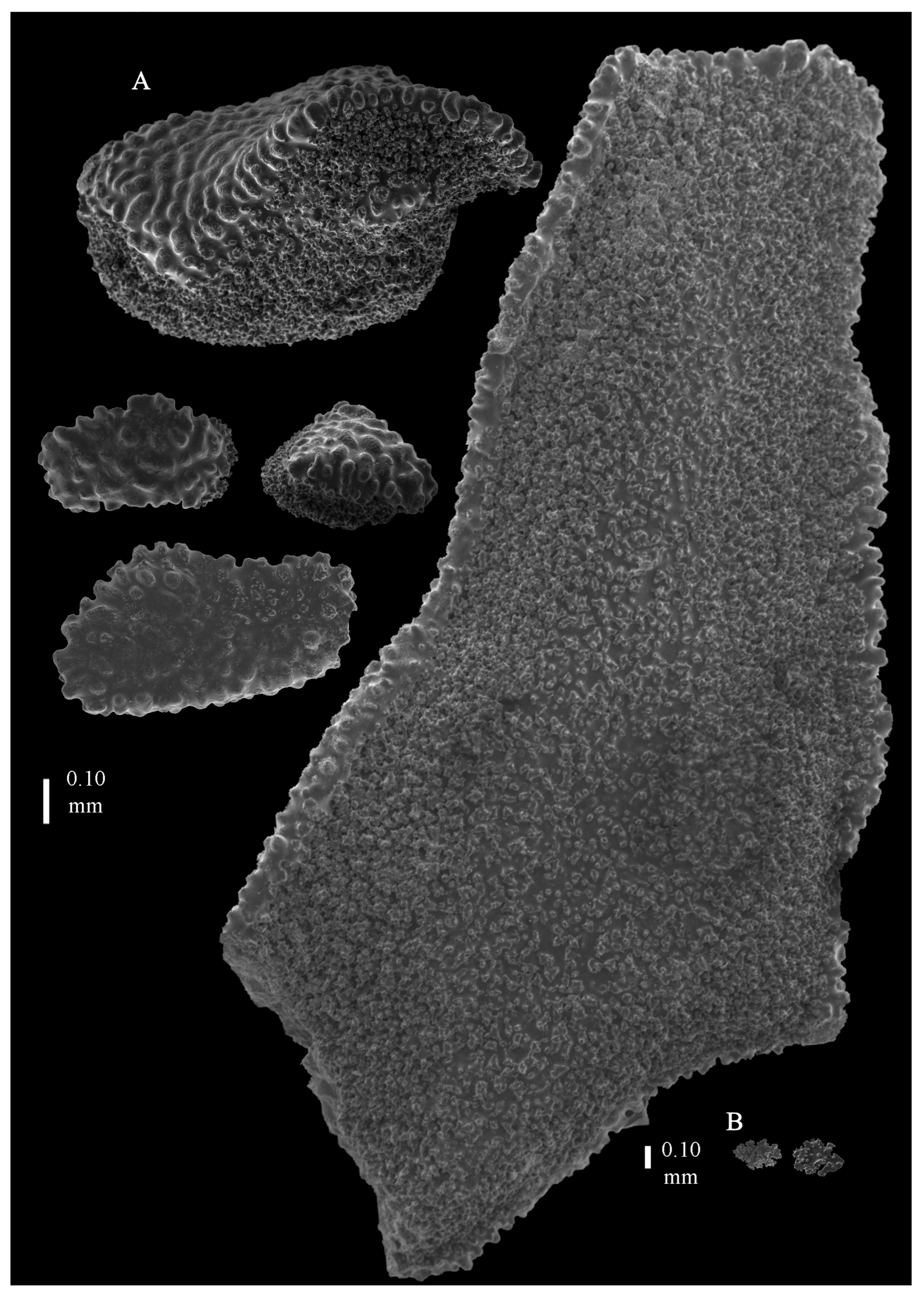
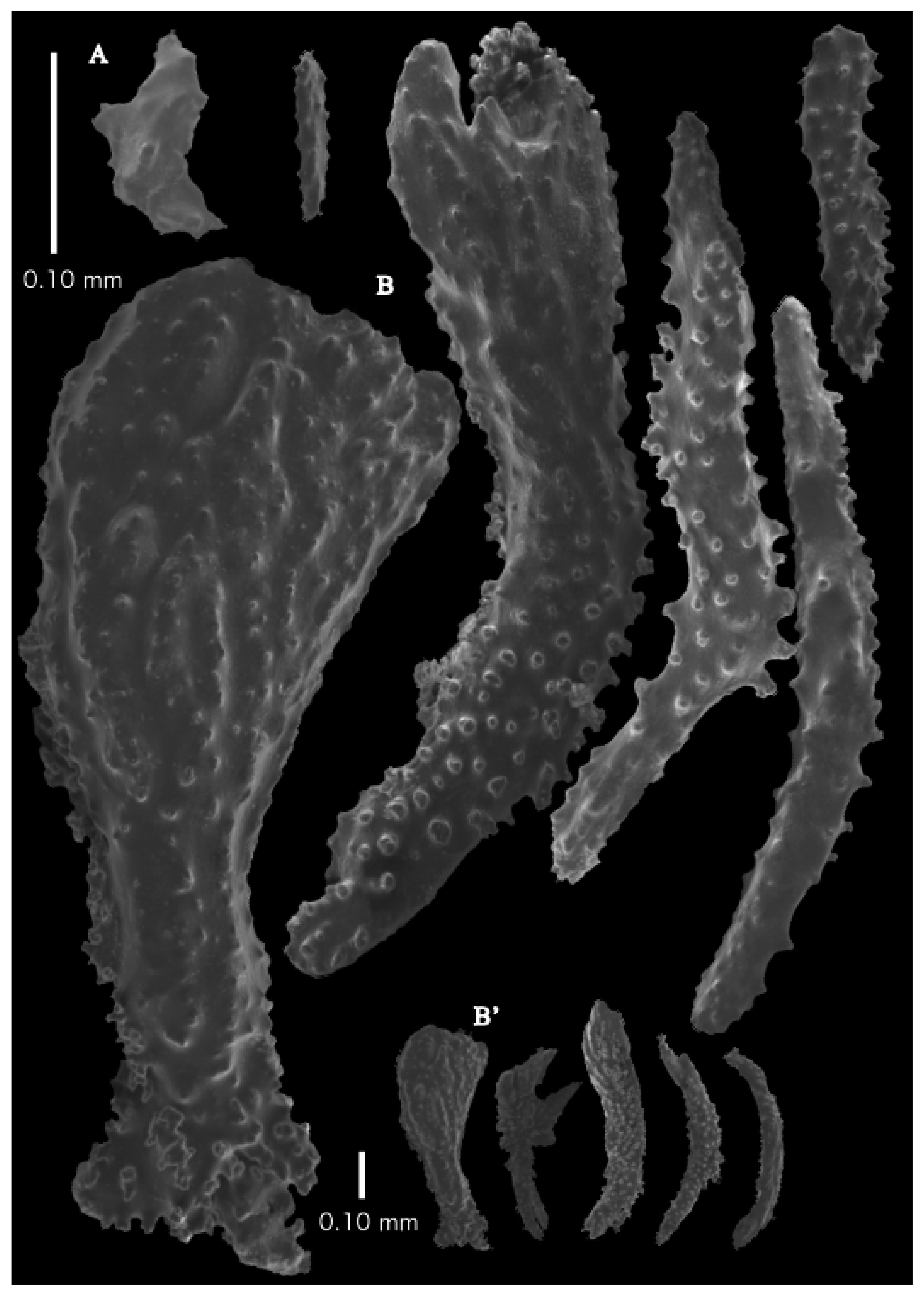
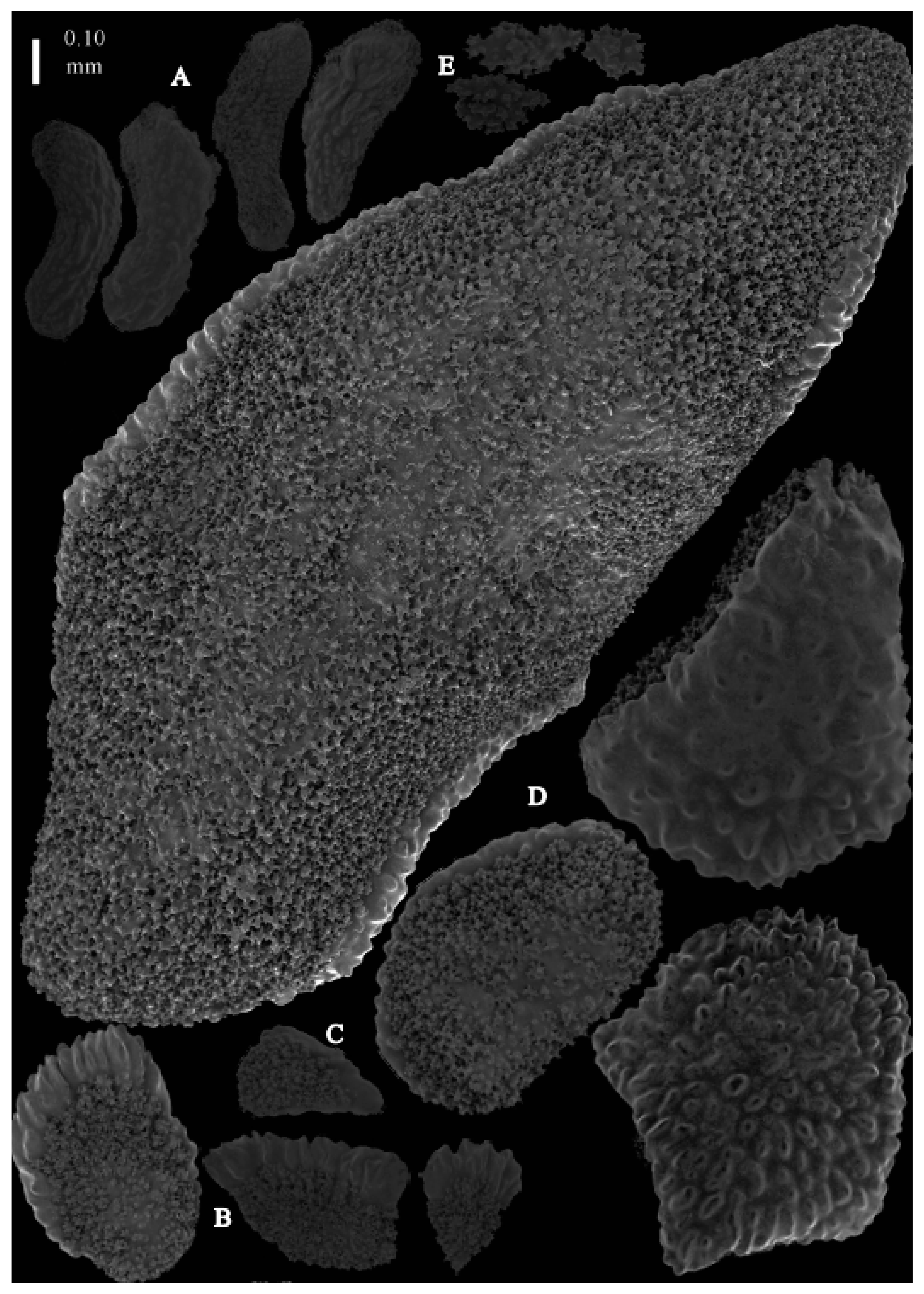
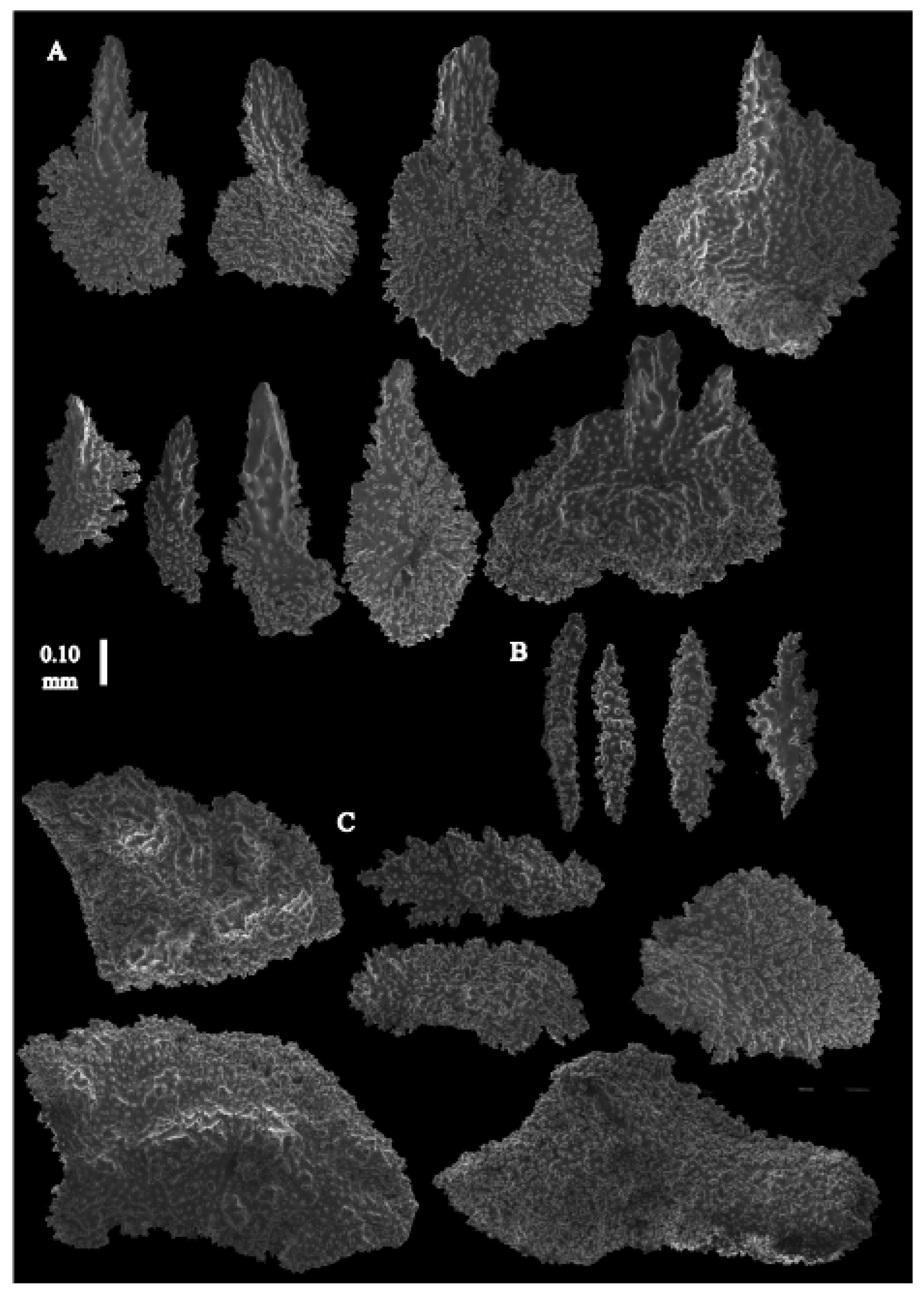
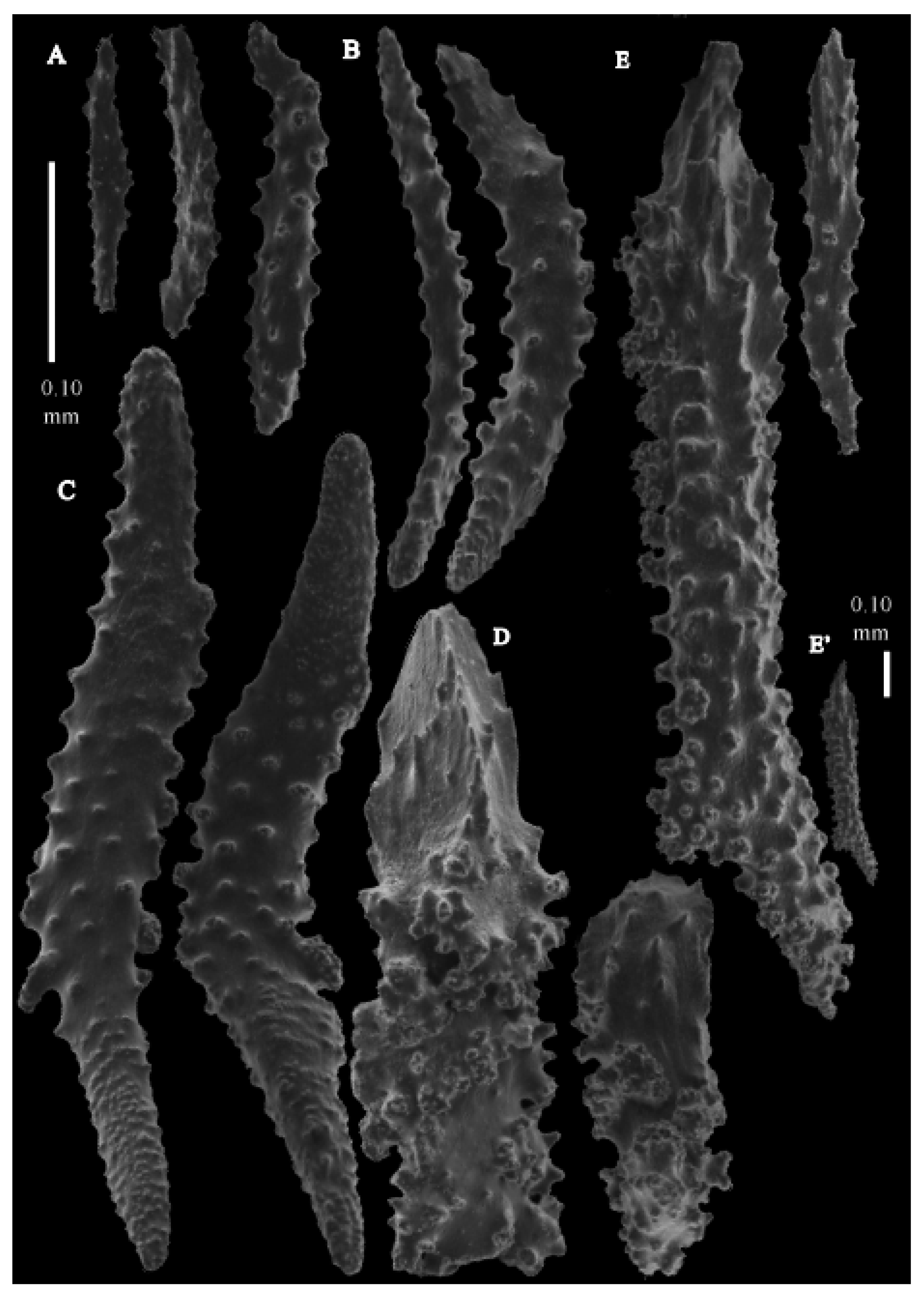
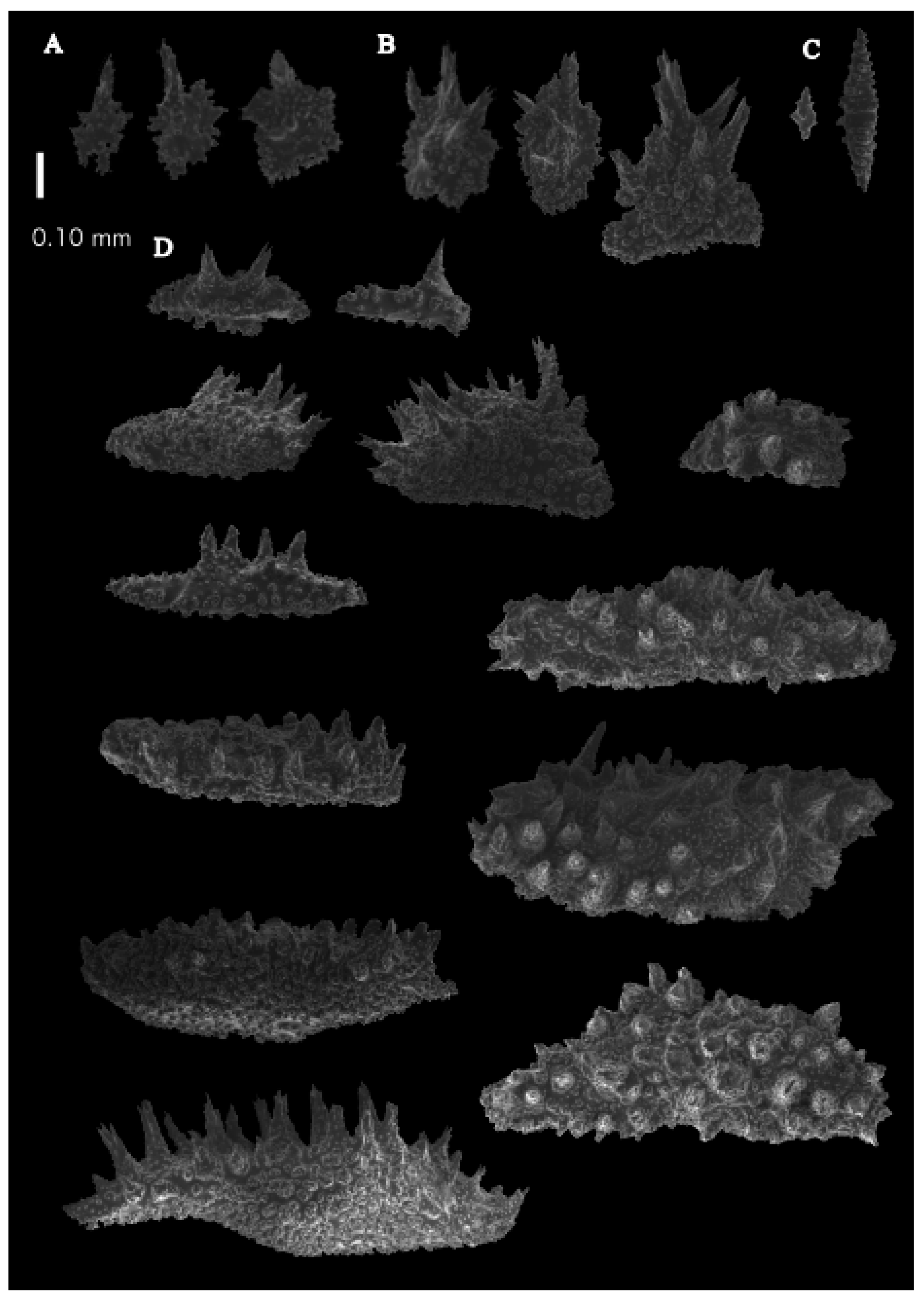
Re-description. The holotype (UMUTZ-G097) is 5.3 cm high and 9.5 cm wide, branched in one plane (
Figure 7B). Stem is 2 mm in diameter and the branches are 1.5 mm in diameter. Calyces are conical, situated on three sides on the main branches, in two rows on the terminal branches. They are separated 1–2 mm from each other (
Figure 19A).
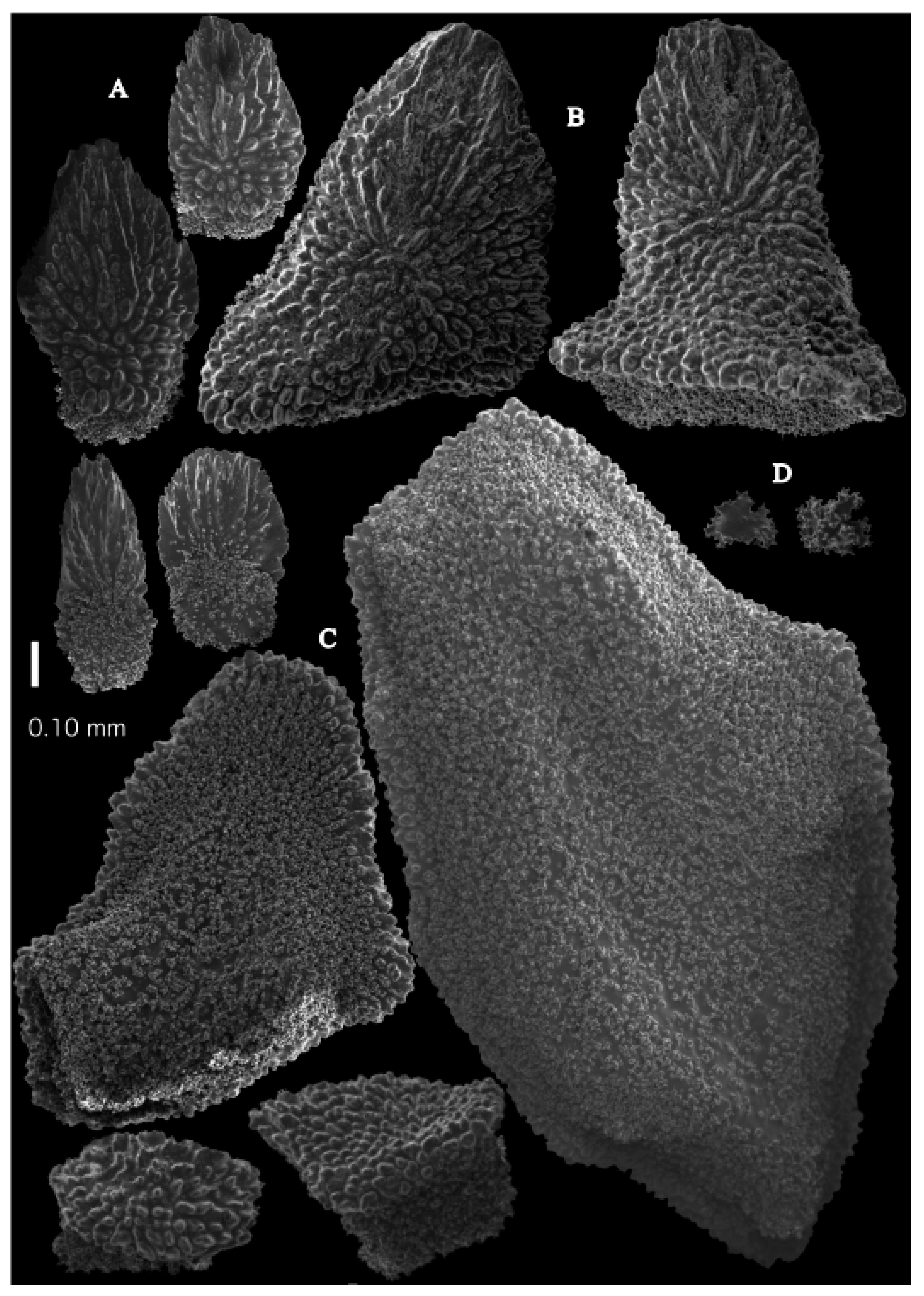
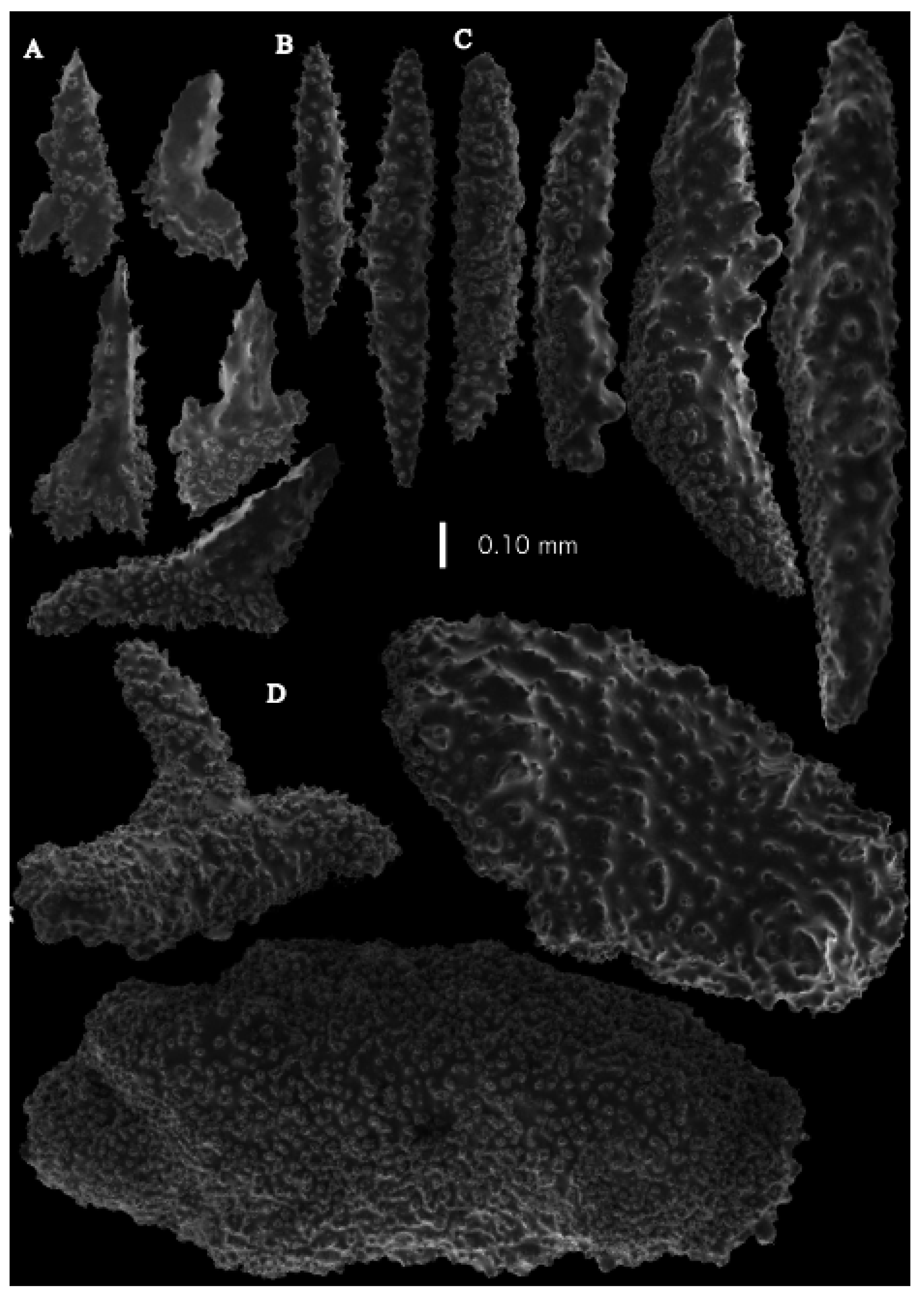
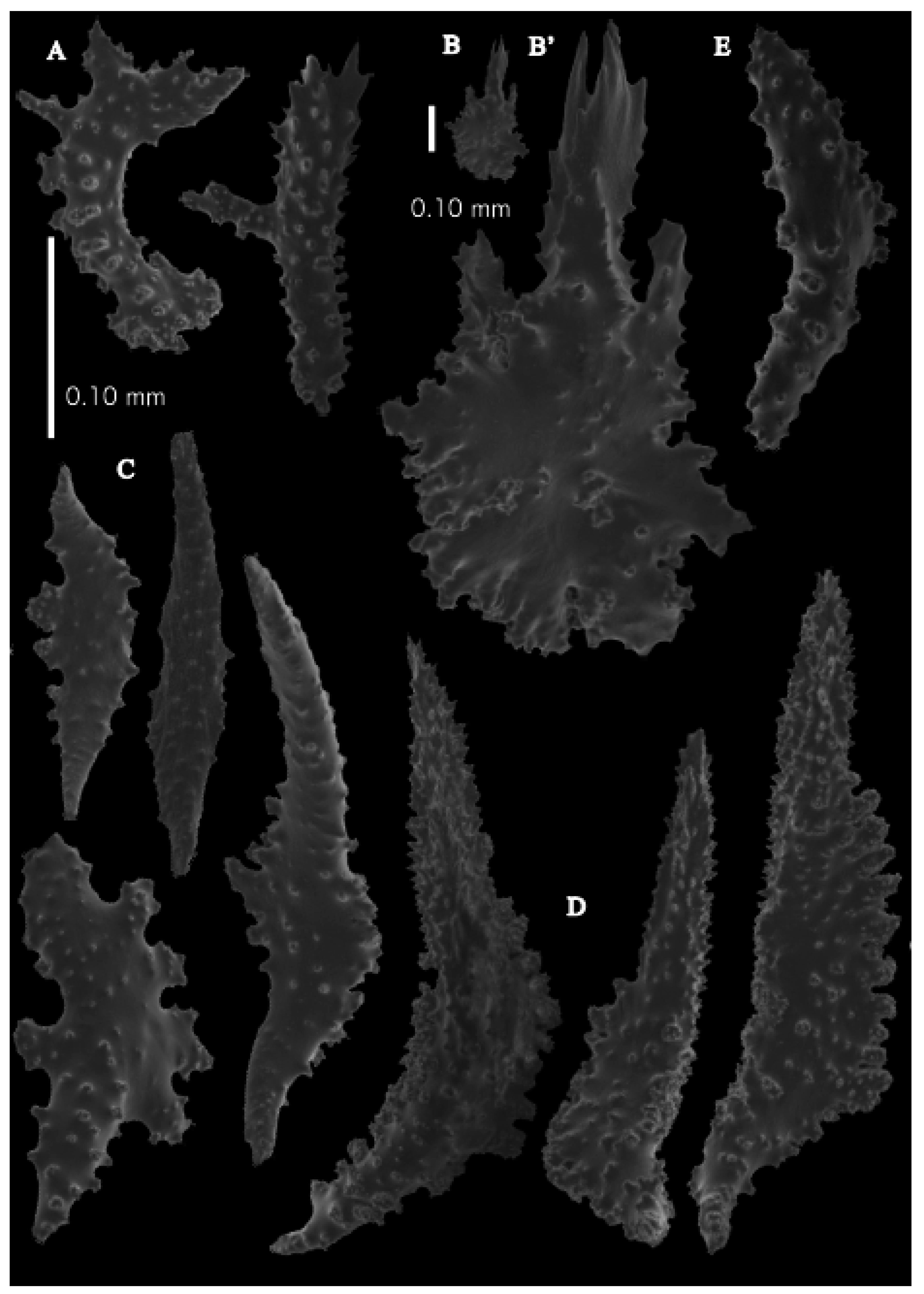
The polyps have large bent spindles, up to 0.6 mm long (
Figure 20B), the upper part with ridges and the lower part with simple tubercles. Smaller tentacle spindles, 0.10–0.15 mm long, have simple tubercles only (
Figure 20A). The calyces are covered in plates (
Figure 21A,B), the distal ones scale-like with rounded tubercles and ridges distally (
Figure 21A), others with rounded tubercles only (
Figure 21B). The calycular plates are 0.3–0.8 mm long. The interior of the plates is granular. The surface layer of the branches with large plates forming a mosaic. The outer side of the plates has rounded tubercles, the inner side is granular (
Figure 21C). These plates are up to 4 mm long. The interior of the coenenchyme has irregular-shaped sclerites with simple and complex tubercles (
Figure 21D). These sclerites are up to 0.4 mm long.
Colours. Colony white with brownish polyps. Anthocodial sclerites colourless. Coenenchymal sclerites white.
Variation. Muzik [
9] mentioned that the Bikini specimen (USNM44077 (
Figure 7C,
