
The Relationship between the Rhizothrichidae Por (Copepoda: Harpacticoida) and the Cletodoidea Bowman & Abele, Including the Establishment of a New Genus and the Description of a New Species †


Abstract
:1. Introduction
2. Material and Methods
3. Results
3.1. Taxonomy
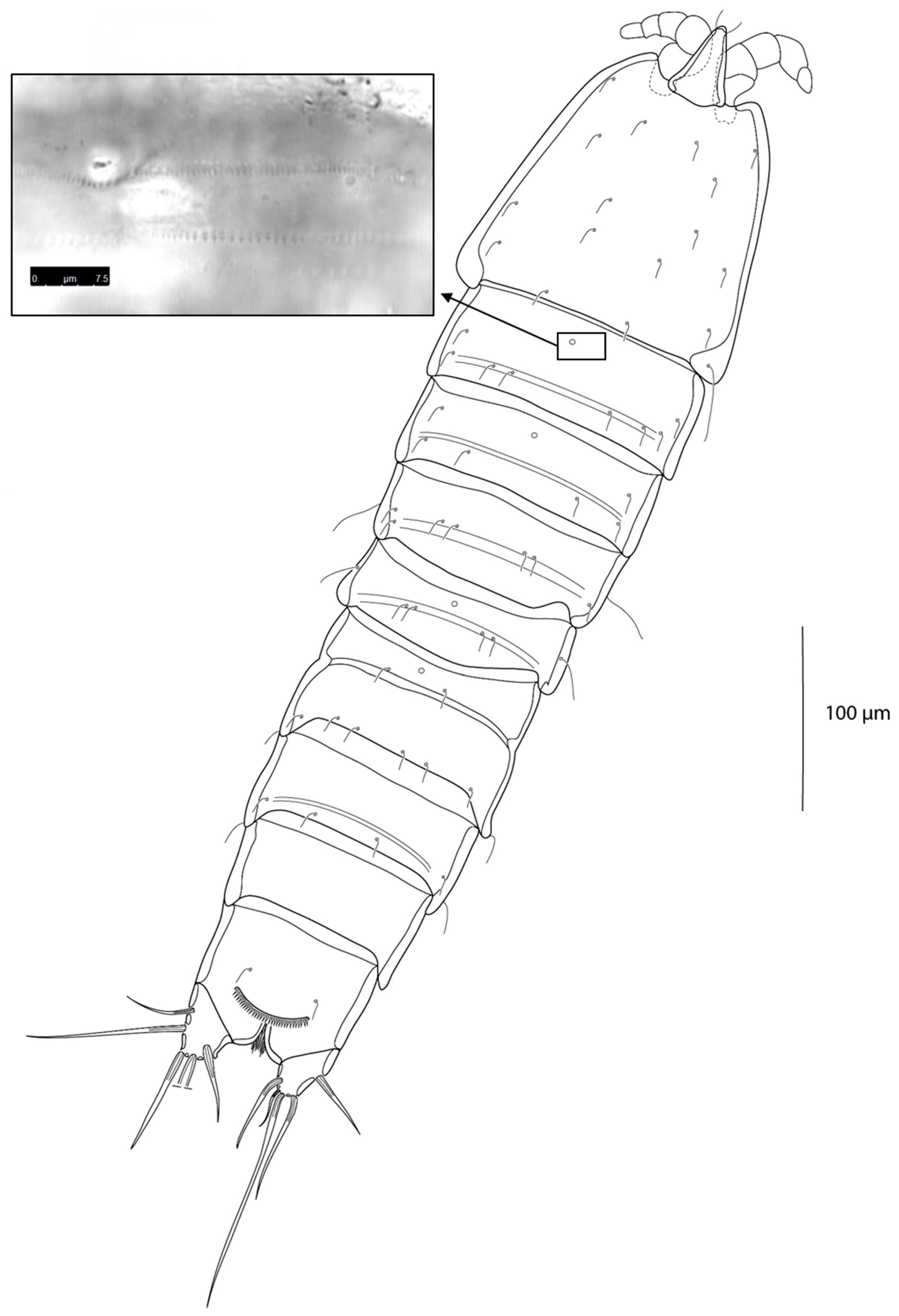
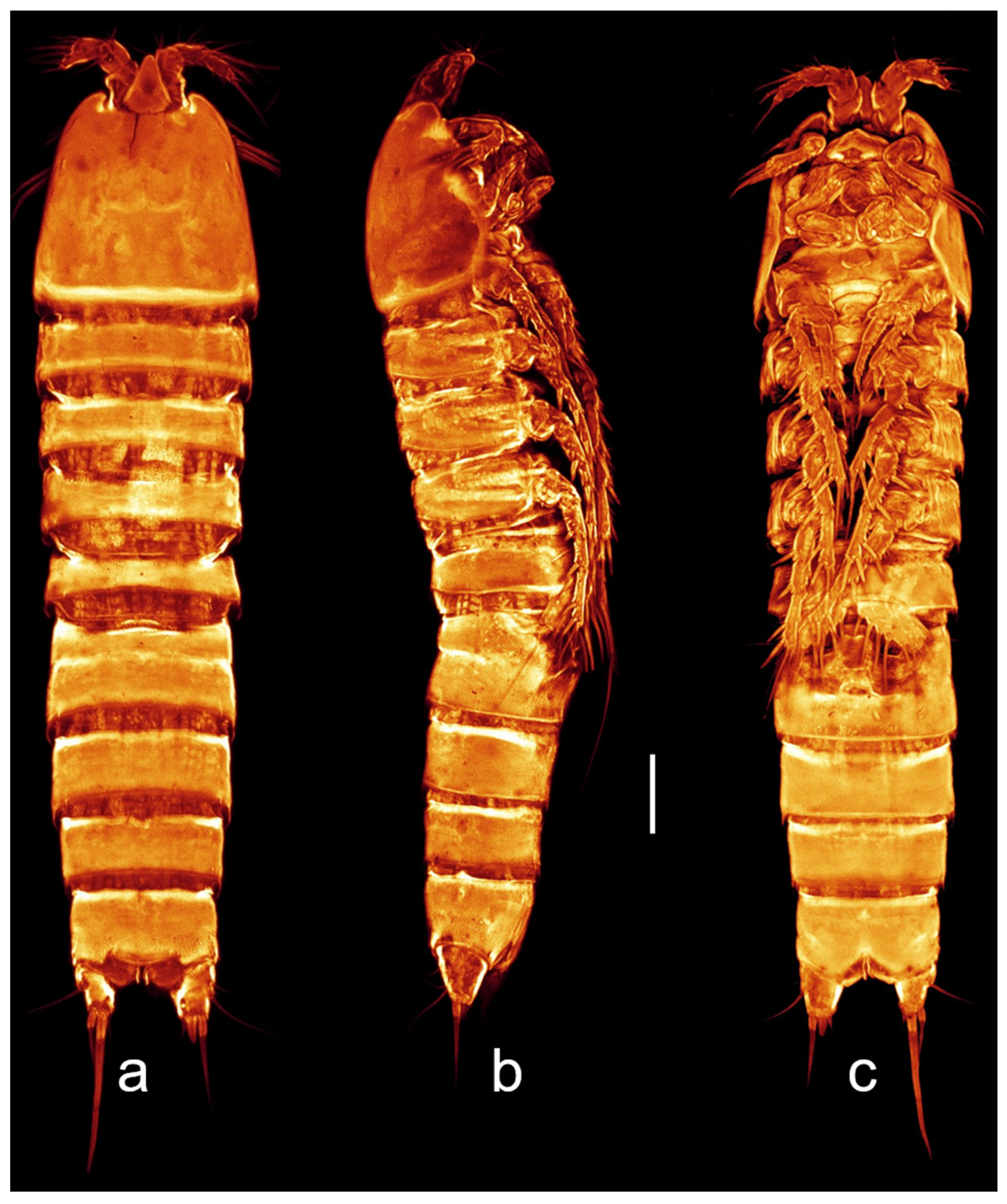
