
Maximal Torque, Neuromuscular, and Potentiated Twitch Responses to Sustained Forearm Flexion Tasks Using Different Anchor Schemes

Abstract
1. Introduction
2. Results
2.1. Reliability
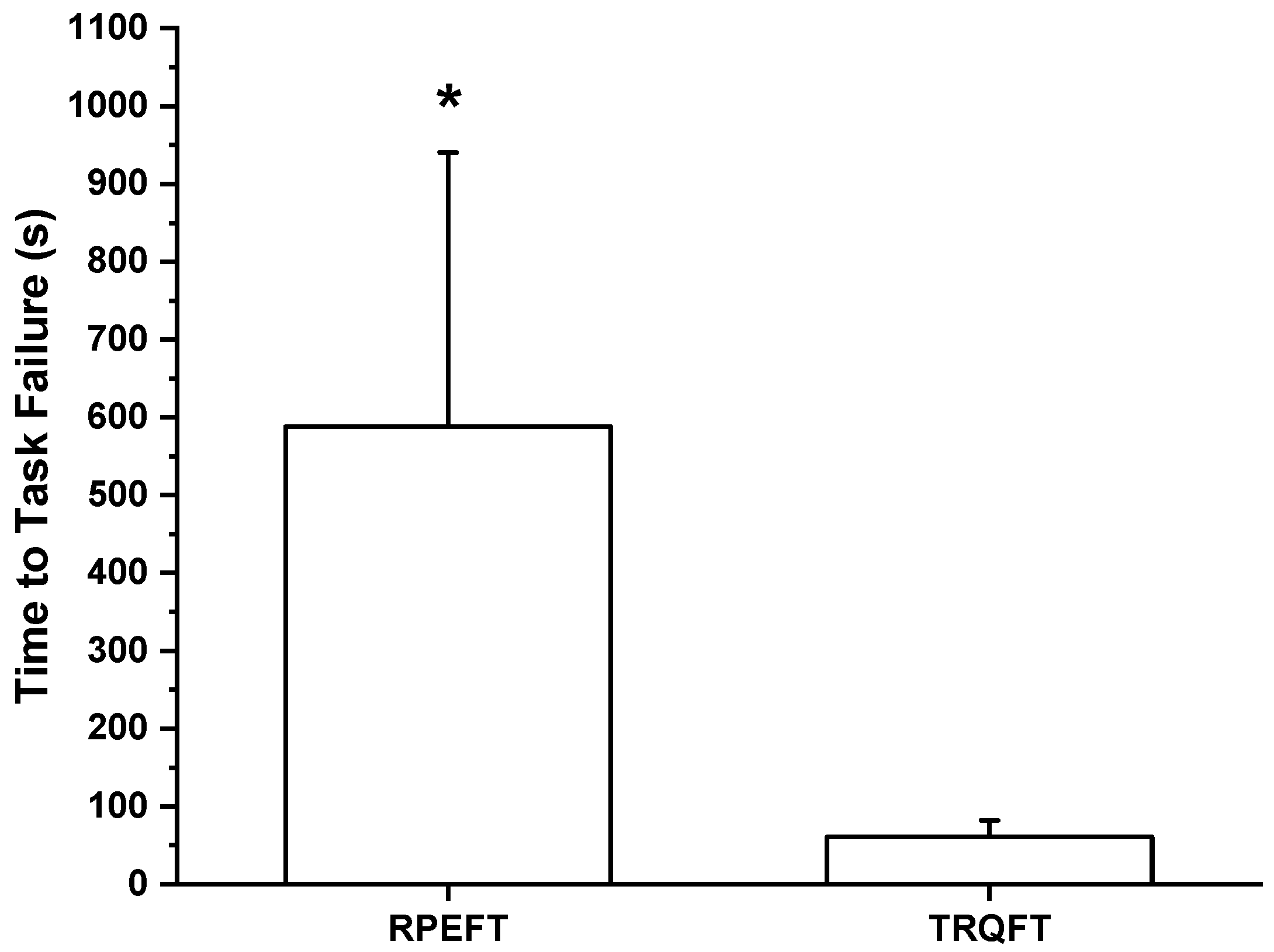
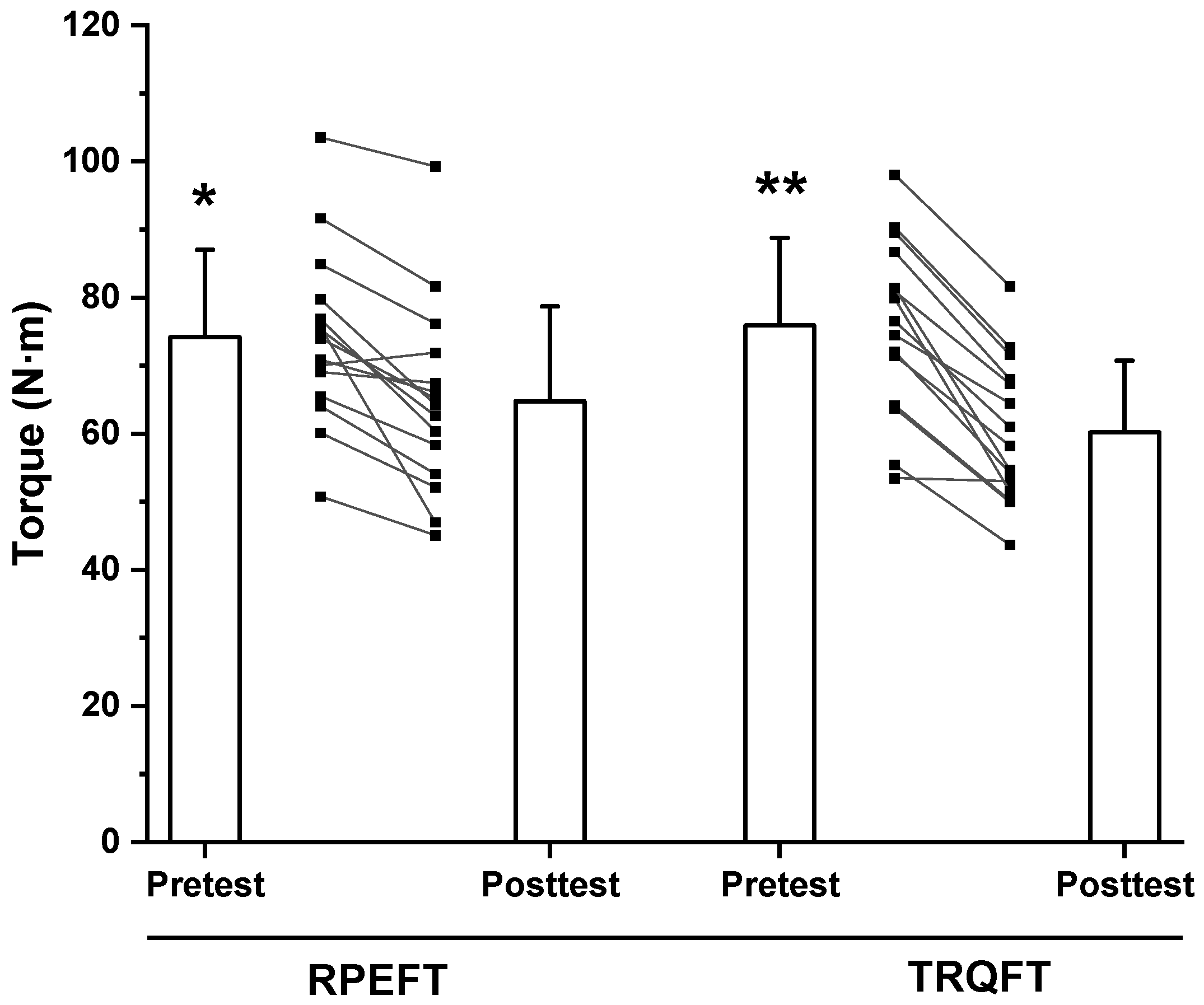
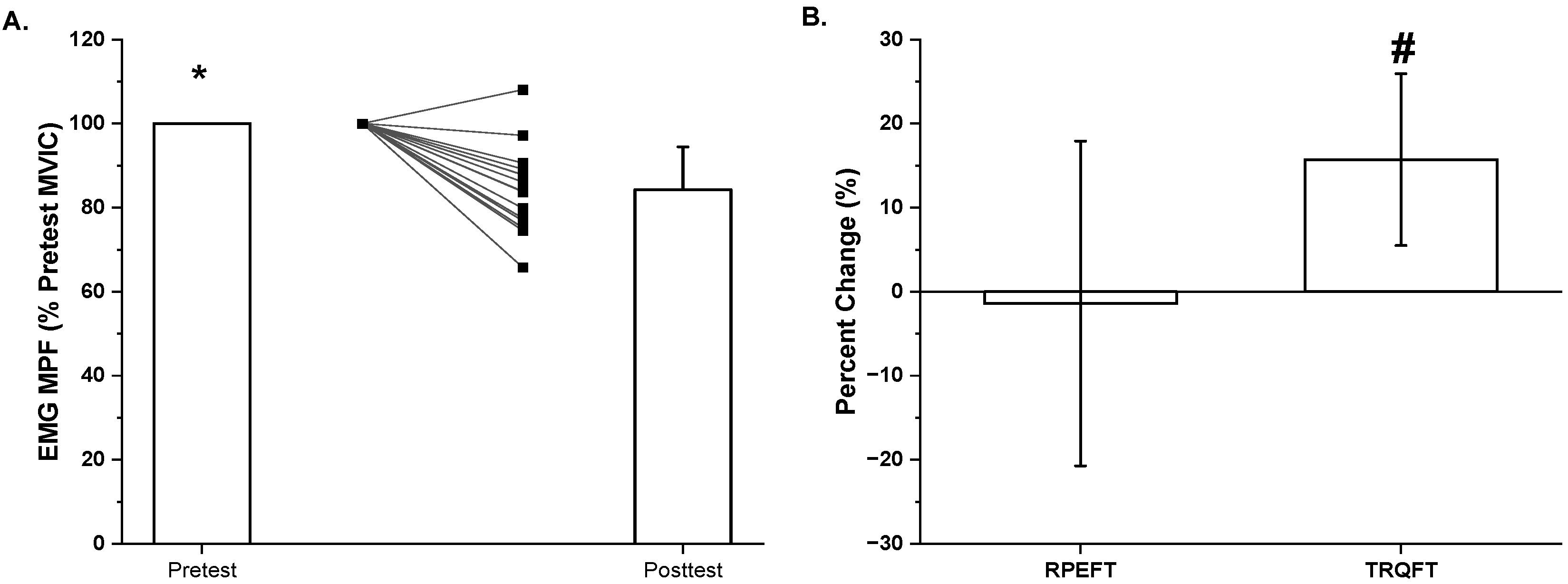
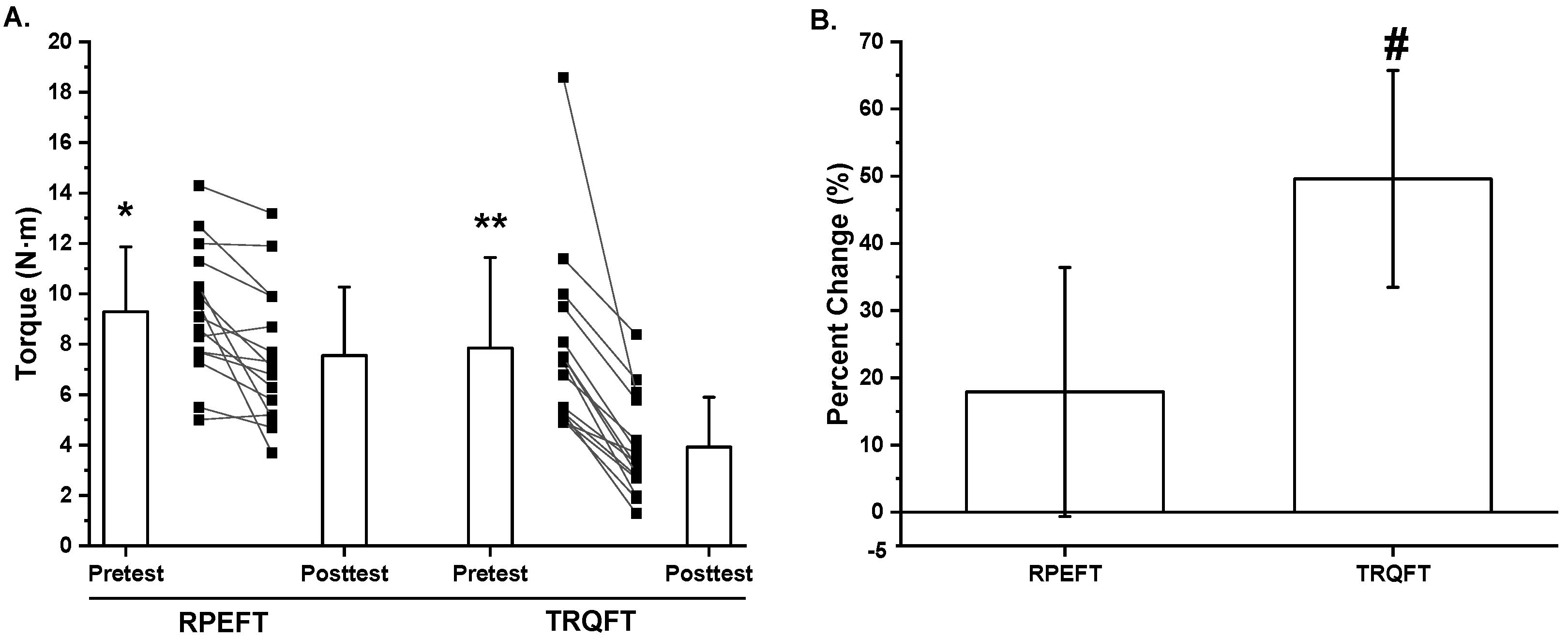
2.2. Time-to-Task Failure, MVIC Torque, Neuromuscular Responses, and Potentiated Twitch Torque
3. Discussion
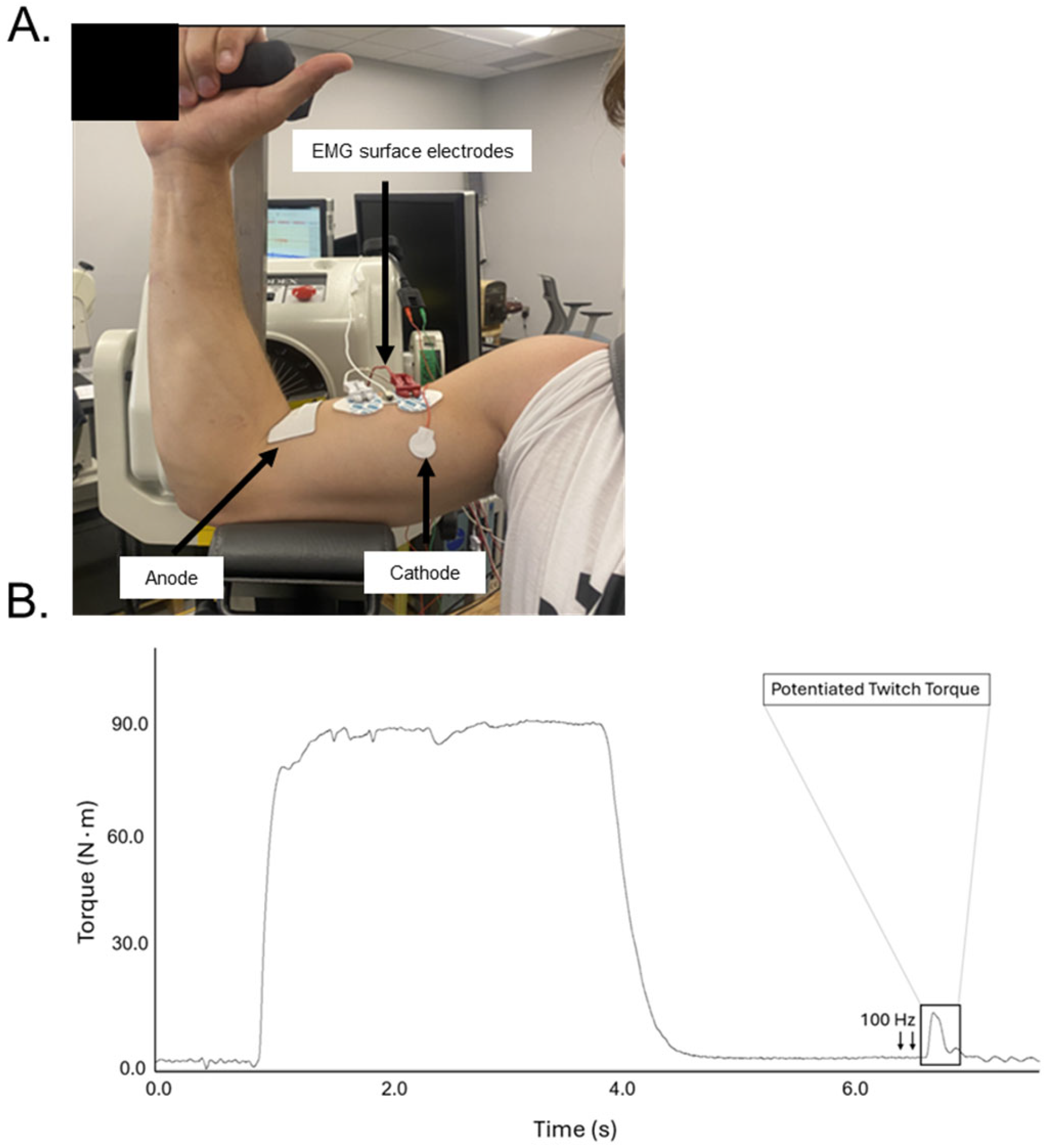
4. Materials and Methods
4.1. Participants
4.2. Orientation Visit
4.3. OMNI-RES Scale Standardized Anchoring Instructions
4.4. Testing Visits
4.5. Evoked Twitch
4.6. Electromyographic and Torque Signal Acquisition
4.7. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Enoka, R.M.; Stuart, D.G. Neurobiology of Muscle Fatigue. J. Appl. Physiol. 1992, 72, 1631–1648. [Google Scholar] [CrossRef]
- Kluger, B.M.; Krupp, L.B.; Enoka, R.M. Fatigue and Fatigability in Neurologic Illnesses: Proposal for a Unified Taxonomy. Neurology 2013, 80, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Enoka, R.M.; Duchateau, J. Translating Fatigue to Human Performance. Med. Sci. Sports Exerc. 2016, 48, 2228–2238. [Google Scholar] [CrossRef] [PubMed]
- Gandevia, S.C. Spinal and Supraspinal Factors in Human Muscle Fatigue. Physiol. Rev. 2001, 81, 1725–1789. [Google Scholar] [CrossRef]
- Hureau, T.J.; Broxterman, R.M.; Weavil, J.C.; Lewis, M.T.; Layec, G.; Amann, M. On the Role of Skeletal Muscle Acidosis and Inorganic Phosphates as Determinants of Central and Peripheral Fatigue: A 31 P-MRS Study. J. Physiol. 2022, 600, 3069–3081. [Google Scholar] [CrossRef]
- Taylor, J.L.; Gandevia, S.C. A Comparison of Central Aspects of Fatigue in Submaximal and Maximal Voluntary Contractions. J. Appl. Physiol. 2008, 104, 542–550. [Google Scholar] [CrossRef]
- De Luca, C.J. Myoelectrical Manifestations of Localized Muscular Fatigue in Humans. Crit. Rev. Biomed. Eng. 1984, 11, 251–279. [Google Scholar]
- Semmler, J.G. Motor Unit Synchronization and Neuromuscular Performance. Exerc. Sport Sci. Rev. 2002, 30, 8–14. [Google Scholar] [CrossRef]
- Lindstrom, L.; Magnusson, R.; Petersén, I. Muscular Fatigue and Action Potential Conduction Velocity Changes Studied with Frequency Analysis of EMG Signals. Electromyography 1970, 10, 341–356. [Google Scholar]
