Abstract
The Ediacaran–Cambrian deposits of Brittany (Brioverian series) contain both a few isolated pluricentimetric discoid structures, dome-shaped or “donut”-shaped, and a multitude of centimetric to infracentimetric more or less elliptical fossils or pseudofossils. The discoid and elliptical Brioverian structures are compared to similar fossils and pseudofossils found worldwide, and interpreted considering both sedimentary and biological hypotheses. This synthesis of more or less enigmatical fossils from the Ediacaran–Fortunian deposits of Brittany completes the previous descriptions of more well-known discoid and elliptical Brioverian structures such as Nimbia-like and Chuaria-like fossils. It provides a better understanding of the diversity of the Brioverian fossils and original sedimentary structures.
1. Introduction
Discoid and elliptical fossils from Ediacaran–Cambrian deposits have been found worldwide [1]. Among these fossils, the smallest and more abundant, ranging in size from millimeters to centimeters, have almost always been referred to the same taxa, Beltanelliformis Menner 1974, Beltanelloides Sokolov 1965, or Nemiana Palij 1976, the two last genera being generally considered as synonymous to the first one [2]. But their biological and taxonomical interpretations have sometimes changed, from algae to cyanobacteria [3]. Larger discoid fossils are also known in numerous Ediacaran–Cambrian deposits, and were assigned to various genera such as Aspidella Bilings 1872, Cyclomedusa Sprigg 1947, Heliomedusa Sun and Hou 1987, or Medusinites Sprigg 1949, previously termed “medusoids” and interpreted as coelenterate-grade organisms [4,5,6,7]. However, Grazhdankin and Gerdes [8] have considered that enigmatic discoidal fossils from the Ediacaran White Sea deposits were in fact microbial mats. Banerjee et al. [9] have also considered that circular fossils similar to Cyclomedusa [10] from Ediacaran deposits in India can be explained as mat fragment impressions. Finally, Young and Hagadorn [11] have concluded that medusan preservation is maximal for Cambrian deposits and then decreases through time, but is not attested for Neoproterozoic strata. In 2007, Grazhdankin and Gerdes [8] indicated the presence of filamentous structures and pyritization in “medusoid” fossils from the White Sea area. Although they described the preservation of microbial colonies, they could not exclude that, in some cases, these microbial colonies developed on medusae corpses. Finally, the existence of fossil medusae in Ediacaran deposits remains possible, but needs to be clearly demonstrated.
In northwestern France, the Neoproterozoic strata have never been completely or clearly described and dated, and the precise location of the Ediacaran/Cambrian boundary is still unknown. Deposits covering the end of the Proterozoic and the base of the Paleozoic are regrouped in the Brioverian facies, which is widespread in Brittany and south-west Normandy [12] (Figure 1). The lower part of the Brioverian deposits is mainly Ediacaran in age, even Cryogenian at the base, according to previous stratigraphical and palynological works [13,14,15,16,17,18]. In Brittany, the Cryogenian deposits are restricted to the northern coast, near Saint-Brieuc, while the Late Ediacaran/Early Cambrian series constitute the upper Brioverian strata of Central Brittany, especially between Néant-sur-Yvel and Nouvoitou, near Rennes [12,19] (Figure 1). The upper Brioverian biostratigraphy is poorly known, although, in recent years, radiochronological dating and paleontological studies have been carried out to place the fossils in a timeframe and to discuss the stratigraphic position of the different facies [19,20,21,22,23,24,25,26,27]. The upper Brioverian paleontological content, previously documented by Lebesconte [28] and recently revised [20,21,22,23,25], is mainly composed of thin ichnofossils (e.g., Helminthoidichnites, Helmintopsis) [29] corresponding to grazing trails of vermiform organisms made on microbial mats in shallow environments. The early Cambrian ichnofossil index, Treptichnus pedum, has never been observed in spite of the richness of the upper Brioverian deposits in ichnofossils. However, radiochrological dating does not allow confirmation whether the upper Brioverian deposits are solely Ediacaran in age or if they include the lower Fortunian (Early Cambrian) strata [21,26].
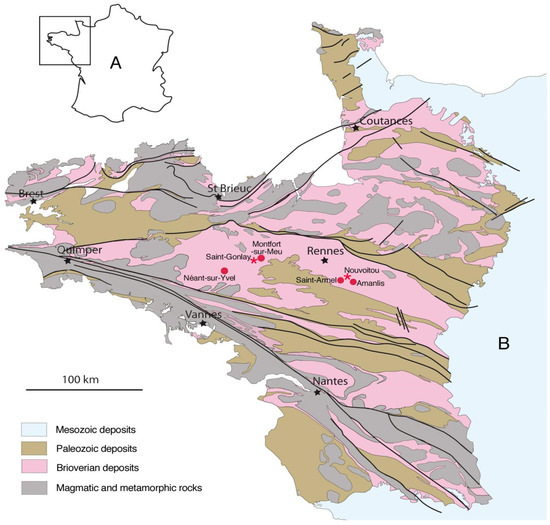
Figure 1.
Location of the Late Brioverian outcrops (Ediacaran–Fortunian) from Brittany. (A) Location of the Armorican Massif in NW France; (B) geological map of the Armorican Massif with location (red spots) of the main Brioverian localities in northwestern France mentioned in the text: Montfort-sur-Meu, locality where the first Brioverian ichnofossils were discovered by Lebesconte [30]; Néant-sur-Yvel, locality where the first Brioverian microbial mats were discovered by Lebesconte [30]; Saint-Armel and Amanlis, localities where Lebesconte [30] discovered the enigmatical fossils Armelia and Amanlisia; Nouvoitou and Saint-Gonlay (red stars), localities where the fossils described in the present paper have been collected; the black stars indicate the main big cities from northwestern France (from Néraudeau et al. [26]).
Following previous discoveries of ichnofossils, the Ediacaran–Cambrian deposits in Brittany were searched for body fossils. Two kinds of small elliptical fossils have been recently described [24,26]: centimetric elliptical structures, with a slight relief, related to the genus Nimbia Fedonkin 1980 [26] and small flattened glossy carbonaceous structures, 5 to 8 mm in size, identified as Chuaria Walcott 1899 [24]. During the last five years, two categories of new discoid or elliptical structures have been found. The first type is represented by clusters of hundreds of more or less elliptical centimetric to infracentimetric forms. The second type corresponds to a few isolated pluricentimetric dome-shaped or “donut”-shaped discoid structures. The first objective of the present work is to describe these new elliptical or discoid structures from the Brioverian of the Armorican Massif. Secondly, various hypotheses are examined to explain the origins of the Breton structures and to assign them, when possible, to fossils or pseudofossils.
2. Geological Setting
Geological context—The elliptical and discoid structures (fossils or pseudofossils) presented in this study were sampled by the authors from 2018 to 2023 in two localities of Brittany (NW France), Saint-Gonlay and Nouvoitou (Figure 1).
At Saint-Gonlay, the shale with ichnofossils, body fossils and sedimentary figures were collected on a slag heap located on the edge of a backfilled and vegetated quarry. At Nouvoitou, the fossiliferous siltites and fine sandstones were collected on a pile of rocks dug for the foundations of a house. It was therefore not possible to construct a stratigraphical column in the two localities.
The rocks containing the elliptical and discoid structures are grey-green fine sands and siltites (Nouvoitou) to grey-blue shales (Saint-Gonlay). Because of the way the samples were collected (slag heap and rock fragment), it was not possible to determine with certainty the stratigraphic top and bottom (younging direction) of each sample.
Geochronological constraints—Gougeon et al. [21,23] reported geochronological constraints obtained by the U-Pb method on detrital zircon grains from the sandstones interbedded in the fossiliferous Brioverian shale series of Néant-sur-Yvel and Saint-Gonlay (Figure 1). The U-Pb data argue for maximum deposition ages between 551 ± 7 Ma and 540 ± 5 Ma. The radiochronological dating of a sandstone from the Nouvoitou locality is presented in this paper.