Figure 22 and
Figure 23)) and
Alb -Philippine specimen (USNM49909 (
Figure 7D,
Figure 24 and
Figure 25)) are different from each other, and the Philippine specimen resembles
P. squamata. We interpret these differences as an intraspecific variation because USNM49909 lacks the pronounced ridges on the calycular sclerites (Figure 35B) and peculiar rod-like calycular sclerites (Figure 35A) that are diagnostic of
I. squamata.
Distribution: Japan, Northern Mariana Islands, Philippines: Northwest Pacific; Kermadec Islands: South Pacific.
Remarks. The smooth rods of the pinnules mentioned by Kinoshita [
1] were not observed by us. Though we did not re-examine the material of
Paracis cf.
ijimai Aurivillius [
6], according to the following reasons, we consider that it probably is a species of
Pseudoparacis gen. nov. Aurivillius described that (1) it has a collaret which is not present in
Imbricacis gen.nov. and (2) it does not have the dragon wing sclerites which are present in
Neoacis. He also mentioned that (3) the polyps of
P. ijimai are distributed all around the stem, but
I. ijimai has polyps distributed around three sides of the main branches.
Imbricacis foliatasp. nov.
urn:lsid:zoobank.org:act:B367267A-1979-47A0-B458-2D1F9BF544CC
Material examined. Holotype USNM89348, 60 Naut. Mile NE of Heard Island, Indian Ocean, depth 500 m, coll. date unknown.
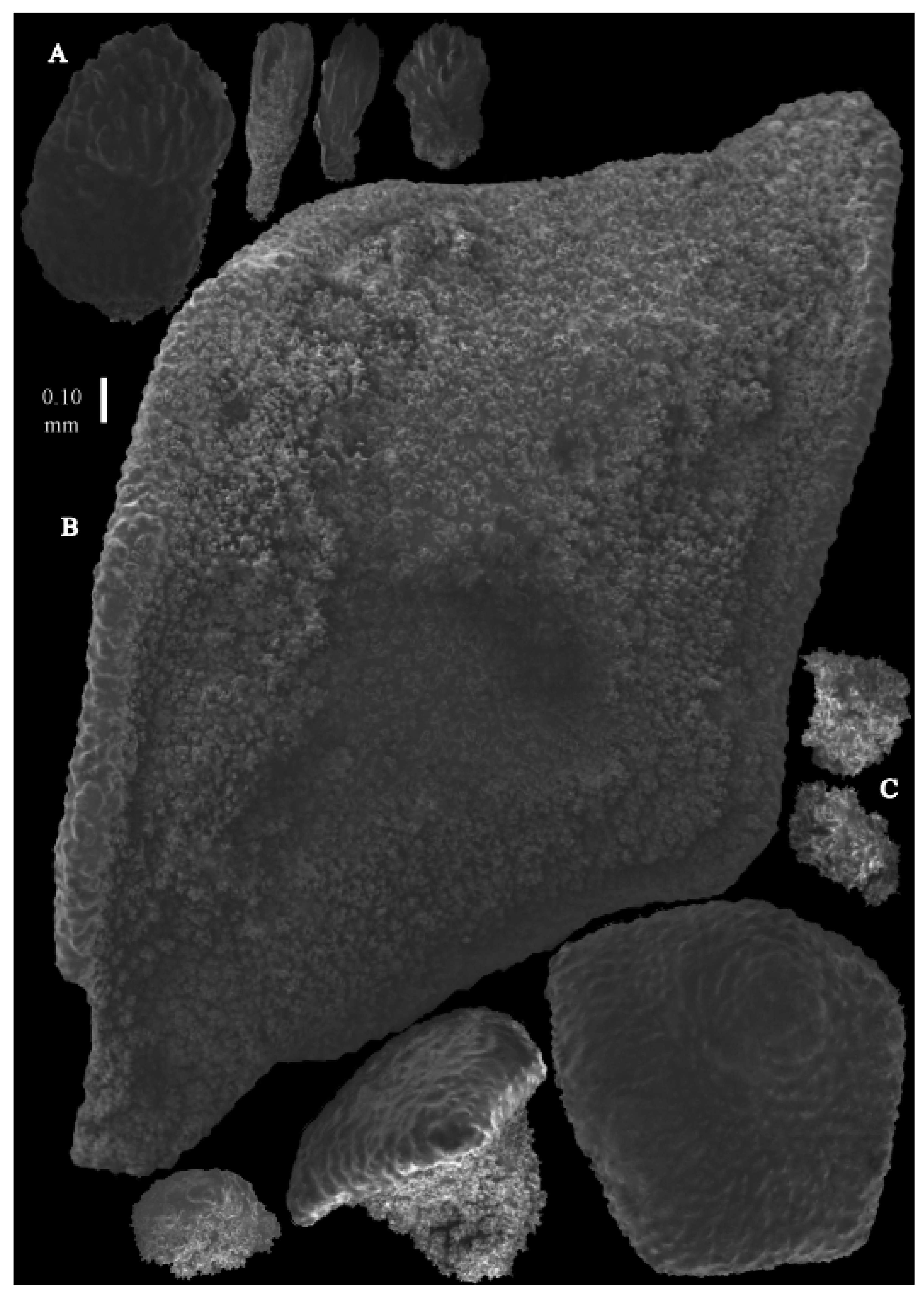
Description. The holotype is 4.6 cm high and 1.8 cm wide, branched in one plane (
Figure 26A). Stem is 2.1 mm in diameter. Stem and larger branches flattened in a plane perpendicular to that of the branches. Terminal branches cylindrical, 1.6 mm in diameter. Calyces are conical, situated on three sides of the branches, closely set to each other (
