
Reconciling Divergent Ages for the Oldest Recorded Air-Breathing Land Animal, the Millipede, Pneumodesmus newmani Wilson & Anderson, 2004: A Review of the Geology and Ages of the Basal Old Red Sandstone Stonehaven Group (Silurian–Early Devonian), Aberdeenshire, Scotland




Abstract
:
1. Introduction
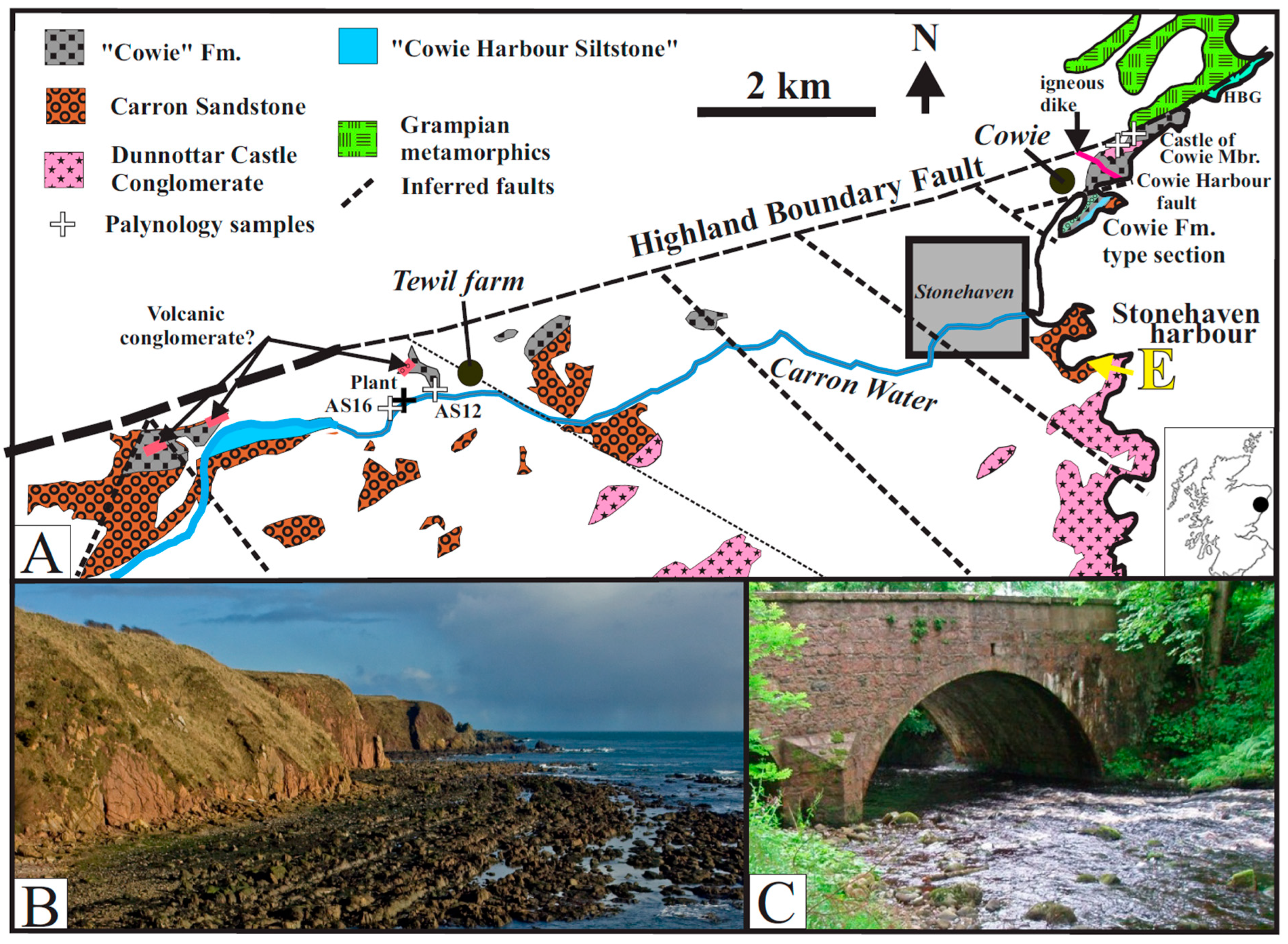
2. Structural Units
2.1. Inland Exposures
2.2. Coastal Exposures
3. Descriptions and Interpretations of the Northern and Southern Blocks
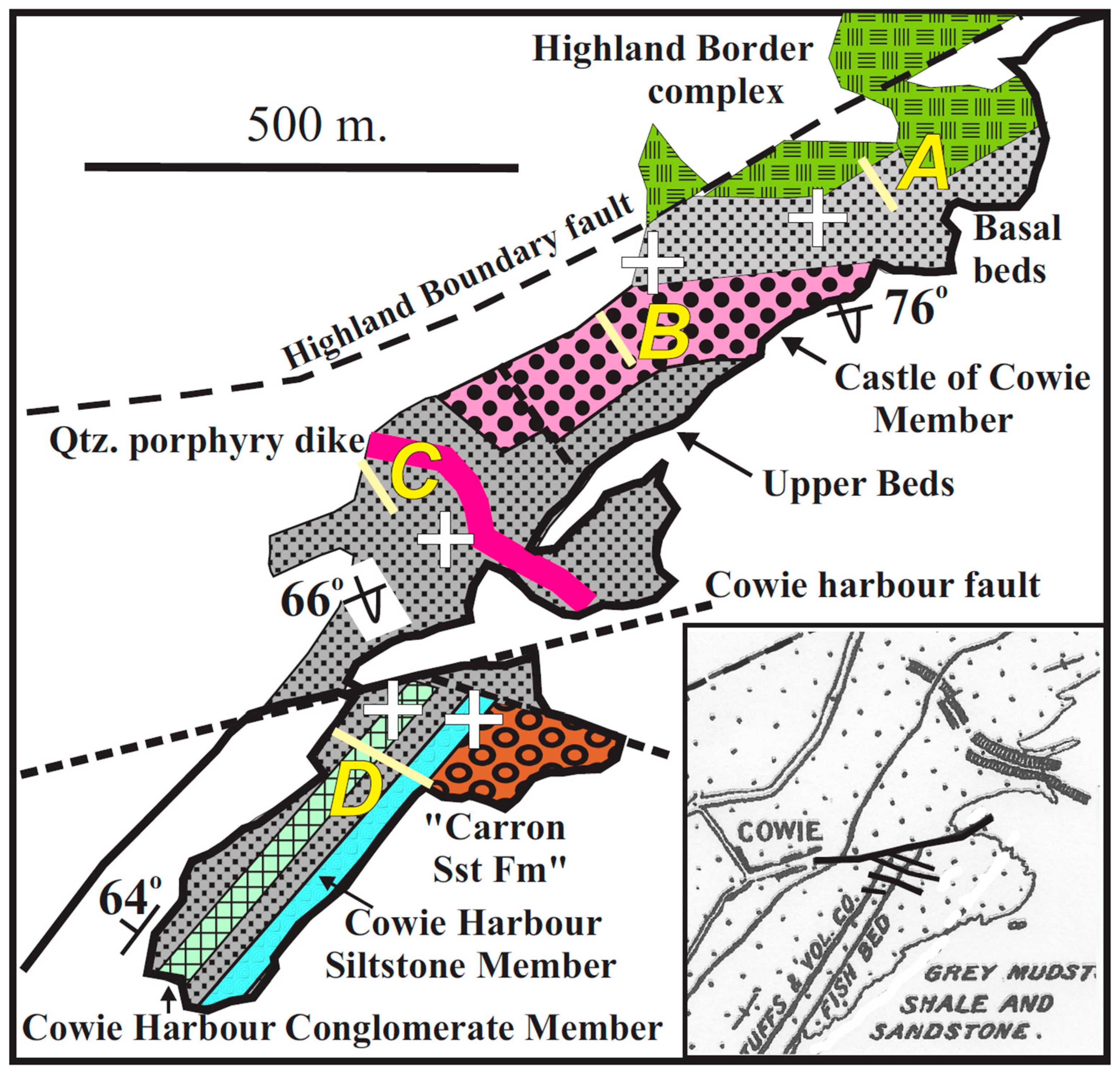
3.1. Northern Block
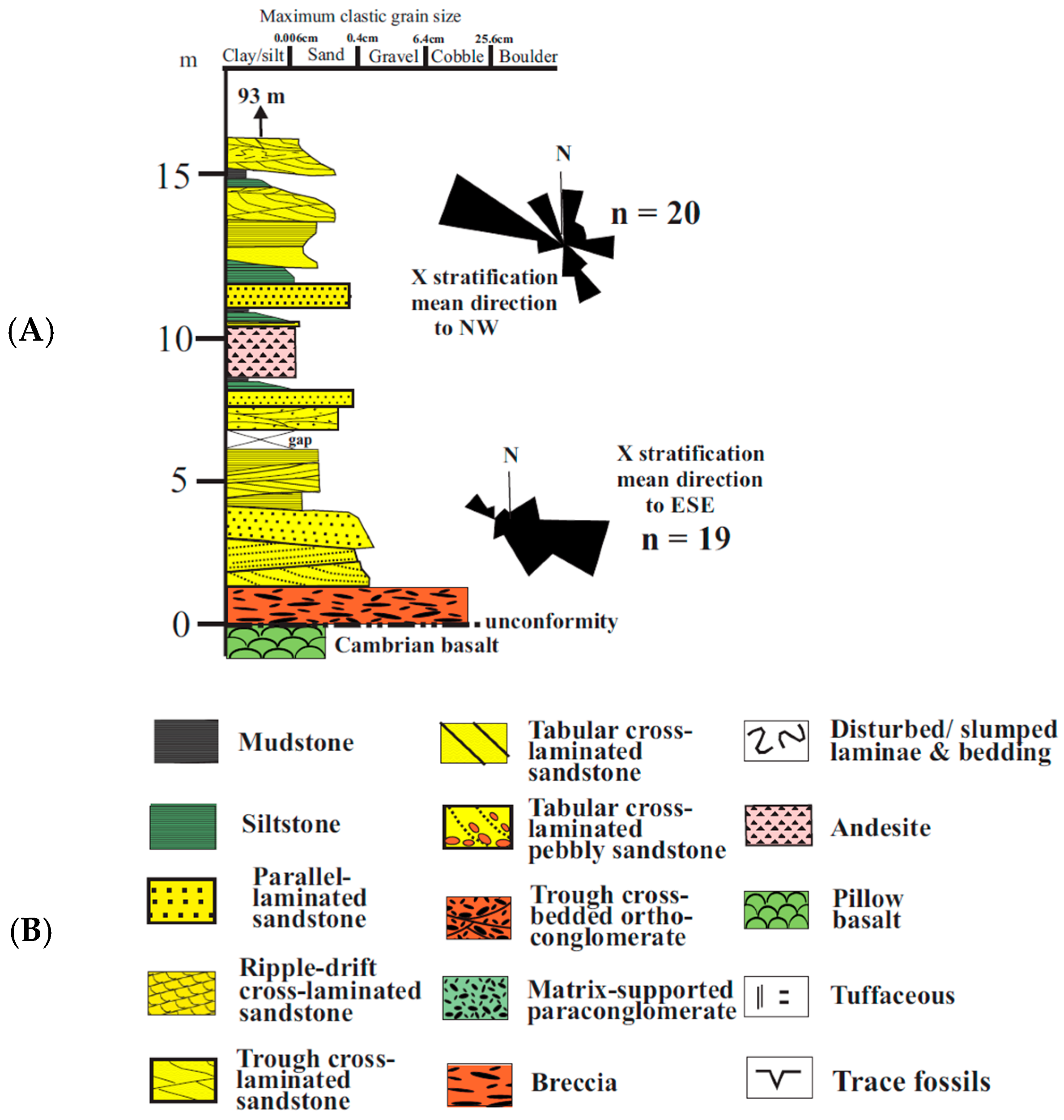
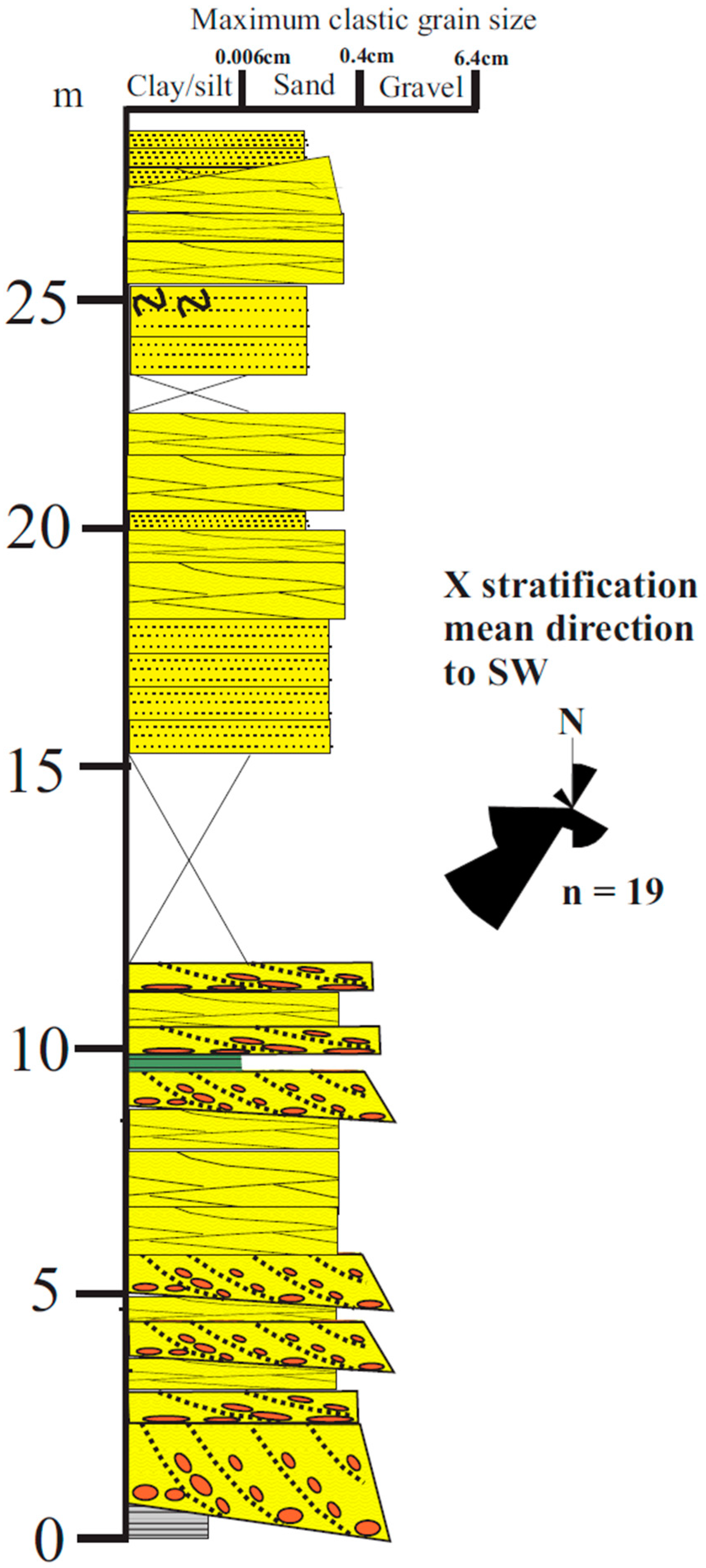
3.1.1. Sediments
3.1.2. Fossils
3.1.3. Detrital Zircon U/Pb Dates
3.1.4. Summary
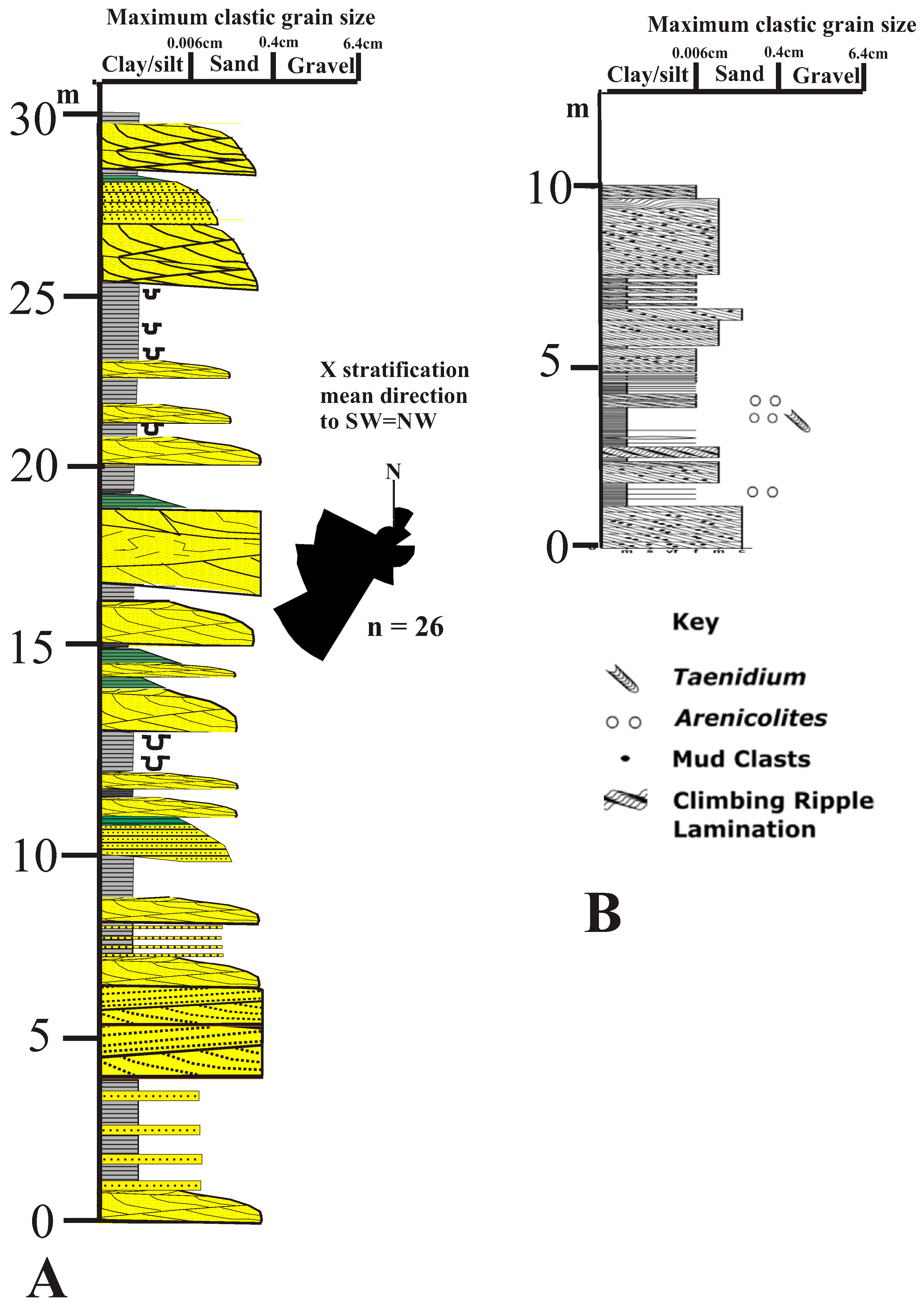
3.2. Southern Block
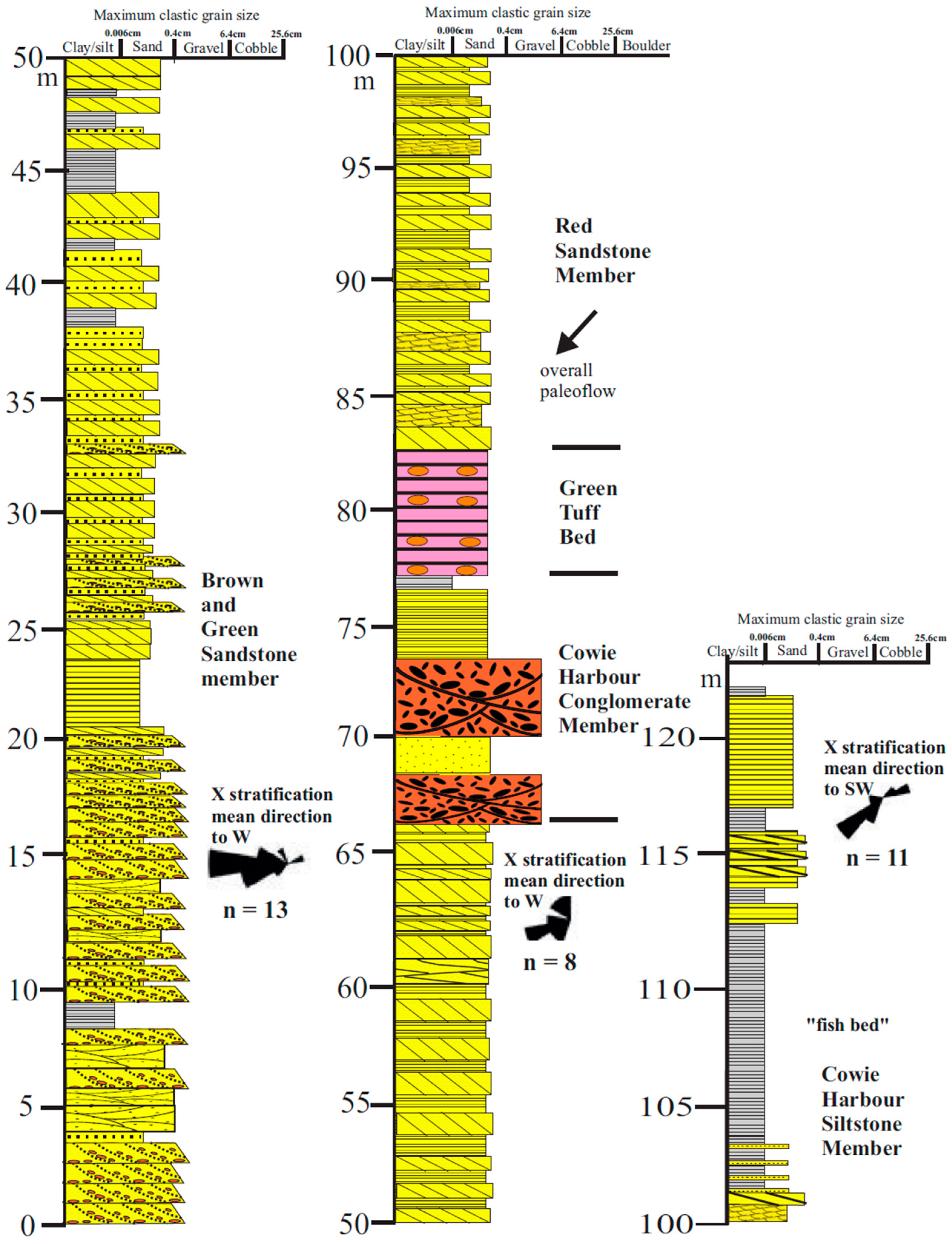
3.2.1. Sediments
3.2.2. Fossils
3.2.3. Detrital Zircon U/Pb Dates
3.2.4. Summary
4. Conclusions
5. Future Work
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilson, H.; Anderson, L. Morphology and taxonomy of Paleozoic millipedes (Diplopoda: Chilognatha: Archipolypoda) from Scotland. J. Paleontol. 2004, 78, 169–184. [Google Scholar] [CrossRef]
- Brookfield, M.E.; Catlos, E.J.; Suarez, S.E. Myriapod divergence times differ between molecular clock and fossil evidence: U/Pb zircon ages of the earliest fossil millipede-bearing sediments and their significance. Hist. Biol. 2020, 33, 2014–2018. [Google Scholar] [CrossRef]