Paleontological context—Ichnofossils are abundant both on shale and sandstone surfaces [21,25,26]. The main part of the ichnofossils have a constant diameter between 1 to 2 mm and are horizontal, elongated with straight to meandering trails. These trace fossils were historically described by Lebesconte (1886) within the ichnogenera Montfortia Lebesconte 1886, in reference to the locality Montfort-sur-Meu (Figure 1). Gougeon et al. [21] revised the ichnogenera and have demonstrated that, according to the morphology of these ichnofossils, they are junior synonyms of Helminthoidichnites Fitch 1850 (millimetric and sub-straight), Helminthopsis Heer 1877 (millimetric and meandering), and Gordia Emmons 1844 (millimetric and intertwined) [31]. These thin ichnofossils are interpreted as trails of locomotion or nutrition (grazing trace) made by vermiform organisms (e.g., polychaetes, priapulids, nematods) [32,33]. In addition, larger sub-horizontal burrows with variable diameters of several millimeters, previously named Amanlisia by Lebesconte [30] (referring to the locality Amanlis, Figure 1), were synonymized with Palaeophycus Hall 1847 by Néraudeau [24]. The shale surfaces that bear elliptical or discoid fossils are generally poor in ichnofossils [26]. The absence of typical Cambrian fossils and ichnofossils and the radiochronological dating give a Late Ediacaran maximum deposition age for the fossiliferous beds, although an Early Cambrian age (Fortunian) cannot be disregarded.
At Saint-Gonlay, the shale surfaces that bear elliptical fossils are often rich in Microbially Induced Sedimentary Surfaces (MISS), previously named “Neantia” by Lebesconte [30] with reference to the locality of Néant-sur-Yvel (Figure 1). These MISS are conspicuous when wrinkled ([21]: Figure 7; [25]: Figures 7 and 10), and can be highlighted by thin sections for more smooth shales [21,27].
3. Material and Methods
Radiochronological dating—For the purpose of this study, we selected one sandstone sample from the Nouvoitou deposit in order to perform U-Pb dating on detrital zircon grains (as mentioned earlier, Saint-Gonlay dating was previously published by Gougeon et al. [23]). U-Pb dating was conducted in-situ by laser ablation inductively coupled plasma mass spectrometry (LA-ICP-MS) at the GeOHeLiS analytical platform (Université de Rennes) using an ESI NWR193UC Excimer laser coupled to an Agilent 7700x quadrupole ICP-MS equipped with a dual pumping system to enhance sensitivity [34]. The analytical procedure followed the method described in Nosenzo et al. [35]. The analytical protocol is provided in Supplementary Table S1, while the zircon standard analyses (GJ-1 [36] and Plešovice [37]) are listed in Supplementary Table S2. KDE and Concordia diagrams (Figure 2) were generated using IsoplotR [38].
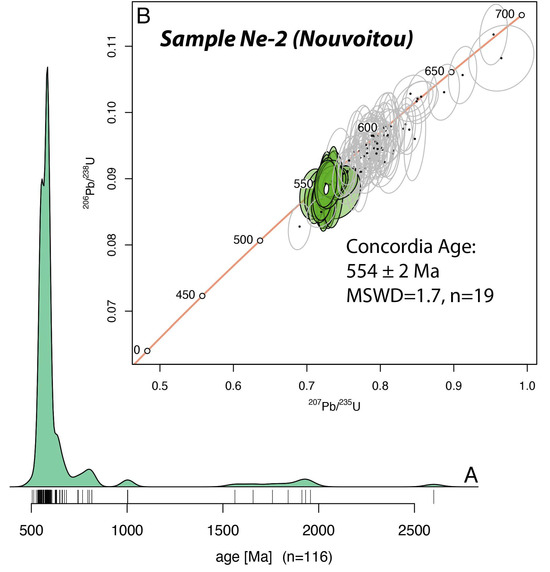
Figure 2.
(A) KDE diagram (Kernel Density Estimation) for the more than 90% concordant data from the U-Pb dating of detrital zircon grains from Nouvoitou. (B) Wetherill concordia diagram for the zircon grains that are younger than 700 Ma. The calculated age of 554 ± 2 Ma is given by the youngest cluster of concordant data.
One hundred and forty-two grains were analyzed (Supplementary Table S2), out of which 116 were 90% concordant or more (see Figure 2A). Most of the analyses yield apparent ages between ca. 800 and 550 Ma, while a few others are Palaeo- to Meso-Proterozoic, between 2000 and 1000 Ma (Figure 2A). The youngest group of nineteen concordant data yield a concordia date of 554 ± 2 Ma (MSWD = 1.7; n = 19) that we interpret as the maximum age of deposition for this sandstone (Figure 2B).
Fossil analysis and storage—The samples were simply washed in water before being measured and photographed. The elliptical and discoid structures were measured with a caliper and photographed with a Panasonic DMC-FZ62 camera. Thirty specimens found on the shale sample as a cloud of elliptical structures were measured (length and width). Measurements were also realized on every isolated large specimen of the other shale samples. Polished sections were realized to observe the potential 3D structures present in the cluster of elliptical structures and to define the relationship between the sediment grain size, the microbial mats and the specimens. However, if only one single specimen was available, or for the isolated discoid structures, thin slides or polished sections were not made in order to preserve them.
The samples are housed at the Geological Institute of the University of Rennes under the numbers IGR-PAL-90201 to IGR-PAL-90208.
4. Results
4.1. Clusters of “Nimbia-like” Structures
The clusters of “Nimbia-like” fossils from Saint-Gonlay: Elliptical centimetric structures with a peripheral bulge, close to the Nimbia morphology, have been recently described for small clusters of specimens from the Late Ediacaran/Fortunian deposits of Brittany [26] and from the Fortunian deposits of Normandy [27]. The discovery, in the Late Ediacaran/Fortunian deposits of Saint-Gonlay, of dense clusters including thousands of centimetric elliptical structures (Figure 3A,B) gives a new insight on these fossils, previously observed in the Armorican Massif (Brittany and Normandy).

Figure 3.
Clusters of Nimbia-like fossils from the Late Brioverian (Ediacaran–Fortunian) of Saint-Gonlay, Brittany, NW France. (A) General view of a shale bearing numerous Nimbia-like fossils (IGR-PAL-90201); (B–D) high magnification of three specimens showing their relatively regular shape and size, from the previous sample; (E) polished section of a second sample (IGR-PAL-90202) showing that the Nimbia-like fossils (pointed at by white arrows) are 3D structures with a pancake shape deposited on fine laminae. Scale bar: 1 cm.
Material: Three shales (IGR-PAL-90201 to IGR-PAL-90203) that bear between 100 and 150 more or less conspicuous specimens (e.g., Figure 3A), separated from each other by 0.5 to 2 cm.
Description: The best specimens show clearly an elliptical and acuminated outline, a peripheral rim and a central depression (Figure 3B–D). The size of the more conspicuous specimens is always about 7–8 mm in length and 4 to 5 mm in width. The peripheral rim is about 1.4 mm in width, and its maximum elevation above the lamina surface is about 0.4–0.5 mm. On the same fossiliferous shale, conspicuous elliptical specimens are associated to more elongated ones, but all are (1) oriented in the same direction, (2) more or less inclined in the sediment with one side more elevated on the sediment surface than the other side, and (3) with a subrounded extremity opposite to a more acuminated extremity, the last one being opposed to the flow of current, and constituting a small “tail” for some elongated specimens (Figure 3B,C). Some specimens are more or less tilted in the sediments. This explains why parts of these structures are more visible at the surface of the sediment, giving an asymmetrical and sometimes crescent-like depression (Figure 3A).
Discussion: These dense clusters of elliptical structures could be interpreted either as sedimentary features (tool mark, flute cast, load cast) or as raindrop impressions or gas bubbles.
But the relatively regular size and elliptical acuminated outline of the fossils, preserved in epirelief, is clearly different from flute casts or tool marks, which are elongated erosion figures. Their horizontal pancake profile covering the sediment laminae does not correspond to load casts either, as they would have deformed the sedimentary substrate (Figure 3E).
Raindrop impressions are generally subcircular, crateriform or concave–convex (e.g., [39,40,41]). For the crateriform ones, raised rims around depressions that are up to 2 cm in diameter are typical of raindrop impressions [42]. The size of the Brioverian specimens is congruent with the previous characteristics, but raindrop impressions frequently coalesce to form linear furrows [43] and frequently show overlap between two or several imprints. None of these last characteristics can be observed on the Brioverian surfaces bearing elliptical structures.