Figure 19D). The polyps have large bent spindles, up to 0.5 mm long. The upper part has ridges, some leafy, the lower part with simple and complex tubercles (
Figure 27B,B’). Tentacle spindles, 0.10–0.20 mm long, have simple tubercles only (
Figure 27A). The calyces are covered in plates, the distal ones with distal ridges (
Figure 27A). The plates are 0.4–0.6 mm long. The interior of all plates is granular. The surface layer of the branches with large plates forming a mosaic. The outer side of the plates is smooth, the inner side is granular (
Figure 28B). These plates are up to 5 mm long. The interior of the coenenchyme has spindles with complex tubercles (
Figure 28C). These sclerites are up to 0.4 mm long.
Colour. Colony white. Anthocodial sclerites colourless. Coenenchymal sclerites white.
Distribution. The species is only known from the type of locality, off Heard Island, Indian ocean.
Etymology. From the Latin folia (leaf), referring to the presence of leafy distal ends of the polyp sclerites that distinguish it from I. miyajimai.
Remarks. The species differs from I. miyajimai, the other species with coenenchymal plates with a smooth outer surface, in having point sclerites with a leafy distal end.
Imbricacis hawaiiensis sp. nov.
urn:lsid:zoobank.org:act:72D8495E-BCD9-4B80-B666-99437DF20052
New genus species A: Muzik, 1979: 56-62, Pls. VII, VIII.
Material examined. Holotype, BPBM-D529, Makapuu, depth 1200 ft (=365.76 m), coll. R. Grigg, id: KMM as new genus, new species, 1977; paratype, USNM1010725 Kahoolawe, Maui County, Hawaii, North Pacific Ocean, 20.4863′ N, 156.66′ W, depth 403 m, DSR/V Pices 4, KOK/Pices 4 cruise, Dive P4-047, 5 September 2002, coll. Hawaii Undersea Research Laboratory.
