
1H-NMR-Based Plasma Metabolomic Profiling of Crossbred Beef Cattle with Divergent RFI Phenotype

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Feeding, RFI Determination
2.2. NMR-Based Metabolome Analysis of Plasma Samples
2.3. Data Modelling and Statistical Analysis
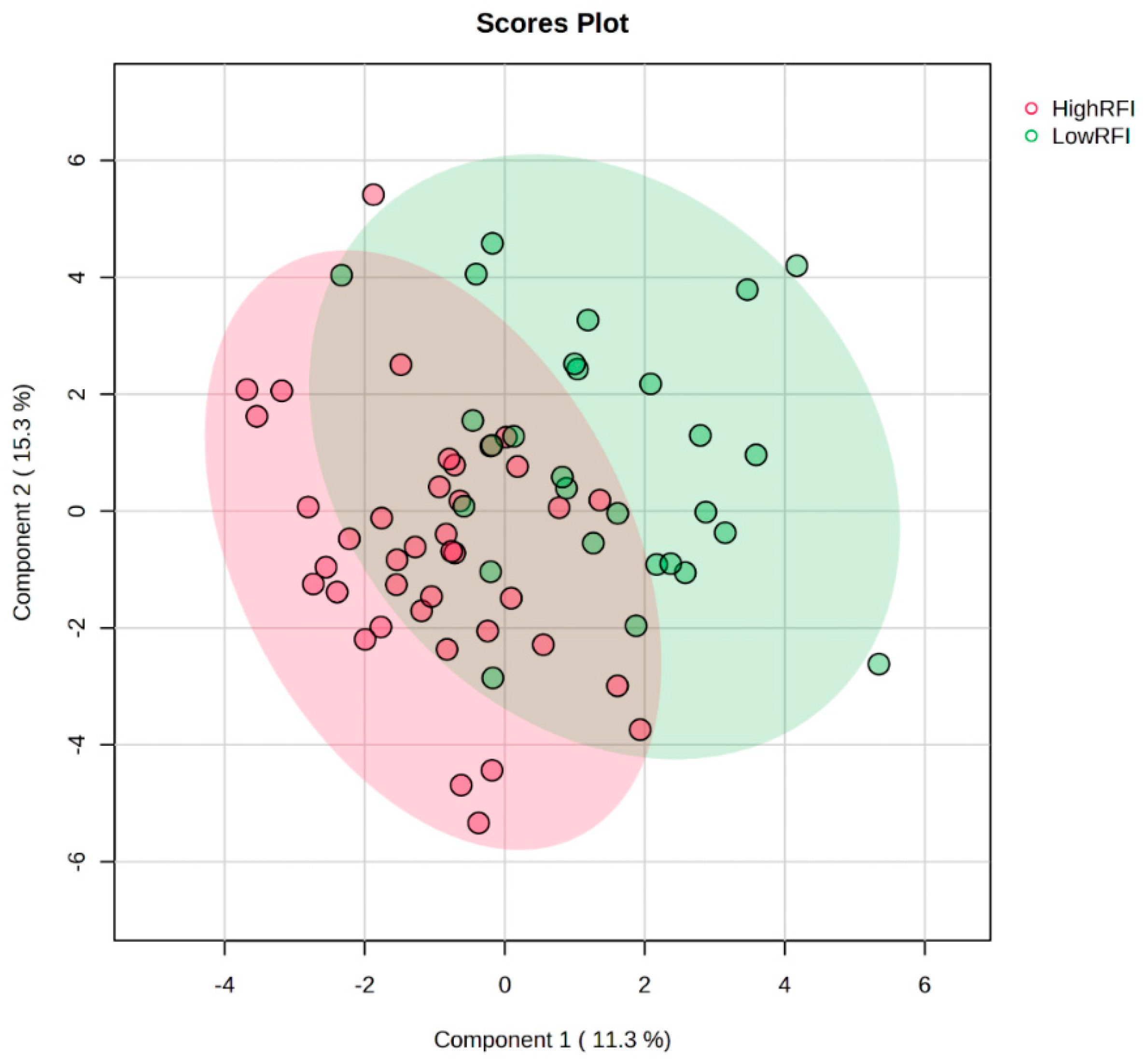
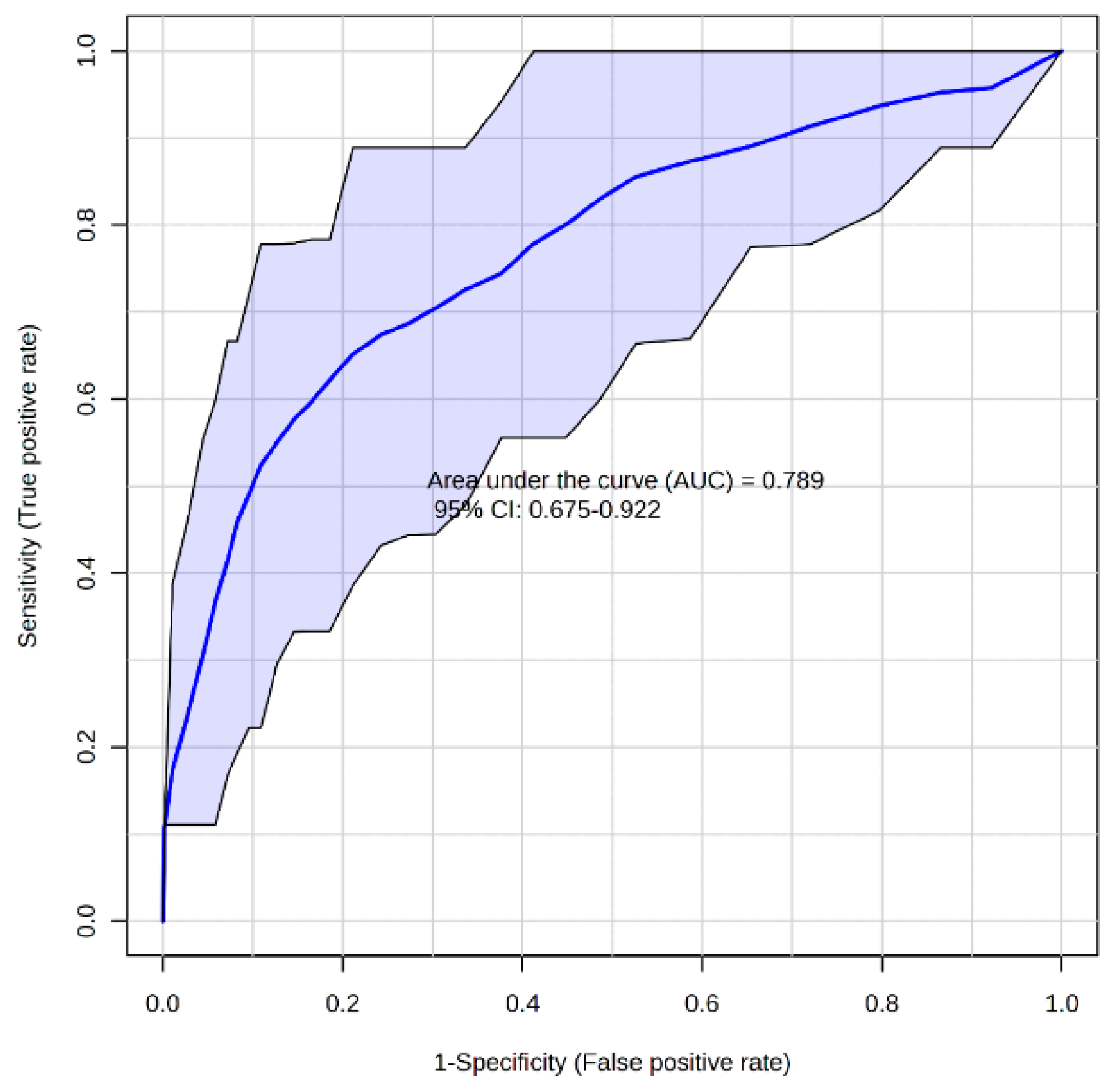
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arthur, P.F.; Archer, J.A.; Johnston, D.J.; Herd, R.M.; Richardson, E.C.; Parnell, P.F. Genetic and phenotypic variance and covariance components for feed intake, feed efficiency, and other postweaning traits in Angus cattle. J. Anim. Sci. 2001, 79, 2805. [Google Scholar] [CrossRef]
- Maia de Souza, D.; Petre, R.; Jackson, F.; Hadarits, M.; Pogue, S.; Carlyle, C.N.; Bork, E.; McAllister, T. A Review of Sustainability Enhancements in the Beef Value Chain: State-of-the-Art and Recommendations for Future Improvements. Animals 2017, 7, 26. [Google Scholar] [CrossRef]
- Taiwo, G.; Idowu, M.; Collins, S.; Sidney, T.; Wilson, M.; Pech-Cervantes, A.; Ogunade, I.M. Chemical Group-Based Metabolome Analysis Identifies Candidate Plasma Biomarkers Associated with Residual Feed Intake in Beef Steers. Front. Anim. Sci. 2022, 2, 783314. [Google Scholar] [CrossRef]
- Koch, R.M.; Swiger, L.A.; Chambers, D.; Gregory, K.E. Efficiency of Feed Use in Beef Cattle. J. Anim. Sci. 1963, 22, 486–494. [Google Scholar] [CrossRef]
- Archer, J.A.; Richardson, E.C.; Herd, R.M.; Arthur, P.F. Potential for selection to improve efficiency of feed use in beef cattle: A review. Aust. J. Agric. Res. 1999, 50, 147. [Google Scholar] [CrossRef]