- Buatois, L.A.; Davies, N.S.; Gibling, M.R.; Krapovickas, V.; Labandeira, C.C.; MacNaughton, R.B.; Mángano, M.G.; Minter, N.J.; Shillito, A.P. The Invasion of the Land in Deep Time: Integrating Paleozoic Records of Paleobiology, Ichnology, Sedimentology, and Geomorphology. Integr. Comp. Biol. 2022, 62, 297–331. [Google Scholar] [CrossRef]
- Wellman, C.H.; Lopes, G.; McKellar, Z.; Hartley, A. Age of the basal ‘Lower Old Red Sandstone’ Stonehaven Group of Scotland: The oldest reported air-breathing land animal is Silurian (late Wenlock) in age. J. Geol. Soc. 2023, 181, jgs2023-138. [Google Scholar] [CrossRef]
- Melchin, M.J.; Sadler, P.M.; Cramer, B.D. Chapter 21—The Silurian period. In Geologic Time Scale 2020; Gradstein, F.M., Ogg, J.G., Schmitz, M.D., Ogg, G.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 695–732. [Google Scholar] [CrossRef]
- Becker, R.T.; Marshall, J.A.E.; Da Silva, A.-C.; Agterberg, F.P.; Gradstein, F.M.; Ogg, J.G. Chapter 22—The Devonian Period. In Geologic Time Scale 2020; Gradstein, F.M., Ogg, J.G., Schmitz, M.D., Ogg, G.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 733–810. [Google Scholar] [CrossRef]
- Westoll, T.S. The vertebrate-bearing strata of Scotland. In Proceedings of the 18th International Geological Congress, London, UK, 31 December 1951; Volume 2, pp. 5–21. [Google Scholar]
- Marshall, J.E.A. Palynology of the Stonehaven Group, Scotland: Evidence for a mid-Silurian age and its geological implications. Geol. Mag. 1991, 128, 283–286. [Google Scholar] [CrossRef]