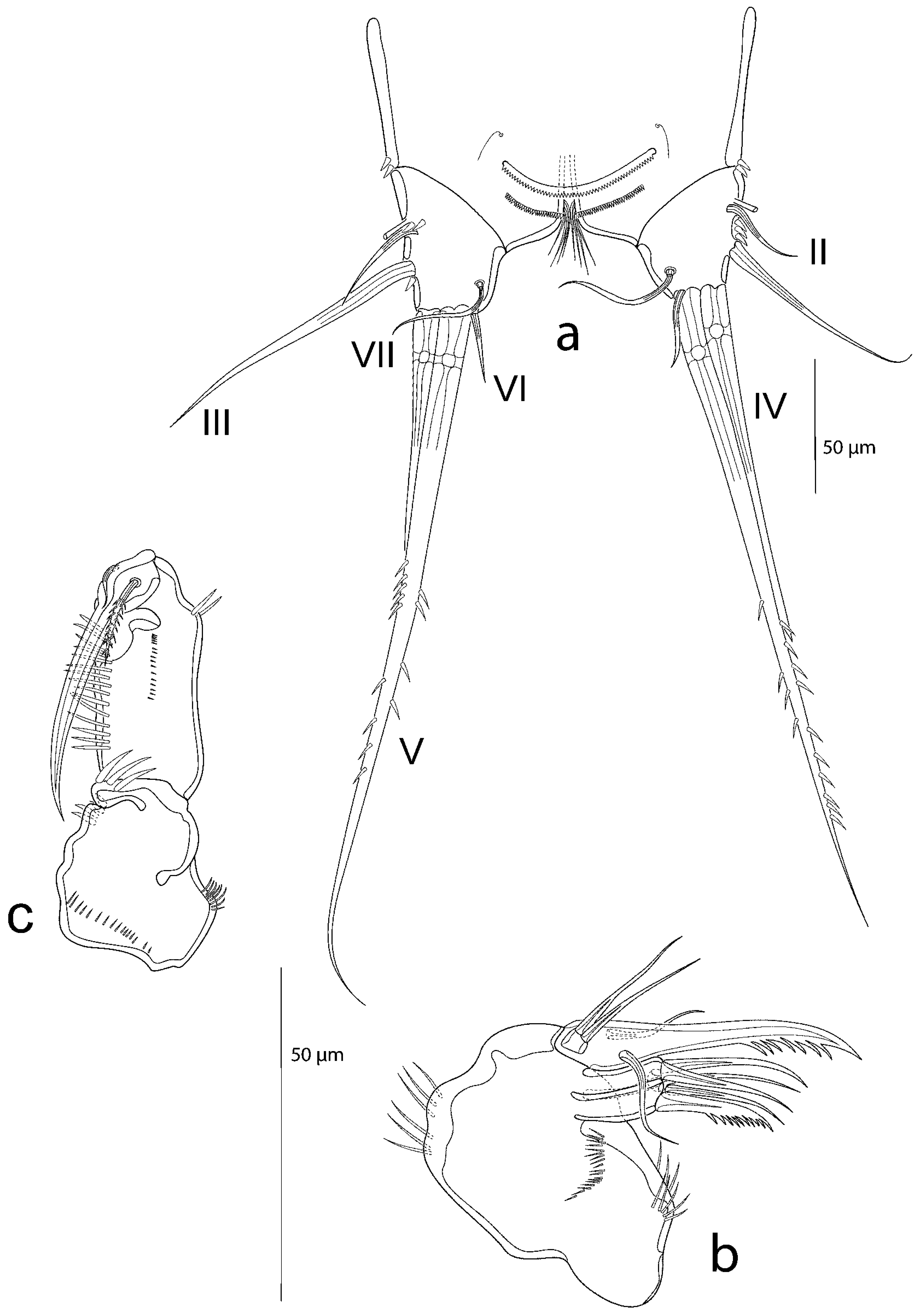
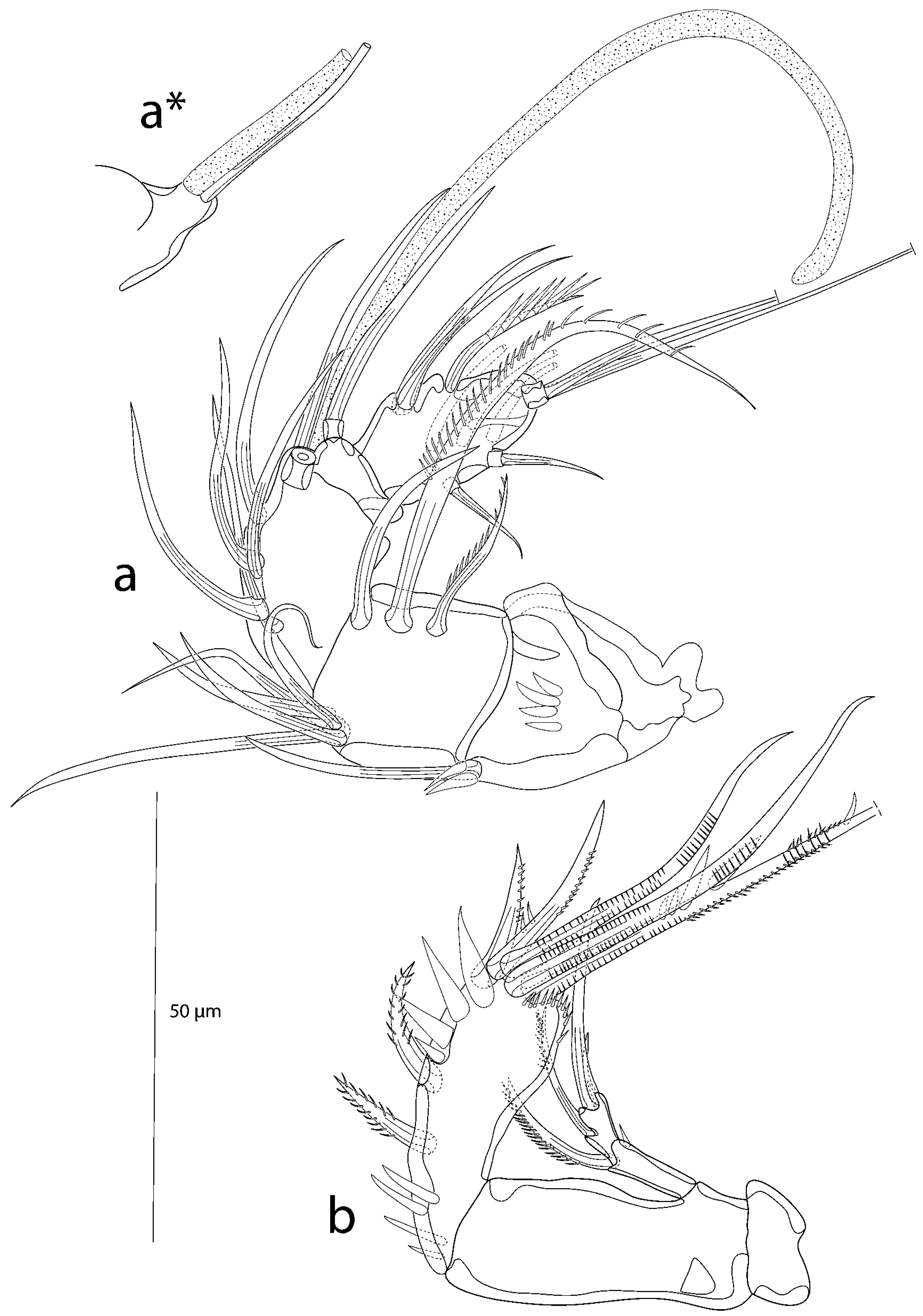
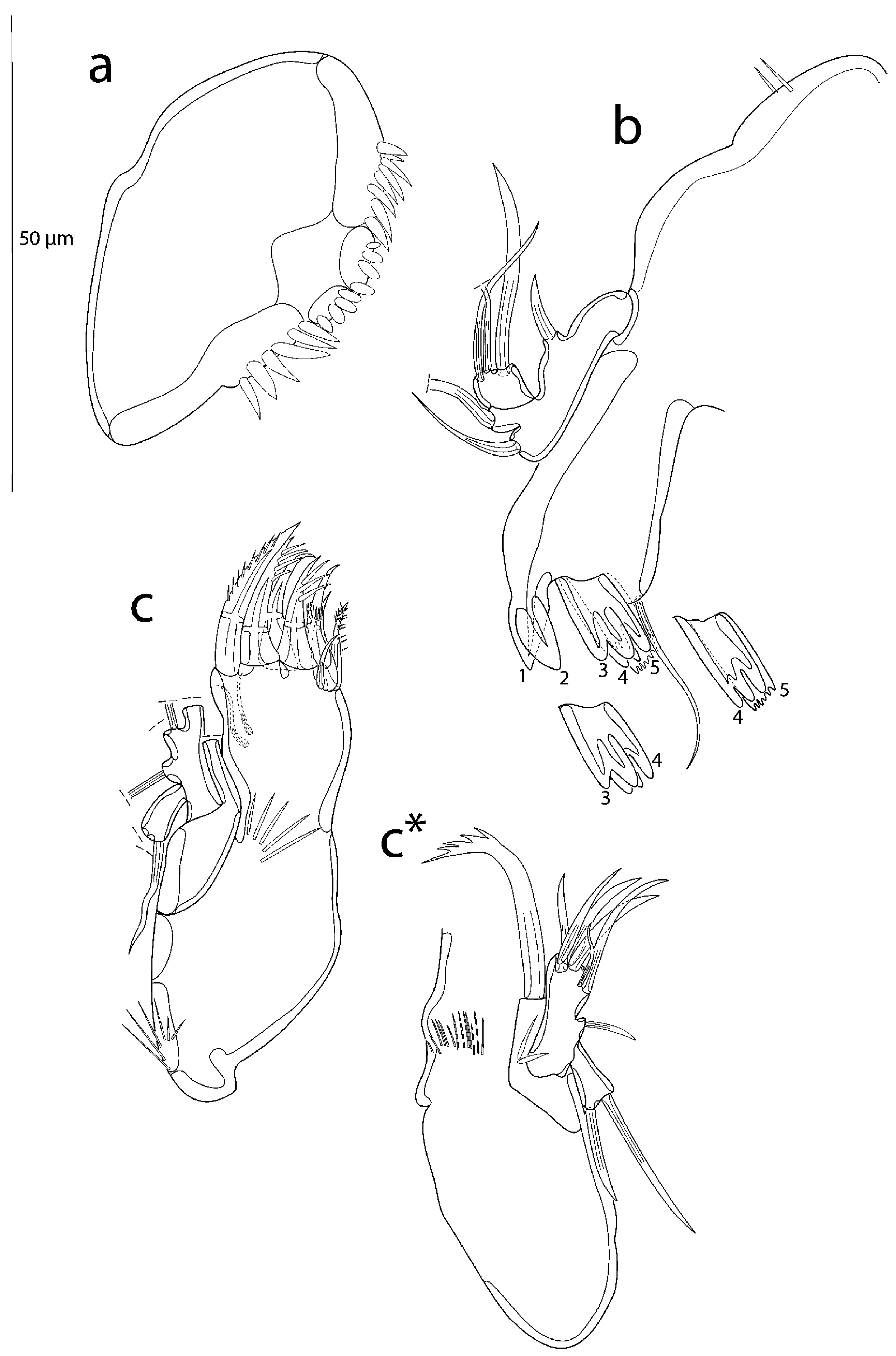
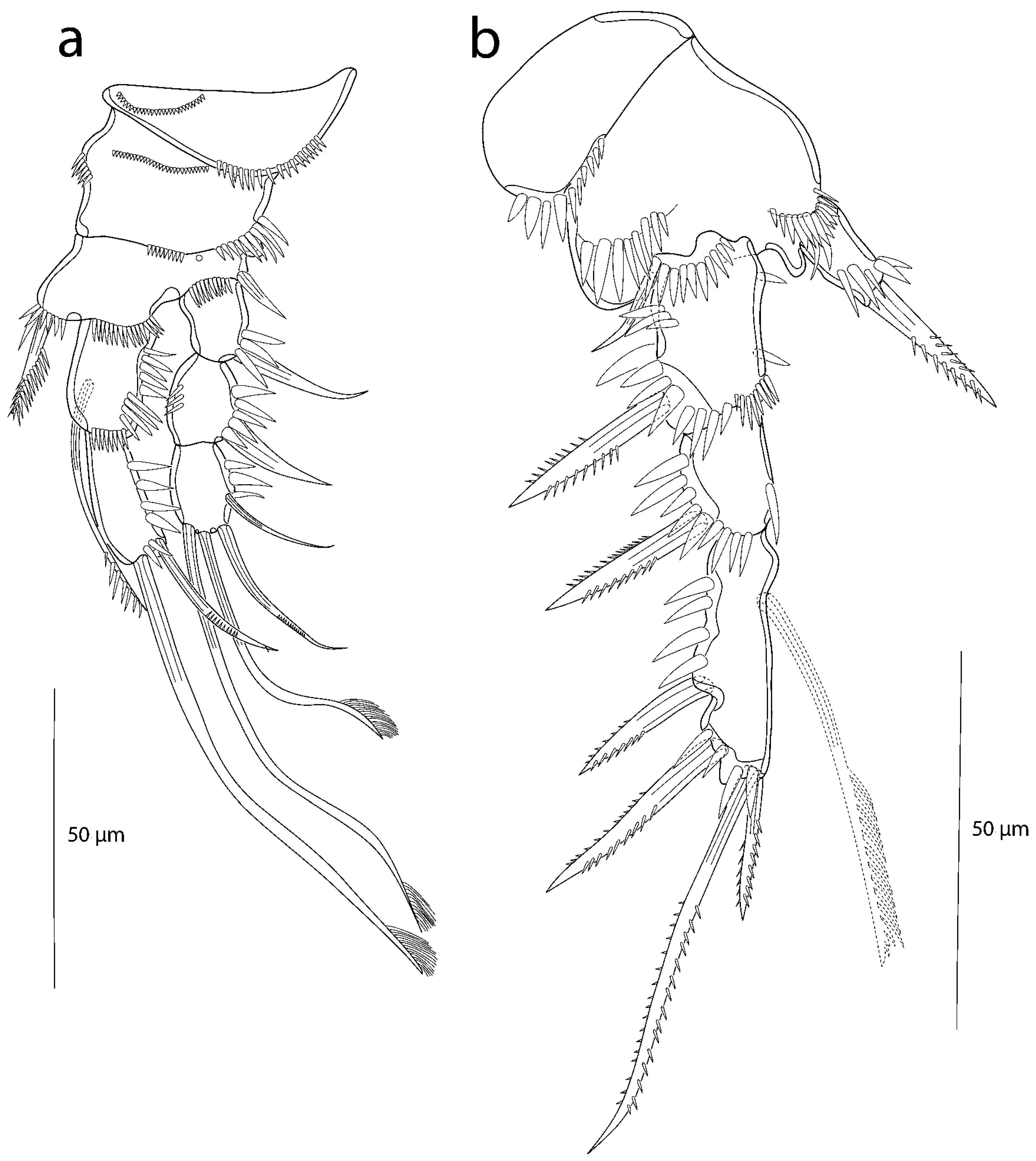
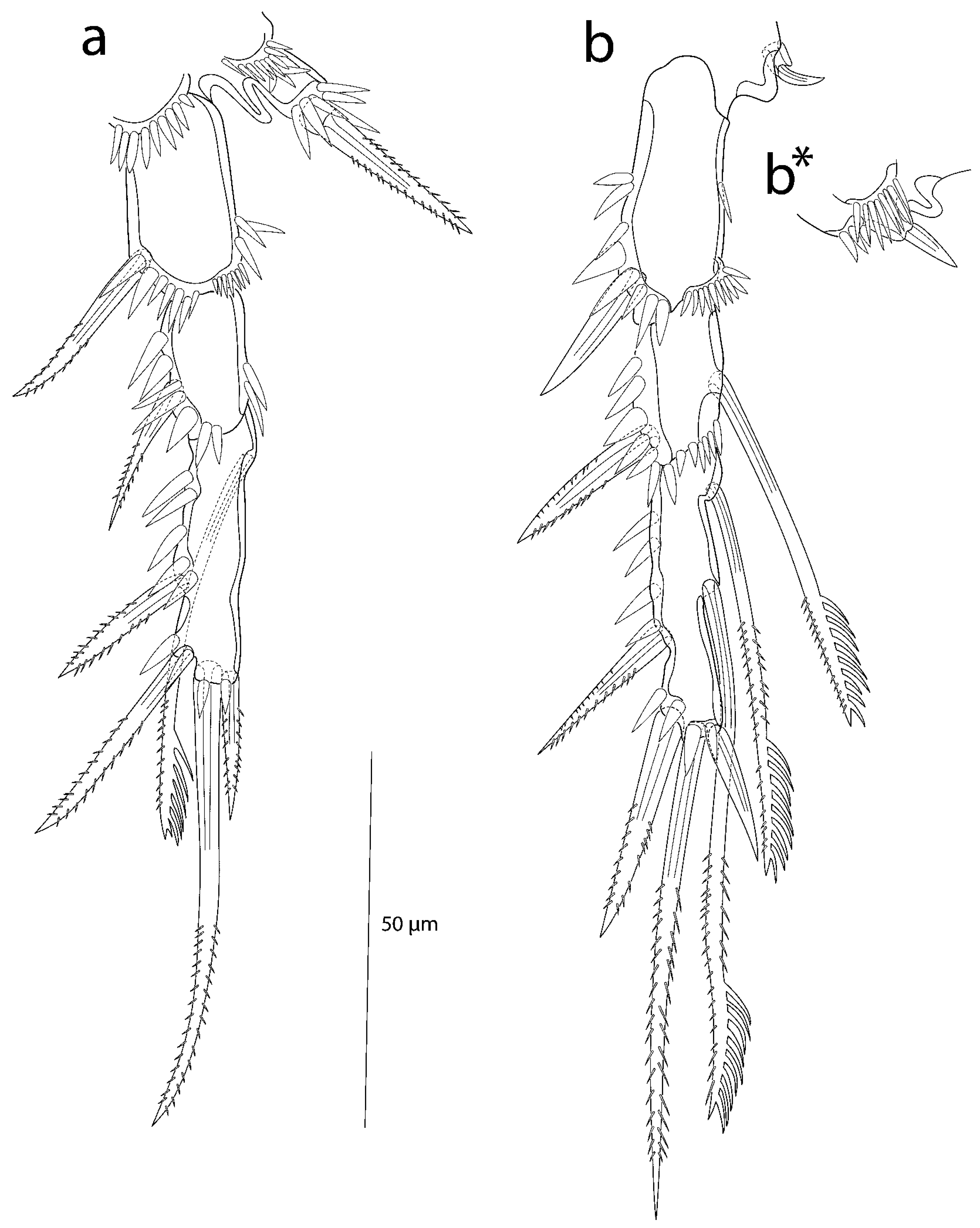
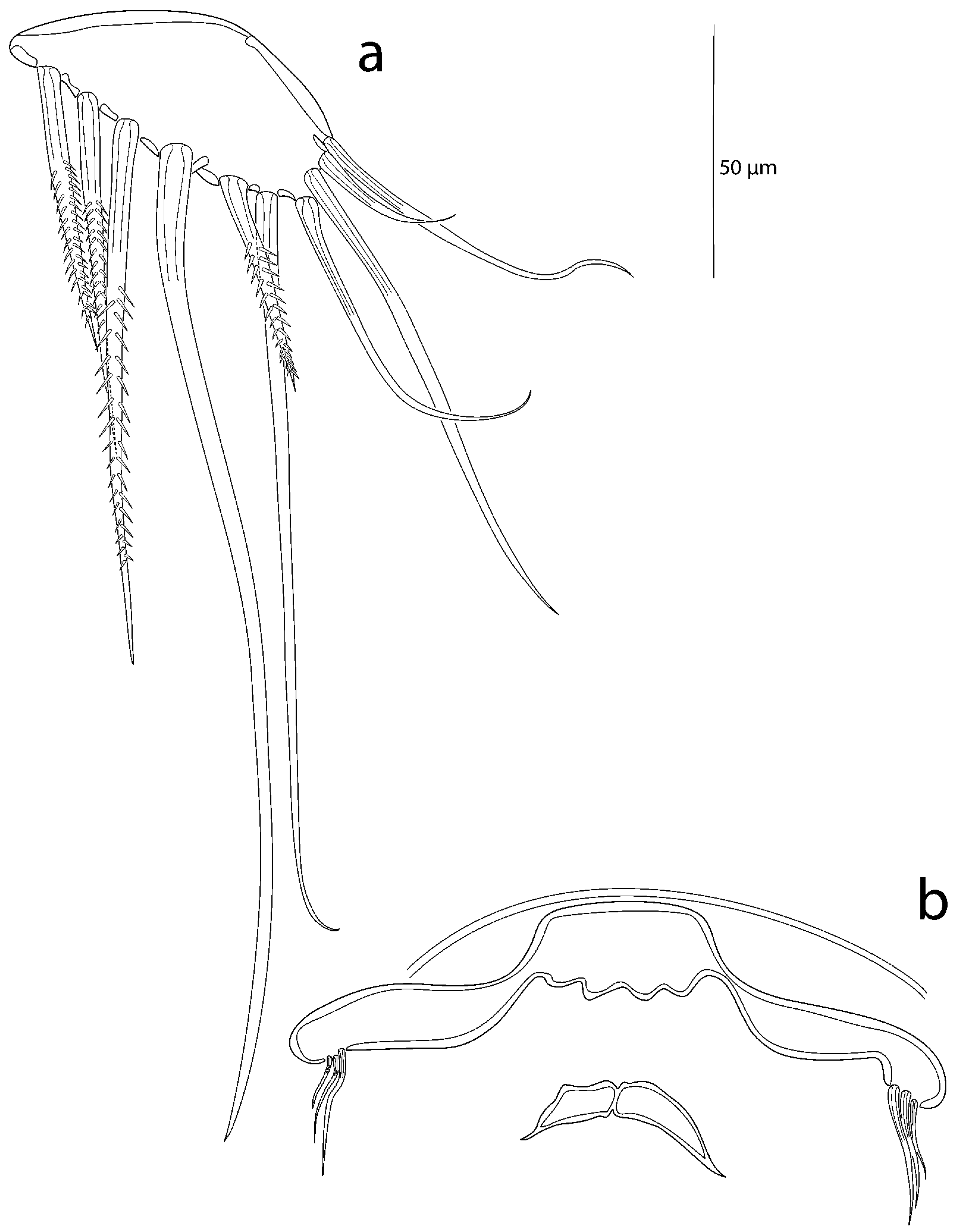
3.2. Description of the Female
- A2 allobasis with abexopodal seta; A2 exopod 1-segmented; P1 enp-1 without setae; P1 enp-2 inner and outer apical seta brush-like; 2
