This study provides comprehensive information on the biology and population dynamics of an important foraging species and a component of the trophic chain in the Bay of Biscay [
37,
56],
M. muelleri. In the absence of exploratory campaigns capable of sampling the entire annual cycle of the species, we provide, for the first time, both information on the biology of this species and a global view of the population’s temporal dynamics. We are aware that some of the models fitted for the estimation of population parameters would be more reliable and robust if the quality of sampling was better. However, despite this setback, we obtained estimates of key demographic parameters for the population of
M. muelleri inhabiting the Bay of Biscay that largely resemble those previously reported for this species in other areas of the northeast Atlantic (as we discuss below). Based on three months of sampling and the size distribution and ages, we have been able to discern a temporal cycle for
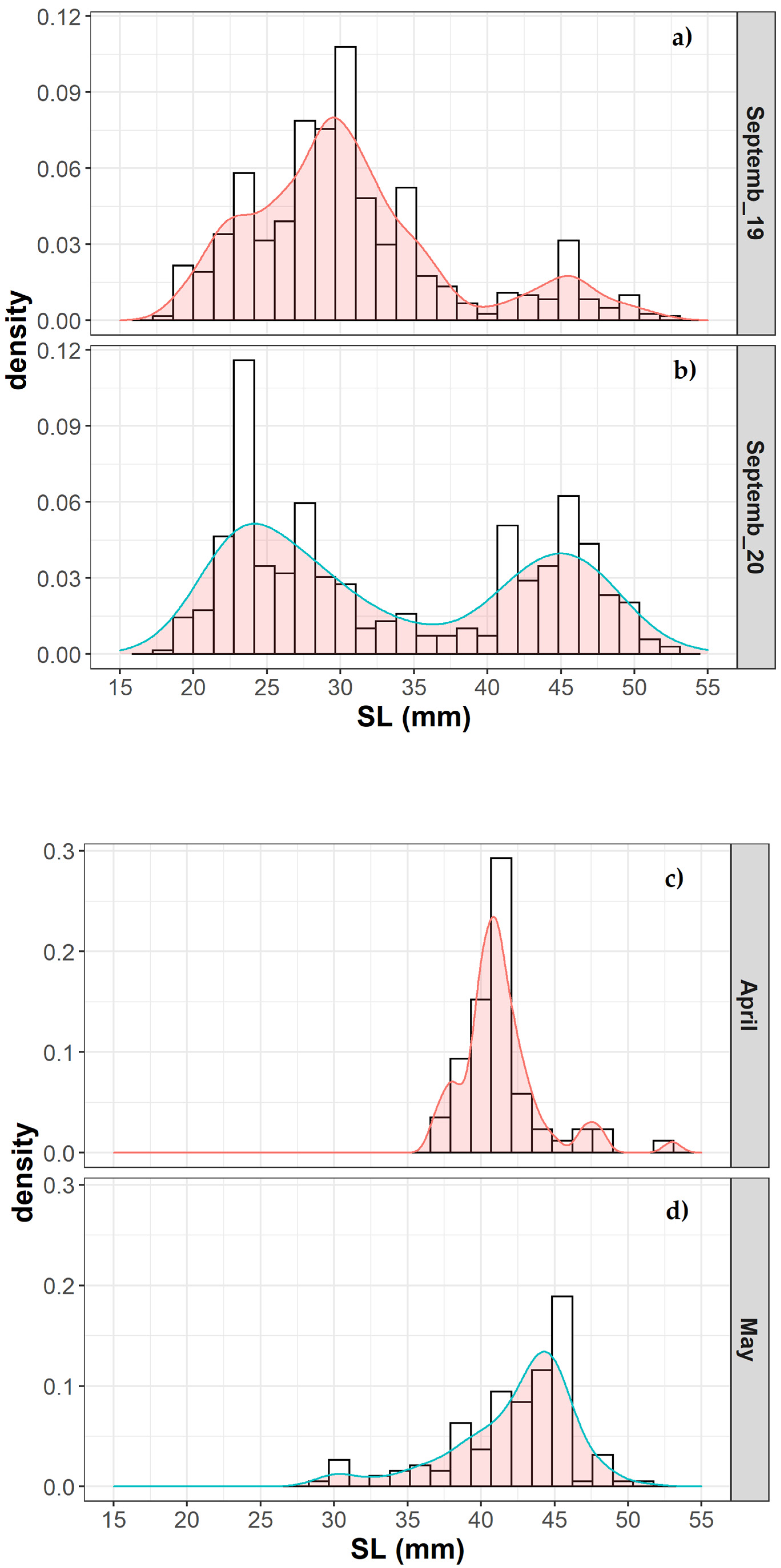
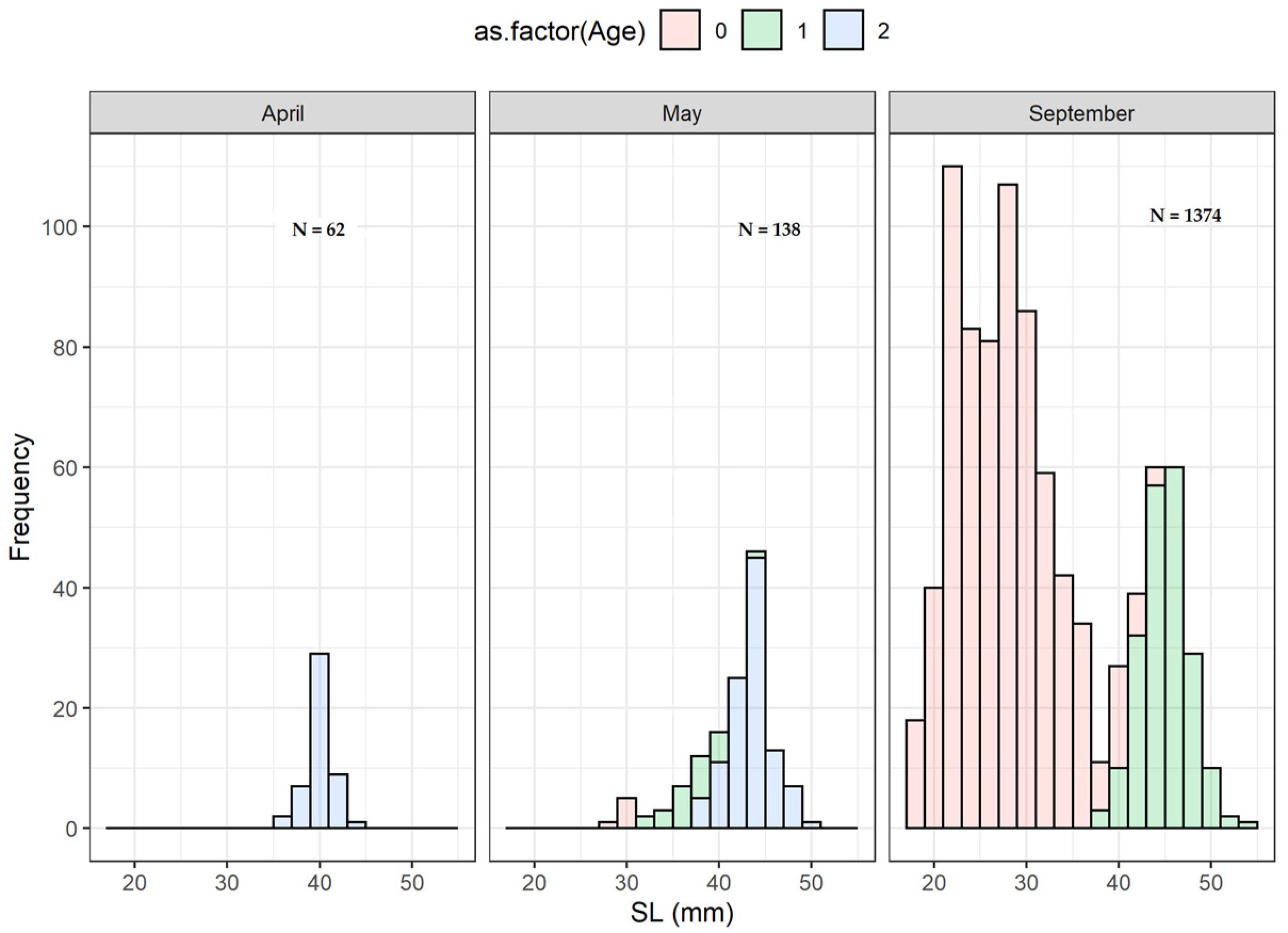
M. muelleri. Thus, the average length of the sampled individuals was different in spring and summer. While in September, most of the population consisted of juveniles (immatures, age group 0) or young adults (age group 1), adults dominated the population in April–May (mature, age groups 1 and 2).