- Richardson, E.C.; Herd, R.M.; Oddy, V.H.; Thompson, J.M.; Archer, J.A.; Arthur, P.F. Body composition and implications for heat production of Angus steer progeny of parents selected for and against residual feed intake. Aust. J. Exp. Agric. 2001, 41, 1065. [Google Scholar] [CrossRef]
- Wang, J.H.; Byun, J.; Pennathur, S. Analytical approaches to metabolomics and applications to systems biology. Semin. Nephrol. 2010, 30, 500–511. [Google Scholar] [CrossRef]
- Qiu, S.; Cai, Y.; Yao, H.; Lin, C.; Xie, Y.; Tang, S.; Zhang, A. Small molecule metabolites: Discovery of biomarkers and therapeutic targets. Signal Transduct. Target. Ther. 2023, 8, 132. [Google Scholar] [CrossRef]
- Jones, O.A.; Cheung, V.L. An introduction to metabolomics and its potential application in veterinary science. Comp. Med. 2007, 57, 436–442. [Google Scholar] [PubMed]
- Goldansaz, S.A.; Guo, A.C.; Sajed, T.; Steele, M.A.; Plastow, G.S.; Wishart, D.S. Livestock metabolomics and the livestock metabolome: A systematic review. PLoS ONE 2017, 12, e0177675. [Google Scholar] [CrossRef] [PubMed]
- Casadei-Gardini, A.; Del Coco, L.; Marisi, G.; Conti, F.; Rovesti, G.; Ulivi, P.; Canale, M.; Frassineti, G.L.; Foschi, F.G.; Longo, S.; et al. (1)H-NMR Based Serum Metabolomics Highlights Different Specific Biomarkers between Early and Advanced Hepatocellular Carcinoma Stages. Cancers 2020, 12, 241. [Google Scholar] [CrossRef]
- Karisa, B.; Moore, S.; Plastow, G. Analysis of biological networks and biological pathways associated with residual feed intake in beef cattle. Anim. Sci. J. 2013, 85, 374–387. [Google Scholar] [CrossRef] [PubMed]
- Jorge-Smeding, E.; Renand, G.; Centeno, D.; Pétéra, M.; Durand, S.; Polakof, S.; Cantalapiedra-Hijar, G. Metabolomics reveals changes in urea cycle associated to residual feed intake in growing heifers. In Energy and Protein Metabolism and Nutrition; Wageningen Academic Publishers: Wageningen, The Netherlands, 2019. [Google Scholar]
