
Mitochondrial Effects, DNA Damage, and Antioxidant Enzyme Activity in Cryopreserved Human Sperm Samples: A Pilot Study


 ,
,  ,
, 
Abstract
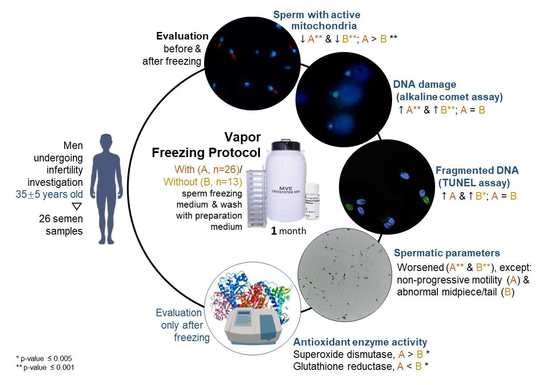
1. Introduction
2. Materials and Methods
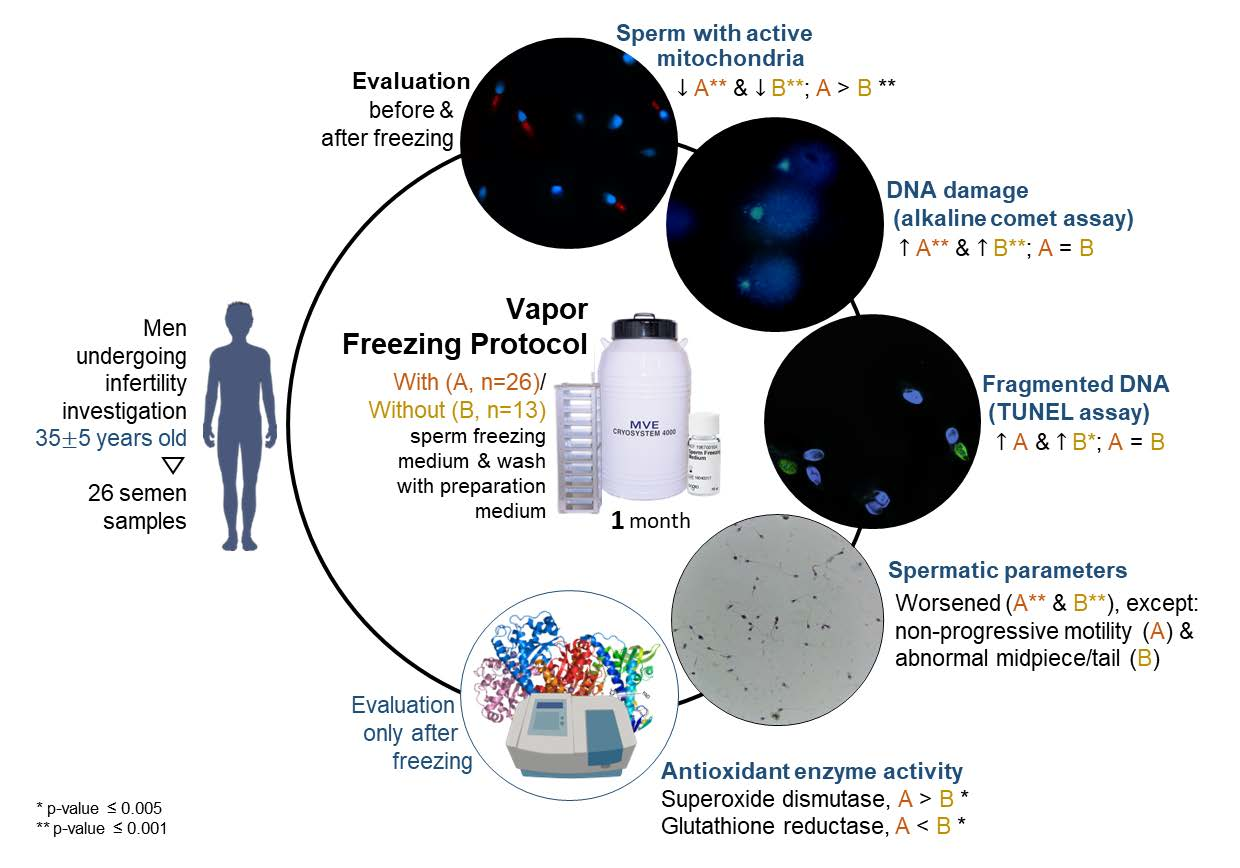
2.1. Study Design and Sample Collection
2.2. Vapor Freezing and Thawing
2.3. Semen Analysis
2.4. Percentage of Spermatozoa with Active Mitochondria
2.5. DNA Integrity
2.5.1. ACA
2.5.2. TUNEL
2.6. Antioxidant Enzymes Activity
2.6.1. Superoxide Dismutase (SOD)
2.6.2. Glutathione Reductase (GR)
2.6.3. Protein Quantification Assay
2.7. Statistical Analysis
3. Results
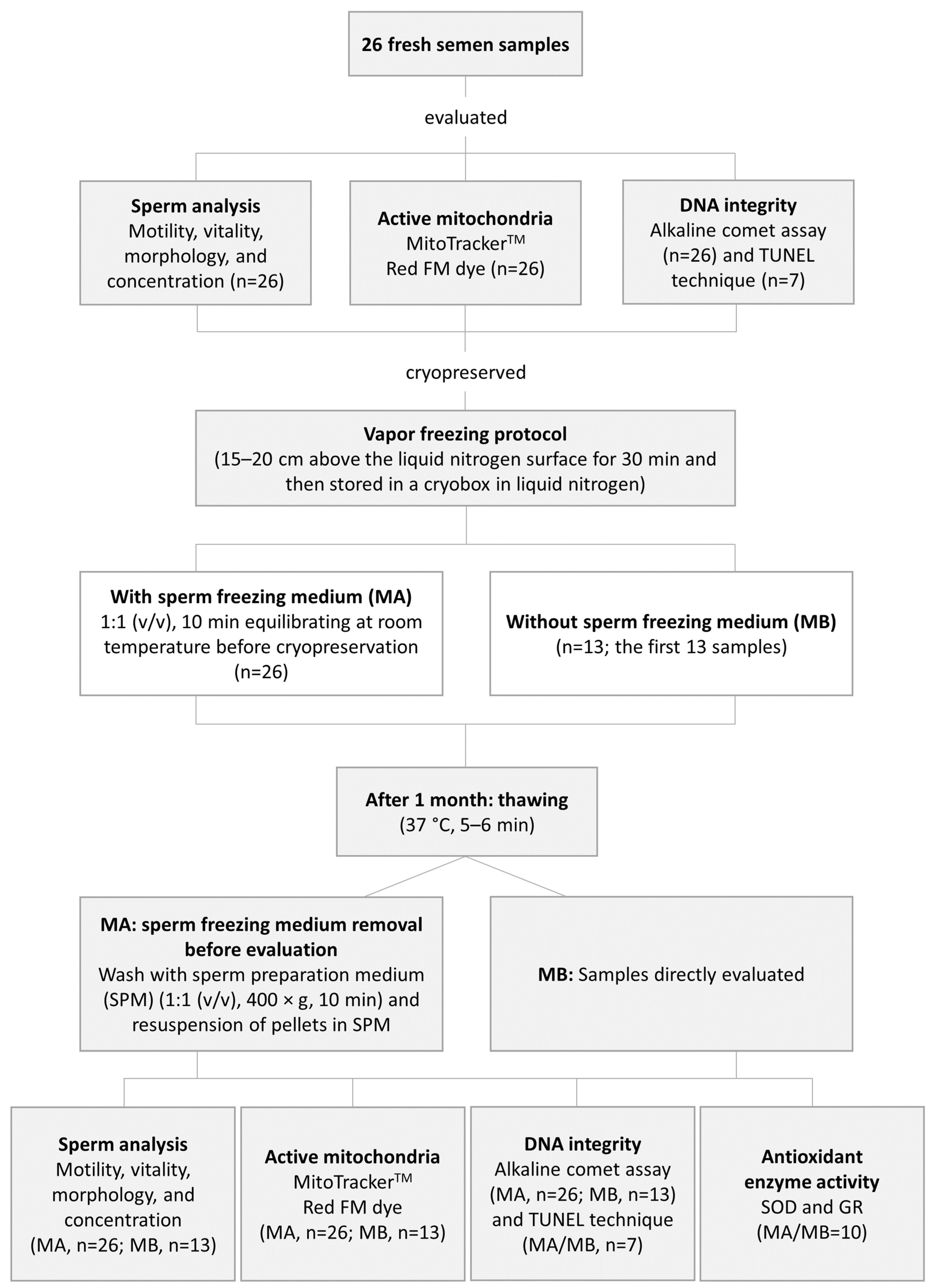
3.1. Spermatic Parameters
3.2. Percentage of Spermatozoa with Active Mitochondria
3.3. DNA Integrity
3.4. Antioxidant Enzymes Activity
3.5. The Influence of Spermatic Quality on Cryopreservation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Best, B.P. Cryoprotectant Toxicity: Facts, Issues, and Questions. Rejuvenation Res. 2015, 18, 422–436. [Google Scholar] [CrossRef] [PubMed]
- Majzoub, A.; Argawal, A. Preface. In The Complete Guide to Male Fertility Preservation; Springer International: Cham, Switzerland, 2018. [Google Scholar]
- Creemers, E.; Nijs, M.; Vanheusden, E.; Ombelet, W. Cryopreservation of human sperm: Efficacy and use of a new nitrogen-free controlled rate freezer versus liquid nitrogen vapour freezing. Andrologia 2011, 43, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Talwar, P.; Ghosh, P. Sperm Freezing Injuries. In The Complete Guide to Male Fertility Preservation; Springer International: Cham, Switzerland, 2018; pp. 205–226. [Google Scholar]
