

Exploring Emerging Therapeutic Targets and Opportunities in Neuroendocrine Tumors: Updates on Receptor Tyrosine Kinases

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. EGFR
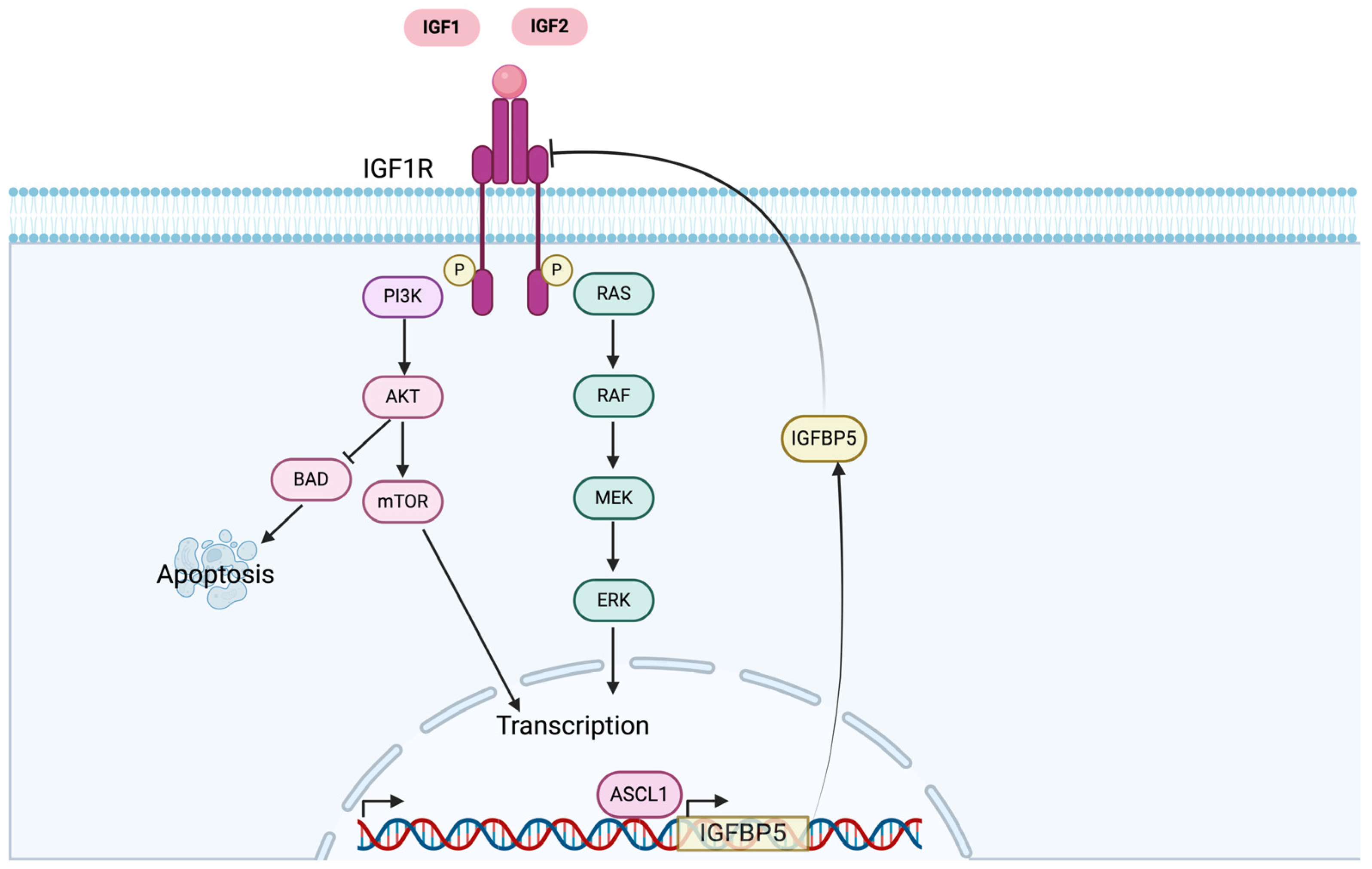
3. IGF-1R
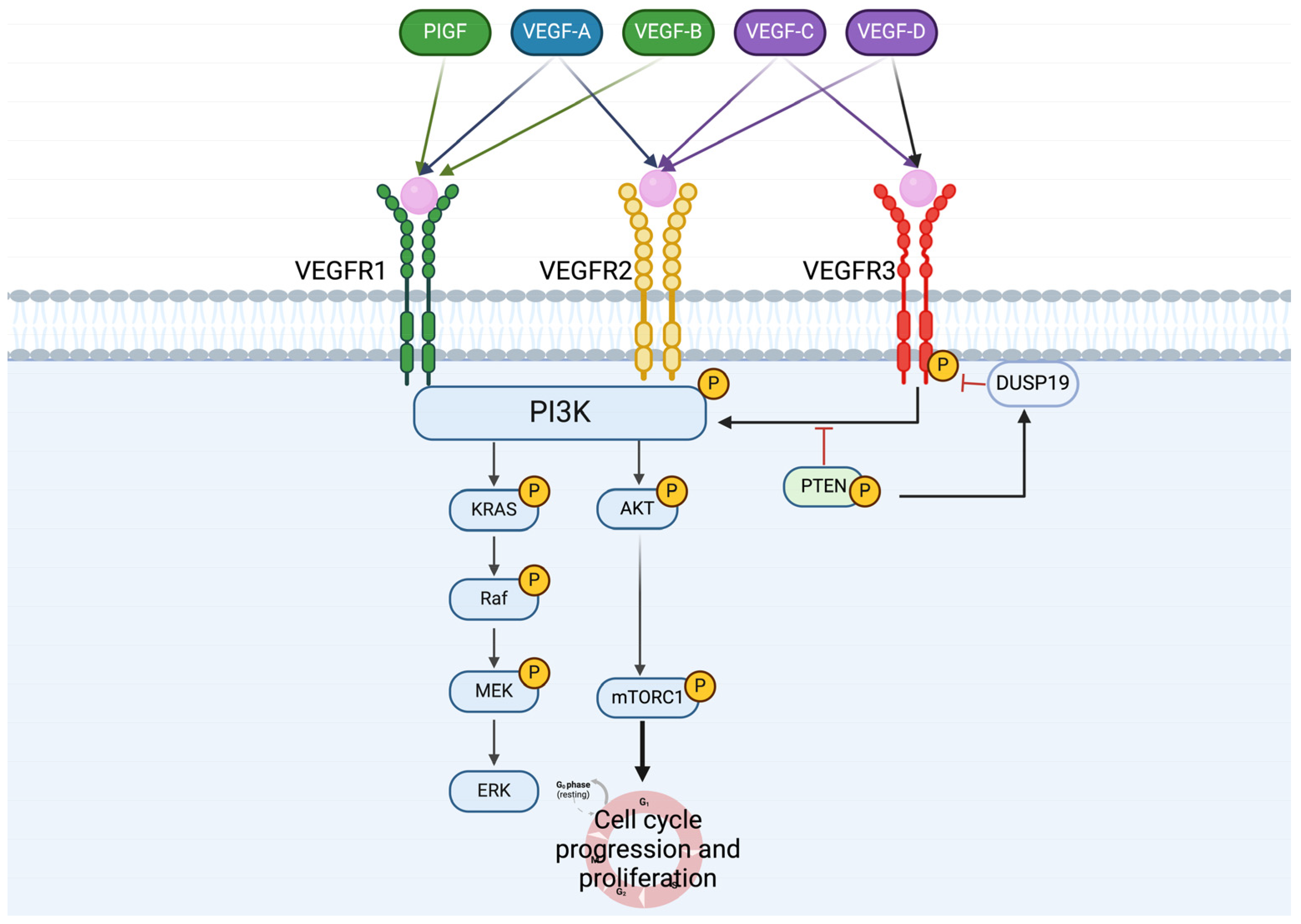
4. VEGFRs

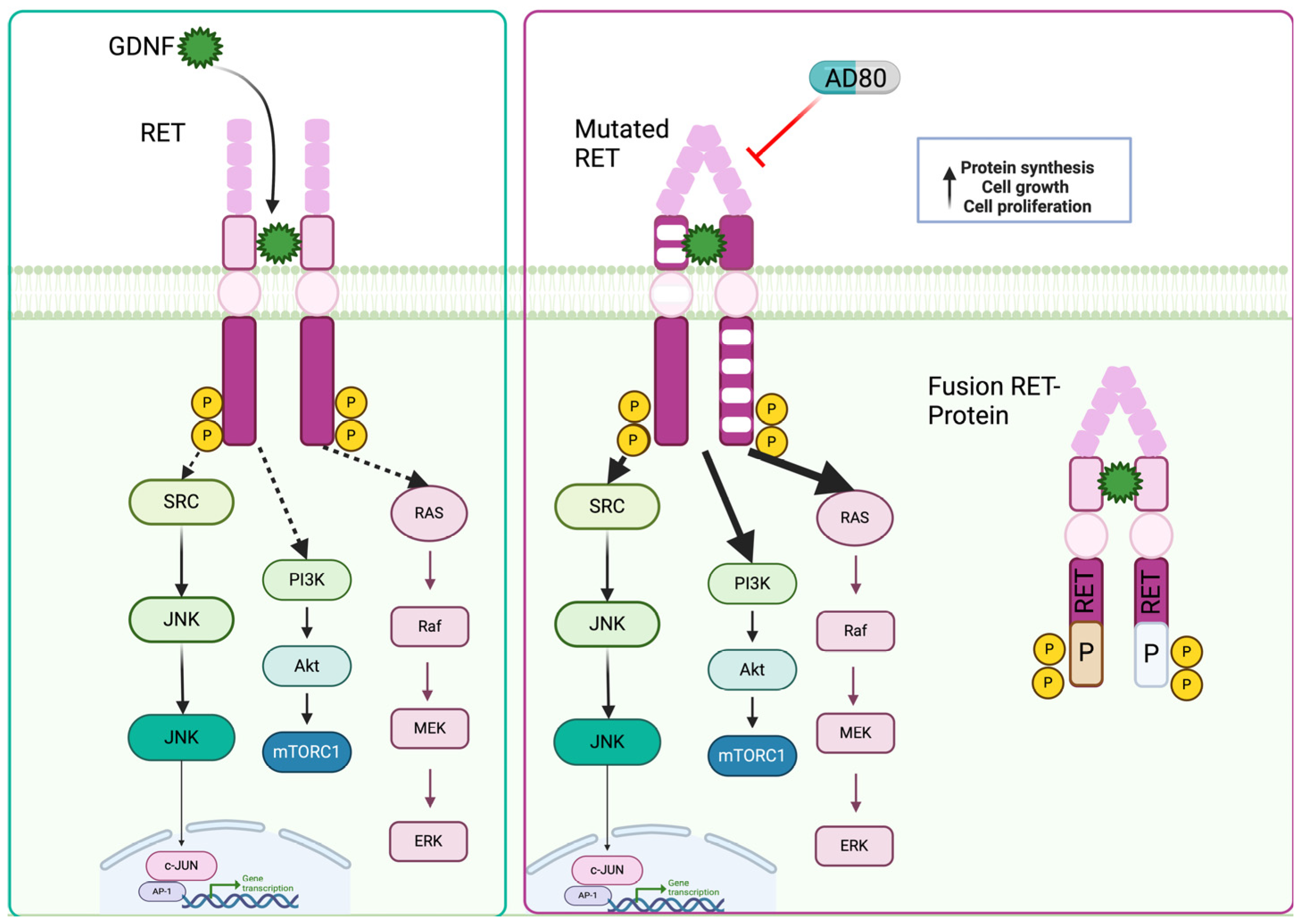
5. RET

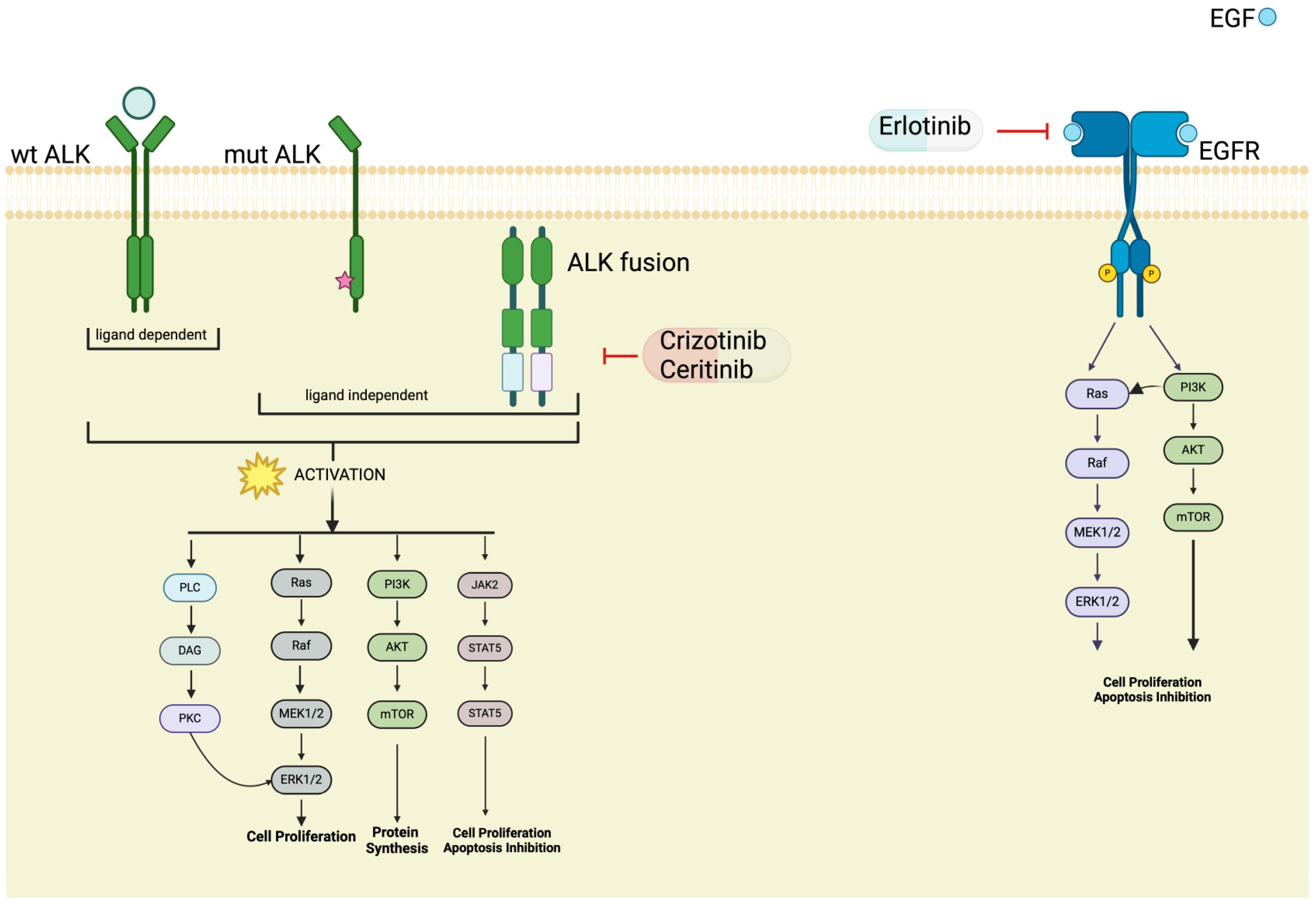
6. ALK

7. Discussion
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wu, H.; Yu, Z.; Liu, Y.; Guo, L.; Teng, L.; Guo, L.; Liang, L.; Wang, J.; Gao, J.; Li, R.; et al. Genomic Characterization Reveals Distinct Mutation Landscapes and Therapeutic Implications in Neuroendocrine Carcinomas of the Gastrointestinal Tract. Cancer Commun. 2022, 42, 1367–1386. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, S.E.; Dowrey, T.W.; Martin, C.V.; Bi, K.; Titchen, B.; Johri, S.; DelloStritto, L.; Patel, M.; Mackichan, C.; Inga, S.; et al. Intertumoral Lineage Diversity and Immunosuppressive Transcriptional Programs in Well-Differentiated Gastroenteropancreatic Neuroendocrine Tumors. Sci. Adv. 2023, 9, eadd9668. [Google Scholar] [CrossRef] [PubMed]
- Pavel, M.; Öberg, K.; Falconi, M.; Krenning, E.P.; Sundin, A.; Perren, A.; Berruti, A. Gastroenteropancreatic Neuroendocrine Neoplasms: ESMO Clinical Practice Guidelines for Diagnosis, Treatment and Follow-Up. Ann. Oncol. 2020, 31, 844–860. [Google Scholar] [CrossRef] [PubMed]
- Di Domenico, A.; Wiedmer, T.; Marinoni, I.; Perren, A. Genetic and Epigenetic Drivers of Neuroendocrine Tumours (NET). Endocr. Relat. Cancer 2017, 24, R315–R334. [Google Scholar] [CrossRef] [PubMed]
- Scarpa, A. The Landscape of Molecular Alterations in Pancreatic and Small Intestinal Neuroendocrine Tumours. Ann. Endocrinol. 2019, 80, 153–158. [Google Scholar] [CrossRef]
- Sakane, T.; Sakamoto, Y.; Masaki, A.; Murase, T.; Okuda, K.; Nakanishi, R.; Inagaki, H. Mutation Profile of Thymic Carcinoma and Thymic Neuroendocrine Tumor by Targeted Next-Generation Sequencing. Clin. Lung Cancer 2021, 22, 92–99.e4. [Google Scholar] [CrossRef] [PubMed]