- Wellman, C.H. A land plant microfossil assemblage of Mid Silurian age from the Stonehaven Group, Scotland. J. Micropalaeontol. 1993, 12, 47–66. [Google Scholar] [CrossRef]
- Suarez, S.E.; Brookfield, M.E.; Catlos, E.J.; Stöckli, D.F. A U-Pb zircon age constraint on the oldest-recorded air-breathing land animal. PLoS ONE 2017, 12, e0179262. [Google Scholar] [CrossRef]
- Gillen, G.; Trewin, N.H. Dunnottar to Stonehaven and the Highland Boundary Fault. In Excursion Guide to the Geology of the Aberdeen Area; Trewin, N.H., Kneller, B.C., Gillen, C., Eds.; Scottish Academic Press: Edinburgh, UK, 1986; pp. 265–273. [Google Scholar]
- McKellar, Z. Sedimentology of the Lower Old Red Sandstone of the Northern Midland Valley Basin and Grampian Outliers, Scotland: Implications for Post-Orogenic Basin Development. Ph.D. Thesis, University of Aberdeen, Aberdeen, UK, 2017; 303p. [Google Scholar]
- McKellar, Z.; Hartley, A.J. Caledonian foreland basin sedimentation: A new depositional model for the Upper Silurian-Lower Devonian Lower Old Red Sandstone of the Midland Valley Basin, Scotland. Basin Res. 2021, 33, 754–778. [Google Scholar] [CrossRef]
- Campbell, R. The geology of northeastern Kincardineshire. Trans. R. Soc. Edinb. 1913, 48, 1–37. [Google Scholar] [CrossRef]
- MacGregor, A.R. Fife and Angus Geology: An Excursion Guide. Excursion 1. Arbroath, Crawton and Stonehaven, 3rd ed.; The Pentland Press: Edinburgh, UK, 1996. [Google Scholar]
