
Effect of CdSTe QDs’ Crystal Size on Viability and Cytochrome P450 Activity of CHO-K1 and HEP-G2 Cells



Abstract
1. Introduction
2. Materials and Methods
2.1. Microwave-Assisted Synthesis and Characterization of CdSTe QDs
2.2. Cell Culture
2.3. Fluorescence Microscopy
2.4. Cell Viability Using Trypan Blue Exclusion Assay
2.5. CellTiter-Glo® Luminescent Cell Viability Assay
2.6. RealTime-Glo™ MT Cell Viability Assay
2.7. Apoptosis and Necrosis Assay
2.8. P450-Glo™ Assays
2.9. Statistical Analysis
3. Results and Discussion
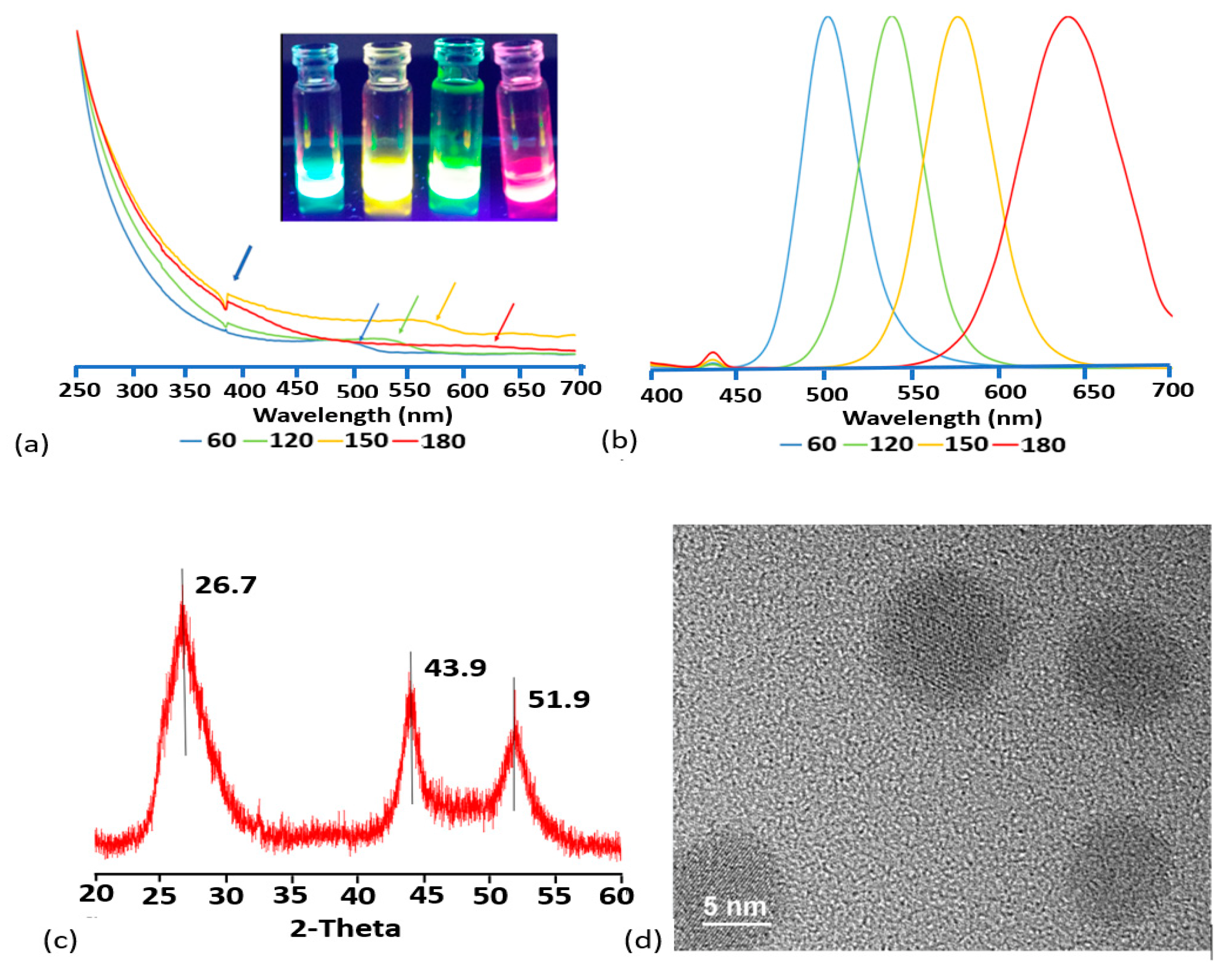
3.1. Synthesis and Characterization of CdSTe QDs
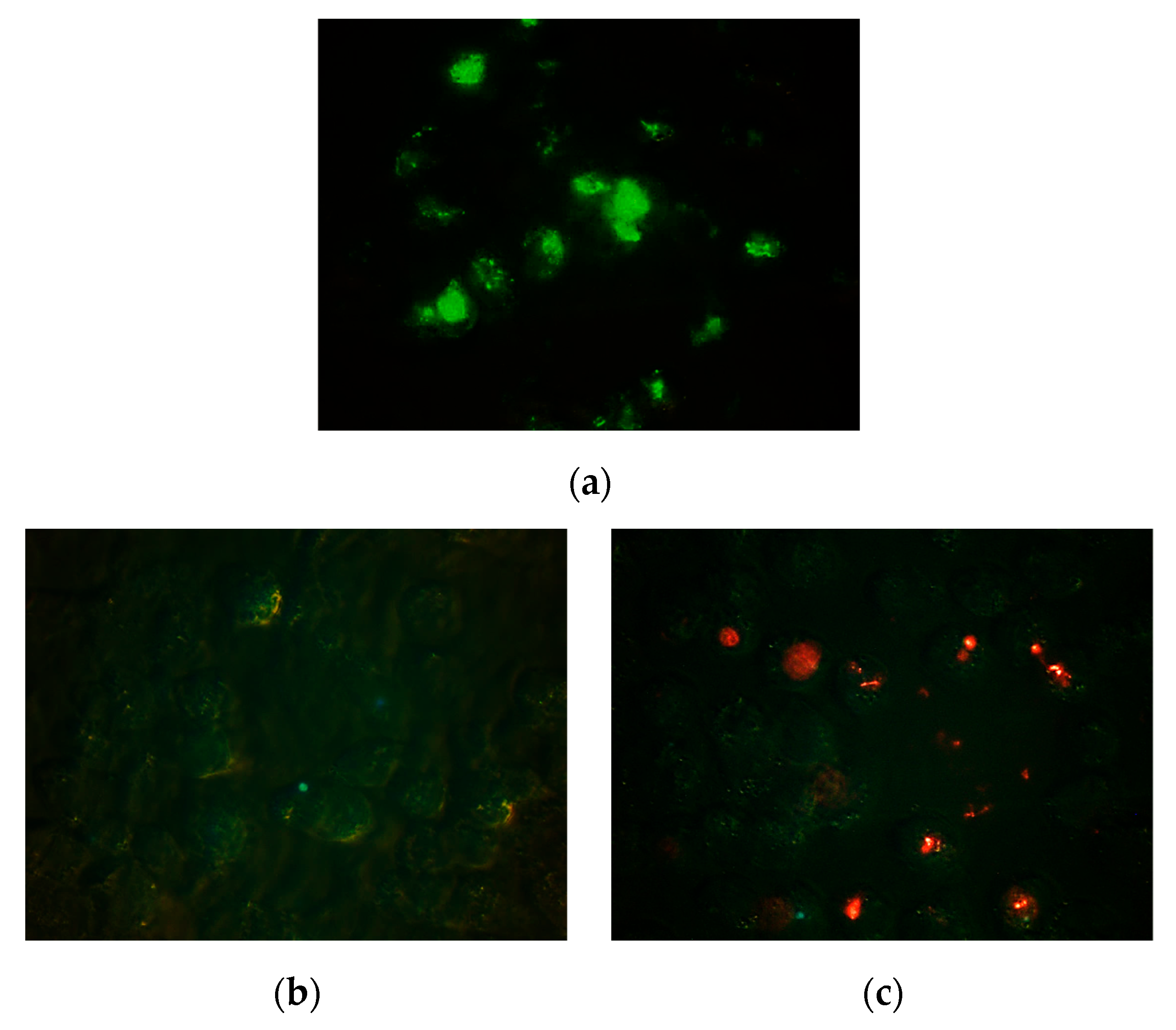
3.2. Fluorescence Microscopy
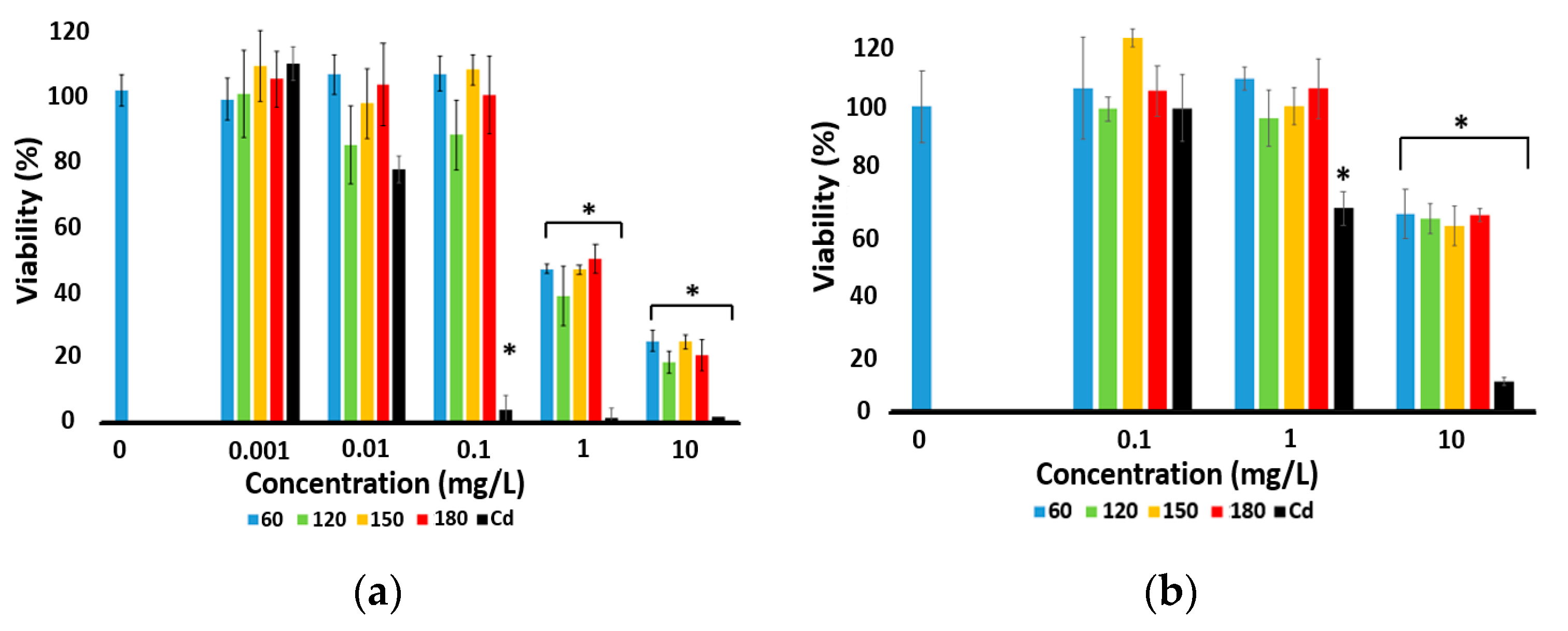
3.3. Viability Assays
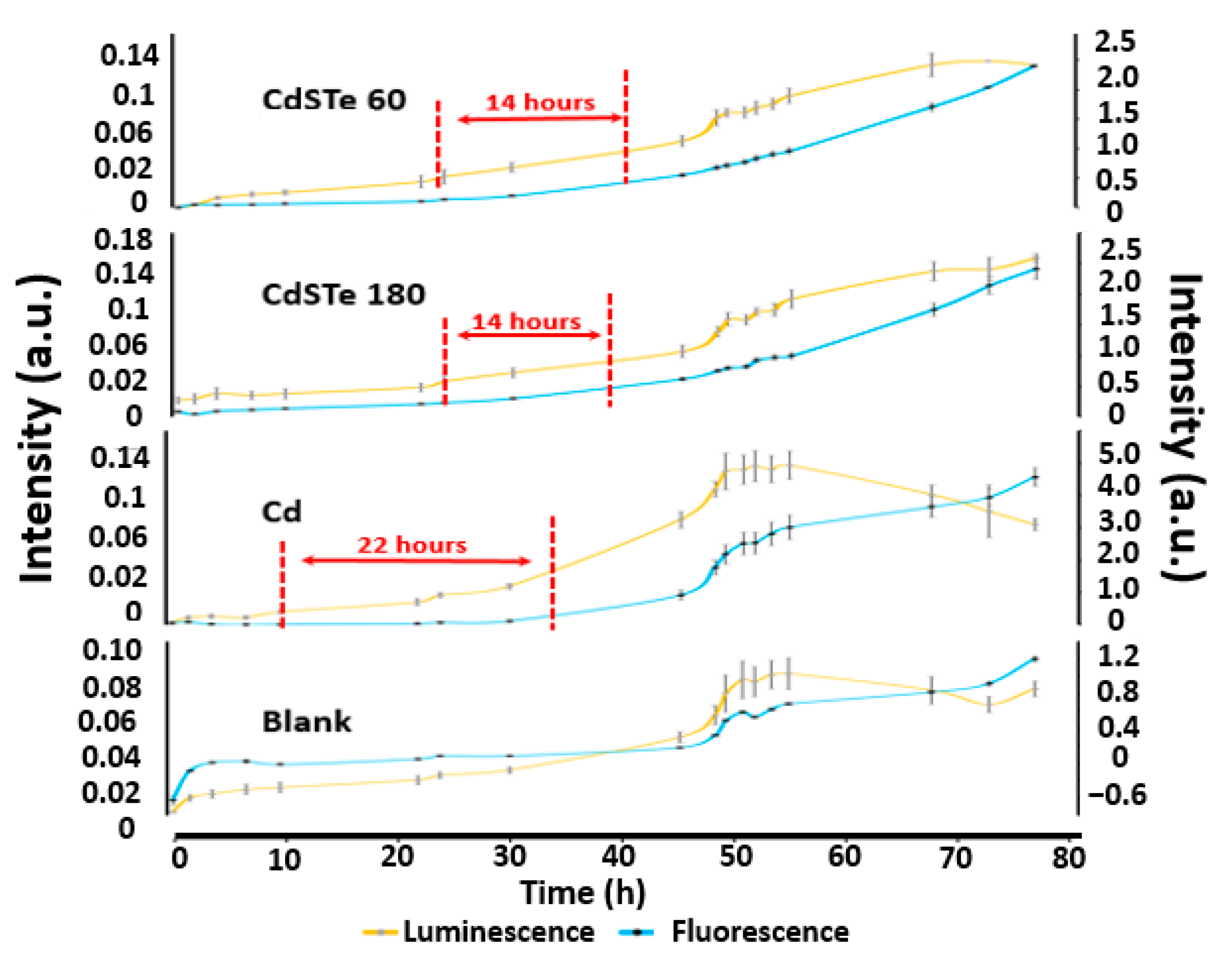
3.4. Apoptosis and Necrosis Assay
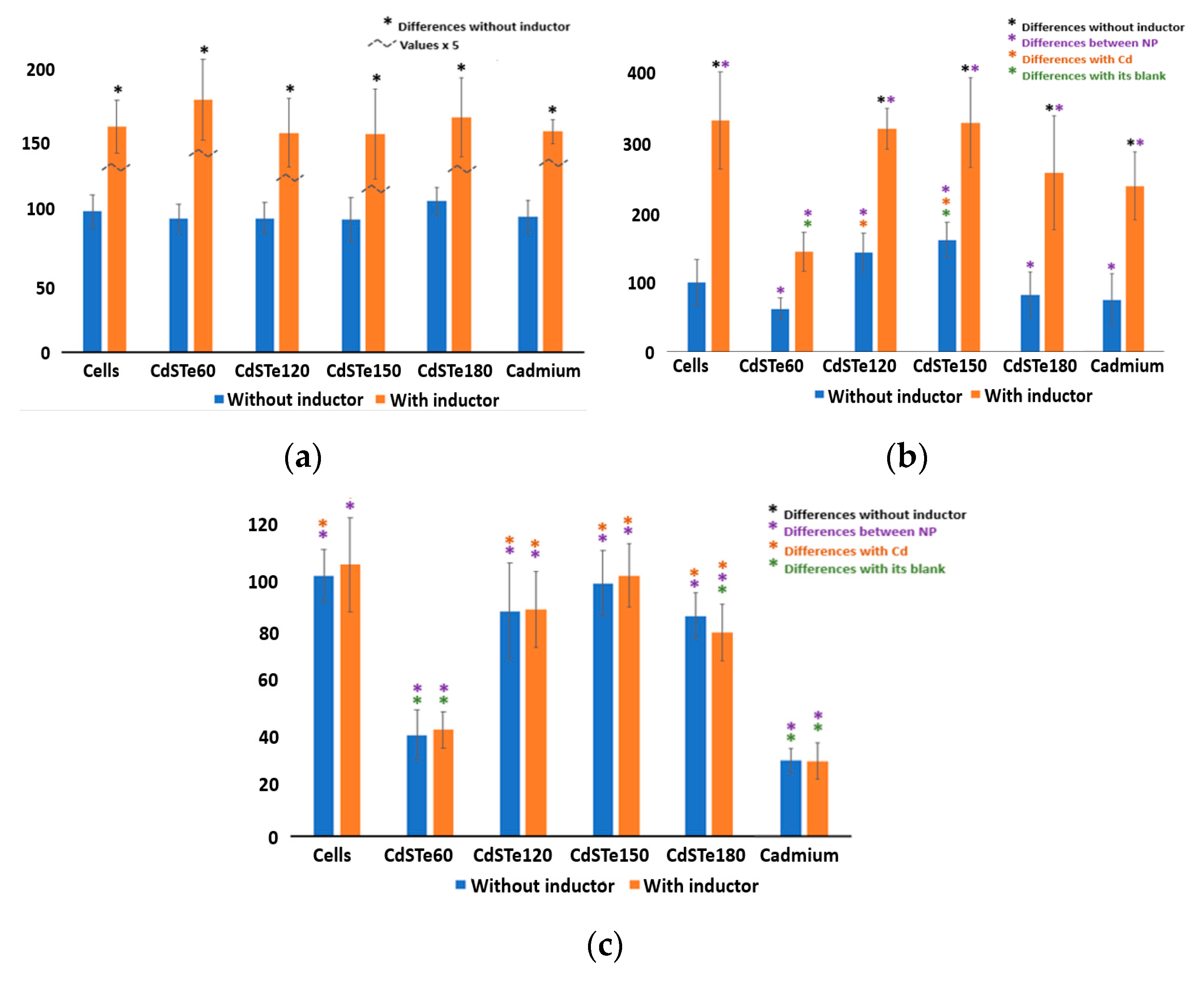
3.5. The Activity of CYP450 Isoenzymes
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, J.; Zhou, X.; Ni, S.; Wang, X. One-pot synthesis of strongly luminesencing CdTe quantum dots and their conjugation with mouse antibody to alpha-fetoprotein. Colloid J. 2010, 72, 710–715. [Google Scholar] [CrossRef]