- Gupta, S.; Sharma, R.; Agarwal, A. The Process of Sperm Cryopreservation, Thawing and Washing Techniques. In The Complete Guide to Male Fertility Preservation; Majzoub, A., Argawal, A., Eds.; Springer International Springer International: Cham, Switzerland, 2018; pp. 183–204. [Google Scholar]
- Nallella, K.P.; Sharma, R.K.; Allamaneni, S.S.; Aziz, N.; Agarwal, A. Cryopreservation of human spermatozoa: Comparison of two cryopreservation methods and three cryoprotectants. Fertil. Steril. 2004, 82, 913–918. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, C.; Forell, F.; Oliveira, A.; Rodrigues, J. Current status of sperm cryopreservation: Why isn’t it better? Theriogenology 2002, 57, 327–344. [Google Scholar] [CrossRef]
- Lopes, F.; Pinto-Pinho, P.; Gaivão, I.; Martins-Bessa, A.; Gomes, Z.; Moutinho, O.; Oliveira, M.M.; Peixoto, F.; Pinto-Leite, R. Sperm DNA damage and seminal antioxidant activity in subfertile men. Andrologia 2021, 53, e14027. [Google Scholar] [CrossRef]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen; WHO: Geneva, Switzerland, 2010. [Google Scholar]
- Amaral, A.; Ramalho-Santos, J. Assessment of mitochondrial potential: Implications for the correct monitoring of human sperm function. Int. J. Androl. 2010, 33, e180–e186. [Google Scholar] [CrossRef]
- Invitrogen. MitoTracker® Mitochondrion—Selective Probes; Thermo Fisher Scientific: Waltham, MA, USA, 2008. [Google Scholar]