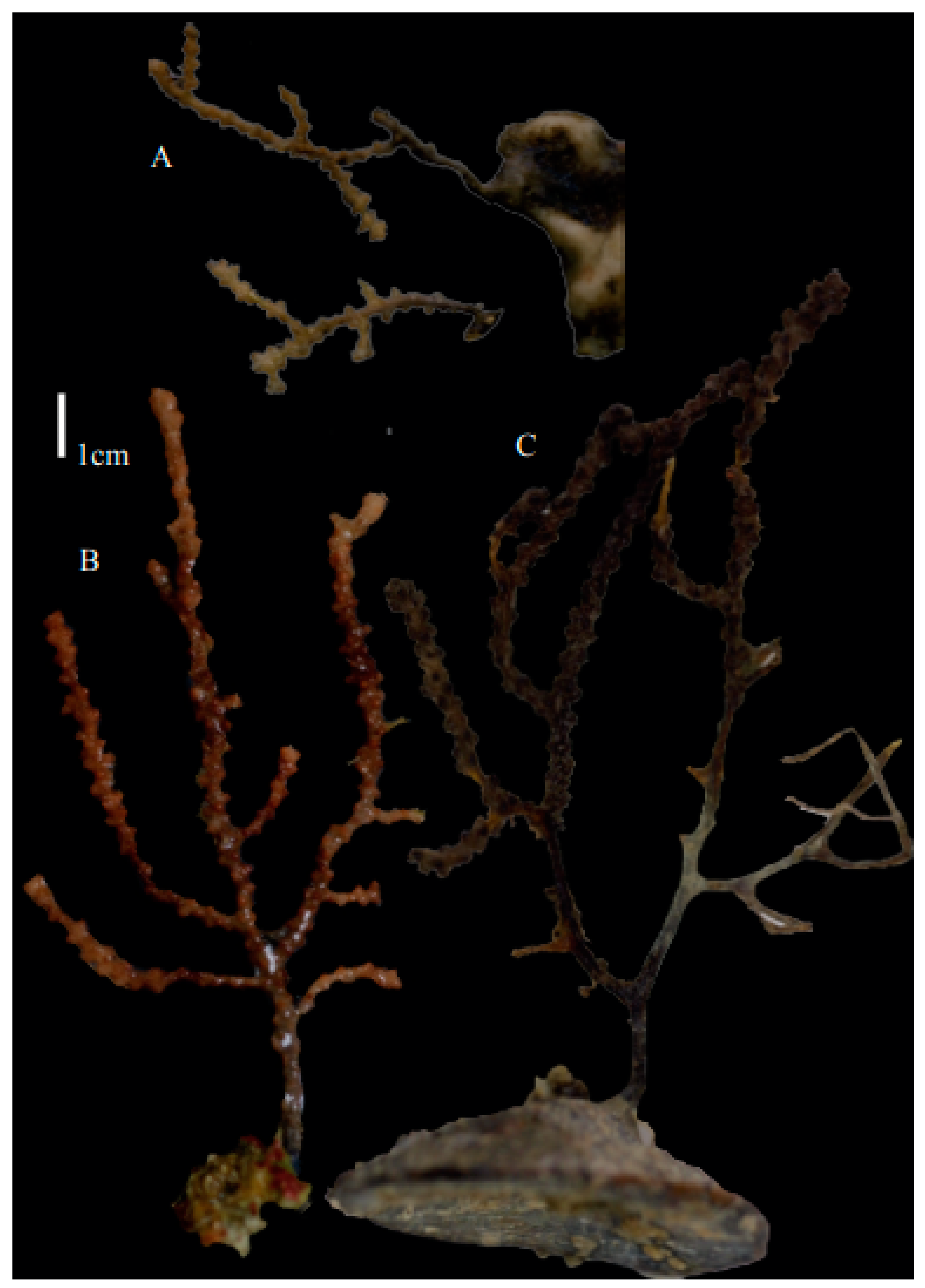
Description. The holotype (BPBM-D529) is 20 cm long and 7 cm wide, holdfast included (
Figure 26B). Stem is 6.8 mm in diameter, and branches are 3 mm in diameter. Calyces are conical, situated on three sides on the main branches, in two rows on the terminal branches. They are separated 2.8–3.7 mm from each other (
Figure 19E). The polyps have large bent spindles, up to 0.55 mm long, with simple tubercles and widened distal end (
Figure 29B,B’). Smaller tentacle spindles, 0.10–0.20 mm long, have simple tubercles only (
Figure 29A). The calyces are covered in plates (
Figure 30A), the distal ones are scale-like with rounded tubercles and ridges distally. The plates are 0.30–0.50 mm long. The interior of the plates is granular. The surface layer of the branches with large plates forming a mosaic (
Figure 30B). The outer side of the plates has rounded tubercles, and the inner side is granular. These plates are up to 7 mm long. The interior of the coenenchyme has irregular-shaped sclerites with simple and complex tubercles (
Figure 30C). These sclerites are up to 0.30 mm long.
Colour. Dried colony looks to be of a dark colour, but the original colony was white. Anthocodial sclerites colourless. Coenenchymal sclerites white.
Variation. Paratype (USNM1010725) is an intact colony, 26.4 cm long and 17.8 cm wide, holdfast included (
Figure 19F and
Figure 26C).
Distribution. The species is only known from the type locality, Hawaii, North Pacific.
Etymology. The species is named after its type locality, Hawaii.
Remarks. The species differs from I. ijimai and I. squamata, the two other species with rounded tubercles on the outer side of the coenenchymal plates, in lacking point sclerites with distal spiny ends. The other materials from Hawaii in USNM (98805, 98806, 1071439), which have been previously identified as Paracis cf. squamata by F. M. Bayer on 1 May 1997 or Paracis miyajimai, we have not examined, but they could also be this species.
Imbricacis miyajimai (Kinoshita, 1909)
Acis miyajimai Kinoshita, 1909: 10, Pl. 1 Figures 4–6, pl. 2 Figures 17–20 (off Province of Satsuma).
Paracis miyajimai Kükenthal, 1924: 158.
Material examined. holotype UMUTZ-G211 Acis miyajimai Coral ground of Kagoshima Pref., Japan, East China Sea, depth unknown. Obtained 1 July 1898. Bought at Kataura market, Kagoshima Pref. Japan by Mikinosuke Miyajima. Dry.
Re-description. The dry holotype is 70 cm high and 50 cm wide, branched in one plane (
Figure 26D). Most of the coenenchyme is lost. Stem is 2 cm in diameter. Stem and larger branches flattened in a plane perpendicular to that of the branches. Terminal branches cylindrical, 1.5 mm in diameter. Calyces are conical, situated on three sides of the branches, closely set to each other (
Figure 19G). The polyps have large bent spindles, up to 0.6 mm long. The upper part is rounded and the lower part has simple tubercles (
Figure 31B). Tentacle spindles, 0.10–0.20 mm long, have simple tubercles only or are almost smooth (
Figure 31A). The calyces are covered in plates (
Figure 32A), the distal ones are granular but smooth distally. The plates are 0.3–1 mm long. The interior of all plates is granular. The surface layer of the branches has large plates forming a mosaic. The outer side of the plates is smooth, but the inner side is granular (
Figure 32B). These plates are up to 3 mm long. The interior of the coenenchyme has spindles with simple tubercles (
Figure 31C,C’). These sclerites are up to 0.4 mm long. The interior of the coenenchyme also has irregular-shaped sclerites with simple and complex tubercles (
Figure 32C). These sclerites are up to 0.19 mm long.
Colours. Dried colony looks to be of a dark colour. Sclerites are all white.
Distribution. Only known from type of locality.
Remarks. This species differs from I. ijimai, the most widespread Japanese species, by its very large colony size and quite solid and smooth sclerites on the surface.
Imbricacis squamata (Nutting, 1910)
Acis squamata Nutting, 1910: 42, pl. VII Figure 2, 2a, pl. XX Figure 50 (Kei islands Indonesia, SIBOGA st. 253); Thomson and Dean 1931: 200, pl. 5, Figure 2, pl. 16, Figure 9.
Paracis squamata; Kükenthal 1924: 158, Figure 108, 109; Aurivillius 1931: 145 (in Key only); van Soest 1979: 92.
? Acis squamata Nutting, 1912: 81 (Japan).
NOT Acis squamata Nutting, 1910: 42, st. 65A (Kei islands), st. 305 (off Kampong Menanga).
Material examined. lectotype, ZMA Coel. 2142b (syntype on the label) SIBOGA st. 253, 5°48′.2 S., 132°13′ E. near Kei Islands, depth 304 m, clay; non-types ZMA Coel. 2143 SIBOGA st. 105, 6°8′ N., 121°19′ E. Sulu Sea, 275 m. Coral. bottom; ZMA Coel. 2144, SIBOGA st. 251, 5°28.4′ S., 132°0.2′ E. Kei Islands, depth 204 m, Harter coral sand. 1 Ex.
Re-description. The lectotype (ZMA Coel. 2142b) is 10 cm long and 3 cm wide, holdfast included (
Figure 33A). Stem is 5 mm in diameter, and the branches are 2 mm. Calyces are conical, situated on three sides on the main branches, in two rows on the terminal branches. They are separated 1–2 mm from each other (
Figure 19H). The polyps have large bent spindles, up to 0.40 mm long, the upper part with ridges and the lower part with simple tubercles (
Figure 34B,B’). Smaller tentacle spindles, 0.10–0.20 mm long, have simple tubercles only (
Figure 34A). The calyces are covered in plates (
Figure 35); the distal ones are scale-like with rounded tubercles and ridges distally (
Figure 35B), whereas others are with rounded tubercles only or with a smooth distal end (
Figure 35C). Several of these calycular plates are rod-like (
Figure 35A). The plates are 0.30–0.50 mm long. The interior of the plates is granular. The surface layer of the branches with large plates forms a mosaic. The outer side of the plates has rounded tubercles; the inner side is granular (
Figure 35D). These plates are up to 3 mm long. The interior of the coenenchyme has irregular-shaped sclerites with simple and complex tubercles (
Figure 35E). These sclerites are up to 0.25 mm long.
Colours. Colony white with brownish polyps. Anthocodial sclerites colourless. Coenenchymal sclerites white. Live colour of I. squamata is not clear. Nutting mentioned that the SIBOGA material he examined has been longer preserved in alcohol.
Distribution. Kei Islands, Sulu Sea, Indonesia.
Remarks. Nutting [
4] mentioned type specimens from three localities, st. 65A, st. 253, and st. 305. In the RMNH, two jars are present: syntype ZMA Coel. 2142b: Stat. 253. 5°48′.2 S., 132°13′ E. near Kei Islands. 304 m. Clay; the other material, ZMA Coel. 2142a, with the label “cotype” from station 251, not mentioned by Nutting, is a species of
Keroeides. ZMA Coel. 2143 from station 105 (
Figure 33B) and ZMA Coel. 2144 from station 251 (
Figure 33C) are not mentioned by Nutting as types. The material from SIBOGA Station 305 (Mid channel, Solor Strait, off Kampong Menanga, depth 113 m, stony) and station 65A (7°0′ S., 120., 120°34′. 5E. near Saleyer Island, depth 400 m, mud and coral) are not present anymore in the RMNH. In London, a fragment (BMNH1925.7.25.38) from station 305 is present, which also is
Keroeides. The type specimen from station 65A (near Selayar Island, Greater Sunda Islands, Celebes Island, Sulawesi, Indonesia, South Pacific Ocean, 7°0′ S., 120°34.5′ (120.575) E., depth 300–400 m, Mud and coral, 6 May 1899) has been donated to the Iowa State University (van Soest 1979 [
16]) and is now deposited in the Smithsonian Institution (USNM91922); it is not
I. squamata.
We designate here ZMA Coel. 2142b as the lectotype of
Acis squamata (
Figure 33A,
Figure 34 and
Figure 35). The species mostly resembles
Imbricacis ijimai Kinoshita, 1909 regarding colony shape and sclerites. It differs from that species in having more pronounced ridges on the calycular sclerites (
Figure 31B), while some calycular sclerites have a smooth distal end. It also differs in having several peculiar rod-like calycular sclerites (
Figure 31A). Though Nutting [
5] reported
Acis squamata of
Alb-USFC/A4936, we could not find the specimen in the Smithsonian (USNM) and could not examine it.
Genus PSEUDOPARACIS gen. nov.
urn:lsid:zoobank.org:act:C7C763FA-DAE2-49A0-A687-E412D0158E1B
Type species. Acis spinifera Nutting, 1912.
Diagnosis. Colonies branched in one plane. Coenenchyme with large polygonal thick plates. Calycular sclerites are thornscales. Points with spindles with upper end spiny and lower end tuberculate. Collaret with bent spindles, more tuberculate in the middle part. Tentacles with rods.
Etymology. A combination of Pseudo (Greek for false) and the genus Paracis.
Remarks. The genus mostly resembles Paracis but differs in lacking dragon wing sclerites in the tentacles and rounded tubercles on the outside of the coenenchymal plates.
Key to the species of Pseudoparacis (three species)
| 1. Coenenchymal plates with spines and ridges and small range of sizes, thornscales with blunt spine, polyps all around at the tip. | P. japonica |
| -. Coenenchymal plates without spines or ridges and large range of sizes, thornscales with sharp/blunt spine. | 2 |
| 2. Coenenchymal plates with rounded elevations, thornscales with blunt spine, polyps on three sides of branches. | P. tanseii sp. nov. |
| -. Coenenchymal plates with tubercles only, thornscales with sharp spine, polyps all around branches. | P. spinifera |
Pseudoparacis spinifera (Nutting, 1912)
Acis spinifera Nutting, 1912: 82, pl. 13 Figures 2, 2a, pl. 21 Figure 1.
Paracis spinifera; Kükenthal, 1924: 161 Figure 110.
Stenella horrida Kükenthal, 1924: 304.
Paracis horrida; Muzik, 1979: 45, pl. III, IV, USNM56845, USNM56849 (Hawaii).
?Paracis cf. ijimai; Aurivillius, 1931: 146, Figure 28, pl. 3 Figure 6 (Sagami Misaki, Okinose).
Not Stenella horrida Thomson and Henderson, 1906:36 pl. V, Figure 13, pl. IX, Figure 3.
? Paracis horrida; Aurivillius, 1931: 145.