- Asiedu, M.K.; Thomas, C.F.; Dong, J.; Schulte, S.C.; Khadka, P.; Sun, Z.; Kosari, F.; Jen, J.; Molina, J.; Vasmatzis, G.; et al. Pathways Impacted by Genomic Alterations in Pulmonary Carcinoid Tumors. Clin. Cancer Res. 2018, 24, 1691–1704. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, Y.; Umemura, S.; Ishii, G.; Tsuta, K.; Matsumoto, S.; Aokage, K.; Hishida, T.; Yoshida, J.; Ohe, Y.; Suzuki, H.; et al. Expression Profiling of Receptor Tyrosine Kinases in High-Grade Neuroendocrine Carcinoma of the Lung: A Comparative Analysis with Adenocarcinoma and Squamous Cell Carcinoma. J. Cancer Res. Clin. Oncol. 2015, 141, 2159–2170. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Lovly, C.M. Mechanisms of Receptor Tyrosine Kinase Activation in Cancer. Mol. Cancer 2018, 17, 58. [Google Scholar] [CrossRef] [PubMed]
- Uri, I.; Grozinsky-Glasberg, S. Current Treatment Strategies for Patients with Advanced Gastroenteropancreatic Neuroendocrine Tumors (GEP-NETs). Clin. Diabetes Endocrinol. 2018, 4, 16. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, F.; Albertelli, M.; Grillo, F.; Mohamed, A.; Saveanu, A.; Barlier, A.; Ferone, D.; Florio, T. Neuroendocrine Tumors: Insights into Innovative Therapeutic Options and Rational Development of Targeted Therapies. Drug Discov. Today 2014, 19, 458–468. [Google Scholar] [CrossRef] [PubMed]
- IEO ENETS Center of Excellence for GEP NETs; Martins, D.; Spada, F.; Lambrescu, I.; Rubino, M.; Cella, C.; Gibelli, B.; Grana, C.; Ribero, D.; Bertani, E.; et al. Predictive Markers of Response to Everolimus and Sunitinib in Neuroendocrine Tumors. Target. Oncol. 2017, 12, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Marinović, S.; Cigrovski Berković, M.; Zjačić-Rotkvić, V.; Kapitanović, S. Analysis of Polymorphisms in EGF, EGFR and HER2 Genes in Pancreatic Neuroendocrine Tumors (PNETs). Cancer Genet. 2022, 266–267, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Papouchado, B.; Erickson, L.A.; Rohlinger, A.L.; Hobday, T.J.; Erlichman, C.; Ames, M.M.; Lloyd, R.V. Epidermal Growth Factor Receptor and Activated Epidermal Growth Factor Receptor Expression in Gastrointestinal Carcinoids and Pancreatic Endocrine Carcinomas. Mod. Pathol. 2005, 18, 1329–1335. [Google Scholar] [CrossRef] [PubMed]