- Alamo-Nole, L.; Bailon-Ruiz, S.; Cruz-Acuña, R.; Perales-Pérez, O.; Román, F.R. Quantum dots of ZnSe(S) doped with copper as nanophotocatalyst in the degradation of organic dyes. J. Nanosci. Nanotechnol. 2014, 14, 7333–7339. [Google Scholar] [CrossRef]
- Muhammad, F.; Tahir, M.; Zeb, M.; Wahab, F.; Kalasad, M.N.; Khan, D.N.; Karimov, K.S. Cadmium selenide quantum dots: Synthesis, characterization and their humidity and temperature sensing properties with poly-(dioctylfluorene). Sens. Actuators B Chem. 2019, 285, 504–512. [Google Scholar] [CrossRef]
- Auffan, M.; Rose, J.; Wiesner, M.R.; Bottero, J.-Y. Chemical stability of metallic nanoparticles: A parameter controlling their potential cellular toxicity in vitro. Environ. Pollut. 2009, 157, 1127–1133. [Google Scholar] [CrossRef]
- Sotomayor, C.G.; Groothof, D.; Vodegel, J.J.; Eisenga, M.F.; Knobbe, T.J.; IJmker, J.; Lammerts, R.G.M.; de Borst, M.H.; Berger, S.P.; Nolte, I.M.; et al. Plasma cadmium is associated with increased risk of long-term kidney graft failure. Kidney Int. 2021, 99, 1213–1224. [Google Scholar] [CrossRef]
- Abbas, Q.; Yousaf, B.; Amina; Ali, M.U.; Munir, M.A.M.; El-Naggar, A.; Rinklebe, J.; Naushad, M. Transformation pathways and fate of engineered nanoparticles (ENPs) in distinct interactive environmental compartments: A review. Environ. Int. 2020, 138, 105646. [Google Scholar] [CrossRef]