Material examined. Holotype USNM 30088, Alb-USFC/A4936, 54°40′ N 130°37′30 E, Sata Misaki Light, 5.7 miles, off Kagoshima prefecture, North Pacific Ocean, 103 fathoms (188 m), Northwestern Pacific Expedition, 16 August 1906; other materials: USNM49341, Alb-USFC/A3742, 35°02′ N 139° 50′ E, Suno Saki, Chiba Prefecture, Sagami Bay, Honshu Island, Japan, North Pacific Ocean, depth, 104–161 m, Trawl–Blake Beam, 5.5 ft, coll. 19 May 1900, id by Bayer, dry; RMNH.COEL.47988 (AKM554) Off Takato, Tachibana-ura, Otsuki, Tosa, Japan, 132°48.06′ E, 32°43.08′ N–132°47.68′ E, 32°43.12′ N, Kiryo-maru, st.3, depth 84.75–83.1 m, 7 October 2004.
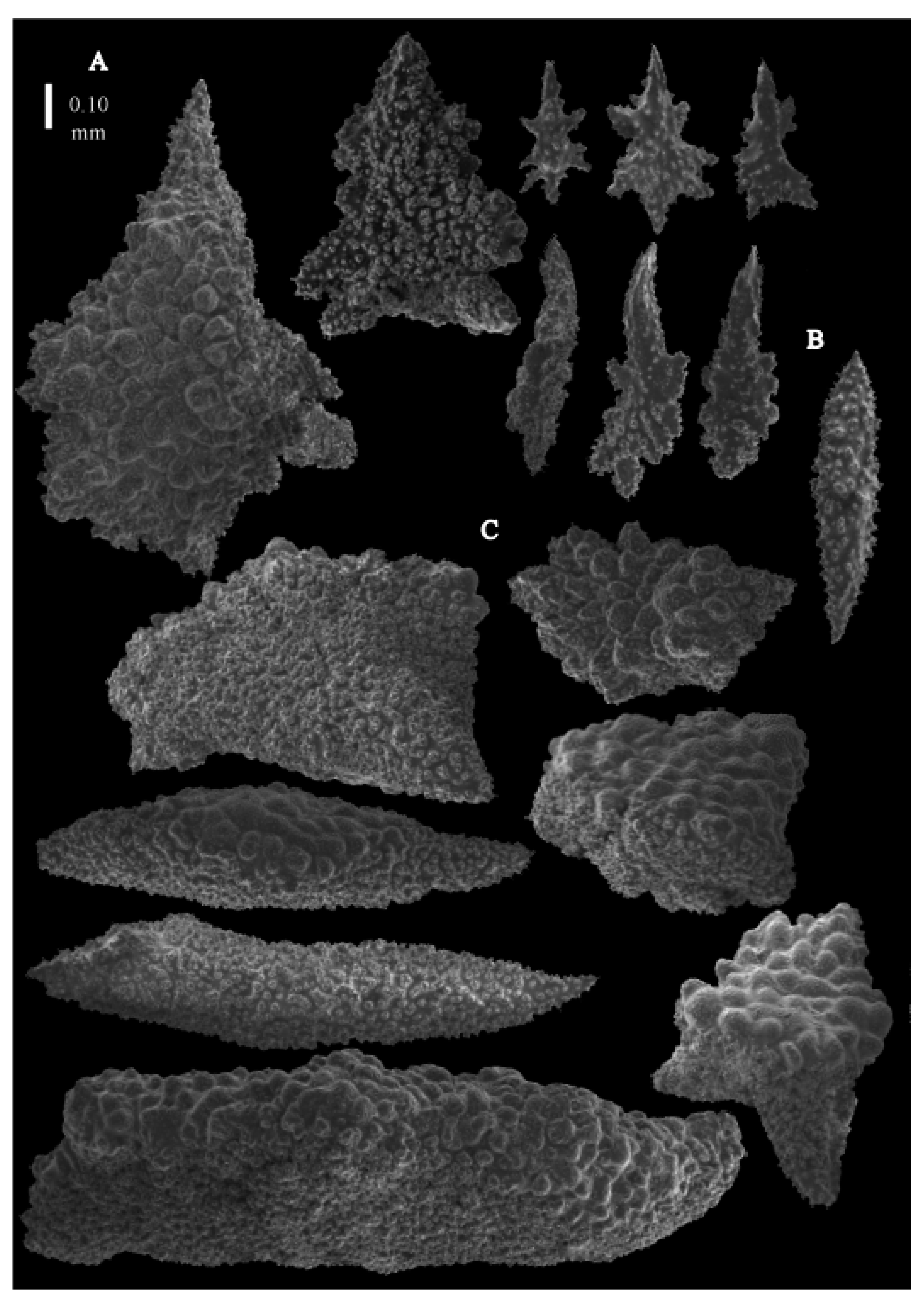
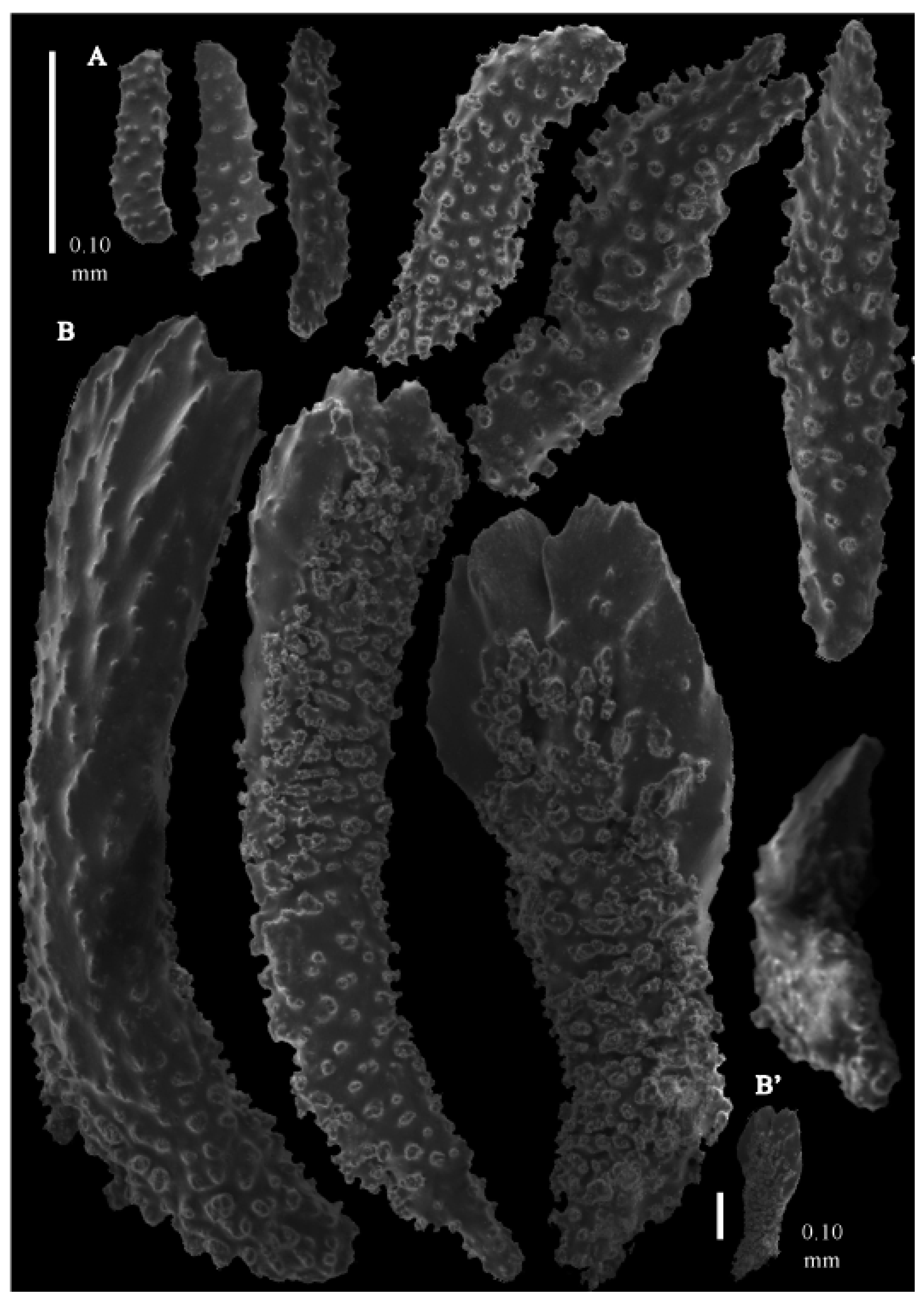
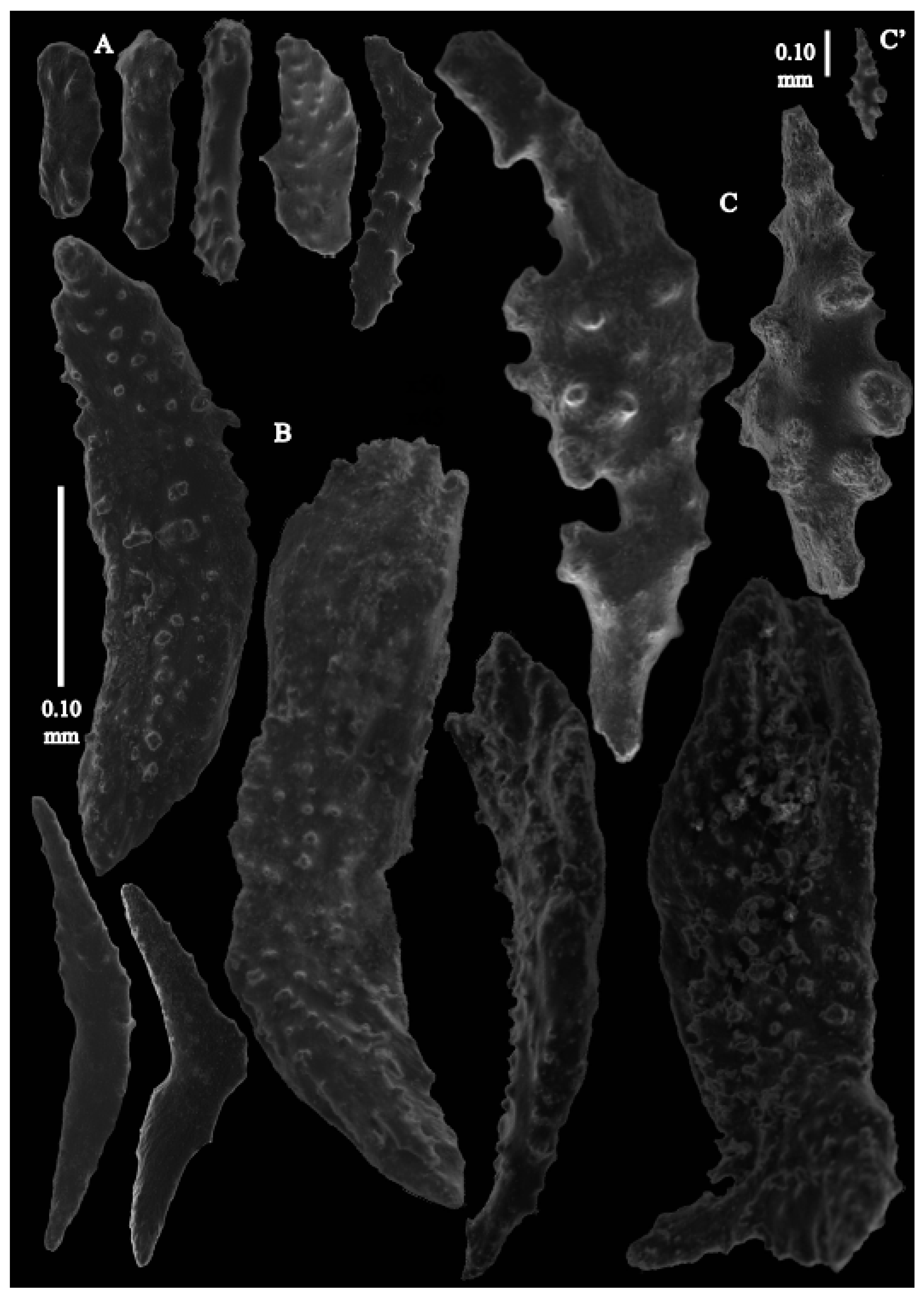
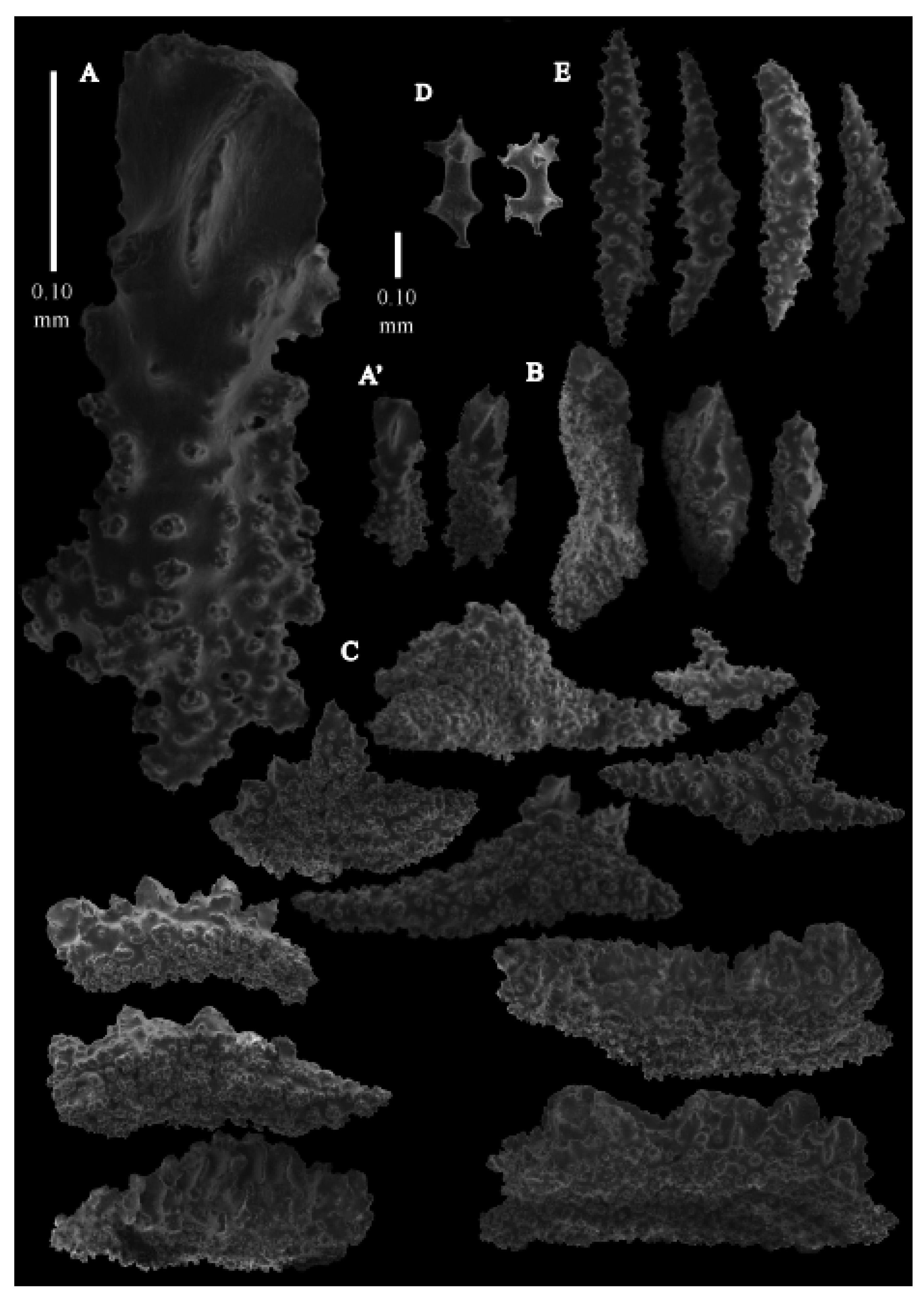
Re-description. Colony 9.6 cm high and 10 cm wide, branched in one plane (
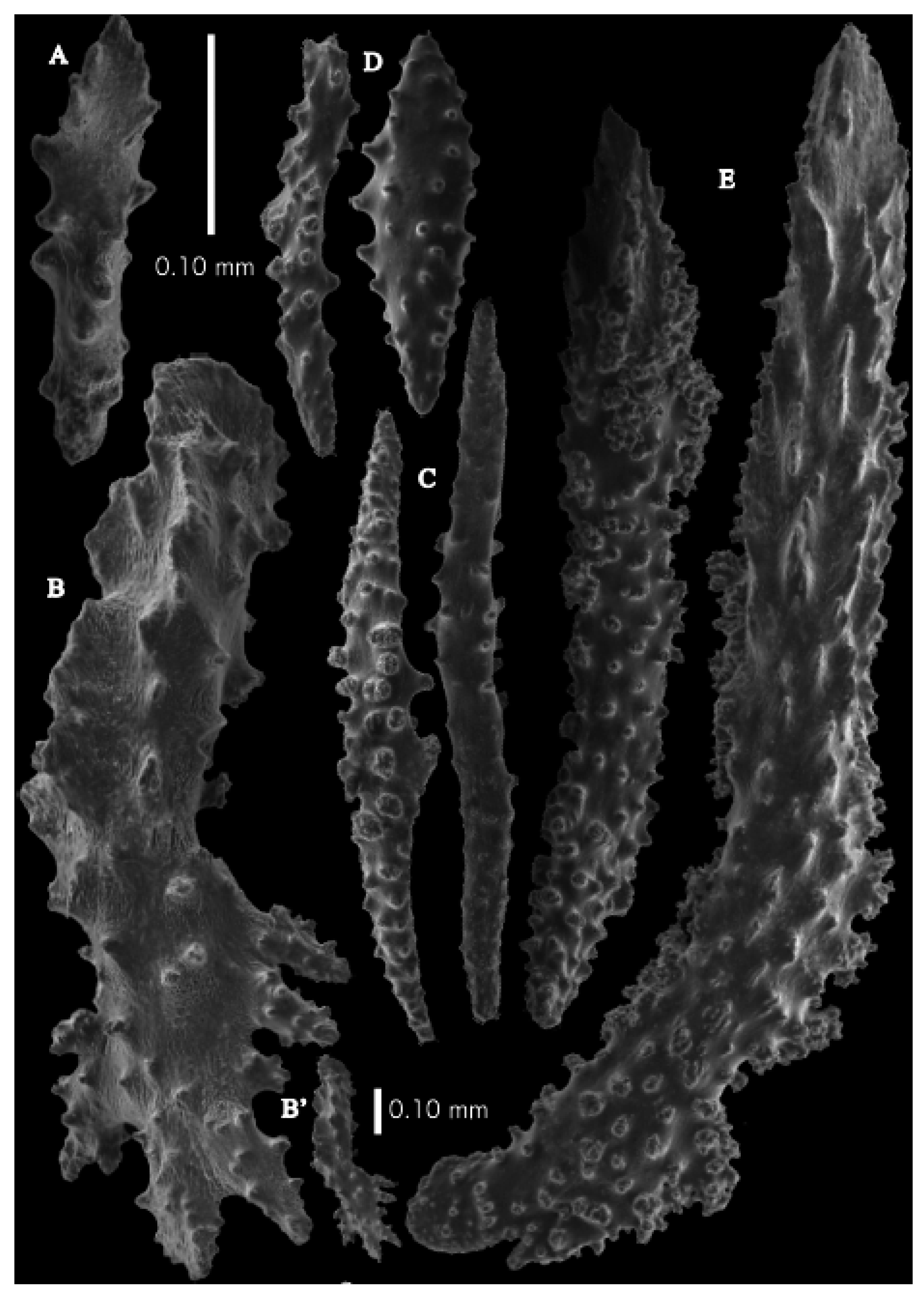
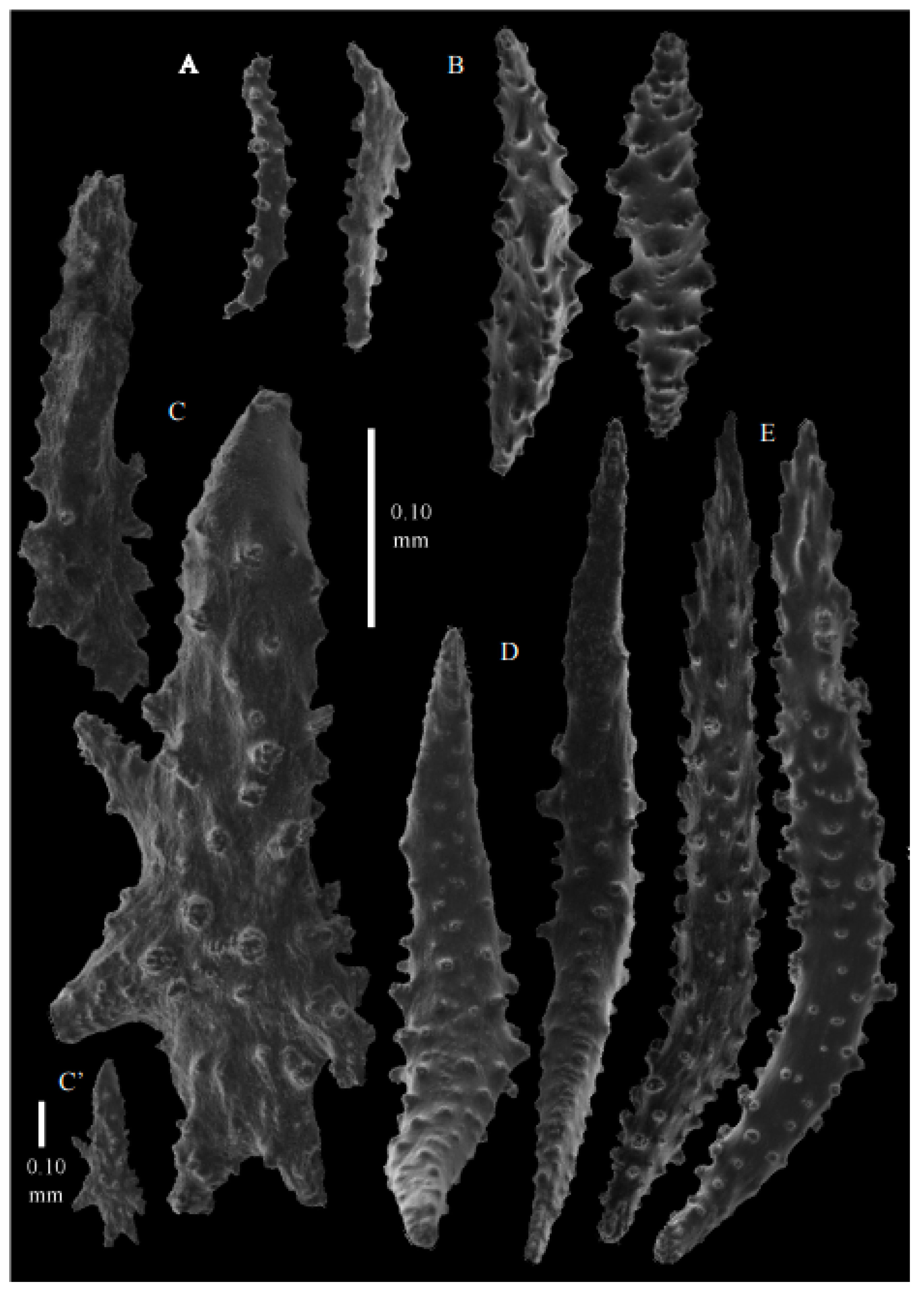
Figure 33D). Calyces conical, about 2 mm high and wide, present on all sides of the colony, often less than 1 mm apart. Polyps with collaret and points (