- Place, N. Quantification of Central Fatigue: A Central Debate. Eur. J. Appl. Physiol. 2021, 121, 2375–2376. [Google Scholar] [CrossRef]
- Ortega, D.G.; Smith, R.W.; Arnett, J.E.; Neltner, T.J.; Schmidt, R.J.; Johnson, G.O.; Housh, T.J. Are Performance and Perceived Fatigability Dependent on the Anchor Scheme of Fatiguing Isometric Tasks in Men? J. Sports Med. Phys. Fitness 2024, 64, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Ortega, D.G.; Housh, T.J.; Smith, R.W.; Arnett, J.E.; Neltner, T.J.; Anders, J.P.V.; Schmidt, R.J.; Johnson, G.O. Fatiguing Joint Angle Does Not Influence Torque and Neuromuscular Responses Following Sustained, Isometric Forearm Flexion Tasks Anchored to Perceptual Intensity in Men. J. Funct. Morphol. Kinesiol. 2023, 8, 114. [Google Scholar] [CrossRef] [PubMed]
- Place, N.; Bruton, J.D.; Westerblad, H. Mechanisms of Fatigue Induced by Isometric Contractions in Exercising Humans and in Mouse Isolated Single Muscle Fibres. Clin. Exp. Pharmacol. Physiol. 2009, 36, 334–339. [Google Scholar] [CrossRef] [PubMed]
- Alway, S.E.; Hughson, R.L.; Green, H.J.; Patla, A.E.; Frank, J.S. Twitch Potentiation after Fatiguing Exercise in Man. Eur. J. Appl. Physiol. 1987, 56, 461–466. [Google Scholar] [CrossRef]
- Bigland-Ritchie, B.; Johansson, R.; Lippold, O.C.; Woods, J.J. Contractile Speed and EMG Changes during Fatigue of Sustained Maximal Voluntary Contractions. J. Neurophysiol. 1983, 50, 313–324. [Google Scholar] [CrossRef]
- Neyroud, D.; Rüttimann, J.; Mannion, A.F.; Millet, G.Y.; Maffiuletti, N.A.; Kayser, B.; Place, N. Comparison of Neuromuscular Adjustments Associated with Sustained Isometric Contractions of Four Different Muscle Groups. J. Appl. Physiol. 2013, 114, 1426–1434. [Google Scholar] [CrossRef]
- Bülow, P.M.; Nørregaard, J.; Mehlsen, J.; Danneskiold-Samsøe, B. The Twitch Interpolation Technique for Study of Fatigue of Human Quadriceps Muscle. J. Neurosci. Methods 1995, 62, 103–109. [Google Scholar] [CrossRef]
- Koo, T.K.; Li, M.Y. A Guideline of Selecting and Reporting Intraclass Correlation Coefficients for Reliability Research. J. Chiropr. Med. 2016, 15, 155–163. [Google Scholar] [CrossRef]
- Hill, E.C.; Housh, T.J.; Keller, J.L.; Smith, C.M.; Anders, J.V.; Schmidt, R.J.; Johnson, G.O.; Cramer, J.T. Low-Load Blood Flow Restriction Elicits Greater Concentric Strength than Non-Blood Flow Restriction Resistance Training but Similar Isometric Strength and Muscle Size. Eur. J. Appl. Physiol. 2020, 120, 425–441. [Google Scholar] [CrossRef]