- Hartley, A.J.; Leleu, S. Sedimentological constraints on the late Silurian history of the Highland Boundary Fault, Scotland: Implications for Midland Valley Basin development. J. Geol. Soc. 2015, 172, 213–217. [Google Scholar] [CrossRef]
- Shillito, A.P.; Davies, N.S. Archetypically Siluro-Devonian ichnofauna in the Cowie Formation, Scotland: Implications for the myriapod fossil record and Highland Boundary Fault Movement. Proc. Geol. Assoc. 2017, 128, 815–828. [Google Scholar] [CrossRef]
- Anderson, J.G.C. The Geology of the Highland Border: Stonehaven to Arran. Trans. R. Soc. Edinb. 1947, 61, 479–515. [Google Scholar] [CrossRef]
- McKay, L.; Shipton, Z.K.; Lunn, R.J.; Andrews, B.; Raub, T.D.; Boyce, A.J. Detailed internal structure and along-strike variability of the core of a plate boundary fault: The Highland Boundary fault, Scotland. J. Geol. Soc. 2020, 177, 283–296. [Google Scholar] [CrossRef]
- Phillips, E.R. Petrology and Provenance of the Siluro-Devonian (Old Red Sandstone Facies) Sedimentary Rocks of the Midland Valley, Scotland; British Geological Survey Internal Report IR/07/040; British Geological Survey: Nottingham, UK, 2007; 65p.
- Smith, N.D. Transverse bars and braiding in the lower Platte River, Nebraska. Geol. Soc. Am. Bull. 1971, 82, 3407–3420. [Google Scholar] [CrossRef]
- Miall, A.D. Fluvial Depositional Systems; Springer International: Cham, Switzerland, 2014; 316p. [Google Scholar]