- -
- A2 allobasis without abexopodal seta; A2 exopod 2-segmented; P1 enp-1 with inner seta; P1 enp-2 inner apical seta brush-like; outer seta bare, short, geniculated, not brush-like; P2–P4 endopods 1-segmented, with 1 seta Monopenicillus gen. nov.
- 2.
- P2–P4 exopods 2-segmented; P2–P4 endopods 1-segmented, with at least 2 setae Tryphoema
- -
- P2–P4 exopods 3-segmented; P2–P4 endopods 2-segmented Rhizothrix.
4. Discussion
4.1. Outgroup and Character Selection
4.2. Phylogenetic Analysis
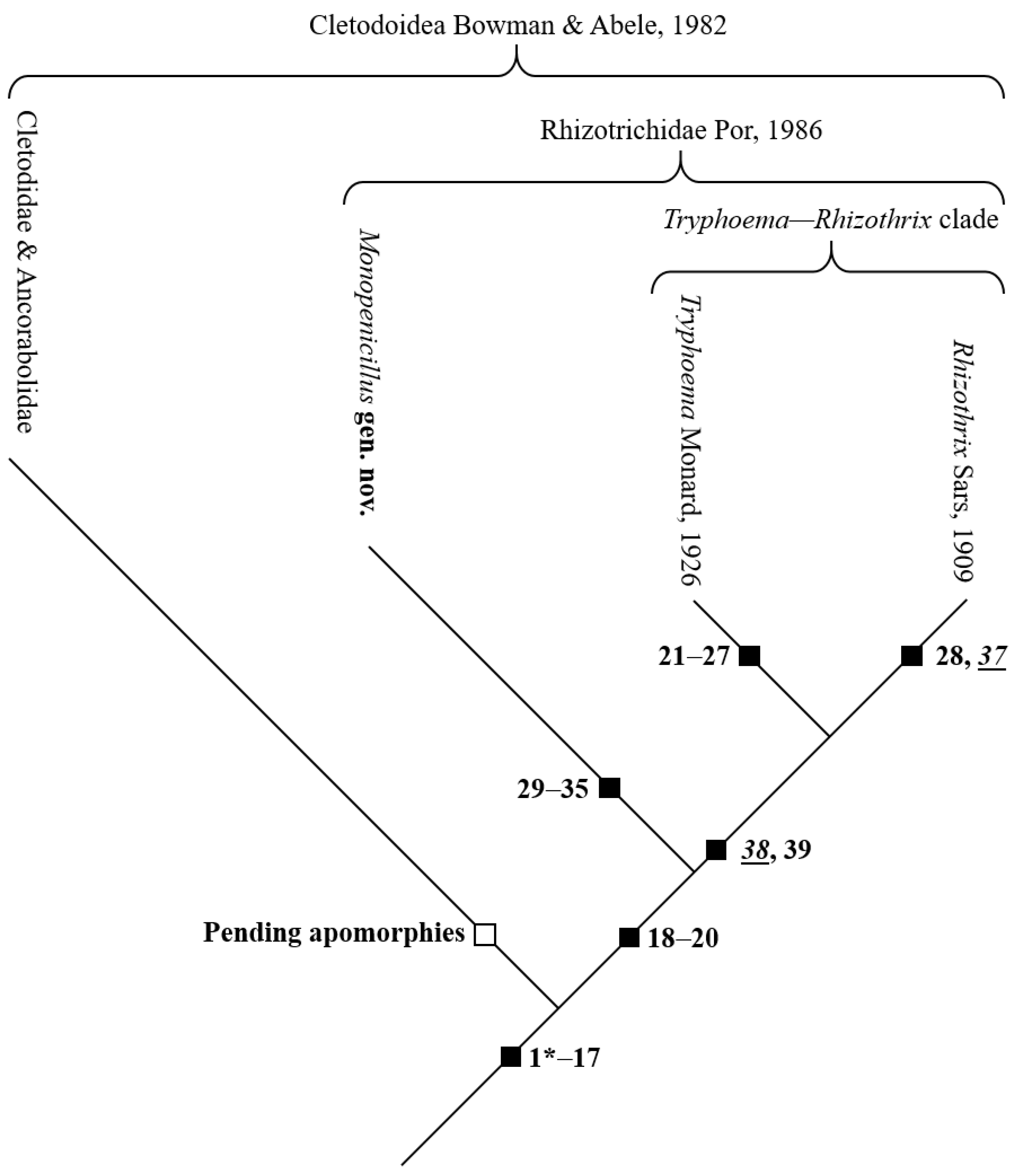
4.2.1. Assignment of the Rhizothrichidae to the Cletodoidea
- Body covered by a dense pubescence [body not completely covered with setules, denticles, or spinules];
- Female A1 distal segment apically with 1 strong spine [apical seta of distal segment of female A1 not remarkably short and thickened];
- P1 exopod distal segment with 2 long apical brush-like setae [P1 setae without apical brush];
- P1 endopod distal segment with 2 long apical brush-like setae [P1 setae without apical brush].