- Xue, M.Y.; Xie, Y.Y.; Zhong, Y.; Ma, X.J.; Sun, H.Z.; Liu, J.X. Integrated meta-omics reveals new ruminal microbial features associated with feed efficiency in dairy cattle. Microbiome 2022, 10, 32. [Google Scholar] [CrossRef] [PubMed]
- Kaur, H.; Kaur, G.; Gupta, T.; Mittal, D.; Ali, S.A. Integrating Omics Technologies for a Comprehensive Understanding of the Microbiome and Its Impact on Cattle Production. Biology 2023, 12, 1200. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Bu, D.; Ma, L. Integration of multiplied omics, a step forward in systematic dairy research. Metabolites 2022, 12, 225. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Mukiibi, R.; Wang, Y.; Plastow, G.S.; Li, C. Identification of candidate genes and enriched biological functions for feed efficiency traits by integrating plasma metabolites and imputed whole genome sequence variants in beef cattle. BMC Genom. 2021, 22, 832. [Google Scholar] [CrossRef] [PubMed]
- Foroutan, A.; Fitzsimmons, C.; Mandal, R.; Piri-Moghadam, H.; Zheng, J.; Guo, A.; Li, C.; Guan, L.L.; Wishart, D.S. The Bovine Metabolome. Metabolites 2020, 10, 233. [Google Scholar] [CrossRef] [PubMed]
- Pal, M.; Bharati, P. Introduction to Correlation and Linear Regression Analysis. In Applications of Regression Techniques; Springer: Singapore, 2019; pp. 1–18. [Google Scholar]
- Wells, R.S.; Interrante, S.M.; Sakkuma, S.S.; Walker, R.S.; Butler, T.J. Accuracy of the VYTELLE SENSE In-Pen Weighing Positions. Appl. Anim. Sci. 2021, 37, 626–634. [Google Scholar] [CrossRef]