- Malakar, A.; Kanel, S.R.; Ray, C.; Snow, D.D.; Nadagouda, M.N. Nanomaterials in the environment, human exposure pathway, and health effects: A review. Sci. Total Environ. 2021, 759, 143470. [Google Scholar] [CrossRef]
- Naito, M.; Yokoyama, T.; Hosokawa, K.; Nogi, K. (Eds.) Chapter 1—Basic Properties and Measuring Methods of Nanoparticles. In Nanoparticle Technology Handbook, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 3–47. ISBN 978-0-444-64110-6. [Google Scholar]
- Rivera-Rodriguez, G.; Peralez-Perez, O.; Su, Y.-F.; Alamo-Nole, L. Effect of the Reaction Temperature on the Optical Properties of CdSTe Quantum Dots Synthesized Under Microwave Irradiation. MRS Adv. 2016, 1, 2207–2212. [Google Scholar] [CrossRef]
- Sharifi, S.; Behzadi, S.; Laurent, S.; Laird Forrest, M.; Stroeve, P.; Mahmoudi, M. Toxicity of nanomaterials. Chem. Soc. Rev. 2012, 41, 2323–2343. [Google Scholar] [CrossRef]
- Behzadi, S.; Serpooshan, V.; Tao, W.; Hamaly, M.A.; Alkawareek, M.Y.; Dreaden, E.C.; Brown, D.; Alkilany, A.M.; Farokhzad, O.C.; Mahmoudi, M. Cellular uptake of nanoparticles: Journey inside the cell. Chem. Soc. Rev. 2017, 46, 4218–4244. [Google Scholar] [CrossRef]