4.2.2. Phylogenetic Status of the Rhizothrichidae inside the Cletodoidea
4.2.3. Phylogenetic Characterization of the Rhizothrichidae
Remarks on the Phylogenetic Relationships within the Rhizothrichidae
The monophylum Tryphoema Monard, 1926
The monophylum Rhizothrix Sars, 1909
Establishment of Monopenicillus gen. nov
4.2.4. Notes on Incongruent Characters
4.2.5. Remarks on Possible Sister–Group Relationships
A Possible Tryphoema–Rhizothrix Clade
A Possible Tryphoema–Monopenicillus gen. nov. Clade
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Por, F.D. A re-evaluation of the Cletodidae Sars, Lang (Copepoda, Harpacticoida). Syllogeous 1986, 58, 420–425. [Google Scholar]
- Wells, J.B.J. An annotated checklist and keys to the species of Copepoda Harpacticoida (Crustacea). Zootaxa 2007, 1568, 1–872. [Google Scholar] [CrossRef]
- Gómez, S. A new record and a new species of the genus Rhizothrix (Copepoda: Harpacticoida: Rhizothrichidae) from the deep sea of the Gulf of California. Proc. Biol. Soc. Wash. 2018, 131, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Walter, T.C.; Boxshall, G.A. Rhizothrix Sars G.O., 1909. In World Register of Marine Species; World of Copepods Database: 2023. Available online: https://www.marinespecies.org/aphia.php?p=taxdetails&id=534122 (accessed on 9 October 2023).
- Walter, T.C.; Boxshall, G.A. Tryphoema Monard, 1926. In World Register of Marine Species; World of Copepods Database: 2023. Available online: https://www.marinespecies.org/aphia.php?p=taxdetails&id=115437 (accessed on 9 October 2023).
- Arlt, G. Taxonomy and ecology of some Harpacticoids (Crustacea, Copepoda) in the Baltic Sea and Kattegat. Zool. Jahrb. Syst. 1983, 110, 45–85. [Google Scholar]
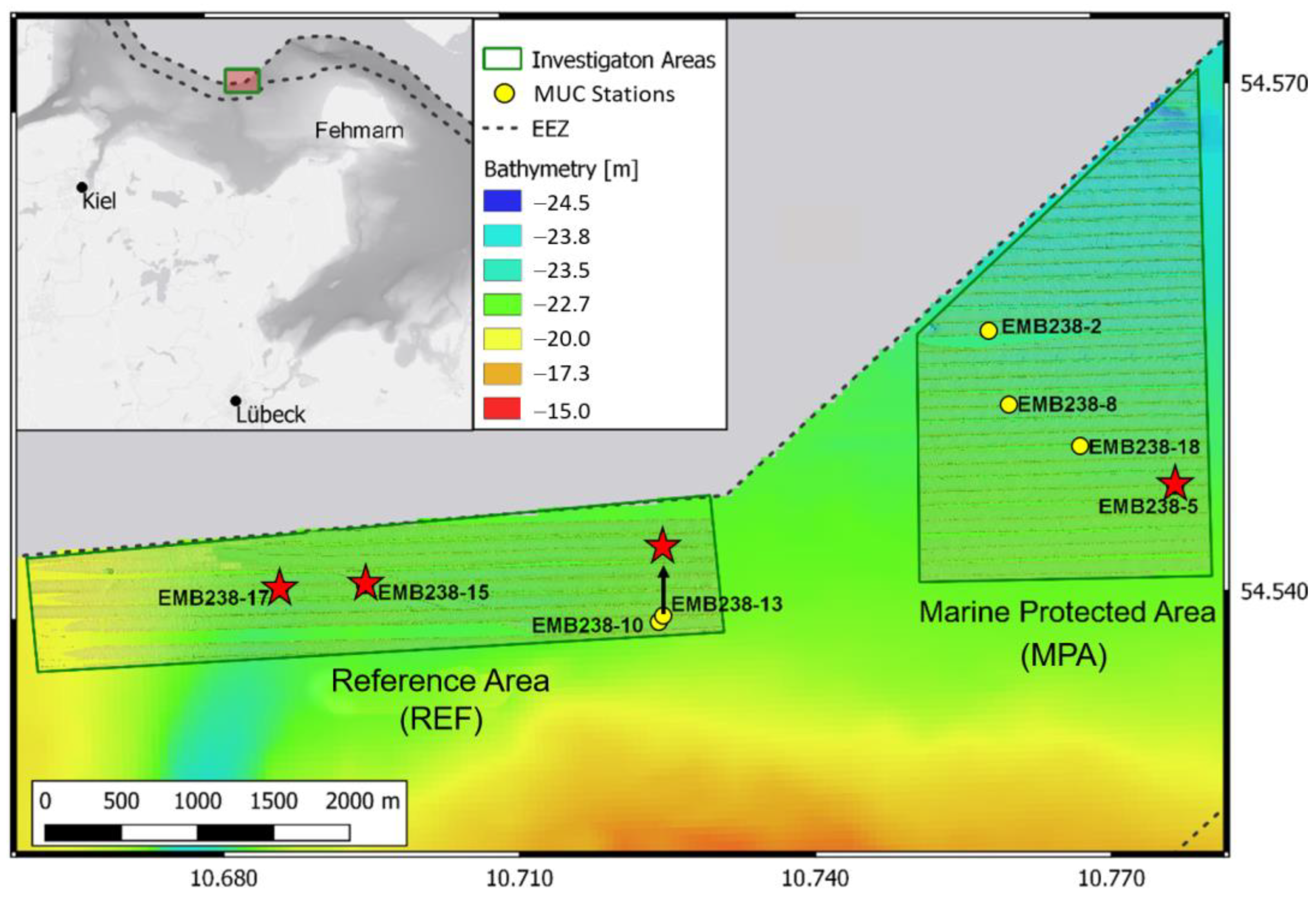
- Gogina, M.; Schönke, M. MGF-Ostsee Project: Potential effects of closure for bottom fishing in the marine protected areas (MPAs) of the western Baltic Sea—baseline observations, Cruise No. EMB238/Leg1+2, 26.05.2020–09.06.2020. In ELISABETH MANN BORGESE-Berichte; Institut für Ostseeforschung Warnemünde an der Universität Rostock: Rostock, Germany, 2020; pp. 1–43. [Google Scholar]
- Somerfield, P.J.; Warwick, R.M.; Moens, T. 6. Meiofauna techniques. In Methods for the Study of Marine Benthos; Eleftheriou, A., McIntyre, A., Eds.; Blackwell Scientific Ltd.: Oxford, UK, 2005; pp. 229–272. [Google Scholar]
- Lang, K. Monographie der Harpacticiden I & II; Hǎkan Ohlssons Boktryckeri: Lund, Sweden, 1948; pp. 1–1682. [Google Scholar]
- Huys, R.; Gee, J.M.; Moore, C.G.; Hamond, R. Marine and brackish water harpacticoid copepods. Part 1. Synop. Br. Fauna 1996, 51, 1–352. [Google Scholar]
- Boxshall, G.A.; Halsey, S.H. An Introduction to Copepod Diversity I & II; The Ray Society: London, UK, 2004; pp. 1–966. [Google Scholar]
- George, K.H.; Khodami, S.; Kihara, T.C.; Martínez Arbizu, P.; Martínez, A.; Mercado-Salas, N.F.; Pointner, K.; Veit-Köhler, G. Copepoda. In Guide to the Identification of Marine Meiofauna; Schmidt-Rhaesa, A., Ed.; Verlag Dr. Friedrich Pfeil: München, Germany, 2020; pp. 465–533. [Google Scholar]
- George, K.H.; Lehmanski, L.M.A.; Kihara, T.C. Revision of the genus Laophontodes T. Scott (Copepoda, Harpacticoida, Ancorabolidae), including the description of a new species and a key to species. ZooKeys 2020, 997, 17–46. [Google Scholar] [CrossRef]
- Hennig, W. Phylogenetische Systematik; Pareys Studientexte 34: Berlin/Hamburg, Germany, 1982; pp. 1–246. [Google Scholar]
- Ax, P. Das Phylogenetische System; Gustav Fischer Verlag: Stuttgart, Germany, 1984; pp. 1–349. [Google Scholar]
- Schminke, H.K. The ubiquitous telson and the deceptive furca. Crustaceana 1976, 30, 292–300. [Google Scholar] [CrossRef]
- Nam, E.J.; Lee, W. A new species of the genus Rhizothrix (Copepoda: Harpacticoida: Rhizothricidae) from Korean waters. Proc. Biol. Soc. Wash. 2005, 118, 692–705. [Google Scholar] [CrossRef]
- Packmor, J. Rhizothrichidae Por, 1986 (Copepoda: Harpacticoida) of Madeira and Porto Santo: Description of two new species of the genus Tryphoema Monard, 1926. Mar. Biodiv. 2013, 43, 341–361. [Google Scholar] [CrossRef]
- Alper, A.; Sak, S.; Metin, O. First record of the family Rhizothrichidae (Copepoda, Harpacticoida) from Turkey with description of a new species. Mar. Biodiv. 2018, 48, 357–365. [Google Scholar] [CrossRef]