- Ribas-Maynou, J.; García-Peiró, A.; Fernández-Encinas, A.; Abad, C.; Amengual, M.J.; Prada, E.; Navarro, J.; Benet, J. Comprehensive analysis of sperm DNA fragmentation by five different assays: TUNEL assay, SCSA, SCD test and alkaline and neutral Comet assay. Andrology 2013, 1, 715–722. [Google Scholar] [CrossRef]
- Sipinen, V.; Laubenthal, J.; Baumgartner, A.; Cemeli, E.; Linschooten, J.O.; Godschalk, R.W.L.; Van Schooten, F.J.; Anderson, D.; Brunborg, G. In vitro evaluation of baseline and induced DNA damage in human sperm exposed to benzo[a]pyrene or its metabolite benzo[a]pyrene-7,8-diol-9,10-epoxide, using the comet assay. Mutagenesis 2010, 25, 417–425. [Google Scholar] [CrossRef]
- Collins, A.R. The Comet Assay for DNA Damage and Repair: Principles, Applications, and Limitations. Mol. Biotechnol. 2004, 26, 249–261. [Google Scholar] [CrossRef]
- Muratori, M.; Piomboni, P.; Baldi, E.; Filimberti, E.; Pecchioli, P.; Moretti, E.; Gambera, L.; Baccetti, B.; Biagiotti, R.; Forti, G.; et al. Functional and Ultrastructural Features of DNA-Fragmented Human Sperm. J. Androl. 2000, 21, 903–912. [Google Scholar]
- Payá, M.; Halliwell, B.; Hoult, J. Interactions of a series of coumarins with reactive oxygen species: Scavenging of superoxide, hypochlorous acid and hydroxyl radicals. Biochem. Pharmacol. 1992, 44, 205–214. [Google Scholar] [CrossRef]
- Carlberg, I.; Mannervik, B. [59] Glutathione reductase. Methods Enzymol. 1985, 113, 484–490. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- CooperSurgical. Sperm Freezing Medium. Available online: https://fertility.coopersurgical.com/art_media/sperm-freezing-medium/ (accessed on 30 May 2020).