- Demir, E.; Qin, T.; Li, Y.; Zhang, Y.; Guo, X.; Ingle, T.; Yan, J.; Orza, A.I.; Biris, A.S.; Ghorai, S.; et al. Cytotoxicity and genotoxicity of cadmium oxide nanoparticles evaluated using in vitro assays. Mutat. Res. Toxicol. Environ. Mutagen. 2020, 850–851, 503149. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chen, S.; Ding, Y.; Zhu, Q.; Zhang, N.; Yu, S. Biofabrication of morphology improved cadmium sulfide nanoparticles using Shewanella oneidensis bacterial cells and ionic liquid: For toxicity against brain cancer cell lines. J. Photochem. Photobiol. B Biol. 2018, 178, 424–427. [Google Scholar] [CrossRef] [PubMed]
- Kaviyarasu, K.; Kanimozhi, K.; Matinise, N.; Maria Magdalane, C.; Mola, G.T.; Kennedy, J.; Maaza, M. Antiproliferative effects on human lung cell lines A549 activity of cadmium selenide nanoparticles extracted from cytotoxic effects: Investigation of bio-electronic application. Mater. Sci. Eng. C 2017, 76, 1012–1025. [Google Scholar] [CrossRef] [PubMed]
- Lovrić, J.; Cho, S.J.; Winnik, F.M.; Maysinger, D. Unmodified cadmium telluride quantum dots induce reactive oxygen species formation leading to multiple organelle damage and cell death. Chem. Biol. 2005, 12, 1227–1234. [Google Scholar] [CrossRef] [PubMed]