- Li, Z.; Wang, Y.; Zhu, W.; Gao, H.; Liu, F.; Xing, W.; Zhang, C.; Qiao, Q.; Lei, X. Fluvial Responses to Late Quaternary Climate Change in a Humid and Semi-Humid Transitional Area: Insights from the Upper Huaihe River, Eastern China. Water 2023, 15, 1767. [Google Scholar] [CrossRef]
- Finotello, A.; Ielpi, A.; Lapôtre, M.G.A.; Lazarus, E.D.; Ghinassi, M.; Carniello, L.; Favaro, S.; Tognin, D.; D’Alpaos, A. Vegetation enhances curvature-driven dynamics in meandering rivers. Nat. Commun. 2024, 15, 1968. [Google Scholar] [CrossRef]
- Wang, S.A. Method for Estimating the Hydrodynamic Values of Anastomosing Rivers: The Expression of Channel Morphological Parameters. Water 2024, 16, 163. [Google Scholar] [CrossRef]
- Makaske, B. Anastomosing rivers: A review of their classification, origin and sedimentary products. Earth-Sci. Rev. 2001, 53, 149–196. [Google Scholar] [CrossRef]
- Ferguson, R.I. The Threshold between Meandering and Braiding. In Channels and Channel Control Structures; Smith, K.V.H., Ed.; Springer: Berlin/Heidelberg, Germany, 1984; pp. 749–763. [Google Scholar] [CrossRef]
- Gibling, M.R.; Davies, N.S. Palaeozoic landscapes shaped by plant evolution. Nat. Geosci. 2012, 5, 99–105. [Google Scholar] [CrossRef]
- Retallack, G.J. Chapter 21—Paleozoic paleosols. In Developments in Earth Surface Processes; Martini, I.P., Chesworth, W., Eds.; Elsevier: Amsterdam, The Netherlands, 1992; Volume 2, pp. 543–564. [Google Scholar] [CrossRef]