- Townsend, C.M.; Ishizuka, J.; Thompson, J.C. Studies of Growth Regulation in a Neuroendocrine Cell Line. Acta Oncol. 1993, 32, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.C. Molecular Targeted Therapy for Carcinoid and Islet-Cell Carcinoma. Best Pract. Res. Clin. Endocrinol. Metab. 2007, 21, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Bresciani, G.; Ditsiou, A.; Cilibrasi, C.; Vella, V.; Rea, F.; Schiavon, M.; Cavallesco, N.G.; Giamas, G.; Zatelli, M.C.; Gagliano, T. EGF and IGF1 Affect Sunitinib Activity in BP-NEN: New Putative Targets beyond VEGFR? Endocr. Connect. 2019, 8, 680–690. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Xu, H.; Strosberg, J.R.; Lu, R.; Zhu, X.; Deng, S.; Ding, L.; Ni, Q.; Warshaw, A.L.; Yu, X.; et al. EGFR Is a Potential Therapeutic Target for Highly Glycosylated and Aggressive Pancreatic Neuroendocrine Neoplasms. Int. J. Cancer 2023, 153, 164–172. [Google Scholar] [CrossRef]
- Kogo, M.; Shimizu, R.; Uehara, K.; Takahashi, Y.; Kokubo, M.; Imai, Y.; Tomii, K. Transformation to Large Cell Neuroendocrine Carcinoma as Acquired Resistance Mechanism of EGFR Tyrosine Kinase Inhibitor. Lung Cancer 2015, 90, 364–368. [Google Scholar] [CrossRef] [PubMed]
- Benyoucef, S.; Surinya, K.H.; Hadaschik, D.; Siddle, K. Characterization of Insulin/IGF Hybrid Receptors: Contributions of the Insulin Receptor L2 and Fn1 Domains and the Alternatively Spliced Exon 11 Sequence to Ligand Binding and Receptor Activation. Biochem. J. 2007, 403, 603–613. [Google Scholar] [CrossRef] [PubMed]
- Galal, M.A.; Alouch, S.S.; Alsultan, B.S.; Dahman, H.; Alyabis, N.A.; Alammar, S.A.; Aljada, A. Insulin Receptor Isoforms and Insulin Growth Factor-like Receptors: Implications in Cell Signaling, Carcinogenesis, and Chemoresistance. Int. J. Mol. Sci. 2023, 24, 15006. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Li, K.; Li, W.; Wang, C.; Liu, Y.; Zhang, H.; Pan, J.; Qi, S.; Peng, J. Expression of Insulin-Like Growth Factor Type 1 Receptor Is Linked to Inflammation in Adamantinomatous Craniopharyngioma. Neuroendocrinology 2022, 112, 917–926. [Google Scholar] [CrossRef] [PubMed]