In temperate and subtropical waters,
M. muelleri generally spawns from late winter to early autumn (see
Table 10). References are scarce for the Bay of Biscay, but eggs and larvae occur from February to June according to Valencia et al. [
33], Rodriguez et al. [
34], and Rodriguez [
35]. In the survey data used for this study (JUVENA, MEGS, and BIOMAN surveys), early life stages were observed in March, April, and May, but not in September. However, mature individuals with spawning markers, i.e., hydrated oocytes, were found in September of 2020 (
Table 7), indicating a spawning capability in late summer. Kawaguchi and Mauchline [
13] identified a prolonged spawning season of about six months for this species in the area of Rockall Trough, describing the presence of juveniles in autumn and winter, mixed with late larvae or adults, respectively. The range of modes observed in the juvenile size distributions in September in the Bay of Biscay and the presence of juveniles in April suggest a potentially protracted spawning season. The poor representation of larger fish in late summer could be explained by different causes. On the one hand,
M. muelleri are known to perform DVMs related to foraging [
16]. DVMs were also detected in the Bay of Biscay [
56], although our stomach content index did not mirror an obvious relation with the DVM, likely because of the absence of nocturnal samples (
Figure S2). Some authors have found that larger adult fish do not perform DVMs [
14,
15], which could lead to reduced accessibility to these larger fish if the net does not reach the depth where they are found. This does not seem to be the situation in the Bay of Biscay, since the vertical distribution range of this species mainly comprises the first 400 m of the water column [