- Smith, R.W.; Neltner, T.J.; Anders, J.P.V.; Keller, J.L.; Housh, T.J.; Schmidt, R.J.; Johnson, G.O. Fatigability, Coactivation, and Neuromuscular Responses of the Biceps Brachii and Triceps Brachii Following Sustained, Maximal, Isometric Forearm Flexion to Task Failure. J. Exerc. Physiol. Online 2021, 24, 55–74. [Google Scholar]
- Ollivier, K.; Portero, P.; Maïsetti, O.; Hogrel, J.-Y. Repeatability of Surface EMG Parameters at Various Isometric Contraction Levels and during Fatigue Using Bipolar and Laplacian Electrode Configurations. J. Electromyogr. Kinesiol. Off. J. Int. Soc. Electrophysiol. Kinesiol. 2005, 15, 466–473. [Google Scholar] [CrossRef] [PubMed]
- Arnett, J.E.; Smith, R.W.; Neltner, T.J.; Anders, J.P.V.; Ortega, D.G.; Housh, T.J.; Schmidt, R.J.; Johnson, G.O. The Effects of Joint Angle and Anchoring Scheme on Performance Fatigability and Neuromuscular Responses Following Isometric Forearm Flexion Tasks to Failure. J. Soc. NeuroSports 2023, 1, 7. [Google Scholar]
- Weir, J.P. Quantifying Test-Retest Reliability Using the Intraclass Correlation Coefficient and the SEM. J. Strength Cond. Res. 2005, 19, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Halaki, M.; Ginn, K.; Halaki, M.; Ginn, K. Normalization of EMG Signals: To Normalize or Not to Normalize and What to Normalize To? IntechOpen: London, UK, 2012; ISBN 978-953-51-0805-4. [Google Scholar]
- Ibitoye, M.O.; Hamzaid, N.A.; Zuniga, J.M.; Hasnan, N.; Wahab, A.K.A. Mechanomyographic Parameter Extraction Methods: An Appraisal for Clinical Applications. Sensors 2014, 14, 22940–22970. [Google Scholar] [CrossRef]
- Soderberg, G.L. Selected Topics in Surface Electromyography for Use in the Occupational Setting: Expert Perspectives; Soderberg, G.L., Ed.; U.S. Department of Health and Human Services, Public Health Service, Centers for Disease Control, National Institute for Occupational Safety and Health: Washington, DC, USA, 1992. [Google Scholar]
- Allen, G.M.; Gandevia, S.C.; McKenzie, D.K. Reliability of Measurements of Muscle Strength and Voluntary Activation Using Twitch Interpolation. Muscle Nerve 1995, 18, 593–600. [Google Scholar] [CrossRef]
- Behrens, M.; Husmann, F.; Gube, M.; Felser, S.; Weippert, M.; Bruhn, S.; Zschorlich, V.; Mau-Moeller, A. Intersession Reliability of the Interpolated Twitch Technique Applied during Isometric, Concentric, and Eccentric Actions of the Human Knee Extensor Muscles. Muscle Nerve 2017, 56, 324–327. [Google Scholar] [CrossRef]