- Ludovini, V.; Bellezza, G.; Pistola, L.; Bianconi, F.; Di Carlo, L.; Sidoni, A.; Semeraro, A.; Del Sordo, R.; Tofanetti, F.R.; Mameli, M.G.; et al. High Coexpression of Both Insulin-like Growth Factor Receptor-1 (IGFR-1) and Epidermal Growth Factor Receptor (EGFR) Is Associated with Shorter Disease-Free Survival in Resected Non-Small-Cell Lung Cancer Patients. Ann. Oncol. 2009, 20, 842–849. [Google Scholar] [CrossRef]
- Wulbrand, U.; Remmert, G.; Zöfel, P.; Wied, M.; Arnold, R.; Fehmann, H.C. mRNA Expression Patterns of Insulin-like Growth Factor System Components in Human Neuroendocrine Tumours. Eur. J. Clin. Investig. 2000, 30, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Van Adrichem, R.C.S.; De Herder, W.W.; Kamp, K.; Brugts, M.P.; De Krijger, R.R.; Sprij-Mooij, D.M.; Lamberts, S.W.J.; Van Koetsveld, P.M.; Janssen, J.A.M.J.L.; Hofland, L.J. Effects of Somatostatin Analogs and Dopamine Agonists on Insulin-Like Growth Factor 2-Induced Insulin Receptor Isoform A Activation by Gastroenteropancreatic Neuroendocrine Tumor Cells. Neuroendocrinology 2016, 103, 815–825. [Google Scholar] [CrossRef] [PubMed]
- Afargan, M. Novel Long-Acting Somatostatin Analog with Endocrine Selectivity: Potent Suppression of Growth Hormone But Not of Insulin. Endocrinology 2001, 142, 447–486. [Google Scholar] [CrossRef] [PubMed]
- Gentilin, E.; Di Pasquale, C.; Rossi, M.; Tagliati, F.; Gagliano, T.; Rossi, R.; Pelizzo, M.; Merante Boschin, I.; Degli Uberti, E.C.; Zatelli, M.C. IGF-I Influences Everolimus Activity in Medullary Thyroid Carcinoma. Front. Endocrinol. 2015, 6, 63. [Google Scholar] [CrossRef] [PubMed]
- Chitnis, M.M.; Yuen, J.S.P.; Protheroe, A.S.; Pollak, M.; Macaulay, V.M. The Type 1 Insulin-Like Growth Factor Receptor Pathway. Clin. Cancer Res. 2008, 14, 6364–6370. [Google Scholar] [CrossRef] [PubMed]
- Henfling, M.E.R.; Perren, A.A.; Schmitt, A.M.; Saddig, C.M.; Starke, A.A.; Riedl, R.G.; Versleijen-Jonkers, Y.M.H.; Sprij-Mooij, D.M.; Ramaekers, F.C.S.; Hofland, L.J.; et al. The IGF Pathway Is Activated in Insulinomas but Downregulated in Metastatic Disease. Endocr. Relat. Cancer 2018, 25, 1005–1018. [Google Scholar] [CrossRef]