56], which is usually sampled with the net. However, net avoidance may be higher at these size ranges than at smaller sizes (faster swimming of big-sized specimens), which would also contribute to a lower accessibility [
57]. On the other hand, if the higher post-breeding mortality observed by Gjøsæter [
28] in Norwegian waters also occurs in southern areas (as in Rockall Trough [
13]), it would be expected that the proportion of adult fish of 1 or 2 years old in late summer would be reduced. Interannual differences can also occur, as reflected by the higher percentage of larger fish (SL > 40 mm, age group 1) in 2020 compared to 2019. In order to explain these differences, some hypotheses can be put forward. In line with the observations of other authors regarding post-spawning mortality, this interannual change in the adult fish proportion might be interpreted in terms of lower mortality rates for adult spawners in 2020 at the end of the spawning season. In this case, we might attribute particularly unfavourable post-spawning hydrographic conditions in one year compared to another as the driver of these changes. For instance, Rosland and Giske [
58] stated that turbidity close to the surface would benefit
M. muelleri, since the visibility of piscivores with long reaction distances would decrease more than the visibility of zooplanktivores with short reaction distances. The estimated wind-induced mixing in the Bay of Biscay for these years (
Figure S3) showed no appreciable variability between years, so this mixing effect was insufficient to cause a singular effect on the M. muelleri mortality. An alternative hypothesis, and likely the simplest and most reasonable, is that differences in the population biomass are responsible for this perception (the estimates of biomass in the area were 157 and 208 ktons for 2019 and 2020, respectively—Guillermo Boyra, personal communication). Assuming a similar proportion of adults each year, it would be expected that the probability of catching adult fish in 2020 would be higher, as we indeed observed for this year.
4.2. GSI and Sex Ratio
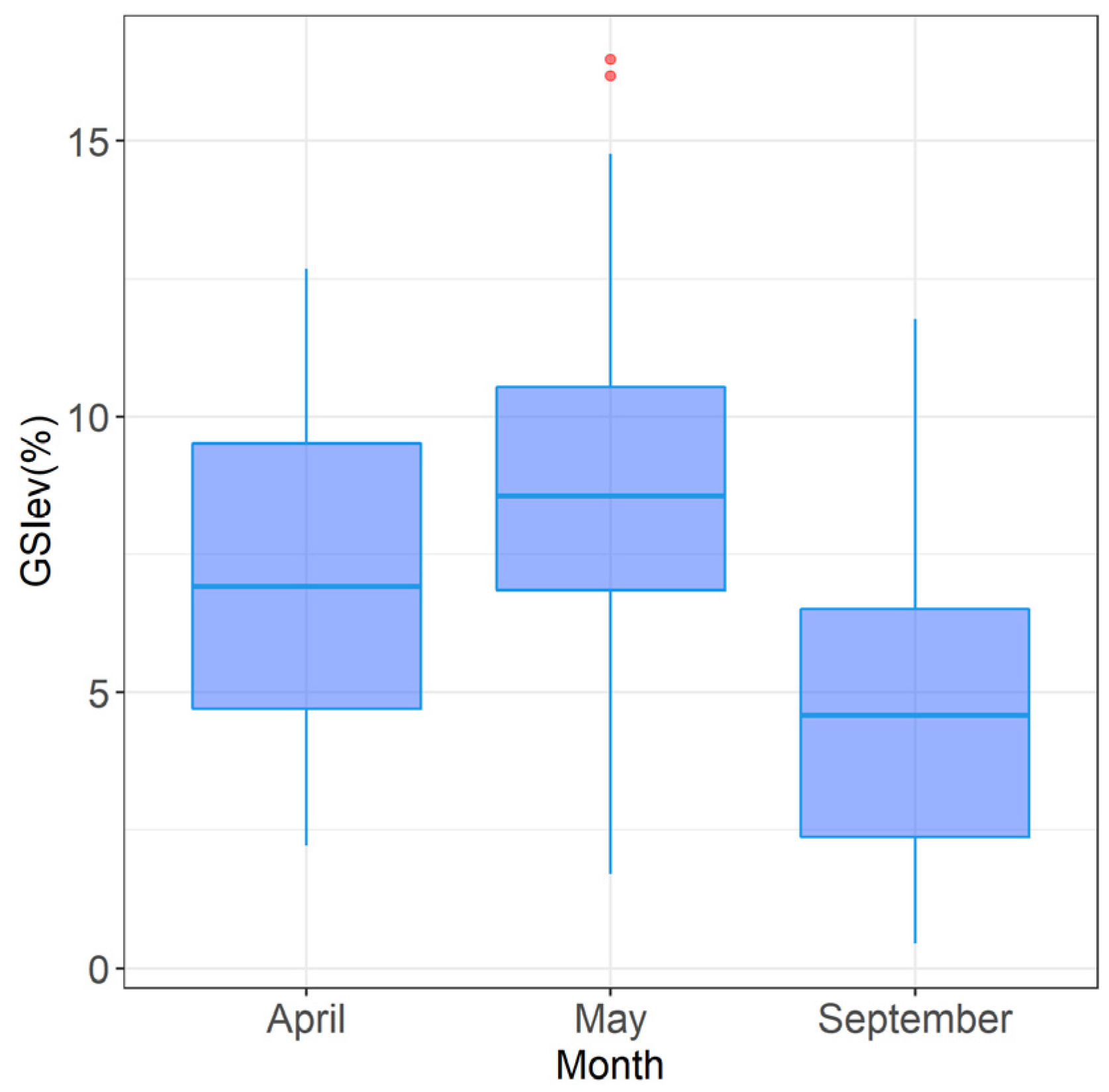
The range of values of the GSI estimated here was significantly higher than those described in the literature. The maximum value of the GSI (16%, in May) contrasts with those reported by Rasmussen and Giske [
17] and Salvanes and Stockley [
60] in Norway (7% and 5.6%, respectively), and Clark [
6] in Australia (9.6%). Salvanes and Stockley [
60] found that the highest averages for the GSI were closely related to the diameter of the oocytes, that is to say, to the maturation process of the fish gonads. In this study, the occurrence of fish with high GSI values (about 8%) in September as well denoted that a small proportion of
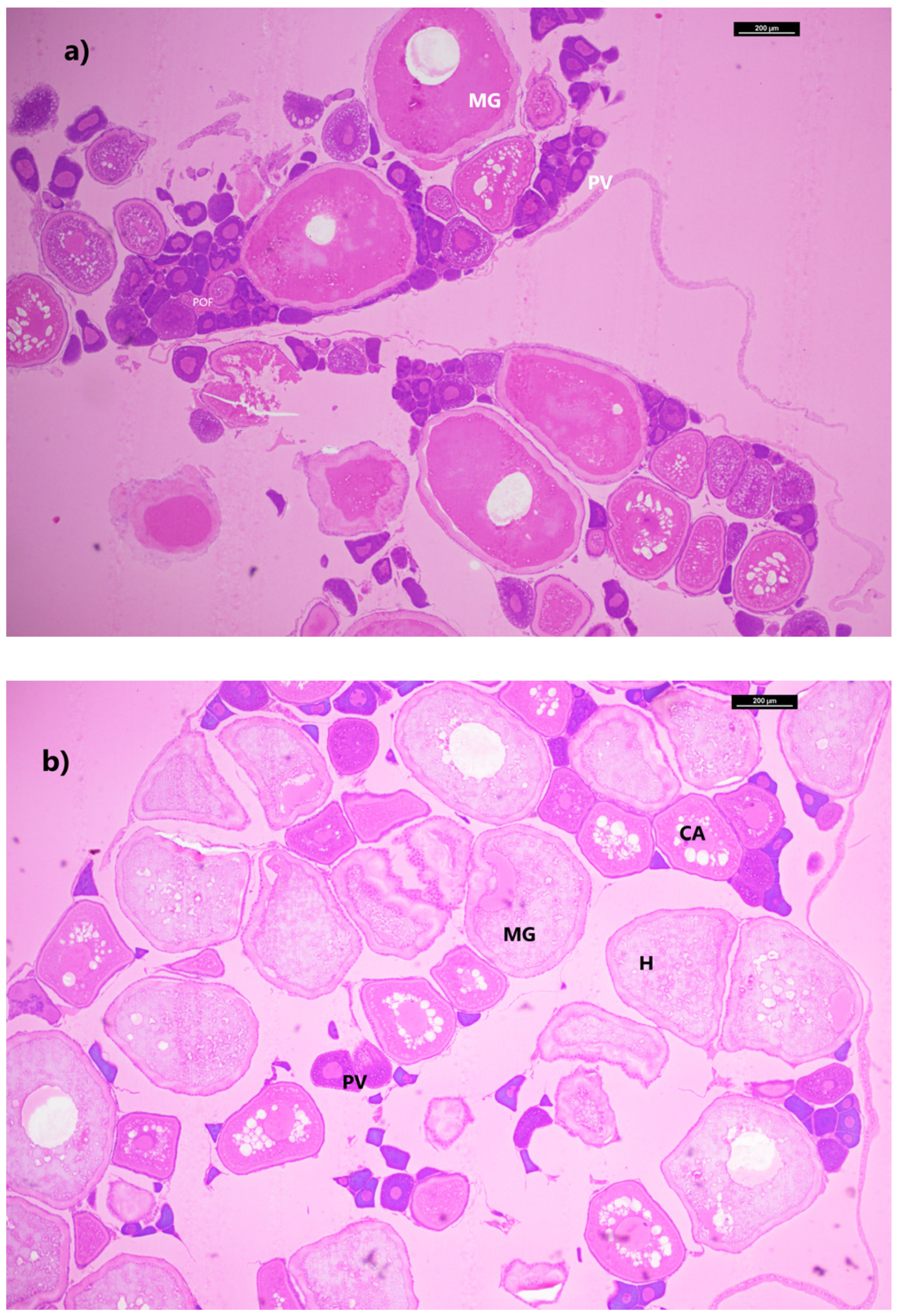
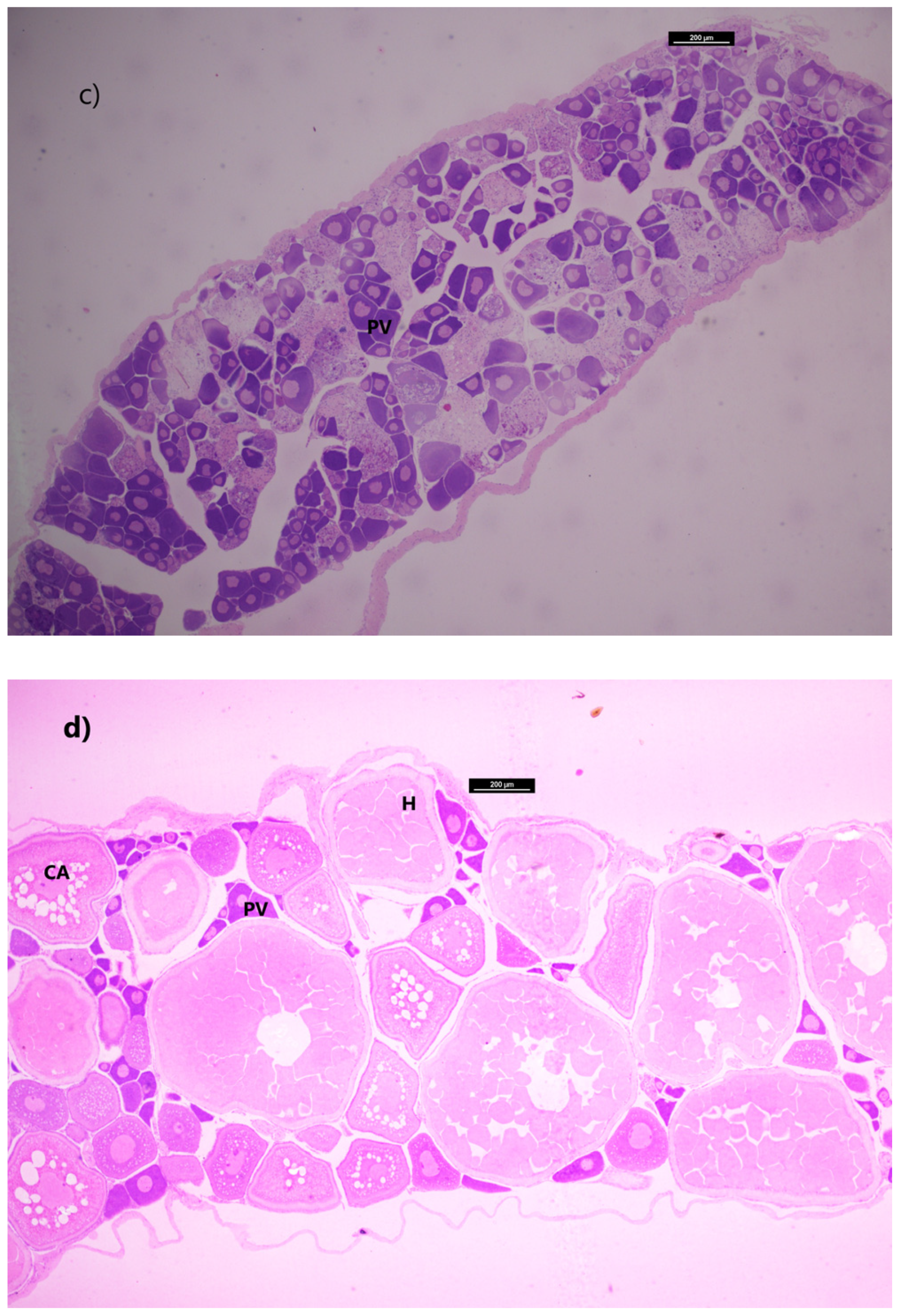
M. muelleri exhibited reproductive activity in late summer. Higher GSI values in May indicated that there was a greater investment in reproduction during this period, which coincided with the presence of the more advanced stages of maturation in the gonad (
Figure 6). This result demonstrates that the GSI may be used as a suitable tool for the evaluation of gonadal maturation in