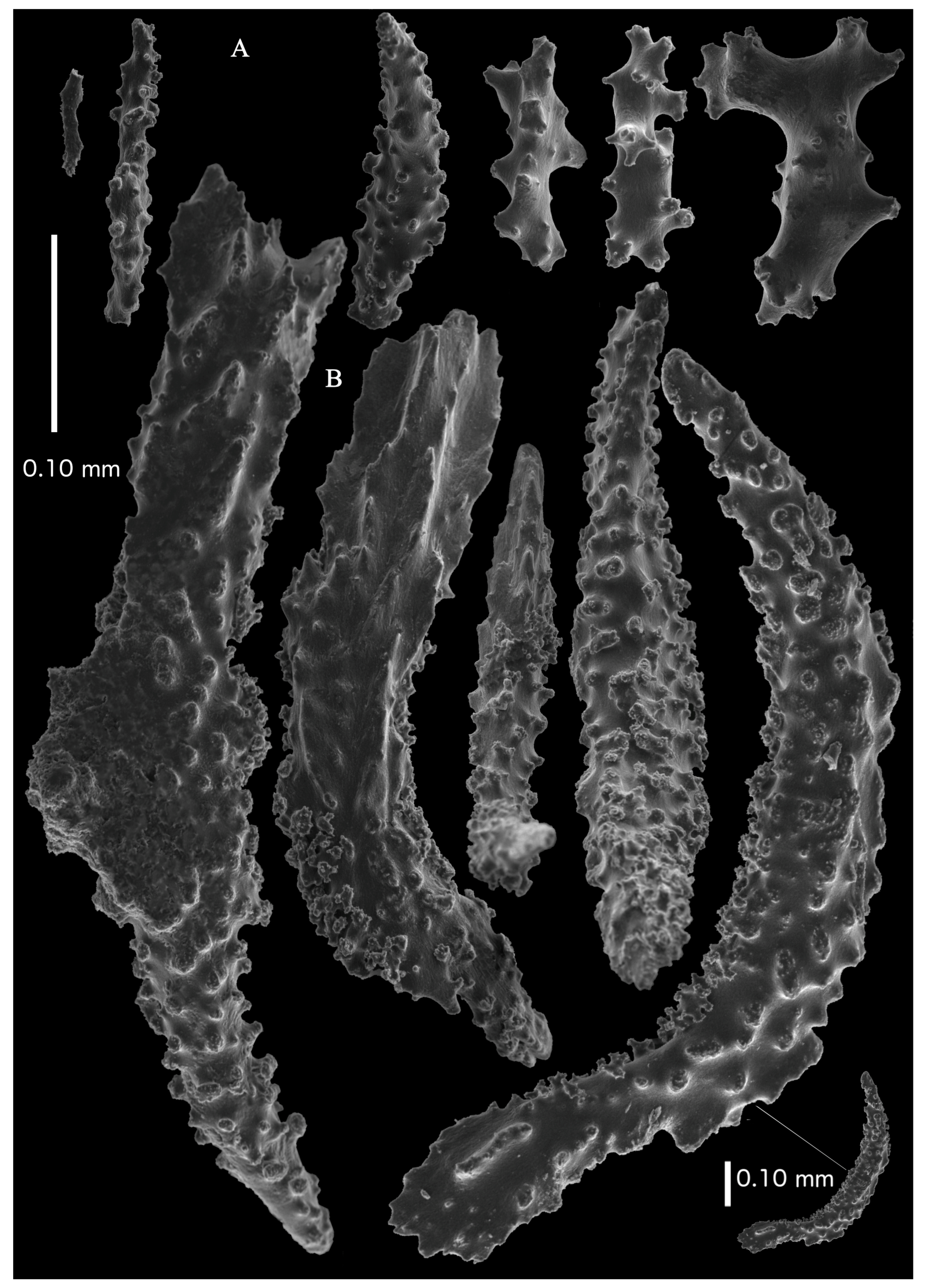
Figure 36A). Point sclerites are bent spindles (
Figure 37D,D’), up to 0.8 mm long, with lower part with simple tubercles, and spiny upper part. Collaret with bent spindles (
Figure 37B), up to 0.5 mm long, with simple tubercles, most pronounced in the middle. Tentacles with rods (
Figure 37A), up to 0.2 mm long, with simple tubercles. Calyces with thornscales (
Figure 38A), 0.3–1.5 mm long. Lower part of the smaller thornscales branched, with simple tubercles, upper part with simple tubercles and spines. Larger thornscales have a less branched lower part. Coenenchyme with big plates, up to 1.2 mm long (
Figure 38B), with simple tubercles. The size of coenenchyme sclerites extends over a wide range. Interior with spindles, up to 0.4 mm long, with simple tubercles (
Figure 37C).
Colours. The live colour of RMNH.COEL.47988 (AKM554) colony is blue (
Figure 33F) and sclerites are white. Nutting mentioned that the colour of the colony is tan-brown but did not mention if it was the live colour or not. Preserved colony and calyces are brown-grey.