If we consider their organization and the shape of each fossil, the elliptical Brioverian structures are clearly different than the previously published fossil bubbles [44]. Foam bubbles, with several coalescent bubbles in a same area, are totally lacking on the Brioverian fossiliferous surfaces, and the sizes of the elliptical structures are very similar; the fossils are elliptical, acuminated and flat, never hemispherical. Microbial mats are frequent in the fossiliferous layers of Saint-Gonlay, but the polished perpendicular sections of the surfaces bearing clusters of elliptical fossils do not show hemispherical structures such as the ones published by Arrouy et al. ([3]: Figure 3) and Bozak et al. [45] for fossil or modern bubbles preserved by microbial laminae.
These new elliptical fossils from Saint-Gonlay correspond in polished sections to flat “pancakes” regularly spaced and resemble the biological Nimbia-like structures from the Fortunian of Normandy ([27]: Figure 6C–F). But their true nature and origin remain uncertain. Indeed, depending on the authors and the year of publication, Nimbia sensu lato are interpreted as cnidarians [46,47,48,49], microbial mat structures ([8,50], scratch circles formed by the attachment site of organisms [51] or remain uncertain [31,52]. Our elliptical structures are similar to Nimbia-like structures, but their size and shape are more irregular, making it even more difficult to interpret their origin.
4.2. Clusters of “Beltanelliformis-like” Structures
Elliptical flat structures have been discovered in the Brioverian deposits of Nouvoitou (Figure 4A–D). The presence of ichnofossils close to treptichnids in the local sedimentary succession, but not in association to the elliptical structures, could indicate a Fortunian age for part of the deposits. But these treptichnids are not typical of Treptichnus pedum, the index of the Cambrian base [53,54], and even with the radiochronological dating it is difficult to conclude either on a late Ediacaran or early Fortunian (earliest Cambrian) age for the fossiliferous deposits. Either way, the elliptical structures are numerous (several dozens) and present in successive silty laminae of a fine sandstone.

Figure 4.
Clusters of Beltanelliformis from the Late Brioverian (Ediacaran–Fortunian) of Nouvoitou (A–D) and Saint-Gonlay (E,F), Brittany, NW France. (A) Sample with spaced tiny specimens (IGR-PAL-90205); (B) sample very rich in large specimens, overlaid in successive laminae (IGR-PAL-90204); (C) large specimens surrounded by a brownish oxidized halo (bh) (IGR-PAL-90205); (D) zoom on specimens from (B) with arrows indicating the direction of the current according to three specimens deposited in successive laminae: specimen 1 is located in the upper lamina and separated from specimen 2 by a thin layer of sediment (s); specimen 3 is located in a lower lamina separated from specimen 2 by a thin sediment layer (s’); (E) dense cluster of tiny specimens (IGR-PAL-90203); (F) zoom on a part of the cluster of (E). Scale bar: 5 mm.
Material: One piece of sandstone (13 cm in length by 5.5 cm in width) with three dozen large specimens (Figure 4B,D) (IGR-PAL-90204); one piece of sandstone (5 cm in length by 3 cm in width) with a dozen of large specimens (IGR-PAL-90206); a third piece of sandstone with both dozens of small specimens in a part of the laminae (Figure 4A) and five large specimens in another laminae (8 cm in length by 6.5 cm in width) (Figure 4C) (IGR-PAL-90205). The previous measurements of sandstone pieces with elliptical structures give an idea of the density of the elliptical structures on each sample, the different pieces being collected ex situ in a pile of dug rock.
Description: The structures are flat and have an elliptical outline, with a regular size in the same cluster, about 3–3.5 mm in length by 2–2.5 mm in width for the biggest ones (Figure 4B–D), about 1 mm in length by 0.7 mm for small ones (Figure 4A), and are made of a black oxidized material (iron and manganese). The large specimens have almost the same orientation in the fossiliferous laminae (Figure 4B–D). On one piece of sandstone, the large specimens are surrounded by a brown halo of oxides (Figure 4C).
Discussion: The specimens of Nouvoitou, which are very flat, regular in size and shape, with a conspicuous outline and a dark color, cannot be interpreted as sedimentary structures made by the sinking or the groove of natural objects (e.g., load casts or tool marks). Also, because of their regularity and peculiar shapes, they cannot be interpreted as microbial mat spots or as simple traces of oxidation. They differ from the Nimbia-like fossils of the Ediacaran–Fortunian deposits of the Armorican Massif [25,27] (see previous Section 4.1) and various other localities (e.g., [55]) by lacking a peripheral rim. They are similar in shape (elliptical, flattened with local overlapping), size and color to the specimens illustrated by Brasier et al. [56], Vidal et al. ([57]: Figure 16b), McCall ([58]: Figure 110B,C) or Jensen et al. ([59]: Figure 3a). But these last specimens are carbonaceous discs, corresponding more likely to Chuaria, whereas our specimens are ferruginous. Moreover, our specimens do not have the glossy surface and the conspicuous concentric ridges of other Brioverian fossils from Brittany assigned to Chuaria algae [24]. Finally, they are more similar in size and shape to the “Beltanelloides sorichevae” illustrated by Ivantsvov et al. ([2]: Figure 3) and synonymized by the same authors with Beltanelliformis brunsae (or B. minutae). Indeed, these authors indicate that the lower size known from Beltanelliformis is about a few millimeters, like our specimens. Moreover, our specimens are flattened and elliptical such as the specimens published by Ivantsov et al. ([2]: Figures 3, 6–8) or Saint Martin and Saint Martin [60], and not dome-shaped (convex) and contiguous like the typical Beltanelliformis published by McCall ([58]: Figure 89) or Ivantsov ([61]: Figure 2a). Ivantsvov et al. [2] mention that the “accumulation of these fossils to thin interbeds inside more or less homogeneous strata and accumulation on one or closely positioned bedding planes suggest that remains were transported and accumulated rather than dropped from plankton. It is plausible that multilayer burials of Beltanelloides were formed as a result of transportation and accumulation”. Therefore, we interpret the elliptical structures from Nouvoitou as “Beltanelliformis-like” fossils.
4.3. Clusters of Micro-Elliptical Structures
Inframillimetric elliptical structures have been discovered in the Brioverian deposits of Saint-Gonlay, near Rennes, in a piece of greenish shale (Figure 4E,F). These structures are preserved in a piece of shale devoid of ichnofossils. However, in the same locality, the shales generally show a great abundance of ichnofossils (Helminthoidichnites, Helminthopsis, Palaeophycus; see Section 4.1 above), but without Treptichnus pedum, the index of the Cambrian base, it is difficult to conclude either on a late Ediacaran or early Fortunian (earliest Cambrian) age for the fossiliferous deposits. Either way, the tiny elliptical imprints are very numerous and present in successive silty laminae of fine sediment (Figure 4E,F).
Material: A single piece of shale (14 cm in length by 12.5 cm in width) with more than one thousand specimens.
Description: The fossils are preserved on the upper surfaces of successive laminae. They are flat and have a slightly elliptical outline, with a regular size in the same cluster part, about 0.7 mm in length by 0.5 mm in width for the biggest ones (Figure 4E right part and Figure 4F), about 0.4 mm in length by 0.3 mm for small ones (Figure 4E, left part), and are made of dark material. The orientation of the specimens from the same lamina is rather variable, according to their main axis of elongation.
Discussion: These tiny specimens are close in color, elliptical outline, flat shape and presence in successive laminae to the “Beltanelliformis-like” fossils found at Nouvoitou (see Section 4.2 above). But they are much smaller, more rounded, and lack any unidirectional orientation. In their synthesis about Beltanelliformis-like fossils, Ivantsov et al. ([2]: Table 1) give the size range known and previously published for this kind of rounded to elliptical fossil: from 2 mm [62] to 44 mm [63], and varying from circular to elliptical in outline. Consequently, the inframillimetric elliptical structures from Saint-Gonlay are clearly too small to correspond to the classical “Beltanelliformis-like” fossils. Their tiny size, very thin thickness and dense accumulation in successive laminae differentiate them from subcircular sedimentary features (e.g., gas bubbles or load casts) or rounded ichnological structures (e.g., scratch circles or burrow openings). If they are body fossils, their accumulation as thin interbeds may correspond to multilayer burials, formed as a result of transportation and accumulation. We can consider the possibility that it is an accumulation of unicellular algae. But a hypothesis of MISS spots can be proposed as well, with colonies of cyanobacteria developed on successive sedimentary layers. Finally, their rarity (a single piece of shale) and their original shape and accumulation encourage us to interpret them as possible fossils of biological (organic) origins.