- George, K.H. Restructuring the Ancorabolidae Sars (Copepoda, Harpacticoida) and Cletodidae T. Scott, with a new phylogenetic hypothesis regarding the relationships of the Laophontoidea T. Scott, Ancorabolidae and Cletodidae. Zoosyst Evol. 2020, 96, 455–498. [Google Scholar] [CrossRef]
- Lee, W.; Huys, R. New Normanellidae (Copepoda: Harpacticoida) from western Pacific cold seeps including a review of the genus Normanella. Cah. Biol. Mar. 1999, 40, 203–262. [Google Scholar]
- Galhano, M.H. Contribuição para o conhecimiento da fauna intersticial em Portugal. Publ. Inst. Zool. Dr. Augusto Nobre Fac. Ciênc. Porto 1970, 110, 9–207. [Google Scholar]
- Björnberg, T. Three new species of benthonic Harpacticoida (Copepoda, Crustacea) from São Sebastião Channel. Nauplius 2014, 22, 75–90. [Google Scholar] [CrossRef]
- Gómez, S. Three new species of Enhydrosoma and a new record of Enhydrosoma lacunae (Copepoda: Harpacticoida: Cletodidae) from the Eastern tropical Pacific. J. Crust. Biol. 2003, 23, 94–118. [Google Scholar] [CrossRef]
- Noodt, W. Die Copepoda Harpacticoidea des Brandungsstrandes von Teneriffa (Kanarische Inseln). Abh. Math.-Naturw. Kl. Akad. Wiss. Mainz 1958, 1958, 53–116. [Google Scholar]
- Mielke, W. Systematik der Copepoda eines Sandstrandes der Nordseeinsel Sylt. Mikrofauna Meeresbod. 1975, 52, 1–134. [Google Scholar]
- Lee, W.; Park, E.; Song, S.J. Marine Harpacticoida. Invertebr. Fauna Korea 2012, 21, 1–276. [Google Scholar]
- Huys, R.; Lee, W. On the relationships of the Normanellidae and the recognition of Cletopsyllidae grad. nov. (Copepoda, Harpacticoida). Zool. Anz. 1998/1999, 237, 267–290. [Google Scholar]
- Wells, J.B.J. The littoral Copepoda (Crustacea) of Inhaca Island, Mozambique. Trans. R. Soc. Edinb. 1967, 67, 189–358. [Google Scholar] [CrossRef]
- Wilson, C.B. The copepods of the Woods Hole region, Massachusetts. Bull. U.S. Natn. Mus. 1932, 158, 1–635. [Google Scholar] [CrossRef]
- Noodt, W. Marine Harpacticiden (Cop.) aus dem eulitoralen Sandstrand der Insel Sylt. Abh. Math.-Naturw. Kl. Akad. Wiss. Mainz 1952, 3, 105–142. [Google Scholar]
- Huys, R.; Boxshall, G.A. Copepod Evolution; The Ray Society: London, UK, 1991; pp. 1–468. [Google Scholar]
- Lang, K. Die während der schwedischen Expedition nach Spitzbergen 1898 und nach Grönland 1899 eingesammelten Harpacticiden. K. Sven. Vetenskakad. Handl. 3 1936, 15, 1–55. [Google Scholar]
- Bozic, B. Sur quelques copépodes harpacticoïdes des sables de Roscoff. Arch. Zool. Exp. Gen. 1953, 91, 17–24. [Google Scholar]
- Por, F.D. Harpacticoide noi (Crustacea, Copepoda) din Mîlurile Mârii negre. Acad. Republ. Pop. Rom. 1959, 11, 347–368. [Google Scholar]
- Coull, B.C. Meiobenthic Harpacticoida (Crustacea, Copepoda) from the North Carolina continental shelf. Cah. Biol. Mar. 1971, 12, 195–237. [Google Scholar]
- Bodin, P. Copépodes Harpacticoïdes marins des environs de la Rochelle. 5. Espéces nouvelles ou incertaines. Vie Milieu 1979, 27, 311–357. [Google Scholar]
- Kornev, P.N.; Chertoprud, E.S. Harpacticoid Copepods from the White Sea: Morphology, Systematics, Ecology; KMK Scientific Press: Moscow, Russia, 2008; pp. 1–367. (In Russian) [Google Scholar]
- Monard, A. Descriptions de quelques espèces nouvelles d’harpacticides marins de la région de Banyuls. Rev. Suisse Zool. 1926, 33, 619–628. [Google Scholar]
- Por, F.D. A study of the Levantine and Pontic Harpacticoida (Crustacea, Copepoda). Zool. Verh. 1964, 64, 3–128. [Google Scholar]
- Wells, J.B.J. New and rare Copepoda Harpacticoida From the Isles of Scilly. J. Nat. Hist. 1968, 2, 397–424. [Google Scholar] [CrossRef]
- Pennak, R.W. Harpacticoid copepods from some intertidal beaches near Woods Hole, Massachusetts. Trans. Am. Microsc. Soc. 1942, 61, 274–285. [Google Scholar] [CrossRef]
- Wells, J.B.J.; Clark, M.E. The interstitial Crustacea of two beaches in Portugal. Rev. Biol. 1965, 5, 87–108. [Google Scholar]
- Huys, R. A new family of harpacticoid copepods and an analysis of the phylogenetic relationships within the Laophontoidea T. Scott. Bijdr. Dierk. 1990, 60, 79–120. [Google Scholar] [CrossRef]
- Boxshall, G.A.; Ferrari, F.D.; Tiemann, H. The ancestral copepod: Towards a consensus of opinion at the First International Conference on Copepoda. Crustaceana 1984, 7, 68–84. [Google Scholar]