- Dutra, Y.M.; Lopes, J.P.F.; Murias, J.M.; Zagatto, A.M. Within- and between-Day Reliability and Repeatability of Neuromuscular Function Assessment in Females and Males. J. Appl. Physiol. 2023, 135, 1372–1383. [Google Scholar] [CrossRef]
- Place, N.; Maffiuletti, N.A.; Martin, A.; Lepers, R. Assessment of the Reliability of Central and Peripheral Fatigue after Sustained Maximal Voluntary Contraction of the Quadriceps Muscle. Muscle Nerve 2007, 35, 486–495. [Google Scholar] [CrossRef]
- Todd, G.; Gorman, R.B.; Gandevia, S.C. Measurement and Reproducibility of Strength and Voluntary Activation of Lower-Limb Muscles. Muscle Nerve 2004, 29, 834–842. [Google Scholar] [CrossRef]
- Rodriguez-Falces, J.; Place, N. Determinants, Analysis and Interpretation of the Muscle Compound Action Potential (M Wave) in Humans: Implications for the Study of Muscle Fatigue. Eur. J. Appl. Physiol. 2018, 118, 501–521. [Google Scholar] [CrossRef]
- Dotan, R.; Woods, S.; Contessa, P. On the Reliability and Validity of Central Fatigue Determination. Eur. J. Appl. Physiol. 2021, 121, 2393–2411. [Google Scholar] [CrossRef] [PubMed]
- Keller, J.L.; Housh, T.J.; Anders, J.P.V.; Neltner, T.J.; Schmidt, R.J.; Johnson, G.O. Anchor Scheme, Intensity, and Time to Task Failure Do Not Influence Performance Fatigability or Changes in Neuromuscular Responses Following Bilateral Leg Extensions. J. Exerc. Physiol. Online 2020, 23, 119–134. [Google Scholar]
- Thomas, K.; Goodall, S.; Howatson, G. Performance Fatigability Is Not Regulated to A Peripheral Critical Threshold. Exerc. Sport Sci. Rev. 2018, 46, 240–246. [Google Scholar] [CrossRef]
- Hureau, T.J.; Romer, L.M.; Amann, M. The “Sensory Tolerance Limit”: A Hypothetical Construct Determining Exercise Performance? Eur. J. Sport Sci. 2018, 18, 13–24. [Google Scholar] [CrossRef]
- Hartman, M.E.; Ekkekakis, P.; Dicks, N.D.; Pettitt, R.W. Dynamics of Pleasure-Displeasure at the Limit of Exercise Tolerance: Conceptualizing the Sense of Exertional Physical Fatigue as an Affective Response. J. Exp. Biol. 2019, 222, jeb186585. [Google Scholar] [CrossRef]
- Tucker, R. The Anticipatory Regulation of Performance: The Physiological Basis for Pacing Strategies and the Development of a Perception-Based Model for Exercise Performance. Br. J. Sports Med. 2009, 43, 392–400. [Google Scholar] [CrossRef]