- Lovrić, J.; Bazzi, H.S.; Cuie, Y.; Fortin, G.R.A.; Winnik, F.M.; Maysinger, D. Differences in subcellular distribution and toxicity of green and red emitting CdTe quantum dots. J. Mol. Med. 2005, 83, 377–385. [Google Scholar] [CrossRef]
- Luo, Y.-H.; Wu, S.-B.; Wei, Y.-H.; Chen, Y.-C.; Tsai, M.-H.; Ho, C.-C.; Lin, S.-Y.; Yang, C.-S.; Lin, P. Cadmium-Based Quantum Dot Induced Autophagy Formation for Cell Survival via Oxidative Stress. Chem. Res. Toxicol. 2013, 26, 662–673. [Google Scholar] [CrossRef]
- Omasa, T.; Onitsuka, M.; Kim, W.-D. Cell engineering and cultivation of chinese hamster ovary (CHO) cells. Curr. Pharm. Biotechnol. 2010, 11, 233–240. [Google Scholar] [CrossRef]
- Donato, M.T.; Tolosa, L.; Gómez-Lechón, M.J. Culture and Functional Characterization of Human Hepatoma HepG2 Cells. Methods Mol. Biol. 2015, 1250, 77–93. [Google Scholar] [CrossRef]
- Guengerich, F.P. Common and Uncommon Cytochrome P450 Reactions Related to Metabolism and Chemical Toxicity. Chem. Res. Toxicol. 2001, 14, 611–650. [Google Scholar] [CrossRef]
- Furge, L.L.; Guengerich, F.P. Cytochrome P450 enzymes in drug metabolism and chemical toxicology: An introduction. Biochem. Mol. Biol. Educ. 2006, 34, 66–74. [Google Scholar] [CrossRef]