- Borges, M.; Linnoila, R.I.; Van De Velde, H.J.K.; Chen, H.; Nelkin, B.D.; Mabry, M.; Baylin, S.B.; Ball, D.W. An Achaete-Scute Homologue Essential for Neuroendocrine Differentiation in the Lung. Nature 1997, 386, 852–855. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-D.; Hu, R.; Ding, Q.; Savage, T.K.; Huffman, K.E.; Williams, N.; Cobb, M.H.; Minna, J.D.; Johnson, J.E.; Yu, Y. Subtype-Specific Secretomic Characterization of Pulmonary Neuroendocrine Tumor Cells. Nat. Commun. 2019, 10, 3201. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.A.; Nilsson, M.B.; Le, X.; Cascone, T.; Jain, R.K.; Heymach, J.V. Molecular Mechanisms and Future Implications of VEGF/VEGFR in Cancer Therapy. Clin. Cancer Res. 2023, 29, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Simon, T.; Gagliano, T.; Giamas, G. Direct Effects of Anti-Angiogenic Therapies on Tumor Cells: VEGF Signaling. Trends Mol. Med. 2017, 23, 282–292. [Google Scholar] [CrossRef] [PubMed]
- Bresciani, G.; Hofland, L.J.; Dogan, F.; Giamas, G.; Gagliano, T.; Zatelli, M.C. Evaluation of Spheroid 3D Culture Methods to Study a Pancreatic Neuroendocrine Neoplasm Cell Line. Front. Endocrinol. 2019, 10, 682. [Google Scholar] [CrossRef] [PubMed]
- Mairinger, F.D.; Walter, R.F.H.; Werner, R.; Christoph, D.C.; Ting, S.; Vollbrecht, C.; Zarogoulidis, K.; Huang, H.; Li, Q.; Schmid, K.W.; et al. Activation of Angiogenesis Differs Strongly Between Pulmonary Carcinoids and Neuroendocrine Carinomas and Is Crucial for Carcinoid Tumourgenesis. J. Cancer 2014, 5, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Dicitore, A.; Gaudenzi, G.; Carra, S.; Cantone, M.C.; Oldani, M.; Saronni, D.; Borghi, M.O.; Grotteschi, J.; Persani, L.; Vitale, G. Antitumor Activity of Axitinib in Lung Carcinoids: A Preclinical Study. Cancers 2023, 15, 5375. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.-M.; Chu, P.-Y.; Lin, H.-Y.; Huang, K.-W.; Hung, W.-C.; Shan, Y.-S.; Chen, L.-T.; Tsai, H.-J. PTEN Regulates Invasiveness in Pancreatic Neuroendocrine Tumors through DUSP19-Mediated VEGFR3 Dephosphorylation. J. Biomed. Sci. 2022, 29, 92. [Google Scholar] [CrossRef] [PubMed]