- Vigotsky, A.D.; Halperin, I.; Lehman, G.J.; Trajano, G.S.; Vieira, T.M. Interpreting Signal Amplitudes in Surface Electromyography Studies in Sport and Rehabilitation Sciences. Front. Physiol. 2017, 8, 985. [Google Scholar] [CrossRef]
- Allen, D.G.; Lamb, G.D.; Westerblad, H. Skeletal Muscle Fatigue: Cellular Mechanisms. Physiol. Rev. 2008, 88, 287–332. [Google Scholar] [CrossRef]
- Bigland-Ritchie, B.; Furbush, F.; Woods, J.J. Fatigue of Intermittent Submaximal Voluntary Contractions: Central and Peripheral Factors. J. Appl. Physiol. 1986, 61, 421–429. [Google Scholar] [CrossRef]
- Matkowski, B.; Place, N.; Martin, A.; Lepers, R. Neuromuscular Fatigue Differs Following Unilateral vs Bilateral Sustained Submaximal Contractions. Scand. J. Med. Sci. Sports 2011, 21, 268–276. [Google Scholar] [CrossRef]
- Lamb, G.D. Mechanisms of Excitation-Contraction Uncoupling Relevant to Activity-Induced Muscle Fatigue. Appl. Physiol. Nutr. Metab. Physiol. Appl. Nutr. Metab. 2009, 34, 368–372. [Google Scholar] [CrossRef] [PubMed]
- McKay, A.K.A.; Stellingwerff, T.; Smith, E.S.; Martin, D.T.; Mujika, I.; Goosey-Tolfrey, V.L.; Sheppard, J.; Burke, L.M. Defining Training and Performance Caliber: A Participant Classification Framework. Int. J. Sports Physiol. Perform. 2022, 17, 317–331. [Google Scholar] [CrossRef] [PubMed]
- ACSM. ACSM’s Guidelines for Exercise Testing and Prescription; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2020; ISBN 978-1-975150-21-1. [Google Scholar]
- Lagally, K.M.; Robertson, R.J. Construct Validity of the OMNI Resistance Exercise Scale. J. Strength Cond. Res. 2006, 20, 252–256. [Google Scholar] [CrossRef]
- Robertson, R.J.; Goss, F.L.; Rutkowski, J.; Lenz, B.; Dixon, C.; Timmer, J.; Frazee, K.; Dube, J.; Andreacci, J. Concurrent Validation of the OMNI Perceived Exertion Scale for Resistance Exercise. Med. Sci. Sports Exerc. 2003, 35, 333–341. [Google Scholar] [CrossRef]
- Kulig, K.; Andrews, J.G.; Hay, J.G. Human Strength Curves. Exerc. Sport Sci. Rev. 1984, 12, 417–466. [Google Scholar] [CrossRef]
- Gearhart, R.E.; Goss, F.L.; Lagally, K.M.; Jakicic, J.M.; Gallagher, J.; Robertson, R.J. Standardized Scaling Procedures for Rating Perceived Exertion during Resistance Exercise. J. Strength Cond. Res. 2001, 15, 320–325. [Google Scholar]