M. muelleri.
M. muelleri exhibits a positive bias in the ratio of females to males in larger fish groups [
6,
17,
18,
29,
61]. In the samples taken in this study, males were most abundant at smaller size ranges, while females dominated progressively among larger sizes at a rate of 11% with the size. The overall ratio was slightly in favour of females at 0.44:0.56, whereas an equal proportion of males to females was predicted at 41.5 mm. Although this phenomenon seems to be common in mesopelagic species, it has not been observed in other populations of the same gender [
62]. Different theories have been proposed to explain this difference in size. On the one hand, Rasmussen and Giske [
17] suggested different mortality rates associated with sex or sexual differences in growth. On the other hand, Kristoffersen and Salvanes [
61] proposed sex segregation by depth, which would change with age. More recently, Staby et al. [
14] observed in Norwegian fjords that post-larvae and juvenile fish performed a normal crepuscular migration, while most of the adults remained at lower depths throughout the diel period. If male growth is lower than female growth, the juvenile phase of males should last longer, and the sex ratio should be in preference to juvenile males in the lower ranges. No clear evidence regarding a differential growth between sexes was found, probably because it was not possible to sex juvenile individuals. If there was vertical segregation by sex or size in the Bay of Biscay, sampling should cover the entire vertical distribution of the species to avoid bias in the sex ratio estimation. According to Sobradillo et al. [
56],
M. muelleri is preferentially distributed in a range of depths between 50 and 400 m, which corresponds to the usual depth at which the samples were taken in this study. Therefore, if one rules out these factors as likely to be responsible for the change in the sex ratio proportion by size, it seems that differential mortality may be the cause of the higher presence of females in larger fish groups.
4.3. Female Maturity
This is the first time that data on the reproductive parameters in terms of ovarian development, length at maturity, maturity ogive, and batch fecundity are presented for
M. muelleri in the Bay of Biscay. The SL
50 was only slightly larger in females than in males (35.7 mm and 36.1, respectively) and somewhat lower in sex-combined than in sex-separated samples (34.1 mm). There is no information on this parameter for
M. muelleri, but for sex-combined samples, our estimate for SL
50 was slightly larger than that of
M. sthemanni (SL
50 = 32 mm), according to Almeida et al. [
62]. The age and length at maturation show considerable variability within the same species, and this plasticity seems to be genetically defined and modelled by environmental variables [
63]. For instance, the results showed that the size of the smallest mature female was 30 mm (by histological observation), which is within the range reported in the literature for different areas (20 to 40 mm [
6,
14,
17,
29,
64,
65,
66,
67]). Nevertheless, it would have been reasonable to expect a lower size at maturity in the Bay of Biscay if the sampling during the spawning peak (May or June) had been more intense, since in this period, reproductive activity occurs amongst individuals with a wider range of sizes [
29].
Vazzoler (1996, in Almeida et al. [
62]) defined the “critical minimum size” as the length at which the critical reproductive processes start, and proposed that species with a short life span and a small Lmax (r strategists), as is the case with
M. muelleri, achieve mature gonadal stages before those species with a longer life span and a larger Lmax (k strategists).
The reproductive load (RL = Lm/Lmax) defines the relationship between the minimum size at maturation (Lm) and the maximum size a certain fish is likely to reach (Lmax). Froese and Binohland [
68] compiled information from the literature for over 1100 fish and found that the Lm is, above all, a function of size. The values of these parameters are determined mainly by the interaction between the supply of oxygen and its demand [
69]. According to these authors, the relationship varied between 0.4 and 0.9 and tended to be greater in smaller fish than in larger ones. With the aim of testing this hypothesis, we estimated this parameter (RL = 0.55) by taking the recorded minimum size at first maturation (SLm = 30 mm) and the maximum size observed (SLmax = 55 mm) of females as a reference. Our estimate was similar to the values reported by Almeida et al. [
62] (
Table 2) for the Maurolicus genus from different locations (about 0.5), with the exception of the populations located in Japan [
64] and Tasmania [
66]. This value is attributed to the ability of fish to perceive environmental stimuli that induce them to spawn [
70]. This relationship seems to be genus-specific, so it will remain invariant within the same taxonomic group, and though environmental changes may cause modifications in both the SL
50 and SLmax, they will not influence the relationship between these parameters.
4.4. Fecundity Strategy
In general terms, fish undergo two types of fecundity: determinate and indeterminate [
71], and reproductive parameters have to be estimated according to the reproductive strategy of the fish. In the case of determinate fecundity, all oocytes predestined to be spawned would be recognized at the beginning of the spawning season, and no new spawning oocytes would be recruited from the primary growth stocks, as is the case with indeterminate species [
49]. Salvanes and Stockley [
60] assumed an indeterminate fecundity for
M. muelleri and estimated a total fecundity that ranged from 13,331 to 36,848 eggs in specimens from the northern Norwegian sea. Previously, Goodson et al. [
29] found that the total fecundity of
M. muelleri did not decrease when the spawning season progressed, indicating that new oocytes were being recruited. According to this evidence,
M. muelleri should be regarded as an indeterminate species, and for that reason, the yearly fecundity should be calculated by estimating the spawning frequency, i.e., the percentage of females spawning per day, and the batch fecundity [
51].