4.4. The Isolated Pluricentimetric Discoid or Elliptical Structures
Apart from the previous specimens close to the centimetric (or smaller) Beltanelliformis, Chuaria and Nimbia commented in the previous chapters, the Brioverian deposits of Brittany contain a few large isolated elliptical fossils from 2 to 4 cm in size. Three main shapes have been observed in the Brioverian shales of Saint-Gonlay: “elliptical concentric imprint”, “circular donut shape” and “concentric dome”.
4.4.1. Elliptical Concentric Imprint
Material: A single piece of shale (31 cm in length by 14 cm in width) with a conspicuous concentric elliptical structure and an inconspicuous similar imprint associated to six smooth pit shape structures preserved in hyporelief according to geometry of the dawnlap laminae (Figure 5 and Figure 6F).

Figure 5.
Elliptical concentric imprints from the Late Brioverian (Ediacaran–Fortunian) of Saint-Gonlay, Brittany, NW France. (A) General view of the fossiliferous shale (IGR-PAL-90206), with six small smooth cup-shaped structures (numbers 1 to 6), an inconspicuous concentric imprint (7) and a single conspicuous concentrical structure (8); the white arrow indicates both the direction of the major axis of elongation of the elliptical structures and the direction of enlargement of the peripheral ring of the conspicuous concentrical specimen; the double black arrow (S) indicates where the section illustrated on Figure 6F was made; (B) high magnification of the inconspicuous concentrical imprint (7 on (A)); (C) high magnification of the smooth cup-shaped structure (6 on (A)); the white arrow indicates the outer outline of the structure; (D) high magnification of the conspicuous concentrical structure (8 on (A)); the imprint shows a peripheral ring (p.r.) with an enlargement (e.p.r.) and a central pit (c.p.). Scale bar: (A) = 1 cm; (B–D) = 5 mm.
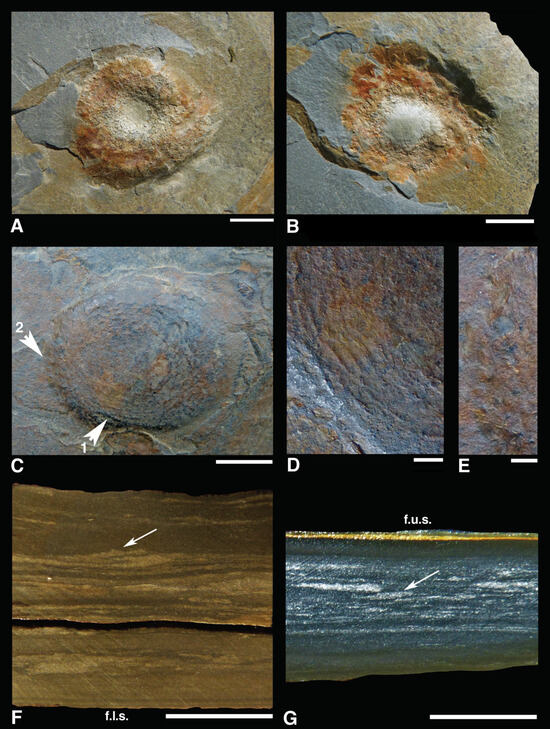
Figure 6.
Discoid or elliptical isolated specimens from the Late Brioverian (Ediacaran–Fortunian) of Saint-Gonlay, Brittany, NW France. (A,B) “Donut shaped” specimen corresponding possibly to a Astropolithon-like sedimentary structure, supposed to be a sand volcano (IGR-PAL-90207); (A) upper face; (B) lower face; (C–E) “dome-shaped” specimen of unknown origin (biological ?, sedimentary ?) (IGR-PAL-90208); (C) general view; (D) side with concentrical ribs (arrow 1 on (C)); (E) slightly pustular side (arrow 2 on (C)); (F) section of the shale sample bearing the structures illustrated on Figure 5; the geometry of the downlap of the laminae (white arrow) indicates that these structures are preserved on the lower surface of the shale (f.l.s. = fossiliferous lower surface); (G) section of the shale bearing the specimen illustrated on pictures (C–E); the geometry of the downlap of the laminae (white arrow) indicates that these structures are preserved on the upper surface of the shale (f.l.s. = fossiliferous lower surface). Scale bar: 1 cm.
Description: The piece of shale bears eight more or less deep elliptical cup-shaped pits, preserved on the upper surface of the bed. The six smaller ones (1 to 6 on Figure 5A,C), of centimetric diameter, are devoid of concentric or radial ornamentation and very similar in size and shape. Another specimen shows a rather conspicuous outer elliptical outline around inconspicuous concentric lines (7 on Figure 5A,B), the central pit being partly filled by the covering sediment. The last specimen is slightly larger, with a large peripheral ring around the central pit (8 on Figure 5A,D), its central pit being also filled by a part of the covering sediment. The maximum diameter of the outer outline (Figure 5D) is about 21 mm when its width is 16 mm. The inner ellipse’s (Figure 5D) dimensions are about 13 mm and 9 mm in length and width, respectively. The specimen shows a conspicuous elliptical outline, with an enlargement of the peripheral ring on one side (5/6 mm on the left part, 4 mm on the right part), as if it had suffered a slip or a tilt. The central pit is partially filled by the sediment of the overlying lamina. The eight specimens show a main axis of elongation in the same direction, as if they were oriented by a current, and this direction is the same for the enlargement axis of the peripheral ring of the more conspicuous concentric specimen (Figure 5A).
Discussion: The six smooth pits are difficult to interpret. But their regular size and shape indicate that they correspond to the same kind of structure, biological or sedimentological. It is impossible to conclude if there were six different specimens or if some specimens correspond only to ricochets of a part of them (for example the specimens 1, 2 and 3 on Figure 5A). These smooth pits could be the imprints of specimens from the underlying deposit, imbedded between two laminae and forming bumps on the sediment surfaces (hyporelief), such as the ones published by Néraudeau et al. ([26]: Figure 4C) for the same outcrop of Saint-Gonlay.
The large concentric specimen (8 on Figure 5A,D) has a more complex structure, and its elliptical peripheral ring, enlarged in an extremity with a sloped position in the sediment and its central pit, recall features of Nimbia-like fossils, such as specimens previously observed in the same Brioverian locality ([26]: Figure 4A). However, the new specimen is twice as big as the previous Nimbia-like specimens found at Saint-Gonlay. Finally, the cup-shape of the central pit filled by the underlying sediment (also visible on specimen 7 in Figure 5B) indicates that the whole structure was probably a bump surface on the epirelief of the underlying layer, as mentioned above for the six associated smooth pits. These characteristics differentiate the concentric specimen(s) from Nimbia-like fossils and correspond more likely to a 3D structure that cannot be interpreted as a simple microbial mat disc or a sedimentary discoid imprint. For the same reason, it cannot be a scratch circle, a structure realized by a tethered organism rotated by currents, with the upper parts of the organism dragged on the substrate surface around the attachment point, leaving circular marks on the sediment [51,64,65]. The similarities in size and shape between the six smooth pits on the one hand, and between the two concentric specimens on the other hand (Figure 5), exclude the hypothesis of load cast imprints. Therefore, they could maybe correspond to body fossils imprints, in what is often called “medusoid” fossils [6]. However, Grazhdankin and Gerdes [8] have considered that many enigmatic discoidal fossils from the Neoproterozoic of the White Sea, such as specimens related to Cyclomedusa or Paliella, are in fact microbial mats. Young and Hagadorn [66] and Van Iten et al. [18] concluded that most Ediacaran “medusoids” represent, in fact, non-medusan fossils, and should rather be interpreted as ichnofossils, inorganic features such as fluid escape structures, microbial decomposition structures or even scratch circles. Also, Young and Hagadorn [11] added that cnidarian medusae preservation is attested for Cambrian strata but not for Neoproterozoic ones. In conclusion, we exclude the possibility that these elliptical concentric imprints correspond to sedimentary structures, microbial mats or scratch circles, but we cannot conclude their biological origin.