- MacNeil, M.D.; Berry, D.P.; Clark, S.A.; Crowley, J.J.; Scholtz, M.M. Evaluation of partial body weight for predicting body weight and average daily gain in growing beef cattle. Transl. Anim. Sci. 2021, 5, txab126. [Google Scholar] [CrossRef]
- Taiwo, G.A.; Idowu, M.; Denvir, J.; Cervantes, A.P.; Ogunade, I.M. Identification of Key Pathways Associated with Residual Feed Intake of Beef Cattle Based on Whole Blood Transcriptome Data Analyzed Using Gene Set Enrichment Analysis. Front. Vet. Sci. 2022, 9, 848027. [Google Scholar] [CrossRef]
- Durunna, O.N.; Mujibi, F.D.N.; Goonewardene, L.; Okine, E.K.; Basarab, J.A.; Wang, Z.; Moore, S.S. Feed efficiency differences and reranking in beef steers fed grower and finisher diets1. J. Anim. Sci. 2011, 89, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Ogunade, I.; Jiang, Y.; Adeyemi, J.; Oliveira, A.; Vyas, D.; Adesogan, A. Biomarker of Aflatoxin Ingestion: 1H NMR-Based Plasma Metabolomics of Dairy Cows Fed Aflatoxin B₁ with or without Sequestering Agents. Toxins 2018, 10, 545. [Google Scholar] [CrossRef] [PubMed]
- Psychogios, N.; Hau, D.D.; Peng, J.; Guo, A.C.; Mandal, R.; Bouatra, S.; Sinelnikov, I.; Krishnamurthy, R.; Eisner, R.; Gautam, B.; et al. The human serum metabolome. PLoS ONE 2011, 6, e16957. [Google Scholar] [CrossRef] [PubMed]
- Ravanbakhsh, S.; Liu, P.; Bjorndahl, T.C.; Mandal, R.; Grant, J.R.; Wilson, M.; Eisner, R.; Sinelnikov, I.; Hu, X.; Luchinat, C.; et al. Accurate, fully-automated NMR spectral profiling for metabolomics. PLoS ONE 2015, 10, e0124219. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Wishart, D.S.; Xia, J. Using MetaboAnalyst 4.0 for Comprehensive and Integrative Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2019, 68, e86. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Mateo, R.D.; Yin, Y.-L.; Wu, G. Functional Amino Acids and Fatty Acids for Enhancing Production Performance of Sows and Piglets. Asian-Australas. J. Anim. Sci. 2006, 20, 295–306. [Google Scholar] [CrossRef]