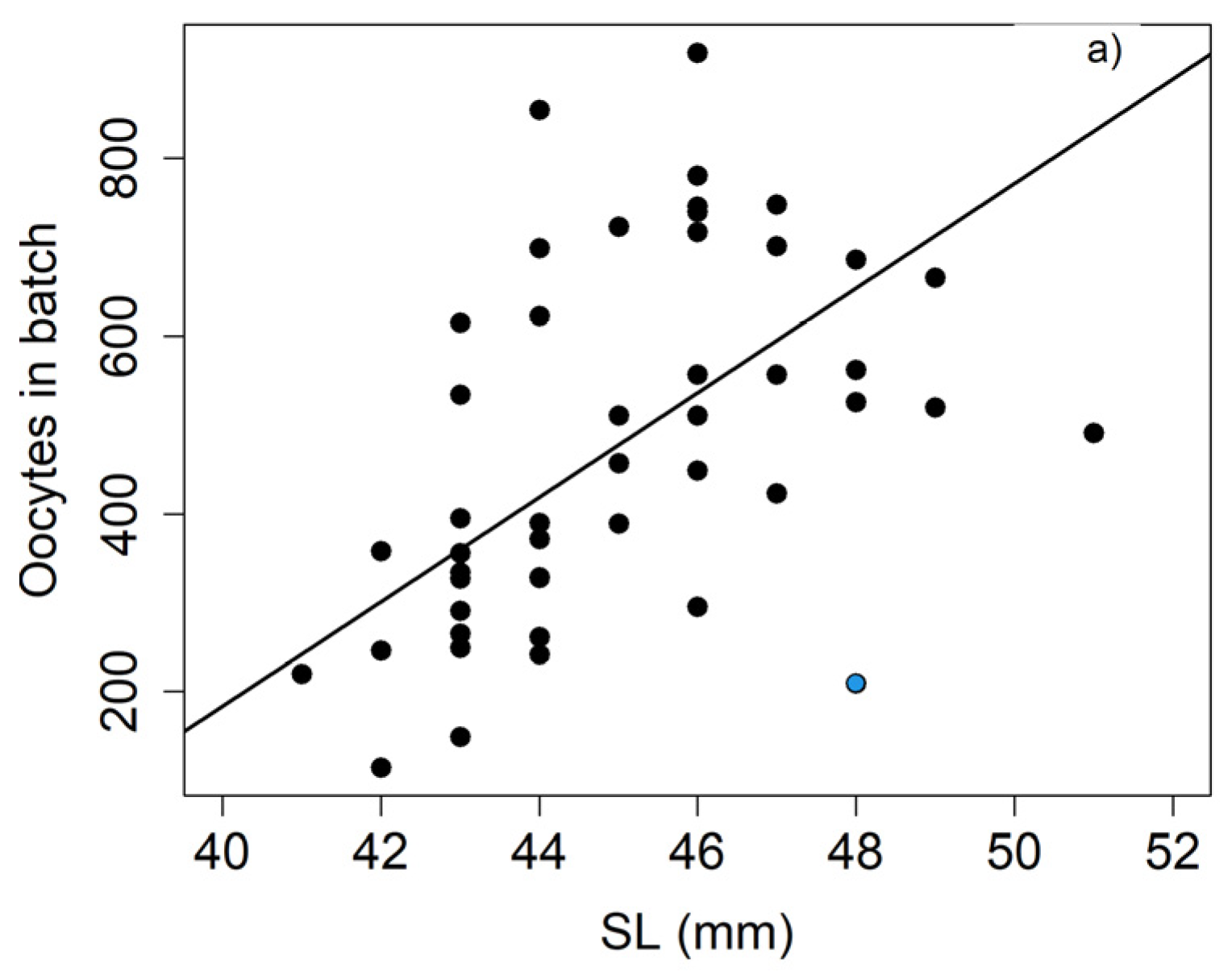
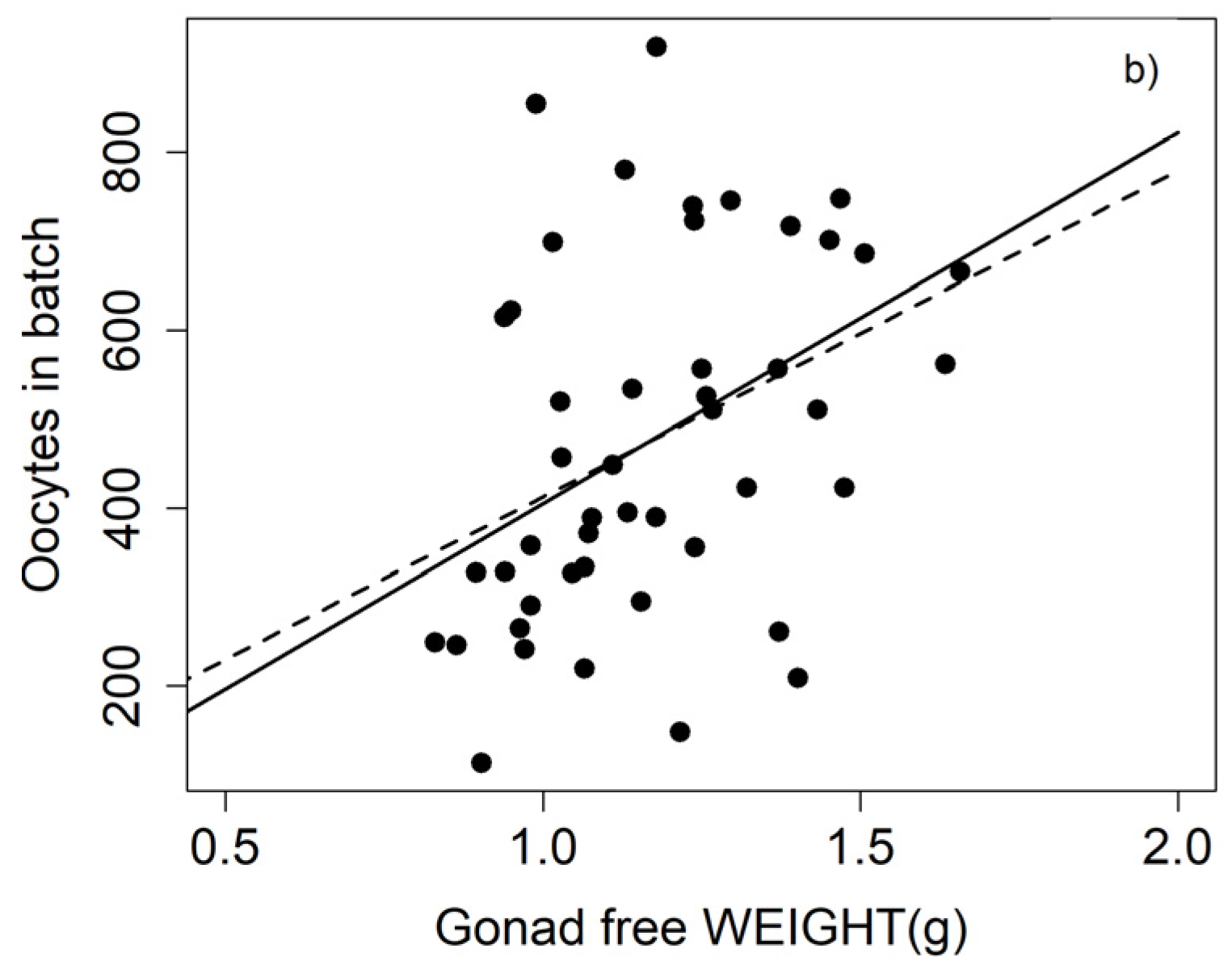
The batch fecundity was found to fluctuate greatly, both among individuals of the same population and between populations from different locations (
Table 10). Our estimates of batch fecundity in May seem to be in the upper range of the values observed in different populations of this species distributed worldwide for individuals of a similar size (
Table 10). This might mirror a differential life history strategy in the Bay of Biscay, where the fish, for the most part, seemingly reproduce no more than twice while alive and then die, while in other areas, fish live longer and take part in more than one spawning season. However, due to variations in the oocyte size thresholds used to determine the batch fecundity on the one hand, and the distinct range of fish analysed in each study on the other, the comparisons should be treated with caution.
We found correlations between the batch fecundity and the size or weight, as has been found by other authors [
6,
13,
17,
72]. However, in other studies [
2,
8,
12,
66,
67], no significant relationship between these variables was noticed. The lack of correlation between batch fecundity and size/weight could be attributed to different factors: firstly, because the range of weights is too short to obtain a relationship between the parameters, and secondly, due to the high dispersion of the batch fecundity, mainly in larger individuals. The high scatter amongst these individuals may indicate that these fish have already begun to spawn, diminishing their fecundity and increasing the variance in the data. This can be largely avoided by selecting only ovaries that do not present spawning markers, such as post-ovulatory follicles, as we did in this study. This seems to be the reason why we obtained a significant correlation, even though the range of fish sizes was narrow.
In this study, we provide some new insights into the biological sustainability of M. muelli as a basis for stock assessment in the Bay of Biscay. However, the exploitation of this resource is delicate and there is a lack of information in this regard. To date, no trial fishing for mesopelagic fish has been conducted in the Bay of Biscay, and it is unknown whether these species are captured as a bycatch and discarded by the trawling fleet that operate in this region. Further actions focused on these gaps are strongly recommended to prevent the misuse of this potential resource.















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}