- Vaahtomeri, K.; Karaman, S.; Mäkinen, T.; Alitalo, K. Lymphangiogenesis Guidance by Paracrine and Pericellular Factors. Genes Dev. 2017, 31, 1615–1634. [Google Scholar] [CrossRef] [PubMed]
- Varney, M.L.; Singh, R.K. VEGF-C-VEGFR3/Flt4 Axis Regulates Mammary Tumor Growth and Metastasis in an Autocrine Manner. Am. J. Cancer Res. 2015, 5, 616–628. [Google Scholar] [PubMed]
- Rudin, C.M.; Drilon, A.; Poirier, J.T. RET Mutations in Neuroendocrine Tumors: Including Small-Cell Lung Cancer. J. Thorac. Oncol. 2014, 9, 1240–1242. [Google Scholar] [CrossRef] [PubMed]
- Salvatore, D.; Santoro, M.; Schlumberger, M. The Importance of the RET Gene in Thyroid Cancer and Therapeutic Implications. Nat. Rev. Endocrinol. 2021, 17, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Tiedje, V.; Ting, S.; Herold, T.; Synoracki, S.; Latteyer, S.; Moeller, L.C.; Zwanziger, D.; Stuschke, M.; Fuehrer, D.; Schmid, K.W. NGS Based Identification of Mutational Hotspots for Targeted Therapy in Anaplastic Thyroid Carcinoma. Oncotarget 2017, 8, 42613–42620. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Carbonero, R.; Anton-Pascual, B.; Modrego, A.; Del Carmen Riesco-Martinez, M.; Lens-Pardo, A.; Carretero-Puche, C.; Rubio-Cuesta, B.; Soldevilla, B. Advances in the Treatment of Gastroenteropancreatic Neuroendocrine Carcinomas: Are We Moving Forward? Endocr. Rev. 2023, 44, 724–736. [Google Scholar] [CrossRef] [PubMed]
- Addeo, A.; Miranda-Morales, E.; Den Hollander, P.; Friedlaender, A.; Sintim, H.O.; Wu, J.; Mani, S.A.; Subbiah, V. RET Aberrant Cancers and RET Inhibitor Therapies: Current State-of-the-Art and Future Perspectives. Pharmacol. Ther. 2023, 242, 108344. [Google Scholar] [CrossRef] [PubMed]
- VanDeusen, H.R.; Ramroop, J.R.; Morel, K.L.; Bae, S.Y.; Sheahan, A.V.; Sychev, Z.; Lau, N.A.; Cheng, L.C.; Tan, V.M.; Li, Z.; et al. Targeting RET Kinase in Neuroendocrine Prostate Cancer. Mol. Cancer Res. 2020, 18, 1176–1188. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.Y.; Bergom, H.E.; Day, A.; Greene, J.T.; Sychev, Z.E.; Larson, G.; Corey, E.; Plymate, S.R.; Freedman, T.S.; Hwang, J.H.; et al. ZBTB7A as a Novel Vulnerability in Neuroendocrine Prostate Cancer. Front. Endocrinol. 2023, 14, 1093332. [Google Scholar] [CrossRef] [PubMed]