- Wynne, K.; Bloom, S.R. The role of oxyntomodulin and peptide tyrosine–tyrosine (PYY) in appetite control. Nat. Clin. Pract. Endocrinol. Metab. 2006, 2, 612–620. [Google Scholar] [CrossRef]
- Church, D.D.; Hirsch, K.R.; Park, S.; Kim, I.-Y.; Gwin, J.A.; Pasiakos, S.M.; Wolfe, R.R.; Ferrando, A.A. Essential Amino Acids and Protein Synthesis: Insights into Maximizing the Muscle and Whole-Body Response to Feeding. Nutrients 2020, 12, 3717. [Google Scholar] [CrossRef] [PubMed]
- Yoo, E.-S.; Yu, J.; Sohn, J.-W. Neuroendocrine control of appetite and metabolism. Exp. Mol. Med. 2021, 53, 505–516. [Google Scholar] [CrossRef]
- Mohagheghpour, N.; Waleh, N.; Garger, S.J.; Dousman, L.; Grill, L.K.; Tusé, D. Synthetic Melanin Suppresses Production of Proinflammatory Cytokines. Cell. Immunol. 2000, 199, 25–36. [Google Scholar] [CrossRef]
- Kin, N.W.; Sanders, V.M. It takes nerve to tell T and B cells what to do. J. Leukoc. Biol. 2006, 79, 1093–1104. [Google Scholar] [CrossRef] [PubMed]
- Konashi, S.; Takahashi, K.; Akiba, Y. Effects of dietary essential amino acid deficiencies on immunological variables in broiler chickens. Br. J. Nutr. 2000, 83, 449–456. [Google Scholar] [PubMed]
- Cooper, J.E.; Cooper, M.E. (Eds.) Importance and Application of Animal Law. In Introduction to Veterinary and Comparative Forensic Medicine; Wiley: Hoboken, NJ, USA, 2007; pp. 42–61. [Google Scholar]