- CooperSurgical. Sperm Preparation Medium. Available online: https://fertility.coopersurgical.com/art_media/sperm-preparation-medium/ (accessed on 30 May 2020).
- Lusignan, M.F.; Li, X.; Herrero, B.; Delbes, G.; Chan, P.T.K. Effects of different cryopreservation methods on DNA integrity and sperm chromatin quality in men. Andrology 2018, 6, 829–835. [Google Scholar] [CrossRef] [PubMed]
- Raad, G.; Lteif, L.; Lahoud, R.; Azoury, J.; Azoury, J.; Tanios, J.; Hazzouri, M.; Azoury, J. Cryopreservation media differentially affect sperm motility, morphology and DNA integrity. Andrology 2018, 6, 836–845. [Google Scholar] [CrossRef] [PubMed]
- Agha-Rahimi, A.; Khalili, M.A.; Nabi, A.; Ashourzadeh, S. Vitrification is not superior to rapid freezing of normozoospermic spermatozoa: Effects on sperm parameters, DNA fragmentation and hyaluronan binding. Reprod. Biomed. Online 2014, 28, 352–358. [Google Scholar] [CrossRef][Green Version]
- Ozkavukcu, S.; Erdemli, E.; Isik, A.; Oztuna, D.; Karahuseyinoglu, S. Effects of cryopreservation on sperm parameters and ultrastructural morphology of human spermatozoa. J. Assist. Reprod. Genet. 2008, 25, 403–411. [Google Scholar] [CrossRef]
- O’Connell, M.; McClure, N.; Lewis, S.E.M. The effects of cryopreservation on sperm morphology, motility and mitochondrial function. Hum. Reprod. 2002, 17, 704–709. [Google Scholar] [CrossRef]
- Pabón, D.; Meseguer, M.; Sevillano, G.; Cobo, A.; Romero, J.L.; Remohi, J.; de Los Santos, M.J. A new system of sperm cryopreservation: Evaluation of survival, motility, DNA oxidation, and mitochondrial activity. Andrology 2019, 7, 293–301. [Google Scholar] [CrossRef]
- Thomson, L.K.; Fleming, S.D.; Aitken, R.J.; De Iuliis, G.N.; Zieschang, J.A.; Clark, A.M. Cryopreservation-induced human sperm DNA damage is predominantly mediated by oxidative stress rather than apoptosis. Hum. Reprod. 2009, 24, 2061–2070. [Google Scholar] [CrossRef]
- Kalthur, G.; Adiga, S.K.; Upadhya, D.; Rao, S.; Kumar, P. Effect of cryopreservation on sperm DNA integrity in patients with teratospermia. Fertil. Steril. 2008, 89, 1723–1727. [Google Scholar] [CrossRef] [PubMed]
- Isachenko, V.; Isachenko, E.; Katkov, I.I.; Montag, M.; Dessole, S.; Nawroth, F.; Van Der Ven, H. Cryoprotectant-Free Cryopreservation of Human Spermatozoa by Vitrification and Freezing in Vapor: Effect on Motility, DNA Integrity, and Fertilization Ability. Biol. Reprod. 2004, 71, 1167–1173. [Google Scholar] [CrossRef] [PubMed]
- Majtnerová, P.; Roušar, T. An overview of apoptosis assays detecting DNA fragmentation. Mol. Biol. Rep. 2018, 45, 1469–1478. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.R.; Oscoz, A.A.; Brunborg, G.; Gaivao, I.; Giovannelli, L.; Kruszewski, M.; Smith, C.C.; Štětina, R. The comet assay: Topical issues. Mutagenesis 2008, 23, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Gregg, X.T.; Prchal, J.T. Red Blood Cell Enzymopathies. In Hematology; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Chatterjee, S.; Gagnon, C. Production of reactive oxygen species by spermatozoa undergoing cooling, freezing, and thawing. Mol. Reprod. Dev. 2001, 59, 451–458. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, J.; Chen, L.; Li, J.; Zhang, H.; Guo, X. Glycine Suppresses AGE/RAGE Signaling Pathway and Subsequent Oxidative Stress by Restoring Glo1 Function in the Aorta of Diabetic Rats and in HUVECs. Oxidative Med. Cell. Longev. 2019, 2019, 4628962. [Google Scholar] [CrossRef]