- Keller, J.L.; Housh, T.J.; Hill, E.C.; Smith, C.M.; Schmidt, R.J.; Johnson, G.O. Neuromuscular Responses of Recreationally Active Women during a Sustained, Submaximal Isometric Leg Extension Muscle Action at a Constant Perception of Effort. Eur. J. Appl. Physiol. 2018, 118, 2499–2508. [Google Scholar] [CrossRef]
- Albertus, Y.; Tucker, R.; St Clair Gibson, A.; Lambert, E.V.; Hampson, D.B.; Noakes, T.D. Effect of Distance Feedback on Pacing Strategy and Perceived Exertion during Cycling. Med. Sci. Sports Exerc. 2005, 37, 461–468. [Google Scholar] [CrossRef]
- Hermens, H.J.; Freriks, B.; Disselhorst-Klug, C.; Rau, G. Development of Recommendations for SEMG Sensors and Sensor Placement Procedures. J. Electromyogr. Kinesiol. Off. J. Int. Soc. Electrophysiol. Kinesiol. 2000, 10, 361–374. [Google Scholar] [CrossRef]
- Kwatny, E.; Thomas, D.H.; Kwatny, H.G. An Application of Signal Processing Techniques to the Study of Myoelectric Signals. IEEE Trans. Biomed. Eng. 1970, 17, 303–313. [Google Scholar] [CrossRef]
- Shechtman, O. The Coefficient of Variation as an Index of Measurement Reliability. In Methods of Clinical Epidemiology; Doi, S.A.R., Williams, G.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 39–49. ISBN 978-3-642-37131-8. [Google Scholar]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences, 2nd ed.; Routledge: New York, NY, USA, 1988; ISBN 978-0-203-77158-7. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Test Visit 1 | Test Visit 2 | P | ICC | ICC95% | SEM | CV (%) |
|---|---|---|---|---|---|---|---|
| MVIC (mean ± SD) | |||||||
| Torque (N∙m) | 74.2 ± 12.8 | 75.9 ± 12.9 | 0.401 | 0.822 | 0.588 to 0.936 | 5.45 | 16.9 |
| Neuromuscular Parameters (mean ± SD) | |||||||
| EMG AMP (µVrms) | 992.7 ± 491.2 | 954.1 ± 553.2 | 0.684 | 0.773 | 0.446 to 0.918 | 248.59 | 52.8 |
| EMG MPF (Hz) | 83.2 ± 17.4 | 83.7 ± 15.6 | 0.927 | 0.321 | −0.245 to 0.712 | 13.77 | 19.5 |
| PTT (N∙m) | 9.3 ± 2.6 | 7.8 ± 3.6 | 0.162 | 0.256 | −0.226 to 0.657 | 2.67 | 36.9 |
| Anchor Scheme | RPEFT | TRQFT | ||||
|---|---|---|---|---|---|---|
| Participants | Pretest | Posttest | % | Pretest | Posttest | % |
| 1 | 91.7 | 81.7 | 10.9 | 90.4 | 72.7 | 19.6 |
| 2 | 75.8 | 47.0 | 38.0 | 74.5 | 64.5 | 13.5 |
| 3 | 50.8 | 45.1 | 11.2 | 53.5 | 53.0 | 0.8 |
| 4 | 70.9 | 66.1 | 6.8 | 79.9 | 51.6 | 35.5 |
| 5 | 79.8 | 64.3 | 19.5 | 72.0 | 54.3 | 24.6 |
| 6 | 103.6 | 99.3 | 4.2 | 89.6 | 71.6 | 20.0 |