- Kander, E.M.; Shah, M.H.; Zhou, Y.; Goyal, A.; Palmer, J.D.; Owen, D.H.; Shilo, K.; Patel, G.; Raval, R.R.; Gonzalez, J.; et al. Response to the Selective RET Inhibitor Selpercatinib (LOXO-292) in a Patient With RET Fusion-Positive Atypical Lung Carcinoid. Clin. Lung Cancer 2021, 22, e442–e445. [Google Scholar] [CrossRef] [PubMed]
- Soda, M.; Choi, Y.L.; Enomoto, M.; Takada, S.; Yamashita, Y.; Ishikawa, S.; Fujiwara, S.; Watanabe, H.; Kurashina, K.; Hatanaka, H.; et al. Identification of the Transforming EML4–ALK Fusion Gene in Non-Small-Cell Lung Cancer. Nature 2007, 448, 561–566. [Google Scholar] [CrossRef] [PubMed]
- Morris, S.W.; Kirstein, M.N.; Valentine, M.B.; Dittmer, K.G.; Shapiro, D.N.; Saltman, D.L.; Look, A.T. Fusion of a Kinase Gene, ALK, to a Nucleolar Protein Gene, NPM, in Non-Hodgkin’s Lymphoma. Science 1994, 263, 1281–1284. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Li, J.; Xia, Q.; Liu, K.; Dong, Z. New Perspectives for Targeting Therapy in ALK-Positive Human Cancers. Oncogene 2023, 42, 1959–1969. [Google Scholar] [CrossRef] [PubMed]
- Shaw, A.T.; Ou, S.H.; Bang, Y.J.; Camidge, D.R.; Solomon, B.J.; Salgia, R.; Riely, G.J.; Varella-Garcia, M.; Shapiro, G.I.; Costa, D.B.; et al. Crizotinib in ROS1-Rearranged Non–Small-Cell Lung Cancer. N. Engl. J. Med. 2015, 372, 683–684. [Google Scholar] [CrossRef] [PubMed]
- Leal, J.L.; Peters, G.; Szaumkessel, M.; Leong, T.; Asadi, K.; Rivalland, G.; Do, H.; Senko, C.; Mitchell, P.L.; Quing, C.Z.; et al. NTRK and ALK Rearrangements in Malignant Pleural Mesothelioma, Pulmonary Neuroendocrine Tumours and Non-Small Cell Lung Cancer. Lung Cancer 2020, 146, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, M.; Uchiyama, N.; Shigemasa, R.; Matsumura, T.; Matsuoka, R.; Nomura, A. Atypical Carcinoid Tumor with Anaplastic Lymphoma Kinase (ALK) Rearrangement Successfully Treated by an ALK Inhibitor. Intern. Med. 2016, 55, 3151–3153. [Google Scholar] [CrossRef] [PubMed]
- Ooki, A.; Osumi, H.; Fukuda, K.; Yamaguchi, K. Potent Molecular-Targeted Therapies for Gastro-Entero-Pancreatic Neuroendocrine Carcinoma. Cancer Metastasis Rev. 2023, 42, 1021–1054. [Google Scholar] [CrossRef] [PubMed]