- Retallack, G.J. Ordovician-Devonian lichen canopies before evolution of woody trees. Gondwana Res. 2022, 106, 211–223. [Google Scholar] [CrossRef]
- Williams, G.E. Flood deposits of the sand-bed ephemeral streams of central Australia. Sedimentology 1971, 17, 1–40. [Google Scholar] [CrossRef]
- Nanson, G.G.; Rust, B.R.; Taylor, G. Coexistent mud braids and anastomosing channel in an arid-zone river: Cooper Creek, central Australia. Geology 1986, 14, 175–178. [Google Scholar] [CrossRef]
- Pickup, G. Event frequency and landscape stability on the flood plain systems of arid central Australia. Quat. Sci. Rev. 1991, 10, 463–473. [Google Scholar] [CrossRef]
- Bourke, M.C. Cyclical Construction and Destruction of Flood Dominated Semiarid Central Australia. In Variability in Stream Erosion and Sediment Transport; Olive, L.J., Loughlan, R., Kesby, J.A., Eds.; International Association of Hydrological Sciences Publication: Wallingford, UK, 1994; Volume 224, pp. 113–123. [Google Scholar]
- Webster, M.; Sadler, P.; Kooser, M.; Fowler, E. Combining stratigraphic sections and museum collections to increase biostratigraphic resolution. In High-Resolution Approaches in Stratigraphic Paleontology; Harries, P.J., Ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 95–128. [Google Scholar] [CrossRef]
- White, J. Palynodata Datafile: 2006 Version. Geological Survey of Canada Open File Report 5793. 2008. Available online: https://paleobotany.ru/palynodata (accessed on 13 February 2025).