4.4.2. “Donut Shape” Specimen
Material: A single piece of shale (16 cm in length by 12.5 cm in width) with a single specimen preserved in epirelief (Figure 6C,D) according to geometry of the dawnlap laminae (Figure 6G).
Description: The only known specimen is about 25 mm in average diameter and about 8.5 mm thick/high (Figure 6A,B). The diameter of the central pit is about 3 to 5 mm, according to its deepness. The structure is in “donut-shape” relief on the bed surface (Figure 6A), and its imprint constitutes a ring bowl on the underlying bed (Figure 6B). Observation with a binocular magnifier allows one to observe that the volume of the fossil is mainly composed of infra-millimeter quartz grains and tiny pyrite crystals, while the surrounding sediment is a siltstone. The central pit is full of fine clayey sediment, mainly preserved on the counter impression (Figure 6B). The lower face of the structure can be partly observed on the other side of the shale and is rounded.
Discussion: A first comparison can be made between the Brioverian specimen and the one published by Arrouy et al. ([67]: Figure 2c) from Ediacaran deposits of South America. Their specimen has the same size range (about 35 mm) and the same donut shape, with a central rounded pit, but it is associated with several other specimens with a more or less hard ornamentation of radial ribs that are lacking on the Brioverian specimen. These both radial and concentric features encourage the authors to place their fossils in the genus form Aspidella [68,69], generally used for disc-like Ediacaran fossils, but it is impossible to propose the same interpretation for our specimen, lacking radial ribs and concentric rings.
The Brioverian specimen is very similar in size and shape to the one published by Hagadorn and Miller ([70]: Figure 2b,d). Like the structures described by Hagadorn and Miller [70], it measures about 30 mm in diameter and is hemisphere-shaped, with a clayey central sediment plug, but without outward-radiating arrays of markings. However, conversely to the second specimen described by these authors ([70]: Figure 2d), the specimen from Brittany lacks conspicuous concentric rings.
Our specimen has also similarities with the specimen published by Farmer et al. ([71]: Figure 6e) and is considered by the author as a (pseudo)fossil resembling Medusinites or Bergaueria. It has almost the same size (about 3 cm versus 2 cm) and a large peripheral bulge around a central depression. However, the specimen of Farmer et al. [71] has a protuberance in the center of the depression, which is lacking on our specimen.
Hagadorn and Miller [70] have considered that most of the structures containing a central sediment plug, radial lineation, concentric rings and a broad trough surrounding or underlying ring margins are not scyphozoan medusae or other circular organisms but sedimentary structures named Astropolithon. They correspond to sand-volcano-like structures linked to upward movement of gases or other fluids from underlying beds. Thus, the Brioverian “donut” structure could be a small sand-volcano such as Astropolithon, according to its general shape and sediment organization. However, deposits bearing Astropolithon generally exhibit several specimens [70,72,73] whereas the Brioverian deposit is a single case observed among thousands of shales extracted from the outcrop. Thus, it is difficult to exclude a biological origin. The presence of pyrite crystals in the fossil, a mineral very uncommon in the Brioverian series and fossils, can indicate either the initial presence of organic matter (biological tissues?) or the circulation of a fluid or a gas rich in iron sulfide (sedimentary phenomenon?).
The similarity of these probable pseudofossils to circular body fossils demonstrates the difficulty in ascribing a biotic origin and taxonomic designation to many circular/elliptical structures in sedimentary units, and the Brioverian fossil remains enigmatical.
4.4.3. “Concentric Dome” Specimen
Material: A single piece of shale (21 cm in length by 19 cm in width) with a single specimen preserved on epirelief (Figure 6G).
Description: The specimen is slightly oval and measures about 37 mm in maximum diameter by about 29 mm in width (Figure 6C). It has the same composition and the same fine (silt) granulometry of the surrounding sediment. In raking light, six to seven irregular and discontinuous fine concentric ribs (Figure 6D) can be distinguished on the longest side, an a more pustular surface on the shortest side (Figure 6E).
Discussion: Like some structures described by Hagadorn and Miller [70], it measures about 30 mm in diameter and has a hemisphere-shape and concentric rings. However, it is different because it lacks a clayey central sediment plug and outward-radiating arrays of markings. The specimen looks like the one published by Farmer et al. ([71]: Figure 4C) under the name “Beltanella sp.”, an elliptical and convex structure, but our material is a third smaller and has irregular concentric ribs.
Rounded subconical structures with an elliptical outline and concentrical lines have been often published as “Scenella-like” fossils [74]. They are generally known as being 2 to 20 mm in maximum size [74,75], such as Scenella tenuistriata Chapman 1911, but larger similar structures, defined as “Scenella-like” fossils, have also been published, raised about 30 mm or more, like our specimen [76]. However, typical Scenella, and similar fossils such as Marocella, are characterized by a blunt subcentral or eccentric prominent apex that is lacking on our specimen [74,75,77,78]. A basal Cambrian fossil published by Narbonne et al. [79], previously interpreted as a medusoid hydrozoan and then reviewed as a helcionacean mollusk [28], has been linked to Scenella and has some similarities with our specimen (in size, outline and ornamentation) but is much more flattened, as in Narbonne et al. ([79]: Figure 2.1). Moreover, these different elliptical fossils have conspicuous and continuous concentric ridges when our specimen shows only discontinuous and weak concentric lines.
The dome shape and the concentric ornamentation of our specimen may look like a microbial or an algal build up, but its concentric ribs are very weak and are clearly not linked to a laminated organization, typical of stromatolitic/oncolitic structures. Secondly, microbial and algal build-ups are generally numerous in the same place (e.g., [80]), whereas our specimen is a single example observed during 10 years of excavation and among thousands of “fossiliferous” shale fragments. Finally, the carbonates, often associated with stromatolitic structures, are lacking in the Brioverian deposits providing the fossils and pseudofossils of Saint-Gonlay. Sand bulges with one or several concentric ridges, resembling Ediacaran fossils, are also known in association with discoidal microbial colonies, but never with the regular dome shape characterizing our specimen [8,9].
Without identical fossils in the literature it is difficult to argue for a biological or a sedimentological structure to interpret our dome-shaped Brioverian structures.
5. Paleoenvironmental Discussion
The lithology of the fossiliferous layers providing circular and elliptical fossils consists of variously laminated alternations of silty clay (Saint-Gonlay) and very fine micaceous arenites (Nouvoitou), with small 2D–3D ripples of unidirectional current [26]. These ripples have straight or sinuous ridges, and their height is small (1 to 8 mm). Stratification is typically flaser, and numerous ichnofossils (mainly Helminthoidichnites and Helminthopsis) are observed associated with elliptical fossils and pseudofossils [21,22]. Macroscopic sedimentary figures [81], such as load casts, flute casts and tool casts, are absent in the laminae that share trace fossils or elliptical structures. When elliptical (pseudo)fossils are numerous, they are generally oriented by a unidirectional current in the same direction of ripples (Nimbia-like specimens from Saint-Gonlay and large Beltanelliformis from Nouvoitou), but not always (tiny Beltanelliformis from Saint-Gonlay). The silty argillites of Saint-Gonlay consist of the repetitive overlapping of numerous millimetric laminae of argillite and arenaceous siltstones (Figure 6F,G), while the fine arenites of Nouvoitou contain only very thin argillaceous laminae. At Saint-Gonlay, the respective thickness of silt and clay laminae vary in an increasing–decreasing way, with deposition cycles referable to semidiurnal and semilunar tidal cycles [26]. These sedimentary characteristics correspond to shallow platform environments subject to unidirectional currents and cyclic processes compatible with a tidal flatland environment. The elliptical (pseudo)fossil preservation took place during mud deposition at low energy levels, while more hydrodynamic conditions were responsible for silt and fine sand depositions (Figure 6F,G). Microbial mats are lacking in the fossiliferous fine sandstones and siltites of Nouvoitou when they have been previously highlighted in the shales of Saint-Gonlay, especially for the preservation of small medusoids [26] and grazing traces [25].