| 7 | 76.9 | 60.4 | 21.5 | 86.7 | 68.1 | 21.5 |
| 8 | 70.0 | 71.9 | −2.7 | 76.6 | 61.0 | 20.3 |
| 9 | 74.0 | 65.2 | 11.9 | 71.5 | 58.3 | 18.5 |
| 10 | 75.4 | 62.7 | 16.9 | 80.9 | 67.4 | 16.8 |
| 11 | 69.1 | 67.5 | 2.4 | 81.5 | 54.7 | 32.8 |
| 12 | 84.9 | 76.2 | 10.3 | 98.1 | 81.7 | 16.7 |
| 13 | 60.2 | 52.2 | 13.3 | 55.4 | 43.7 | 21.1 |
| 14 | 65.5 | 58.4 | 10.9 | 64.2 | 50.3 | 21.7 |
| 15 | 64.1 | 54.1 | 15.6 | 63.8 | 50.0 | 21.6 |
| Mean ± SD | 74.2 ± 12.8 | 64.8 ± 13.9 | 12.7 ± 9.5 | 75.9 ± 12.8 | 60.2 ± 10.5 | 20.3 ± 7.9 |
| Anchor Scheme | RPEFT | TRQFT | ||||
|---|---|---|---|---|---|---|
| Participants | Pretest | Posttest | % | Pretest | Posttest | % |
| 1 | 828.7 | 684.6 | 17.4 | 502.1 | 799.1 | −59.2 |
| 2 | 817.8 | 778.9 | 4.8 | 1064.2 | 1114.3 | −4.7 |
| 3 | 440.8 | 457.7 | −3.8 | 639.9 | 847.8 | −32.5 |
| 4 | 1478.3 | 924.9 | 37.4 | 1454.1 | 1091.5 | 24.9 |
| 5 | 857.4 | 1037.1 | −21.0 | 940.6 | 796.9 | 15.3 |
| 6 | 492.9 | 462.5 | 6.2 | 1016.4 | 463.2 | 54.4 |
| 7 | 1099.1 | 1013.9 | 7.7 | 871.5 | 1233.7 | −41.6 |
| 8 | 349.8 | 374.2 | −7.0 | 617.1 | 743.5 | −20.5 |
| 9 | 1267.4 | 623.4 | 50.8 | 1015.2 | 1024.1 | −0.9 |
| 10 | 1310.8 | 761.0 | 41.9 | 791.8 | 683.4 | 13.7 |
| 11 | 430.0 | 674.8 | −56.9 | 629.5 | 331.5 | 47.3 |
| 12 | 2223.0 | 1977.2 | 11.1 | 2617.0 | 1490.4 | 43.1 |
| 13 | 1267.7 | 1088.6 | 14.1 | 1265.3 | 1675.6 | −32.4 |
| 14 | 1054.7 | 846.6 | 19.7 | 639.2 | 459.4 | 28.1 |
| 15 | 972.4 | 488.7 | 49.7 | 248.1 | 691.2 | −178.6 |
| Mean ± SD | 992.7 ± 491.2 | 812.9 ± 392.6 | 11.5 ± 28.2 | 954.1 ± 553.2 | 896.4 ± 378.5 | −9.6 ± 58.1 |
| Anchor Scheme | RPEFT | TRQFT | ||||
|---|---|---|---|---|---|---|
| Participants | Pretest | Posttest | % | Pretest | Posttest | % |
| 1 | 63.3 | 78.7 | −24.2 | 76.7 | 57.7 | 24.8 |
| 2 | 63.2 | 46.1 | 27.1 | 72.7 | 60.9 | 16.2 |
| 3 | 57.9 | 67.6 | −16.8 | 52.8 | 57.1 | −8.1 |
| 4 | 81.7 | 81.5 | 0.2 | 71.3 | 62.6 | 12.2 |
| 5 | 108.0 | 85.1 | 21.3 | 75.4 | 68.4 | 9.4 |
| 6 | 86.0 | 67.1 | 22.0 | 97.3 | 74.8 | 23.2 |
| 7 | 87.3 | 95.9 | −9.9 | 111.4 | 99.3 | 10.8 |
| 8 | 106.3 | 119.1 | −12.1 | 111.7 | 73.5 | 34.2 |
| 9 | 78.8 | 113.0 | −43.3 | 92.8 | 77.9 | 16.1 |
| 10 | 112.6 | 116.0 | −2.9 | 76.4 | 65.8 | 13.9 |
| 11 | 90.3 | 74.0 | 18.0 | 76.3 | 67.1 | 12.1 |
| 12 | 82.4 | 77.3 | 6.2 | 82.3 | 80.0 | 2.8 |
| 13 | 96.6 | 101.1 | −4.7 | 78.7 | 61.0 | 22.5 |
| 14 | 69.9 | 63.5 | 9.1 | 86.9 | 69.5 | 20.1 |