- Wägele, J.-W. Grundlagen der Phylogenetischen Systematik; Verlag Dr. Friedrich Pfeil: München, Germany, 2001; pp. 1–320. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exopod | Endopod | |
|---|---|---|
| P2 | I; I; II–2–1 | 0–1 |
| P3 | I; I; II–2–1 | 0–1 |
| P4 | I; I–1; II–2–2 | 0–1 |
| No. | Character/Taxon (Apomorphy = 1, Plesiomorphy = 0) | Cletodoidea | Tryphoema | Rhizothrix | Monopenicillus gen. nov. |
|---|---|---|---|---|---|
| 1*–13* | Thirteen autapomorphies of Cletodoidea listed by George (2020), shared with Rhizotrichidae | 1 | 1 | 1 | 1 |
| 14 | P2 exp-3 with at most 1 inner seta [with 2 setae] | 1 | 1 | 1 | 1 |
| 15 | P3 exp-3 with at most 1 inner seta [with 2 setae] | 1 | 1 | 1 | 1 |
| 16 | P3 distal endopodal segment with at most 1 inner seta [with at least 2 inner setae] | 1 | 1 | 1 | 1 |
| 17 | P4 distal endopodal segment with at most 1 inner seta [with at least 2 inner setae] | 1 | 1 | 1 | 1 |
| 18 | Female A1 distal segment (sub-)apically with 1 strong spinulose element [no such element developed] | 0 | 1 | 1 | 1 |
| 19 | P1 exp-3 with at least 1 long seta with apical brush [P1 exp-3 without brush-like setae] | 0 | 1 | 1 | 1 |
| 20 | P1 enp-2 with at least 1 inner apical seta brush-like [seta without apical brush] | 0 | 1 | 1 | 1 |
| 21 | Furcal seta III absent [seta III still present] | 0 | 1 | 0 | 0 |
| 22 | P2 exopod 2-segmented [exopod 3-segmented] | 0 | 1 | 0 | 0 |
| 23 | P3 exopod 2-segmented [exopod 3-segmented] | 0 | 1 | 0 | 0 |
| 24 | P4 exopod 2-segmented [exopod 3-segmented] | 0 | 1 | 0 | 0 |
| 25 | P2 distal exopodal segment without inner seta [with at least 1 inner seta] | 0 | 1 | 0 | 0 |
| 26 | P3 distal exopodal segment without inner seta [with inner seta] | 0 | 1 | 0 | 0 |
| 27 | P4 distal exopodal segment without inner seta [with at least 1 inner seta] | 0 | 1 | 0 | 0 |
| 28 | Female P5 basoendopods fused medially [separated] | 0 | 0 | 1 | 0 |
| 29 | A2 lacking abexopodal seta on allobasis [abexopodal seta still present] | 0 | 0 | 0 | 1 |
| 30 | P1 enp-2 lacking inner subapical seta [inner subapical seta still present] | 0 | 0 | 0 | 1 |
| 31 | P2 distal endopodal segment with 1 seta [with at least 2 setae] | 0 | 0 | 0 | 1 |
| 32 | P3 distal endopodal segment with 1 seta [with at least 2 setae] | 0 | 0 | 0 | 1 |
| 33 | P4 distal endopodal segment with 1 seta [with at least 2 setae] | 0 | 0 | 0 | 1 |
| 34 | P4 endopod minute, knob-like [endopod at least two times longer than broad] | 0 | 0 | 0 | 1 |
| 35 | Female P5 exopod fused with basoendopod [exopod distinct] | 0 | 0 | 0 | 1 |
| Incongruent characters | |||||
| 36 | Body densely covered by pubescense [no pubescense developed] | 0 | 1 | 0 | 0 |
| 37 | A2 exopod 1-segmented [exopod at least 2-segmented] | 1 | 1 | 0 | 0 |
| 38 | P1 enp-1 without inner setae [with inner seta] | 1 | 1 | 1 | 0 |
| 39 | P1 enp-2 outer apical seta brush-like [seta without apical brush] | 0 | 1 | 1 | 0 |
| 40 | P2 endopod 1-segmented [endopod 2-segmented] | 0 | 1 | 0 | 1 |
| 41 | P3 endopod 1-segmented [endopod 2-segmented] | 0 | 1 | 0 | 1 |
| 42 | P4 endopod 1-segmented [endopod 2-segmented] | 0 | 1 | 0 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
George, K.H.; Zey, A.; Packmor, J. The Relationship between the Rhizothrichidae Por (Copepoda: Harpacticoida) and the Cletodoidea Bowman & Abele, Including the Establishment of a New Genus and the Description of a New Species. Taxonomy 2023, 3, 528-550. https://doi.org/10.3390/taxonomy3040030
George KH, Zey A, Packmor J. The Relationship between the Rhizothrichidae Por (Copepoda: Harpacticoida) and the Cletodoidea Bowman & Abele, Including the Establishment of a New Genus and the Description of a New Species. Taxonomy. 2023; 3(4):528-550. https://doi.org/10.3390/taxonomy3040030
Chicago/Turabian StyleGeorge, Kai Horst, Andreina Zey, and Jana Packmor. 2023. "The Relationship between the Rhizothrichidae Por (Copepoda: Harpacticoida) and the Cletodoidea Bowman & Abele, Including the Establishment of a New Genus and the Description of a New Species" Taxonomy 3, no. 4: 528-550. https://doi.org/10.3390/taxonomy3040030
APA StyleGeorge, K. H., Zey, A., & Packmor, J. (2023). The Relationship between the Rhizothrichidae Por (Copepoda: Harpacticoida) and the Cletodoidea Bowman & Abele, Including the Establishment of a New Genus and the Description of a New Species. Taxonomy, 3(4), 528-550. https://doi.org/10.3390/taxonomy3040030