6. Conclusions
At the scale of the Brioverian (Ediacaran–Fortunian) deposits of Brittany, elliptical structures are particularly difficult to interpret, because they are both uncommon and non-typical when compared to what has been previously described. Except for the large elliptical structures from Nouvoitou, corresponding to some Beltanelliformis-like fossils illustrated in the literature, it is difficult to opt either for a body fossil interpretation or a sedimentary structure identification for the elliptical fossils of Saint-Gonlay. The clouds of tiny specimens could be considered as an accumulation of unicellular algae or spots of microbial colonies. The two larger concentrical specimens are similar to Nimbia-like specimens previously found in the same outcrop, but their biological origin remains uncertain. The “donut-shape” structure looks like a sand-volcano Astropolithon, but the fact that the structure was limited to a unique specimen is surprising for sand volcanoes, and a biological origin cannot be excluded. The concentric dome fossil is more problematical and cannot be clearly interpreted without the discovery of other specimens. However, these different findings reveal more and more the potential of the Brioverian strata in providing fossils structures to understand the Ediacaran–Cambrian paleoenvironments of the Armorican Massif. The new radiochronological dating obtained for the outcrop of Nouvoitou (554 ± 2 Ma) clearly gives a late Ediacaran maximum age of deposition for the fossiliferous sandstone, and the lack of the Cambrian fossil index Treptichnus pedum, both at Nouvoitou and Saint-Gonlay, reinforce that Ediacaran age.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/fossils2030006/s1, Table S1: Operating conditions for the LA-ICP-MS equipment. Table S2: Operating conditions for the LA-ICP-MS equipment. References [36,37,82,83] are cited in the Supplementary Materials.
Author Contributions
D.N. has collected the fossils on the field, and made both paleontological and sedimentological analyses; M.P. made the radiochronological analyses, A.L. made the sedimentological analyses; J.C. made paleontological analyses. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Materials, further inquiries can be directed to the corresponding authors.
Acknowledgments
The authors are greatly indebted to the owner of the quarry for the access to his property for the present study. They also thank X. Le Coz (University of Rennes) who made the polished sections of the fossiliferous shales, and Y. Lepagnot (University of Rennes) who realized the rock crushing and the separation of minerals for the U/Pb analysis of zircon grains. Finally, the authors thank B. Coutret, R. Gougeon and P. Votat for their contributions to the field work. This publication is a contribution to the VIBRIO and CAMBRARMOR projects of the CNRS-INSU InterrVie program.
Conflicts of Interest
The authors declare that they have no competing interest.
References
- McGabhann, B.A. Discoidal fossils of the Ediacarian biota: A review of current understanding. In The Rise and Fall of the Ediacarian Biota; Vickers-Rich, P., Komarower, P., Eds.; Geological Society of London: London, UK, 2007; Volume 286, pp. 297–313. [Google Scholar]
- Ivantsov, A.Y.; Gritsenko, V.P.; Konstantinenko, L.I.; Zakrevskaya, M.A. Revision of the Problematic Vendian Macrofossil Beltanelliformis (=Beltanelloides, Nemiana). Paleontol. J. 2014, 48, 1415–1440. [Google Scholar] [CrossRef]
- Bobrovskiy, I.; Hope, J.M.; Krasnova, A.; Ivantsov, I.; Brocks, J. Molecular fossils from organically preserved Ediacara biota reveal cyanobacterial origin for Beltanelliformis. Nat. Ecol. Evol. 2018, 2, 437–440. [Google Scholar] [CrossRef] [PubMed]
- Fedonkin, M.A. Systematic description of Vendian Metazoa. In The Vendian System, Volume 1 Paleontology; Sokolov, B.S., Iwanowski, A.B., Eds.; Springer: Berlin, Germany, 1990; pp. 71–120. [Google Scholar]
- Gehling, J.G.; Narbonne, G.M.; Anderson, M.M. The first named Ediacaran body fossil, Aspidella terranovica. Palaeontology 2000, 43, 427–456. [Google Scholar] [CrossRef]
- Sun, W. Precambrian medusoids: The Cyclomedusa plexus and Cyclomedusa-like pseudofossils. Precambrian Res. 1986, 31, 325–360. [Google Scholar]
- Zhang, Z.; Li, G.; Emig, C.; Han, J.; Holmer, L.E.; Shu, D. Architecture and function of the lophophore in the problematic brachiopod Heliomedusa orienta (Early Cambrian, South China). Geobios 2009, 42, 649–661. [Google Scholar] [CrossRef]
- Grazhdankin, D.; Gerdes, G. Ediacaran microbial colonies. Lethaia 2007, 40, 201–210. [Google Scholar] [CrossRef]
- Banerjee, S.; Sarkar, S.; Eriksson, P.G.; Samanta, P. Microbially related structures in siliciclastic sediment resembling Ediacaran fossils: Examples from India, ancient and modern. In Microbial Mats. Modern and Ancient Microorganisms in Stratified Systems; Seckbach, J., Oren, A., Eds.; Springer: Berlin, Germany, 2010; pp. 111–129. [Google Scholar]
- Sprigg, R.C. Early Cambrian (?) jellyfishes from the Flinders Ranges, South Australia. Trans. R. Soc. South Aust. 1947, 71, 212–224. [Google Scholar]
- Young, G.G.A.; Hagadorn, J.W. Evolving preservation and facies distribution of fossil jellyfish: A slowly closing taphonomic window. Boll. Della Soc. Paleontol. Ital. 2020, 59, 185–203. [Google Scholar]
- Ballèvre, M.; Bosse, V.; Dabard, M.P.; Ducassou, C.; Fourcade, S.; Paquette, J.L.; Peucat, J.J.; Pitra, P. Histoire géologique du massif Armoricain: Actualité de la recherche. Bull. Soc. Géol. Minér. Bretagne 2013, 10–11, 5–96. [Google Scholar]
- Chauvel, J.J.; Mansuy, C. Micropaléontologie du Protérozoïque du Massif armoricain (France). Precambrian Res. 1981, 15, 25–42. [Google Scholar] [CrossRef]
- Chauvel, J.J.; Schopf, J. Late Precambrian microfossils from Brioverian cherts and limestone’s of Brittany and Normandy, France. Nature 1978, 275, 640–642. [Google Scholar] [CrossRef]
- Deflandre, G. Sur l’existence, dès le Précambrien, d’Acritarches du type Acanthomorphitae: Eomocrhystridium nov.gen. Typification du genre Palaeocryptidium. Comptes Rendus Acad. Des. Sci. Paris 1968, 266, 2385–2389. [Google Scholar]
- Mansuy, C. Les Microsphères du Protérozoïque Supérieur Armoricain (Briovérien): Nature, Repartition Stratigraphique, Affinités Biologiques. Doctoral Dissertation, Université de Rennes I, Rennes, France, 1983; 140p. [Google Scholar]
- Roblot, M.M. Sporomorphes du Précambrien armoricain. Ann. Paléontol. 1964, 2, 105–110. [Google Scholar]
- Van Iten, H.; Marques, A.C.; Leme, J.M.; Pacheco, M.L.A.F.; Simões, M.G. Origin and early diversification of the phylum Cnidaria Verrill: Major developments in the analysis of the taxon’s Proterozoic–Cambrian history. Palaeontology 2014, 57, 677–690. [Google Scholar] [CrossRef]
- Dabard, M.P.; Néraudeau, D. Stratotype Briovérien. Collection Stratotypes, MNHN & BRGM, Ed.; 2024; in press.