- Strober, W. Trypan Blue Exclusion Test of Cell Viability. Curr. Protoc. Immunol. 2015, 111, A3.B.1–A3.B.3. [Google Scholar] [CrossRef] [PubMed]
- Derfus, A.M.; Chan, W.C.W.; Bhatia, S.N. Probing the Cytotoxicity of Semiconductor Quantum Dots. Nano Lett. 2004, 4, 11–18. [Google Scholar] [CrossRef]
- Wang, L.; Nagesha, D.K.; Selvarasah, S.; Dokmeci, M.R.; Carrier, R.L. Toxicity of CdSe nanoparticles in Caco-2 cell cultures. J. Nanobiotech. 2008, 6, 11. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Gao, C. Recent advances in interactions of designed nanoparticles and cells with respect to cellular uptake, intracellular fate, degradation and cytotoxicity. Nanotechnology 2016, 27, 412002. [Google Scholar] [CrossRef]
- Shukla, V.; Mishra, S.K.; Pant, H.C. Oxidative stress in neurodegeneration. Adv. Pharmacol. Sci. 2011, 2011, 572634. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zhao, H.; Wang, Y.; Jiang, C.; Xia, P.; Gu, J.; Liu, X.; Bian, J.; Yuan, Y.; Liu, Z. Calcium—Calmodulin signaling elicits mitochondrial dysfunction and the release of cytochrome c during cadmium-induced apoptosis in primary osteoblasts. Toxicol. Lett. 2014, 224, 1–6. [Google Scholar] [CrossRef]
- Messner, B.; Türkcan, A.; Ploner, C.; Laufer, G.; Bernhard, D. Cadmium overkill: Autophagy, apoptosis and necrosis signalling in endothelial cells exposed to cadmium. Cell. Mol. Life Sci. 2016, 73, 1699–1713. [Google Scholar] [CrossRef]