Variation. Paratype (USNM 49341) has both branching and sclerites similar (
Figure 33E). Newly collected material (RMNH.COEL.47988 (AKM554)) is a smaller colony but the sclerites are similar to the holotype (
Figure 33F, 39 and 40).
Distribution. Kagoshima, Chiba, Kouchi Prefectures, Pacific side of Japan, Northwest Pacific.
Remarks. From the description of
Paracis cf.
ijimai by Aurivillius [
6], it is obvious he also had
Pseudoparacis but failed to recognise it. Muzik [
9] identified the Hawaiian specimens as
Paracis horrida (Thomson and Henderson, 1906) in her dissertation (USNM56845, USNM56849). However, Hawaiian materials are more like
Pseudoparacis spinifera. The original type description of
Stenella horrida by Thomson and Henderson, 1906 [
17] is unlikely
Pseudoparacis spinifera. As we did not have a chance to examine the type of species, we cannot say for sure what it is based only on their figures and plates.
Pseudoparacis japonica(Nutting, 1912)
Placogorgia japonica Nutting, 1912: 83, pl. 13 Figure 1, 1a, pl. 21 Figure 2 (Ose Saki, Sata Misaki); Kükenthal, 1924: 211.
Discogorgia japonica Kükenthal 1919: 760.
Pseudothesea foliata Aurivillius, 1931: 182, Figure 35, pl. 4 Figure 1 (HongKong).
Material examined. Paratypes of Placogorgia japonica Nutting, 1912, USNM43128 Alb-USFC/A4933, Sata Misaki Cape, Osumi Strait, Kagoshima Prefecture, Kyushu Isl. Japan, North Pacific Ocean, 30.9833° N, 130.497° E, depth 278 m, Northwestern Pacific Expedition, tangle swabs, 16 August 1906; USNM43117 Alb-USFC/A4936, Kagoshima Gulf, Sata Misaki Cape, Osumi Strait, Kagoshima Prefecture, Kyushu Isl. Japan, North Pacific Ocean, 30.9111° N, 130.625° E, depth 188 m, Northwestern Pacific Expedition, Trawl–Agassiz beam, 9 ft, 16 Aug. 1906; holotype of Pseudothesea foliata Aurivillius, 1931, UPSZTY2154 (UUZM57), 50 mile North of Hong Kong. Depth 60 m, coll. J. Petersen. Previously unidentified material: AKM201 Okinawa, R/V Tansei-maru cruise, KT02-03, st.E5-2, 19 April 2002; AKM202 same data as AKM00201; RMNH.COEL.47989 (AKM551) Off Takato, Tachibana-ura, Otsuki, Tosa, Japan, 132°48.06′ E, 32°43.08′ N–132°47.68′ E, 32°43.12′ N, F/B Kiryo-maru, st.3, depth 84.75–83.1 m, 7 October 2004; RMNH.COEL.47990 (AKM552) Off Takato, Tachibana-ura, Otsuki, Tosa, Japan, 132°48.06′ E, 32°43.08′ N–132°47.68′ E, 32°43.12′ N, Kiryo-maru, st.3, depth 84.75–83.1 m, 7 October 2004; RMNH.COEL.47991 (AKM1489) West of Chichijima Is. Ogasawara Isls. Japan, 142°07.166′ E–142°07.251′ E, 27°02.942′ N–27°02.948′ N, depth 138–136 m, R/V Tansei-maru cruise, KT09-02, st. TW02-04, coll.A.K. Matsumoto, 19 March 2009.
Re-description. Paratype colony (USNM 43128) 9.7 cm high and 6.8 cm wide, branched in one plane (
Figure 41B). Calyces conical, about 1.5 mm high and wide, present on all sides on the end branches, on thicker branches more to one side, often less than 4 mm apart. Polyps with collaret and points (
Figure 36B). Point sclerites are bent spindles (
Figure 42E), up to 0.45 mm long, with lower part with simple tubercles, and spiny upper part. Collaret with bent spindles (
Figure 42C), up to 0.40 mm long, with simple tubercles, most pronounced in the middle. Tentacles with rods (
Figure 42A), up to 0.20 mm long, with simple tubercles. Calyces with thornscales (
Figure 42B,B’ and
Figure 43A), 0.3–0.5 mm long. Lower part with complex tubercles, upper part rather blunt-ended distally, with simple tubercles and spines. Coenenchyme with big plates, up to 1.2 mm long (
Figure 43B), with complex tubercles and outside with spines and ridges. Interior with spindles, up to 0.70 mm long, with simple tubercles (
Figure 42D and
Figure 43C).
Colours. Colony and calyces brown-grey. Sclerites white.
Variation. The colony of the other paratype (USNM43117) is shown in
Figure 41C.
Distribution. Kagoshima Prefecture, Ogasawara Isl., Okinawa, Pacific side of Japan, Northwest Pacific.
Remarks. The condition of the specimen of the holotype (USNM30096) was not suitable for examination (
Figure 41A), so we describe paratype USNM43128 (
Figure 41B). We examined the holotype of
Pseudothesea foliata Aurivillius 1931 (UPSZTY2154 (UUZM57),
Figure 36C,
Figure 41D,
Figure 44 and
Figure 45) and synonymised it as
Pseudoparacis japonica, the most common
Pseudoparacis species in Japanese waters.
Pseudoparacis tanseiisp. nov.
urn:lsid:zoobank.org:act:FE167A86-4BDA-4498-843F-5675DAC9B437
NOT Discogorgia dendritica: Utinomi 1961: 203–204, Figure 3; pl. VII, Figure 4 (SMBL-Cni1069).
Material examined. Holotype RMNH.COEL.47992 (AKM750, larger colony) off Satamisaki Cape, Kagoshima Prefecture, Japan, 30-56.0025 N, 130-44.2299 E–30-56.2953 N, 130-43.3981 E, depth 116–120 m, R/V Tansei-maru cruise KT07-1, st. SM-1, specimen.32, Chain Bag Dredge. Obtained 23 July 2007, coll. A.K. Matsumoto; paratypes RMNH.COEL.47993 (AKM750, smaller colony) same data as holotype; RMNH.COEL.47994 (AKM589), Off Minabe, Wakayama prefecture, Japan, Northwest Pacific, depth ca.40 m, Gill-net, coll. A.K. Matsumoto, 22 March 2005; RMNH.COEL.47995 (AKM1478) West of Chichijima Is. Ogasawara Isls. Japan. 142°07.466′ E–142°07.489′ E, 27°01.371′ N–27°01.344′ N, depth 135–136 m, R/V Tansei-maru cruise, KT09-02, st.TW01-02, 3. 19 March 2009, coll. A.K. Matsumoto; other material SMBL-Cni1069 previously identified as Discogorgia dendritica, 23 km west of Setozaki cape, depth 80–150 m, RV Kaiun-maru, coll. H. Harada, 28 November 1957, two specimens.
Description. Holotype (RMNH.COEL.47992 (AKM750) 6 cm high and 3 cm wide, branched in one plane (
Figure 46A). Calyces conical, about 1.3 mm high and wide, present on three sides of the colony, often less than 1.5 mm apart. Polyps with collaret and points (
Figure 36D). Point sclerites are bent spindles (
Figure 47D), up to 0.50 mm long, with lower part with simple tubercles, and spiny upper part. Collaret with bent spindles (
Figure 47A), up to 0.30 mm long, with simple tubercles, most pronounced in the middle. Tentacles with rods (
Figure 47B), up to 0.15 mm long, with simple tubercles. Calyces with thornscales (
Figure 47C and
Figure 48A), 0.4–0.6 mm long. Lower part with complex tubercles, upper part with simple tubercles and spines. Coenenchyme with big plates arranged longitudinally, up to 1.8 mm long (
Figure 48D) and spindles up to 1.6 mm long (
Figure 48C), all with complex tubercles and an outer side with rounded elevations, smaller plates sometimes with outer spines. The size of coenenchyme sclerites extends over a wide range. Several of the spindles are branched (
Figure 48D). Interior with spindles, up to 1 mm long, with simple or complex tubercles (
Figure 4B).
Colours. Live colony pale brown (
Figure 46A, RMNH.COEL.47992 (AKM750)) or pale reddish-brown with polyps which are purplish brown (
Figure 46B,
Figure 49 and
Figure 50, RMNH.COEL.47995 (AKM1478)). After death, its colour turned dark (
Figure 46C, RMNH.COEL.47994 (AKM589)). Anthocodial sclerites colourless, coenenchymal sclerites white.
Variation.
Discogorgia dendritica (Nutting, 1910) in Utinomi 1961 [
18] (SMBL Cni1069) could be this species with the flattened rods in tentacles, thornscales with wide foliaceous projections, and the coenenchyme plates mostly thorny spindles. Calyces are arranged in four longitudinal rows, not in three sides of branches.
Distribution. Kagoshima, Wakayama Prefectures, Ogasawara Isl., Pacific side of Japan, Northwest Pacific.
Etymology. The species is named after the R/V Tansei-maru.
Remarks. The species is closest to P. spinifera but differs in having coenenchymal plates with rounded elevations and thornscales with blunt spines. The two other species, P. japonica and P. spinifera, have polyps distributed all around the branches, but this species has them only on three sides.
Pseudoparacis erythraeensis (Stiasny, 1938)
Discogorgia erythraeensis Stiasny, 1938:339 (Gulf of Suez); Stiasny 1959: 53, fig.C, pl. 6 Figures 24–27; Grasshoff, M. 2000: 65–67, Figures 110–115.
Remarks. We decided that
Discogorgia erythraeensis has to be moved to the genus
Pseudoparacis gen. nov. as it has thornscales in the calyces which are not present in
Discogorgia (see Grasshoff, 2000 [
19]: p. 65–67). Molecular phylogenetics also confirmed that
D. erythraeensis in McFadden et al. [
7] is not close to our newly collected
Discogorgia spp. or to
Euplexaura but is instead close to the group with thornscales, (i.e.,
Placogorgia) (personal communication: Cathy McFadden, 2 July 2021, 17 April 2023).
D. erythraeensis only fits the description of
Pseudoparacis gen. nov.; currently, there are no other possibilities, although the other species of the genus
Pseudoparacis gen. nov. occurred in the North Pacific Ocean. The sclerites may need additional SEM analysis since the drawings of Grasshoff [
19] are a bit schematic.
Genus PSEUDOTHESEA Kükenthal, 1919
Type of the genus: Pseudothesea placoderma (Nutting, 1910).
Thesea (Indo-Pacific) Nutting, 1910, Siboga Exped. Monogr. 13b: 76.
Pseudothesea Kükenthal 1919, Ergebn. deutschen Tiefsee-Exped. 13(2): 843. (Type species, Thesea placoderma Nutting, by original designation).
Placogorgia Bayer 1959: 55 (Pseudothesea placoderma = Placogorgia atlantica).
Pseudothesea Bayer 1981: 927 (Key and footnote).
Diagnosis. Colonies richly branched in one plane. Polyps situated all around the end branches, on thicker branches more to one side, or on three sides of the end branches. Coenenchyme with surface layer of unilaterally spinose spindles/plates on three sides, the other side with larger and longer spiny spindles/plates tiled as a mosaic. Below these big sclerites lie smaller ones. Calyces cylindrical, with thornscales. Calyces and polyp-side coenenchyme surface look spiny. Polyps with collaret and points, tentacles with dragon wing scales. Sclerites colourless.
Remarks. Kükenthal [
2] established the genus
Pseudothesea with
Thesea placoderma Nutting 1910 as its type species. Bayer [
8] wrote, “the character of its calicular thornscales leaves no doubt that
Thesea placoderma is congeneric with
Placogorgia atlantica Wright and Studer.” He made the key for
Placogorgia atlantica (including
Thesea placoderma) as follows “Cortical spindles small, usually not exceeding a length of 0.5 mm, without a row of prominent external spines [
8]:
Figure 11b. Thorn-scales 0.5 mm long, spine 0.3 mm, sharp and aculeate [
8]:
Figure 11a)”. He synonymised
Pseudothesea Kükenthal, 1919 as the genus
Placogorgia Studer, 1887 (Bayer [
8]). However, he did not mention which specimen he examined for the [
8]:
Figure 11a,b in his paper.
Grasshoff 1977 [
20]: p. 26 examined the type specimen of
Placogorgia atlantica Wright and Studer (St. Paul’s Rocks) and wrote that “it has, as Bayer emphasised [
8], only a few spindles in the coenenchyme and they are normal spindles, so it largely lacks a generic character of
Placogorgia. The species does not seem to have been found again. However, there can be no doubt that Deichmann’s [
21]: p.145 specimens determined as
Echinomuricea atlantica Johnson are identical to the type species
P. atlantica Wright and Studer. The colonies (checked: MCZ No. 4652, 4660, 5837 =? 4655) show the same structure and sclerites as the type specimen in the British Museum, only having more thornspindles in the coenenchyme than the type...(continued)” [translated from German].
The last to discuss the genus was Bayer [
13] in his key to the genera. In his footnote 15, he stated, “Although I have previously synonymised this genus with
Placogorgia (Bayer, 1959), it seems to be generically distinct, at least from the type-species of that genus. It may however, be impossible to distinguish it from
Paracis, from which it was distinguished by Kükenthal [
3] on the basis of having sclerites of very diverse form, including strongly spinose, unilateral thorn-scales, but these occur in both
Pseudothesea and
Paracis.” Bayer mentions for
Placogorgia “Coenenchymal sclerites not conspicuously large, projecting spines of only infrequent occurrence”, and for
Pseudothesea “Coenenchymal sclerites thick, coarse spindles or plates with one to several strong, projecting spines” [
13].
We examined the type material of
Pseudothesea placoderma from the SIBOGA expedition (type ZMA Coel. 3569, BMNH1925.07.25.013 and non-type ZMA Coel. 3570) in detail. However, we did not have a chance to examine the type of
Placogorgia atlantica and Deichmann’s [
20]: p. 145 specimens (MCZ Nr. 4652, 4660, 5837=?4655 [
20]). The drawing of sclerites in Bayer [
8] shows that the coenenchyme sclerites of
P. atlantica [
8]:
Figure 11b are not identical to
P. placoderma [
8]:
Figure 14a,b, especially the spiny outer coenenchyme sclerites. It is also different from the sclerites in the original Wright and Studer
P. atlantica (1989: PL. XXVII.
Figure 2 [
22]).