- Stanley, E.A. The problem of reworked pollen and spores in marine sediments. Mar. Geol. 1966, 4, 397–408. [Google Scholar] [CrossRef]
- Burgess, N.D.; Richardson, J.B. Late Wenlock to Early Přídolí cryptospores and miospores from South and southwest Wales, Great Britain. Palaeontographica B 1995, 236, 1–44. [Google Scholar]
- Rubinstein, C.V.; Steemans, P. Miospore assemblages from the Silurian–Devonian boundary, in borehole A1-61, Ghadamis Basin, Libya. Rev. Palaeobot. Palynol. 2002, 118, 397–421. [Google Scholar] [CrossRef]
- Burgess, N.D.; Richardson, J.B. Silurian cryptospores and miospores from the type Wenlock area, Shropshire, England. Palaeontology 1991, 34, 601–628. [Google Scholar]
- Wetherall, P.M.; Dorning, K.; Wellman, C.H. Palynology, biostratigraphy, and depositional environments around the Ludlow-Pridoli boundary at Woodbury Quarry, Herefordshire, England. Boll. Soc. Paleontol. Ital. 1999, 38, 397–404. [Google Scholar]
- Steemans, P.; Le Hérissé, A.; Bozdogan, N. Ordovician and Silurian cryptospores and miospores from southeastern Turkey. Rev. Palaeobot. Palynol. 1996, 93, 35–76. [Google Scholar] [CrossRef]
- McKellar, Z.; Hartley, A.J.; Morton, A.; Frei, D. A Multidisciplinary Approach to Sediment Provenance Analysis of the Late Silurian-Devonian Lower Old Red Sandstone succession, Northern Midland Valley Basin, Scotland. J. Geol. Soc. 2020, 177, 297–314. [Google Scholar] [CrossRef]
- Horstwood, M.S.A.; Košler, J.; Gehrels, G.; Jackson, S.E.; McLean, N.M.; Paton, C.; Pearson, N.J.; Sircombe, K.; Sylvester, P.; Vermeesch, P.; et al. Community-Derived Standards for LA-ICP-MS U-(Th-)Pb Geochronology—Uncertainty Propagation, Age Interpretation and Data Reporting. Geostand. Geoanalytical Res. 2016, 40, 311–332. [Google Scholar] [CrossRef]
- Schoene, B.; Condon, D.J.; Morgan, L.; McLean, N. Precision and Accuracy in Geochronology. Elements 2013, 9, 19–24. [Google Scholar] [CrossRef]
- Sharman, G.R.; Matthew, A.; Malkowsi, A. Needles in a haystack: Detrital zircon U/Pb ages and the maximum depositional age of modern global sediment. Earth-Sci. Rev. 2020, 203, 103109. [Google Scholar] [CrossRef]
- Garza, H.K.; Catlos, E.J.; Chamberlain, K.R.; Suarez, S.E.; Brookfield, M.E.; Stockli, D.F.; Batchelor, R.A. How old is the Ordovician–Silurian boundary at Dob’s Linn, Scotland? Integrating LA-ICP-MS and CAID-TIMS U-Pb zircon dates. Geol. Mag. 2023, 160, 1775–1789. [Google Scholar] [CrossRef]
- Dickinson, W.R.; Gehrels, G.E. Use of U-Pb ages of detrital zircons to infer maximum depositional ages of strata: A test against a Colorado Plateau Mesozoic database. Earth Planet. Sci. Lett. 2009, 288, 115–125. [Google Scholar] [CrossRef]
- Bridge, J.S.; Lunt, I.A. Depositional models for braided rivers. In Braided Rivers; Sambrook Smith, G.H., Best, J.L., Bristow, C.S., Petts, G.E., Eds.; Blackwell Publishing: Oxford, UK, 2004; pp. 11–50. [Google Scholar]
- Allen, J.R.L. A quantitative model of grain size and sedimentary structures in lateral deposit. Geol. J. 2007, 7, 129–146. [Google Scholar] [CrossRef]
- Fielding, C.R.; Allen, J.P.; Alexander, J.; Gibling, M.R.; Rygel, M.C.; Calder, J.H. Fluvial Systems and their Deposits in Hot, Seasonal Semiarid and Subhumid Settings: Modern and Ancient Examples. In From River to Rock Record: The Preservation of Fluvial Sediments and Their Subsequent Interpretation; Davidson, S.K., Leleu, S., North, C.P., Eds.; Society for Sedimentary Geology Special Publication; SEPM Society for Sedimentary Geology: Claremore, OK, USA, 2011; Volume 97, pp. 89–111. [Google Scholar] [CrossRef]
- Colombera, L.; Mountney, N.P.; McCaffrey, W.D. A quantitative approach to fluvial facies models: Methods and example results. Sedimentology 2013, 60, 1526–1558. [Google Scholar] [CrossRef]
- Dewey, J.F.; Strachan, R.A. Changing Silurian–Devonian relative plate motion in the Caledonides: Sinistral transpression to sinistral transtension. J. Geol. Soc. 2003, 160, 219–229. [Google Scholar] [CrossRef]