- Wu, G.; Bazer, F.W.; Wallace, J.M.; Spencer, T.E. BOARD-INVITED REVIEW: Intrauterine growth retardation: Implications for the animal sciences. J. Anim. Sci. 2006, 84, 2316–2337. [Google Scholar] [CrossRef] [PubMed]
- Kerley, M.S. Nutrition and Feed Efficiency of Beef Cattle. In Feed Efficiency in the Beef Industry; Wiley: Hoboken, NJ, USA, 2012; pp. 75–92. [Google Scholar]
- Cantalapiedra-Hijar, G.; Ortigues-Marty, I.; Sepchat, B.; Titgemeyer, E.; Bahloul, L. Methionine-balanced diets improve cattle performance in fattening young bulls fed high-forage diets through changes in nitrogen metabolism. Br. J. Nutr. 2020, 124, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Sanmartín-Suárez, C.; Soto-Otero, R.; Sánchez-Sellero, I.; Méndez-Álvarez, E. Antioxidant properties of dimethyl sulfoxide and its viability as a solvent in the evaluation of neuroprotective antioxidants. J. Pharmacol. Toxicol. Methods 2011, 63, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Butawan, M.; Benjamin, R.L.; Bloomer, R.J. Methylsulfonylmethane: Applications and Safety of a Novel Dietary Supplement. Nutrients 2017, 9, 290. [Google Scholar] [CrossRef] [PubMed]
- Ponnampalam, E.N.; Kiani, A.; Santhiravel, S.; Holman, B.W.B.; Lauridsen, C.; Dunshea, F.R. The Importance of Dietary Antioxidants on Oxidative Stress, Meat and Milk Production, and Their Preservative Aspects in Farm Animals: Antioxidant Action, Animal Health, and Product Quality-Invited Review. Animals 2022, 12, 3279. [Google Scholar] [CrossRef] [PubMed]
- Lykkesfeldt, J.; Svendsen, O. Oxidants and antioxidants in disease: Oxidative stress in farm animals. Vet. J. 2007, 173, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Celi, P. Oxidative Stress in Ruminants. In Oxidative Stress in Applied Basic Research and Clinical Practice; Humana Press: Totowa, NJ, USA, 2011; pp. 191–231. [Google Scholar]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef]