- Aristizabal Prada, E.T.; Heinzle, V.; Knösel, T.; Nölting, S.; Spöttl, G.; Maurer, J.; Spitzweg, C.; Angele, M.; Schmidt, N.; Beuschlein, F.; et al. Tropomyosin Receptor Kinase: A Novel Target in Screened Neuroendocrine Tumors. Endocr. Relat. Cancer 2018, 25, 547–560. [Google Scholar] [CrossRef] [PubMed]
- Passhak, M.; McNamara, M.G.; Hubner, R.A.; Ben-Aharon, I.; Valle, J.W. Choosing the Best Systemic Treatment Sequence for Control of Tumour Growth in Gastro-Enteropancreatic Neuroendocrine Tumours (GEP-NETs): What Is the Recent Evidence? Best Pract. Res. Clin. Endocrinol. Metab. 2023, 37, 101836. [Google Scholar] [CrossRef] [PubMed]
- Corti, F.; Brizzi, M.P.; Amoroso, V.; Giuffrida, D.; Panzuto, F.; Campana, D.; Prinzi, N.; Milione, M.; Cascella, T.; Spreafico, C.; et al. Assessing the Safety and Activity of Cabozantinib Combined with Lanreotide in Gastroenteropancreatic and Thoracic Neuroendocrine Tumors: Rationale and Protocol of the Phase II LOLA Trial. BMC Cancer 2023, 23, 908. [Google Scholar] [CrossRef] [PubMed]
- Vitale, G.; Cozzolino, A.; Malandrino, P.; Minotta, R.; Puliani, G.; Saronni, D.; Faggiano, A.; Colao, A. Role of FGF System in Neuroendocrine Neoplasms: Potential Therapeutic Applications. Front. Endocrinol. 2021, 12, 665631. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toffoli, L.; Ditsiou, A.; Gagliano, T. Exploring Emerging Therapeutic Targets and Opportunities in Neuroendocrine Tumors: Updates on Receptor Tyrosine Kinases. Receptors 2024, 3, 145-154. https://doi.org/10.3390/receptors3020008
Toffoli L, Ditsiou A, Gagliano T. Exploring Emerging Therapeutic Targets and Opportunities in Neuroendocrine Tumors: Updates on Receptor Tyrosine Kinases. Receptors. 2024; 3(2):145-154. https://doi.org/10.3390/receptors3020008
Chicago/Turabian StyleToffoli, Lara, Angeliki Ditsiou, and Teresa Gagliano. 2024. "Exploring Emerging Therapeutic Targets and Opportunities in Neuroendocrine Tumors: Updates on Receptor Tyrosine Kinases" Receptors 3, no. 2: 145-154. https://doi.org/10.3390/receptors3020008