- Coutret, B.; Néraudeau, D. Quantitative analysis of horizontal bioturbation from Brioverian (Ediacaran–Fortunian) deposits of Brittany (Armorican Massif, NW of France). Ann. Paléontol. 2022, 108, 102543. [Google Scholar] [CrossRef]
- Gougeon, R.; Néraudeau, D.; Dabard, M.P.; Pierson-Wickmann, A.C.; Polette, F.; Poujol, M.; Saint-Martin, J.P. Trace fossils from the Brioverian (Ediacaran-Fortunian) in Brittany (NW France). Ichnos 2018, 25, 11–24. [Google Scholar] [CrossRef]
- Gougeon, R.; Néraudeau, D.; Poujol, M.; Loi, A. Loops, spirals and the appearance of guided behaviors from the Ediacaran-Cambrian of Brittany, NW France. Estud. Geol. 2019, 75, 11–13. [Google Scholar]
- Gougeon, R.; Néraudeau, D.; Loi, A.; Poujol, M. New insights into the early evolution of horizontal spiral trace fossils and the age of the Brioverian series (Ediacaran-Cambrian) in Brittany, NW France. Geol. Mag. 2022, 159, 1284–1294. [Google Scholar] [CrossRef]
- Néraudeau, D. Algal megafossil Chuaria from the Brioverian (Ediacarn-Fortunian) of Britanny, NW France. Rev. Paléobiologie 2023, 42, 237–249. [Google Scholar]
- Néraudeau, D.; Dabard, M.P. Le Briovérien revisité. Bull. Soc. Géol. Minéral. Bretagne 2023, 21, 3–18. [Google Scholar]
- Néraudeau, D.; Dabard, M.P.; El Albani, A.; Gougeon, R.; Mazurier, A.; Pierson—Wickmann, A.C.; Poujol, M.; Saint Martin, J.P.; Saint Martin, S. First evidence of Ediacaran–Fortunian elliptical body fossils in the Brioverian series of Brittany, NW France. Lethaia 2018, 51, 513–522. [Google Scholar] [CrossRef]
- Néraudeau, D.; Coutret, B.; Gendry, D.; Lesage, B.; Loi, A.; Poujol, M. Elliptical body fossils from the Fortunian (Early Cambrian) of Normandy (NW France). CR Palevol 2023, 22, 439–453. [Google Scholar] [CrossRef]
- Landing, E.; Narbonne, G.M. Scenella and « a Chondrophorine (Medusoid HYdrozoan) from the Basal Cambrian (Placentian) of Newfoundland ». J. Paleontol. 1992, 66, 138. [Google Scholar] [CrossRef]
- Jensen, S. The Proterozoic and earliest Cambrian trace fossil record; patterns, problems and perspectives. Integr. Comp. Biol. 2003, 43, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Lebesconte, M. Constitution générale du Massif breton comparée à celle du Finistère. Bull. Soc. Géol. Fr. 1886, 14, 776–820. [Google Scholar]
- Hagadorn, J.W.; Waggoner, B. Ediacaran fossils from the southwestern Great Basin, United States. J. Paleontol. 2000, 74, 349–359. [Google Scholar] [CrossRef]
- Màngano, M.G.; Buatois, L.A. The Trace-Fossil Record of Major Evolutionary Events. Volume 1: Precambrian and Paleozoic. Top. Geobiol. 2016, 39, 358. [Google Scholar]
- Màngano, M.G.; Buatois, L.A. The rise and early evolution of animals: Where do we stand from a trace-fossil perspective? Interface Focus 2020, 10, 20190103. [Google Scholar] [CrossRef] [PubMed]
- Paquette, J.-L.; Piro, J.-L.; Devidal, J.-L.; Bosse, V.; Didier, A.; Sanac, S.; Abdelnous, Y. Sensitivity enhancement in LA-ICP-MS by N2 addition to carrier gas: Application to radiometric dating of U-Th-bearing minerals. Agil. ICP-MS J. 2014, 58, 4–5. [Google Scholar]
- Nosenzo, F.; Manzotti, P.; Poujol, M.; Ballèvre, M.; Langlade, J. A window into an older orogenic cycle: P–T conditions and timing of the pre-Alpine history of the Dora-Maira Massif (Western Alps). J. Metamorph. Geol. 2022, 40, 789–821. [Google Scholar] [CrossRef]
- Jackson, S.E.; Pearson, N.J.; Griffin, W.L.; Belousova, E.A. The application of laser ablation-inductively coupled plasma-mass spectrometry to in situ U–Pb zircon geochronology. Chem. Geol. 2004, 211, 47–69. [Google Scholar] [CrossRef]
- Sláma, J.; Kosler, J.; Condon, D.J.; Crowley, J.L.; Gerdes, A.; Hanchar, J.M.; Whitehouse, M.J. Plešovice zircon—A new natura l reference material for U-Pb and Hf isotopic microanalysis. Chem. Geol. 2008, 249, 1–35. [Google Scholar] [CrossRef]
- Vermeesch, P. IsoplotR: A free and open toolbox for geochronology. Geosci. Front. 2018, 9, 1479–1493. [Google Scholar] [CrossRef]
- Kahle, C.F.J. Subaerial gutter marks. Raindrop impressions, scour marks and other sedimentary structures, SiluriannTymochee Dolomite, Waterville, Ohio. Carbonates Evaporites 2009, 24, 33–44. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kim, K.S.; Kim, T.S. Raindrop imprints from the Late Proterozoic Sangwon System of the Socheong Island of Ongjin-gun, Incheon, Korea. J. Korean Earth Sci. Soc. 1999, 20, 55–60. [Google Scholar]
- Van Der Westhuizen, W.A.; Grobler, N.J.; Loock, J.C.; Tordiffe, E.A.W. Raindrops imprints in the Late Archaean—Early Proterozoic Ventersdorp Supergroup, South Africa. Sediment. Geol. 1989, 61, 303–309. [Google Scholar] [CrossRef]
- Collinson, J.D.; Thompson, D.B. Sedimentary Structures, 2nd ed.; Unwin Hyman: London, UK, 1989; 207p. [Google Scholar]
- Twenhofel, W.H. Principles of Sedimentation, 2nd ed.; McGraw Hill Book Company, Inc.: New York, NY, USA, 1950; 673p. [Google Scholar]
- Arrouy, M.J.; Gomez-Peral, L.E.; Penzo, V.; Ferreyra, C.; Poiré, D.G. Fossil bubble structure related to microbial activity coeval with the Middle Ediacaran oceanic oxygénation évent in the Tandilia system. Lat. Am. J. Sedimentol. Basin Anal. 2021, 28, 99–118. [Google Scholar]
- Bosak, T.; Bush, J.W.M.; Flynn, M.; Liang, B.; Ono, S.; Petroff, A.P.; Sim, M.S. Formation and stability of oxygen-rich bubbles that shape photosynthetic mats. Geobiology 2010, 8, 45–55. [Google Scholar] [CrossRef]
- Fedonkin, M.A. New representatives of the Precambrian coelenterates in the northern Russian platform. Paleontol. Zhurnal 1980, 2, 7–15. (In Russian) [Google Scholar]
- Fedonkin, M.A.; Gehling, J.G.; Grey, K.; Narbonne, G.M.; Vickers-Rich, P. The Rise of Animals: Evolution and Diversification of the Kingdom Animalia; The Johns Hopkins University Press: Baltimore, MD, USA, 2007; 327p. [Google Scholar]
- Meert, J.G.; Gibsher, A.S.; Levashova, N.M.; Grice, W.C.; Kamenov, G.D.; Ryabinin, A.B. Glaciation and 770 Ma Ediacara (?) Fossils from the Lesser Karatau Microcontinent, Kazakhstan. Gondwana Res. 2011, 19, 867–880. [Google Scholar] [CrossRef]
- Sepkoski, J.J., Jr. A compendium of fossil marine animal genera. Bull. Am. Paleontol. 2002, 363, 1–560. [Google Scholar]
- Liu, A.G.; Brasier, M.D.; Bogolepova, O.K.; Raevskaya, E.G.; Gubanov, A.P. First report of a newly discovered Ediacaran biota from the Irkineeva Uplift, East Siberia. Newsl. Stratigr. 2013, 46, 95–110. [Google Scholar] [CrossRef]
- Jensen, S.; Gehlin, G.J.G.; Droser, M.L.; Grant, S.W.F. A scratch circle origin for the medusoid fossil Kullingia. Lethaia 2002, 35, 291–299. [Google Scholar] [CrossRef]
- Bertrand-Sarfati, I.; Moussine-Pouchkine, A.; Amard, B.; Ait Kaci, A.A. First Ediacaran fauna found in Western Africa and evidence for an Early Cambrian glaciation. Geology 1995, 23, 133–136. [Google Scholar] [CrossRef]
- Buatois, L. Treptichnus pedum and the Ediacaran-Cambrian boundary: Significance and caveats. Geol. Mag. 2018, 155, 174–180. [Google Scholar] [CrossRef]