- Cao, S.S.; Kaufman, R.J. Endoplasmic reticulum stress and oxidative stress in cell fate decision and human disease. Antioxid. Redox Signal. 2014, 21, 396–413. [Google Scholar] [CrossRef]
- Coleman, D.N.; Lopreiato, V.; Alharthi, A.; Loor, J.J. Amino acids and the regulation of oxidative stress and immune function in dairy cattle. J. Anim. Sci. 2020, 98 (Suppl. S1), S175–S193. [Google Scholar] [CrossRef]
- Guan, X.-L.; Wu, P.-F.; Wang, S.; Zhang, J.-J.; Shen, Z.-C.; Luo, H.; Chen, H.; Long, L.-H.; Chen, J.-G.; Wang, F. Dimethyl sulfide protects against oxidative stress and extends lifespan via a methionine sulfoxide reductase A-dependent catalytic mechanism. Aging Cell 2017, 16, 226–236. [Google Scholar] [CrossRef]
- Fujisaka, S.; Avila-Pacheco, J.; Soto, M.; Kostic, A.; Dreyfuss, J.M.; Pan, H.; Ussar, S.; Altindis, E.; Li, N.; Bry, L.; et al. Diet, Genetics, and the Gut Microbiome Drive Dynamic Changes in Plasma Metabolites. Cell Rep. 2018, 22, 3072–3086. [Google Scholar] [CrossRef] [PubMed]
- Clemmons, B.A.; Voy, B.H.; Myer, P.R. Altering the Gut Microbiome of Cattle: Considerations of Host-Microbiome Interactions for Persistent Microbiome Manipulation. Microb. Ecol. 2018, 77, 523–536. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
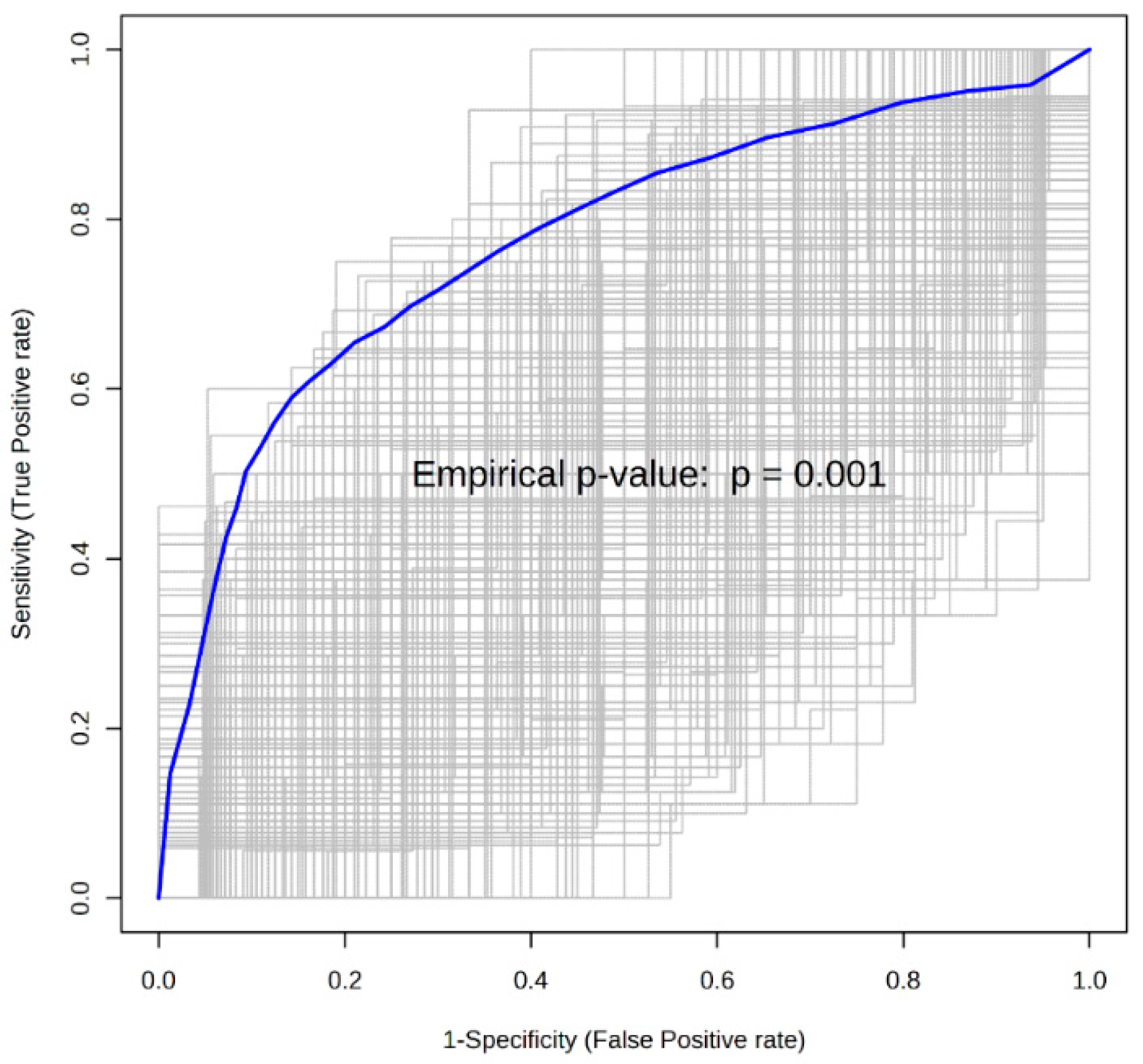{kind=link}
| Item | Low-RFI | High-RFI | SEM | p-Value |
|---|---|---|---|---|
| RFI (kg/d) | −1.54 | 1.49 | 0.03 | 0.01 |
| Initial weight (kg) | 280 | 272 | 8.90 | 0.63 |
| Final weight (kg) | 336 | 327 | 5.93 | 0.41 |
| ADG (kg/d) | 1.13 | 1.11 | 0.04 | 0.86 |
| DMI (kg/d) | 7.08 | 8.23 | 0.38 | 0.01 |