| 15 | 64.2 | 71.2 | −10.9 | 92.5 | 68.9 | 25.5 |
| Mean ± SD | 83.2 ± 17.4 | 83.8 ± 21.2 | −1.4 ± 19.3 | 83.7 ± 15.6 | 69.6 ± 10.8 | 15.7 ± 10.2 |
| Anchor Scheme | RPEFT | TRQFT | ||||
|---|---|---|---|---|---|---|
| Participants | Pretest | Posttest | % | Pretest | Posttest | % |
| 1 | 11.3 | 9.9 | 12.4 | 9.5 | 5.8 | 38.9 |
| 2 | 9.6 | 3.7 | 61.5 | 7.5 | 3.2 | 57.3 |
| 3 | 8.3 | 8.7 | −4.8 | 7.3 | 2 | 72.6 |
| 4 | 7.3 | 5.8 | 20.5 | 5.5 | 3.3 | 40.0 |
| 5 | 7.7 | 7.3 | 5.2 | 8.1 | 3.8 | 53.1 |
| 6 | 8.6 | 6.3 | 26.7 | 5.3 | 1.3 | 75.5 |
| 7 | 5.5 | 4.7 | 14.5 | 5.3 | 2.8 | 47.2 |
| 8 | 7.7 | 6.8 | 11.7 | 7.5 | 2.9 | 61.3 |
| 9 | 9.9 | 7.1 | 28.3 | 6.8 | 4.2 | 38.2 |
| 10 | 12.7 | 9.9 | 22.0 | 4.9 | 3.7 | 24.5 |
| 11 | 14.3 | 13.2 | 7.7 | 10 | 6.6 | 34.0 |
| 12 | 12 | 11.9 | 0.8 | 11.4 | 8.4 | 26.3 |
| 13 | 5 | 5.2 | −4.0 | 5 | 1.9 | 62.0 |
| 14 | 9.1 | 7.7 | 15.4 | 18.6 | 6.1 | 67.2 |
| 15 | 10.3 | 5.1 | 50.5 | 5 | 2.7 | 46.0 |
| Mean ± SD | 9.3 ± 2.6 | 7.6 ± 2.7 | 17.9 ± 18.5 | 7.8 ± 3.6 | 3.9 ± 2.0 | 49.6 ± 16.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smith, R.W.; Arnett, J.E.; Ortega, D.G.; Roberts, T.D.; McCanlies, D.J.; Schmidt, R.J.; Johnson, G.O.; Housh, T.J. Maximal Torque, Neuromuscular, and Potentiated Twitch Responses to Sustained Forearm Flexion Tasks Using Different Anchor Schemes. Physiologia 2025, 5, 15. https://doi.org/10.3390/physiologia5020015
Smith RW, Arnett JE, Ortega DG, Roberts TD, McCanlies DJ, Schmidt RJ, Johnson GO, Housh TJ. Maximal Torque, Neuromuscular, and Potentiated Twitch Responses to Sustained Forearm Flexion Tasks Using Different Anchor Schemes. Physiologia. 2025; 5(2):15. https://doi.org/10.3390/physiologia5020015
Chicago/Turabian StyleSmith, Robert W., Jocelyn E. Arnett, Dolores G. Ortega, Trevor D. Roberts, Dona J. McCanlies, Richard J. Schmidt, Glen O. Johnson, and Terry J. Housh. 2025. "Maximal Torque, Neuromuscular, and Potentiated Twitch Responses to Sustained Forearm Flexion Tasks Using Different Anchor Schemes" Physiologia 5, no. 2: 15. https://doi.org/10.3390/physiologia5020015
APA StyleSmith, R. W., Arnett, J. E., Ortega, D. G., Roberts, T. D., McCanlies, D. J., Schmidt, R. J., Johnson, G. O., & Housh, T. J. (2025). Maximal Torque, Neuromuscular, and Potentiated Twitch Responses to Sustained Forearm Flexion Tasks Using Different Anchor Schemes. Physiologia, 5(2), 15. https://doi.org/10.3390/physiologia5020015