- Narbonne, G.M.; Myrow, P.M.; Landing, E.; Anderson, M.M. A candidate stratotype for the Precambrian-Cambrian boundary, Fortune Head, Burin Peninsula, southeastern New-Foundland. Can. J. Earth Sci. 1987, 24, 1277–1293. [Google Scholar] [CrossRef]
- Ram, H.; Harsh, A.; Kumar, P.; Parihar, V.S. Nimbia: The discoid organisms from Ediacaran Sonia Dansstone of Jodhpur Group, Marwar Supergroup, western India. Curr. Sci. 2023, 125, 999–1004. [Google Scholar]
- Brasier, M.D.; Perejon, A.; San Jose, M.A. Discovery of an important fossiliferous Precambrian-Cambrian sequence in Spain. Estud. Geol. 1979, 35, 379–383. [Google Scholar]
- Vidal, G.; Palacios, T.; Gamez-Vintaned, J.A.; Diez Balda, M.A.; Grant, S.W.F. Neoproterozoic-early Cambrian geology and palaeontology of Iberia. Geol. Mag. 1994, 131, 729–765. [Google Scholar] [CrossRef]
- McCall, G.J.H. The Vendian (Ediacaran) in the geological record: Enigmas in geology’s prelude to the Cambrian explosion. Earth-Sci. Rev. 2006, 77, 1–229. [Google Scholar] [CrossRef]
- Jensen, S.; Palacios, T.; Marti Mus, M. A brief review of the fossil record of the Ediacaran-Cambrian transition in the area of Montes de Toledo-Guadalupe, Spain. In The Rise and Fall of the Ediacaran Biota; Vickers-Rich, P., Komarower, P., Eds.; Geological Society, London, Special Publications: London, UK, 2007; Volume 286, pp. 223–235. [Google Scholar]
- Saint Martin, J.-P.; Saint Martin, S. Beltanelliformis brunsae Menner in Keller, Menner, Stepanov & Chumakov, 1974: And Ediacaran fossil from Neoproterozoic of Dobrogea (Romanian). Geodiversitas 2018, 40, 537–548. [Google Scholar]
- Ivantsov, A.Y. Finds of Ediacaran-Type Fossils in Vendian Deposits of the Yudoma Group, Eastern Siberia. Dokl. Akad. Nauk 2017, 472, 542–545. [Google Scholar] [CrossRef]
- Palij, V.M. Remains of a skeletonless fauna and traces of life ability from deposits of the Upper Precambrian and Lower Cambrian of Podolia. In Paleontology and Stratigraphy of the Upper Precambrian and Lower Paleozoic of the southwestern East European Platform; Naukova Dumka: Kiev, Ukraine, 1976; pp. 63–77. [Google Scholar]
- Sokolov, B.S. Vendian stage in the history of the Earth. In XXIV Session of the International Geological Congress: Reports of Soviet Geologists; Nauka: Moscow, Russia, 1972; pp. 114–125. [Google Scholar]
- Jensen, S.; Högström, A.E.S.; Almond, J.; Taylor, W.L.; Meinhold, G.; Hoyberget, M.; Ebbestad, J.O.R.; Agic, H.; Palacios, T. Scratch circles from the Ediacaran and Cambrian of Arctic Norway and southern Africa, with a review of scratch circle occurrences. Bull. Geosci. 2018, 93, 287–304. [Google Scholar] [CrossRef]
- Tarhan, L.G.; Droser, M.L.; Gehling, J.G. Taphonomic controls on Ediacaran diversity: Uncovering the holdfast origin of morphologically variable enigmatic structures. Palaios 2010, 25, 823–830. [Google Scholar] [CrossRef]
- Young, G.G.A.; Hagadorn, J.W. The fossil record of cnidarian medusae. Palaeoworld 2010, 19, 212–221. [Google Scholar] [CrossRef]
- Arrouy, M.J.; Warren, L.V.; Quaglio, F.; Poiré, D.G.; Guimaraes Simoes, M.; Boselli Rosa, M.; Gomez-Peral, L.E. Ediacaran discs from South America: Probable soft-bodied macrofossils unlock the paleogeography of the Clymene Ocean. Sci. Rep. 2016, 6, 30590. [Google Scholar] [CrossRef] [PubMed]
- Boyce, W.D.; Reynolds, K. The Ediacaran fossil Aspidella terranovica Billings, 1872 from St John’s Convention Centre test pit CjAe-33. Curr. Res. 2008, 8, 55–61. [Google Scholar]
- Tarhan, L.G.; Droser, M.L.; Gehling, J.G.; Dzaugis, M.P. Taphonomy and morphology of the Ediacaran form Genus Aspidella. Precambrian Res. 2015, 257, 124–136. [Google Scholar] [CrossRef]
- Hagadorn, J.W.; Miller, R.F. Hypothesized Cambrian medusae from Saint John, New Brunswick, reinterpreted as sedimentary structures. Atl. Geol. 2011, 47, 66–80. [Google Scholar] [CrossRef][Green Version]
- Farmer, J.; Vidal, G.; Moczydlowska, M.; Strauss, H.; Ahlberg, P.; Siedlecka, A. Ediacaran fossils from the Innerelv Member (late Proterozoic) of the Tanafjorden area, northeastern Finnmark. Geol. Mag. 1991, 129, 181–195. [Google Scholar] [CrossRef]
- Lerner, A.J.; Lucas, S.P. The pseudofossil Astropolithon from the Lower Permian Abo Formation of Socorro County, Central New Mexico. Carbonif.-Permian Transit. Socorro Cty. N. M. 2017, 77, 225–231. [Google Scholar]
- Lucas, S.G.; Lerner, A.J. The rare unusual pseudofossil Astropolithon from the Lower Permian Abo Formation near Socorro, New Mexico. New Mex. Geol. 2017, 39, 40–42. [Google Scholar] [CrossRef]
- Babcock, L.E.; Robison, R.A. Taxonomy and paleobiology of some Middle Cambrian Scenella (cnidaria) and hyolithids (Mollusca) from western North America. The University of Kansas Paleontological Contributions 1988, 121, 1–22. [Google Scholar]
- Chapman, F. New or little known Victorian fossils in the National Museum. Proc. R. Soc. Vic. 1911, 23, 305–324. [Google Scholar]
- Alvaro, J.-J.; Valverde-Vaquero, P.; Jensen, S. Multidisciplinary re-assessment of the Ediacaran-Cambrian interval in south-western Europe. Newsl. Stratigr. 2024. [CrossRef]
- Peel, J.S. An enigmatic cap-shaped fossil from the Middle Cambrian of North Greenland. Bull. Grønl. Geol. Unders. 1994, 169, 149–155. [Google Scholar] [CrossRef]
- Skovsted, C.B.; Brock, G.A.; Lindström, A.; Peel, J.S.; Paterson, J.R.; Fuller, M.K. Early Cambrian record of failed durophagy and shell repair in an epibenthic mollusk. Biol. Lett. 2007, 3, 314–317. [Google Scholar] [CrossRef]
- Narbonne, G.M.; Myrow, P.M.; Landing, E.; Anderson, M.M. A Chondrophorine (Medusoid Hydrozoan) from the Basal Cambrian (Placentian) of Newfoundland. J. Paleontol. 1991, 65, 186–191. [Google Scholar] [CrossRef]
- Shi, X.; Zhang, C.; Jiang, G.; Liu, J.; Wang, Y.; Liu, D. Microbial Mats in the Mesoproterozoic Carbonates of the North China Platform and their potential for hydrocarbon generation. J. China Univ. Geosci. 2008, 19, 549–566. [Google Scholar]
- Martinsen, O.J. Slide and slump structures. In Encyclopedia of Sediments and Sedimentary Rocks; Encyclopedia of Earth Sciences Series Middleton; Church, M., Hardie, L.A., Longstaffe, F.J., Middleton, V., Coniglio, M.A., Eds.; Springer: Dordrecht, The Netherlands, 1978; 821p. [Google Scholar]
- Paton, C.; Woodhead, J.D.; Hellstrom, J.C.; Hergt, J.M.; Greig, A.; Maas, R. Improved laser ablation U-Pb zircon geochronology through robust downhole fractionation correction. Geochem. Geophys. Geosyst. 2010, 11, Q0AA06. [Google Scholar] [CrossRef]
- Horstwood, M.S.A.; Košler, J.; Gehrels , G.; Jackson, S.E.; McLean , N.M.; Paton, C.; Pearson, N.J.; Sircombe, K.; Sylvester, P.; Vermeesch, P.; et al. Community-Derived Standards for LA-ICP-MS U-(Th-)Pb Geochronology—Uncertainty Propagation, Age Interpretation and Data Reporting. Geostand. Geoanal. Res. 2016, 40, 311–332. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).