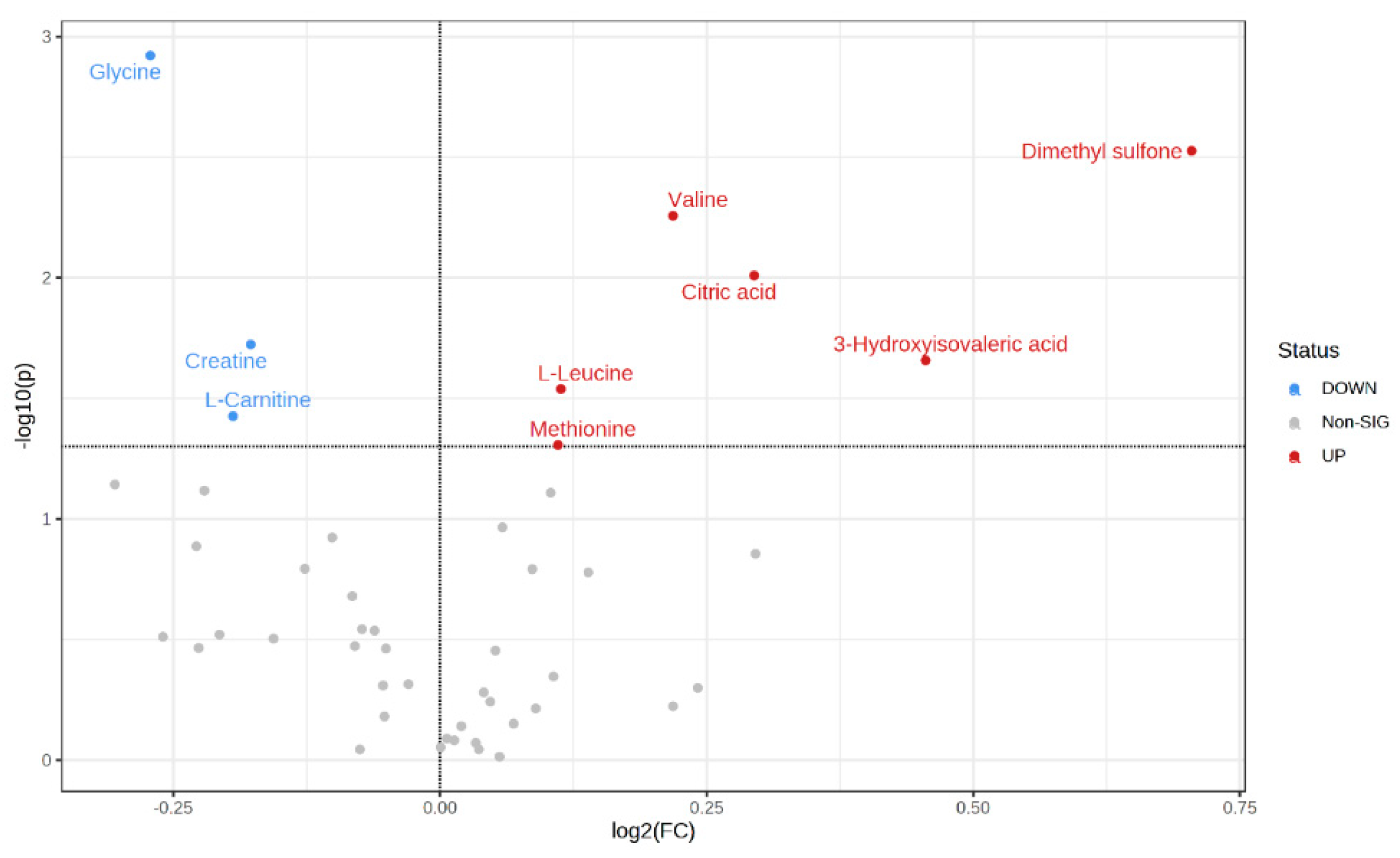
| Metabolites (mM) | FC (Low-RFI/High-RFI) | p-Values |
|---|---|---|
| Dimethyl sulfone | 1.63 | 0.01 |
| 3-Hydroxyisovaleric acid | 1.37 | 0.02 |
| Citric acid | 1.23 | 0.01 |
| Valine | 1.16 | 0.01 |
| L-Leucine | 1.08 | 0.03 |
| Methionine | 1.08 | 0.05 |
| Creatine | 0.88 | 0.02 |
| L-Carnitine | 0.87 | 0.04 |
| Glycine | 0.83 | 0.01 |
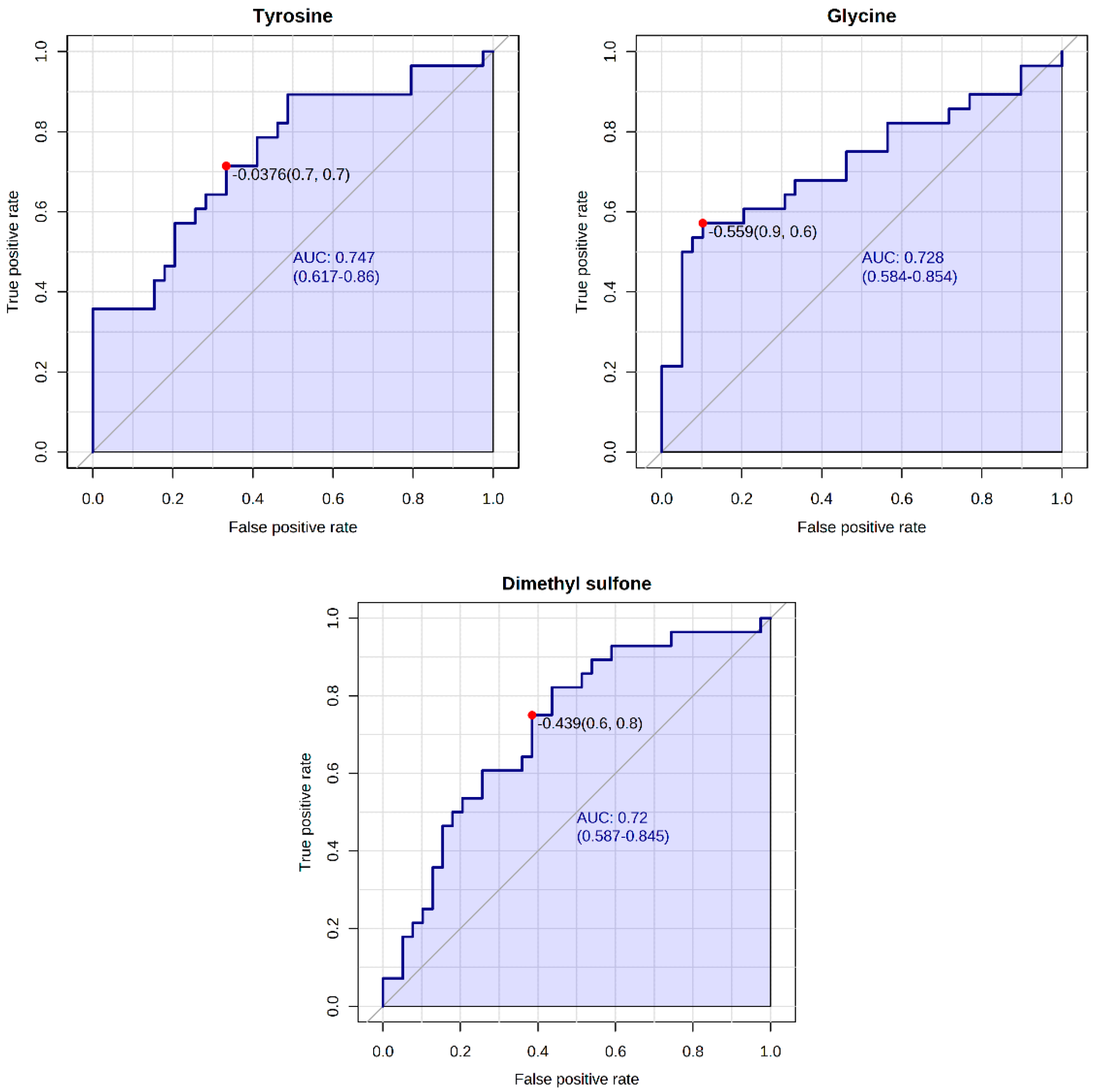
| Intercept | Estimate | Std. Error | z Value | Pr (>|z|) | Odds |
|---|---|---|---|---|---|
| (Intercept) | −0.437 | 0.306 | −1.426 | 0.154 | - |
| Dimethyl sulfone | 1.035 | 0.346 | 2.996 | 0.003 | 2.82 |
| Tyrosine | 0.248 | 0.317 | 0.782 | 0.434 | 1.28 |
| Glycine | −1.152 | 0.354 | −3.256 | 0.001 | 0.32 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taiwo, G.; Idowu, M.; Sidney, T.; Treon, E.; Ologunagba, D.; Leal, Y.; Johnson, S.; Taiwo, R.O.; Adewoye, A.; Ezeigbo, E.; et al. 1H-NMR-Based Plasma Metabolomic Profiling of Crossbred Beef Cattle with Divergent RFI Phenotype. Ruminants 2024, 4, 182-191. https://doi.org/10.3390/ruminants4020012
Taiwo G, Idowu M, Sidney T, Treon E, Ologunagba D, Leal Y, Johnson S, Taiwo RO, Adewoye A, Ezeigbo E, et al. 1H-NMR-Based Plasma Metabolomic Profiling of Crossbred Beef Cattle with Divergent RFI Phenotype. Ruminants. 2024; 4(2):182-191. https://doi.org/10.3390/ruminants4020012
Chicago/Turabian StyleTaiwo, Godstime, Modoluwamu Idowu, Taylor Sidney, Emily Treon, Deborah Ologunagba, Yarahy Leal, Samanthia Johnson, Rhoda Olowe Taiwo, Anjola Adewoye, Ephraim Ezeigbo, and et al. 2024. "1H-NMR-Based Plasma Metabolomic Profiling of Crossbred Beef Cattle with Divergent RFI Phenotype" Ruminants 4, no. 2: 182-191. https://doi.org/10.3390/ruminants4020012