- Ghorbani, M.; Vatannejad, A.; Khodadadi, I.; Amiri, I.; Tavilani, H. Protective effects of glutathione supplementation against oxidative stress during cryopreservation of human spermatozoa. Cryoletters 2016, 37, 34–40. [Google Scholar]
- Boveris, A.; Chance, B. The mitochondrial generation of hydrogen peroxide. General properties and effect of hyperbaric oxygen. Biochem. J. 1973, 134, 707–716. [Google Scholar] [CrossRef]
- Vutyavanich, T.; Piromlertamorn, W.; Nunta, S. Rapid freezing versus slow programmable freezing of human spermatozoa. Fertil. Steril. 2010, 93, 1921–1928. [Google Scholar] [CrossRef]
- Choucair, F.B.; Rachkidi, E.G.; Raad, G.C.; Saliba, E.M.; Zeidan, N.S.; Jounblat, R.A.; Abou Jaoude, I.F.; Hazzouri, M.M. High level of DNA fragmentation in sperm of Lebanese infertile men using Sperm Chromatin Dispersion test. Middle East Fertil. Soc. J. 2016, 21, 269–276. [Google Scholar] [CrossRef]
- Thomson, L.K.; Fleming, S.D.; Schulke, L.; Barone, K.; Zieschang, J.-A.; Clark, A.M. The DNA integrity of cryopreserved spermatozoa separated for use in assisted reproductive technology is unaffected by the type of cryoprotectant used but is related to the DNA integrity of the fresh separated preparation. Fertil. Steril. 2009, 92, 991–1001. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Spermatic Parameters | Method A (n = 26) | Method B (n = 13) | ||||||
|---|---|---|---|---|---|---|---|---|
| Fresh | Cryopreserved | Change | p-Value | Fresh | Cryopreserved | Change | p-Value | |
| PR motility (%) | 57.4 ± 16.3 (13.0–80.0) | 12.6 ± 8.0 (0.7–30) | −78.0% (49.2–98.8) | 0.000 | 59.8 ± 12.8 (36.0–80.0) | 0.6 ± 1.1 (0.0–4.0) | −99.0% (94.9–100.0) | 0.001 |
| NP motility (%) | 12.7 ± 4.9 (6.0–26.0) | 11.7 ± 6.6 (1.7–24.4) | −7.3% (4.3–90.0) | >0.050 | 13.2 ± 4.9 (6.0–20.0) | 0.5 ± 0.6 (0.0–2.0) | −95.9% (77.8–100.0) | 0.001 |
| Total sperm motility (%) | 70.0 ± 14.1 (26.0–86.0) | 24.3 ± 12.3 (3.0–46.0) | −65.2% (37.0–95.9) | 0.000 | 72.9 ± 9.8 (53.0–86.0) | 1.1 ± 1.4 (0.0–5.0) | −98.5% (94.2–100.0) | 0.001 |
| Vitality (%) | 77.3 ± 13.0 (28.0–91.0) | 26.7 ± 12.7 (3.5–47.0) | −65.4% (35.6–95.3) | 0.000 | 80.5 ± 4.1 (73.0–89.0) | 4.7 ± 4.9 (0.6–19.6) | −94.2% (78.0–99.3) | 0.001 |
| Morphologically normal SPZ (%) | 3.6 ± 1.8 (0.0–7.0) | 1.5 ± 1.3 (0.0–4.0) | −59.6% (0.0–100.0) | 0.000 | 4.1 ± 1.7 (1.0–6.0) | 1.1 ± 1.3 (0.0–4.0) | −73.6% (33.3–100.0) | 0.002 |
| SPZ with abnormal head (%) | 96.3 ± 1.8 (93.0–100.0) | 98.5 ± 1.3 (96.0–100.0) | +2.3% (0.0–5.3) | 0.000 | 95.8 ± 1.7 (94.0–99.0) | 98.8 ± 1.6 (95.0–100.0) | +3.0% (1.0–6.4) | 0.006 |