- Bozcaarmutlu, A. Effect of mercury, cadmium, nickel, chromium and zinc on kinetic properties of NADPH-cytochrome P450 reductase purified from leaping mullet (Liza saliens). Toxicol. Vitr. 2007, 21, 408–416. [Google Scholar] [CrossRef]
- Zhang, L.; Gan, J.; Ke, C.; Liu, X.; Zhao, J.; You, L.; Yu, J.; Wu, H. Identification and expression profile of a new cytochrome P450 isoform (CYP414A1) in the hepatopancreas of Venerupis (Ruditapes) philippinarum exposed to benzo[a]pyrene, cadmium and copper. Environ. Toxicol. Pharmacol. 2012, 33, 85–91. [Google Scholar] [CrossRef]
- Baker, J.R.; Satarug, S.; Edwards, R.J.; Moore, M.R.; Williams, D.J.; Reilly, P.E.B. Potential for early involvement of CYP isoforms in aspects of human cadmium toxicity. Toxicol. Lett. 2003, 137, 85–93. [Google Scholar] [CrossRef]
- Chen, N.; He, Y.; Su, Y.; Li, X.; Huang, Q.; Wang, H.; Zhang, X.; Tai, R.; Fan, C. The cytotoxicity of cadmium-based quantum dots. Biomaterials 2012, 33, 1238–1244. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assay | Trypan Blue | AMP Kit | MT Kit | |||
|---|---|---|---|---|---|---|
| CdSTe QDs | 60 | 120 | 60 | 120 | 60 | 120 |