We conclude that Bayer’s ideas about
Paracis were based on his description of a specimen identified by him as
Paracis orientalis (Ridley, 1882) from Bikini Atoll (Bayer [
10]: USNM44078) and may also be affected by a specimen identified by him as
“Paracis” placoderma from Japanese waters (USNM91880, one of the paratype specimens of
Acanthogorgia paradoxa Nutting 1912; note: the other specimens of the type actually belong to the genus
Acanthogorgia and USNM30129, USNM30130, USNM30131). Bayer never checked the type materials of
Neoacis orientalis, P. atlantica, and
P. placoderma. He must have based his description on that of Ridley [
12] which was rather confusing and gave the impression the coenenchymal sclerites had spines. Later on, (1996) Grasshoff [
15] did examine the type of
Neoacis orientalis in order to describe his
P. caecilia from New Caledonia, but he did not provide any images of the coenenchymal sclerites of
N. orientalis.
We re-examined the types of N. orientalis and P. placoderma and presented images of their sclerites. From that, it became obvious that the coenenchymal sclerites of N. orientalis have no spines but are merely protuberances with tubercles, and the outer side of these spindles have rounded tubercles, a character also not seen in Pseudothesea.
We assume that all species from the Indo-Pacific with big unilaterally spinose spindles/plates in the coenenchyme belong to
Pseudothesea and not to
Placogorgia or
Neoacis (
Paracis). McFadden et al. [
7] mentioned that no or only limited molecular data are available to confirm the phylogenetic affinities of
Pseudothesea Kükenthal, 1919.
Key to the species (three species)
1. Collaret spindles with complex tubercles Pseudothesea placoderma (Nutting, 1910);
-. Collaret spindles with simple tubercles.
2. Coenenchyme with unilaterally spinose spindles with complex tubercles P. satsumaensis sp. nov.;
-. Coenenchyme with unilaterally spinose spindles with complex tubercles and rounded elevations with prickles P. tokaraensis sp. nov.
Pseudothesea placoderma (Nutting, 1910)
Thesea placoderma Nutting, 1910: 54, pl. 9 Figure 3, pl. 21 Figure 2 (Flores Sea, 73 m);
not Thesea placoderma Nutting, 1912: 6, 80. (Japan).
Pseudothesea placoderma Kükenthal, 1919: 843; 1924: 228.
Material examined. Holotype ZMA Coel. 3569 Thesea placoderma, SIBOGA st.310, 8°30′ S., 119°7′.5 E. Flores Sea, depth 73 m, sand, with few pieces of dead coral.; Type BMNH1925.07.25.013 wet.
Re-description. Holotype colony (ZMA Coel. 3569) branched in one plane, 7.5 cm high and 6.4 cm wide. Calyces are present on all sides of the branches. Branch ends are swollen (
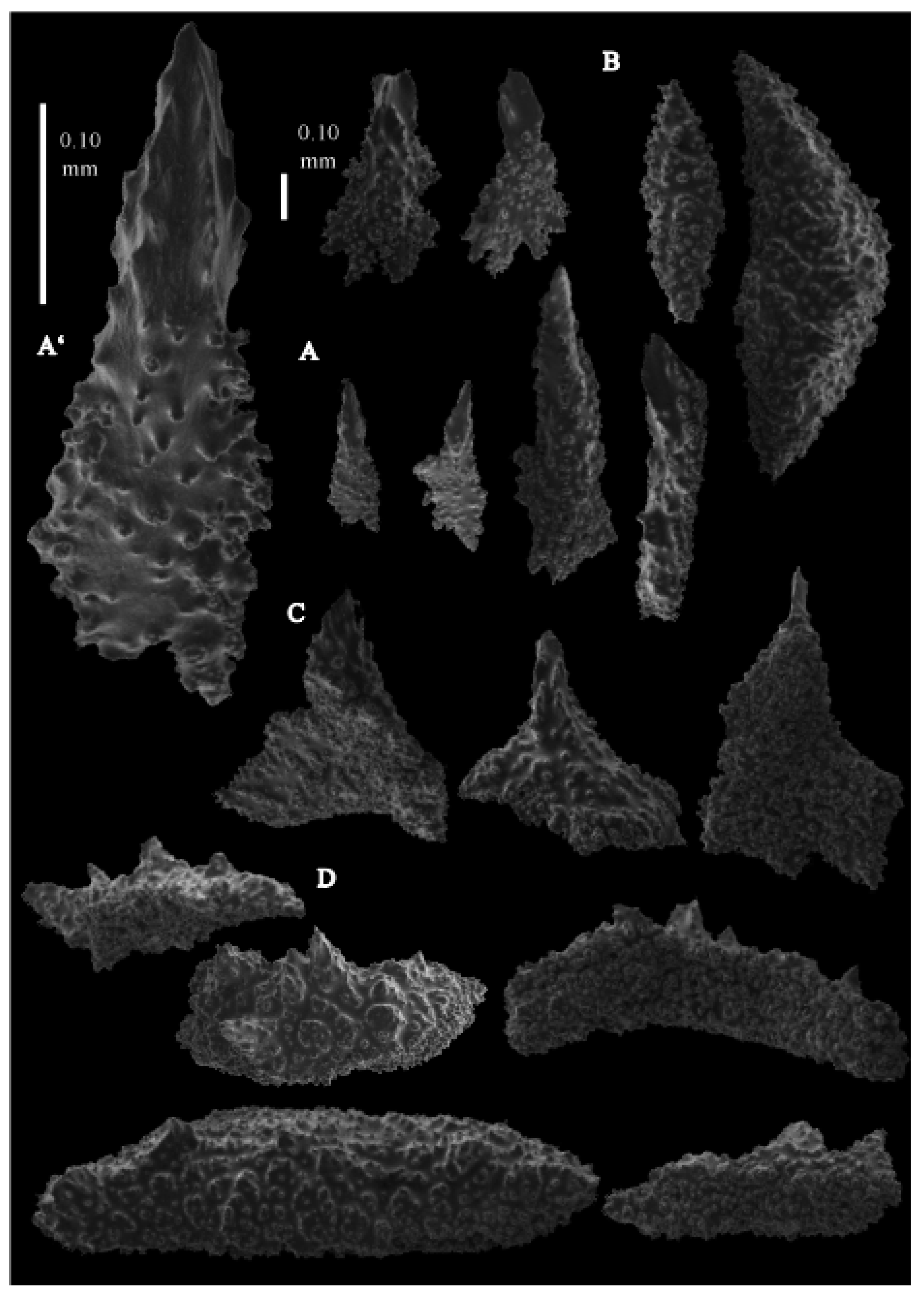
Figure 51A). Calyces are conical, 1 mm high and 1.5 mm wide at their base. Polyps with collaret and points (
Figure 52A). Point sclerites are bent spindles with complex tubercles and distal end spiny, and the basal end is with simple tubercles. These spindles are up to 0.46 mm long (
Figure 53C and
Figure 54A). There is an occasional point sclerite bifurcates at the distal end (
Figure 54B). Collaret with bent spindles, up to 0.58 mm long, middle part with complex tubercles and sometimes small side branches, ends with simple tubercles (
Figure 53B). Tentacles with small spindles, up to 0.13 mm long, with simple tubercles; the largest spindles dragon-wing-shaped (
Figure 53A,D). Calyces with thornscales, up to 0.57 mm long, with simple and complex tubercles (
Figure 54C,D). Surface coenenchyme with unilaterally spinose spindles, up to 1.14 mm long, with complex tubercles (
Figure 54E). Interior with small spindles with simple tubercles (
Figure 54F).
Colours. Preserved colony pale brown, sclerites colourless.
Distribution. The species is only known from its type locality, Flores Sea.
Remarks. Only one interior sclerite was found (
Figure 54F), probably because of the degraded condition of the material. Regarding
Pseudothesea, we should keep
Neoacis and
Pseudothesea separate. However,
Pseudothesea resembles the Atlantic
Placogorgia and
Paramuricea. We also examined non-type material, ZMA Coel. 3570 (SIBOGA Expedition, st. unknown), but it is not
P. placoderma.
Pseudothesea satumaensis sp. nov.
urn:lsid:zoobank.org:act:F2BF8D37-D867-44D6-8648-F599A9753665
NOT
Placogorgia placoderma Bayer, 1959:55
Figure 14ab.
Material examined. Holotype USNM30129 Thesea placoderma by Nutting (1912: 6), Placogorgia placoderma by F.M. Bayer (13 September 1958), Alb-USFC/A4936, Sata Misaki Lt., N.21-E, 5.7 miles, Osumi Strait, Kagoshima, Kyushu Island, Japan, 30°54′40″ N (30.9111° N), 130°37′30″ E (130.625° E), depth 188 m (103 fathoms), Trawl–Agassiz beam, 9 ft, 16 August 1906; paratypes USNM30130 same data as USNM30131 and USNM30129, previously identified as Paracis placoderma by F.M. Bayer (1 July 1994); USNM30131 same data as USNM30130 and USNM30129, previously identified as Paracis placoderma by F.M. Bayer (1 July 1994); RMNH.COEL.47996 (AKM1069) Shin-sone, Danjo Isls., Japan, East China Sea, depth 204–200 m, R/V Tansei-maru cruise KT08-3 (OjiLeg.), GT02(1), specimen no.118, coll. A.K. Matsumoto, 7 March 2008; RMNH.COEL.47997 (AKM1087) Shin-sone, Danjo Isls., Japan, East China Sea, depth 200–210 m, R/V Tansei-maru cruise KT08-3 (OjiLeg.), GT02(2), coll. A.K. Matsumoto, 7 March 2008; RMNH.COEL.47998 (AKM1090) Shin-sone, Danjo Isls., Japan, East China Sea, depth 200–210 m, R/V Tansei-maru cruise KT08-3 (OjiLeg.), GT02(2), specimen no., coll. A.K. Matsumoto, 7 March 2008.
Description. Holotype colony branched in one plane, 5.7 cm high and 7.2 cm wide (
Figure 51B). Calyces are present on three sides of the branches. Branch ends are swollen. Calyces are conical and 0.6 mm high and 0.8 mm wide at their base. Polyps with collaret and points (
Figure 52B). Point sclerites are bent spindles with complex tubercles and distal end spiny, basal end with simple tubercles. These spindles are up to 0.34 mm long (
Figure 55D). Collaret with bent spindles, up to 0.3 mm long, middle part with simple tubercles and sometimes small side branches, ends are smooth (
Figure 55C). Tentacles with small spindles, up to 0.18 mm long, with simple tubercles; the largest spindles dragon-wing-shaped (
Figure 55A). Calyces with thornscales, up to 0.30 mm long, with simple and complex tubercles (
Figure 55B,B’ and
Figure 56A). Surface coenenchyme with unilaterally spinose spindles, up to 1.0 mm long, with complex tubercles (
Figure 56B). Interior with smaller spindles, up to 0.34 mm long, with simple or complex tubercles (
Figure 55E and
Figure 56C).
Colour. Colony colour yellowish white, sclerites colourless.
Variation. Paratype USNM30130 colony is smaller than the holotype but the sclerites are similar (
Figure 51C and
Figure 52C). USNM30131 colony colour slightly darker than USNM30129 and USNM30130 (
Figure 51D and
Figure 52D).
Distribution. The species is only known from its type locality, Kagoshima Prefecture, Japan, Northwest Pacific.
Etymology. The species is named after its type locality, Satsuma, the old name of Kagoshima Prefecture.
Remarks. The label identification is
Paracis placoderma, but that species does not exist. Bayer [
8] once synonymised
Pseudothesea and
Placogorgia, saying the type of the species
Pseudothesea placoderma is identical to
Placogorgia atlantica, but later on, he said that it is different [
13]. Bayer’s identification of materials USNM30130-30131 and USNM91880 may show his confusion in the 1981 paper. Bayer [
13] mentioned that the characters of
Pseudothesea and
Paracis are similar. This confusion by Bayer could be caused by the fact that he never examined the type material of
Acis orientalis (the type of the genus
Neoacis (
Paracis)).
P. satsumaensis sp. nov. resembles
Pseudothesea placoderma but differs in having unilaterally spinose coenenchymal spindles with complex tubercles and collaret spindles with simple tubercles.
Pseudothesea tokaraensissp. nov.
urn:lsid:zoobank.org:act:75526E74-2B17-4BCB-BFBF-704AB51A6213
Material examined. Holotype. RMNH.COEL.47987 (AKM797) Off Takarajima Is., Tokara Isls., East China Sea, 29°14.6410 N, 129°07.8392 E, depth 156 m, R/V Tansei-maru, KT07-2 cruise, st.DT5 (D8), coll. Yokose, 2 March 2007.
Description. Colony branched in one plane, 9 cm high and 12 cm wide (
Figure 51E). Calyces are present on three sides of the branches. Branch ends are swollen. Calyces are conical, 0.63 mm high and 1.3 mm wide at their base. Polyps with collaret and points (
Figure 52E). Point sclerites are bent spindles with complex tubercles and distal end spiny, basal end with simple tubercles. These spindles are up to 0.45 mm long (
Figure 57E). Collaret with bent spindles, up to 0.5 mm long, middle part with simple tubercles and sometimes small side branches, ends are smooth (
Figure 57C). Tentacles with small spindles, up to 0.2 mm long, with simple tubercles; the largest spindles dragon-wing-shaped (
Figure 57A,B). Calyces with thornscales, up to 0.46 mm long, with simple and complex tubercles and rounded elevations with prickles (
Figure 57D,D’ and
Figure 58A). Surface coenenchyme with unilaterally spinose spindles, up to 1.77 mm long, with complex tubercles and rounded elevations with prickles (
Figure 58B). Interior with small spindles, up to 0.3 mm long, with complex tubercles (
Figure 57F and
Figure 58C).
Colour. Colony colour greyish-brown with dark polyps, sclerites colourless.
Distribution. Only known from its type locality, off the Tokara Islands, Japan, East China Sea.
Etymology. The species is named after its type locality, the Tokara Islands.
Remarks. This species with coenenchymal surface sclerites with rounded elevations with prickles shows characters of both Paracis and Pseudothesea. We put this species in Pseudothesea because of the spines on the coenenchyme sclerites.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

























