| SPZ with abnormal midpiece (%) | 46.5 ± 7.8 (28.0–57.0) | 51.8 ± 9.9 (34.0–74.0) | +11.4% (0.0–128.6) | 0.043 | 47.3 51.8 ± 7.8 (33.0–57.0) | 51.5 ± 9.4 (35.0–68.0) | +8.9% (0.0–42.1) | >0.050 |
| SPZ with abnormal tail (%) | 4.8 ± 4.0 (1.0–20.0) | 16.2 ± 10.3 (4.0–52.0) | +239.5% (22.2–1900.0%) | 0.000 | 4.8 ± 2.8 (1.0–10.0) | 8.6 ± 7.9 (2.0–32.0) | +80.6% (25.0–400.0) | >0.050 |
| SPZ with excess residual cytoplasm (%) | 2.1 ± 1.6 (0.0–6.0) | 0.3 ± 0.7 (0.0–3.0) | −85.5% (0.0–100.0) | 0.000 | 2.2 ± 1.5 (0.0–4.0) | 0.4 ± 0.8 (0.0–2.0) | −82.8% (0.0–100.0) | 0.011 |
| Sperm concentration (×106 SPZ/mL) | 80.9 ± 70.1 (3.1–338.5) | 41.9 ± 43.5 (1.3–203.3) | −48.2% (31.9–87.2) | 0.000 | 84.1 ± 88.6 (4.7–338.5) | 74.2 ± 89.1 (3.7–341.5) | −11.8% (6.0–31.7) | 0.007 |
| Cryopreservation Protocol | Sperm with Active Mitochondria (%) | |||
|---|---|---|---|---|
| Fresh | Cryopreserved | Change | p-Value | |
| Method A (n = 26) | 65.7 ± 14.5 (21.0–92.5) | 21.3 ± 10.7 (2.6–39.1) | −67.6 (33.3–95.8) | 0.000 |
| Method B (n = 13) | 72.0 ± 10.6 (57.5–92.5) | 3.0 ± 2.4 (0.4–8.8) | −95.8 (88.8–99.5) | 0.001 |
| Cryopreservation Protocol | Comet Assay (AU) | |||
|---|---|---|---|---|
| Fresh | Cryopreserved | Change | p-Value | |
| Method A (n = 26) | 87.8 ± 50.9 (18–197) | 171.5 ± 57.8 (27–300) | +95.3% (8.1–782.8%) | 0.000 |
| Method B (n = 13) | 96.6 ± 50.3 (47–197) | 182.5 ± 27.9 (99–204) | +88.9% (3.8–304.2%) | 0.001 |
| TUNEL Assay (%) | ||||
| Fresh | Cryopreserved | Change | p-Value | |
| Method A (n = 7) | 5.9 ± 4.5 (1.0–13.4) | 9.5 ± 6.8 (1.0–19.3) | +71.7% (0.0–1440.0%) | >0.050 |
| Method B (n = 7) | 5.9 ± 4.5 (1.0–13.4) | 12.2 ± 8.5 (3.0–28.0) | +119.9% (100.0–336.0%) | 0.043 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinto-Pinho, P.; Arantes-Rodrigues, R.; Gaivão, I.; Peixoto, F.; Gomes, Z.; Brito, M.; Moutinho, O.; Colaço, B.; Pinto-Leite, R. Mitochondrial Effects, DNA Damage, and Antioxidant Enzyme Activity in Cryopreserved Human Sperm Samples: A Pilot Study. Physiologia 2022, 2, 80-93. https://doi.org/10.3390/physiologia2030008
Pinto-Pinho P, Arantes-Rodrigues R, Gaivão I, Peixoto F, Gomes Z, Brito M, Moutinho O, Colaço B, Pinto-Leite R. Mitochondrial Effects, DNA Damage, and Antioxidant Enzyme Activity in Cryopreserved Human Sperm Samples: A Pilot Study. Physiologia. 2022; 2(3):80-93. https://doi.org/10.3390/physiologia2030008
Chicago/Turabian StylePinto-Pinho, Patrícia, Regina Arantes-Rodrigues, Isabel Gaivão, Francisco Peixoto, Zélia Gomes, Miguel Brito, Osvaldo Moutinho, Bruno Colaço, and Rosário Pinto-Leite. 2022. "Mitochondrial Effects, DNA Damage, and Antioxidant Enzyme Activity in Cryopreserved Human Sperm Samples: A Pilot Study" Physiologia 2, no. 3: 80-93. https://doi.org/10.3390/physiologia2030008
APA StylePinto-Pinho, P., Arantes-Rodrigues, R., Gaivão, I., Peixoto, F., Gomes, Z., Brito, M., Moutinho, O., Colaço, B., & Pinto-Leite, R. (2022). Mitochondrial Effects, DNA Damage, and Antioxidant Enzyme Activity in Cryopreserved Human Sperm Samples: A Pilot Study. Physiologia, 2(3), 80-93. https://doi.org/10.3390/physiologia2030008