| 0 | 100 ± 6 | 100 ± 5 | 100 ± 10 | |||
| 0.01 | 105 ± 6 | 83 ± 12 | 91 ± 8 | 93 ± 11 | ||
| 0.1 | 101 ± 2 | 102 ± 5 | 105 ± 5 | 87 ± 10 | 99 ± 14 | 101 ± 10 |
| 1.0 | 70 ± 9 | 69 ± 12 | 46 ± 1 | 38 ± 9 | 76 ± 17 | 67 ± 13 |
| 10 | 62 ± 7 | 66 ± 12 | 24 ± 3 | 18 ± 3 | 6 ± 2 | 14 ± 5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alamo-Nole, L.; Ponton-Almodovar, A.; Ortiz-Laboy, I. Effect of CdSTe QDs’ Crystal Size on Viability and Cytochrome P450 Activity of CHO-K1 and HEP-G2 Cells. Micro 2023, 3, 308-319. https://doi.org/10.3390/micro3010021
Alamo-Nole L, Ponton-Almodovar A, Ortiz-Laboy I. Effect of CdSTe QDs’ Crystal Size on Viability and Cytochrome P450 Activity of CHO-K1 and HEP-G2 Cells. Micro. 2023; 3(1):308-319. https://doi.org/10.3390/micro3010021
Chicago/Turabian StyleAlamo-Nole, Luis, Adriana Ponton-Almodovar, and Ivan Ortiz-Laboy. 2023. "Effect of CdSTe QDs’ Crystal Size on Viability and Cytochrome P450 Activity of CHO-K1 and HEP-G2 Cells" Micro 3, no. 1: 308-319. https://doi.org/10.3390/micro3010021
APA StyleAlamo-Nole, L., Ponton-Almodovar, A., & Ortiz-Laboy, I. (2023). Effect of CdSTe QDs’ Crystal Size on Viability and Cytochrome P450 Activity of CHO-K1 and HEP-G2 Cells. Micro, 3(1), 308-319. https://doi.org/10.3390/micro3010021