- Westoll, T.S. A new cephalaspid fish from the Downtonian of Scotland, with notes on the structure and classification of ostracoderms. Trans. R. Soc. Edinb. 1945, 61, 341–357. [Google Scholar] [CrossRef]
- Ritchie, A. Ateleaspis tessellata Traquair, a non-cornate Cephalaspid from the Upper Silurian of Scotland. J. Linn. Soc. (Zool.) 1967, 47, 69–81. [Google Scholar] [CrossRef]
- Heintz, A. Additional remarks about Hemicyclaspis from Jeløya, southern Norway. Nor. Geol. Tidsskr. 1974, 54, 375–384. [Google Scholar]
- Blieck, A.; Elliott, D.K. Pteraspidomorphs (Vertebrata), the Old Red Sandstone, and the special case of the Brecon Beacons National Park, Wales, U.K. Proc. Geol. Assoc. 2017, 128, 438–446. [Google Scholar] [CrossRef]
- Dineley, D.L. New specimens of Traquairaspis from Canada. Palaeontology 1964, 7, 210–219. [Google Scholar]
- Tarrant, P.R. The ostracoderm Phialaspis from the Lower Devonian of the Welsh Borderland and South Wales. Palaeontology 1991, 34, 399–438. [Google Scholar]
- Elliott, D.K. Siluro-Devonian fish biostratigraphy of the Canadian arctic islands. Proc. Linn. Soc. New South Wales 1984, 107, 197–209. [Google Scholar]
- Dec, M. Traquairaspididae and Cyathaspididae (Heterostraci) from the Lower Devonian of Poland. Ann. Paléontol. 2020, 106, 13–23. [Google Scholar] [CrossRef]
- Talimaa, V. Vertebrate complexes in the heterofacial Lower Devonian deposits of Timan-Pechora Province. In Palaeozoic Microvertebrate Biochronology and Global Marine/Non-Marine Correlation (1991–1995) State of Research; Turner, S., Ed.; UNESCO-I.U.G.S. IGCP No. 328; Ichthyolith Issues, Special Publication 1; J. Zidek Serv: Socorro, NM, USA, 1995; 72p. [Google Scholar]
- Blom, H. A new anaspid fish from the Middle Silurian Cowie Harbour Fish Bed of Stonehaven, Scotland. J. Vertebr. Paleontol. 2008, 28, 594–600. [Google Scholar] [CrossRef]
- Blom, H.; Märss, T.; Miller, C. Silurian and earliest Devonian birkeniid anaspids from the Northern Hemisphere. Trans. R. Soc. Edinb. Earth Sci. 2002, 92, 263–323. [Google Scholar] [CrossRef]
- Davies, N.S.; Turnery, P.; Sansom, I.J. A revised stratigraphy of the Ringerike Group (Upper Silurian, Oslo, Norway). Nor. J. Geol. 2005, 85, 193–201. [Google Scholar]
- Kiaer, J. A new Downtonian fauna in the Sandstone Series of the Kristiania area: A preliminary report. Vidensabers Selskrab Skrifter I Matematisk-Naturvidenskabelig Klasse Kristiania 1911, 1911, 1–22. [Google Scholar]
- Kjellesvig-Waering, E.N. The Silurian Eurypterida of the Welsh Borderland. J. Paleontol. 1961, 35, 789–835. [Google Scholar]
- Tetlie, O.E. Distribution and dispersal history of Eurypterida (Chelicerata). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2007, 252, 557–574. [Google Scholar] [CrossRef]
- Störmer, L. Dictyocaris salter, a large crustacean from the Upper Silurian and Downtonian. Nor. Geol. Tidsskr. 1935, 15, 267–298. [Google Scholar]
- O’Connell, M. The habitat of the Eurypterida. Bull. Buffalo Soc. Nat. Hist. 1916, 11, 277. [Google Scholar]
- Ritchie, A. Palaeontological Studies on Scottish Silurian Fish Beds. Upublished Ph.D. Thesis, University of Edinburgh, Edinburgh, UK, 1963; 170p. [Google Scholar]
- Van der Brugghen, W. Dictyocaris, een enigmatisch fossiel uit het Silur. Grondboor Hamuir 1995, 1, 18–22. [Google Scholar]
- Brookfield, M.E.; Catlos, E.J.; Garca, H.K. The oldest ‘millipede’-plant association? Age, paleoenvironments and sources of the Silurian Lake sediments at Kerrera, Argyll and Bute, Scotland. Hist. Biol. 2024, 1–13. [Google Scholar] [CrossRef]
- Vermeesch, P. On the treatment of discordant detrital zircon U–Pb data. Geochronology 2021, 3, 247–257. [Google Scholar] [CrossRef]
- Kendall, R.S. The Old Red Sandstone of Britain and Ireland: A review. Proc. Geol. Assoc. 2017, 128, 409–421. [Google Scholar] [CrossRef]
- Lehnert, O.; Fryda, J.; Joachimski, M.; Meinhold, G.; Calner, M.; Čáp, P. The ‘Přídolí hothouse’, a trigger of faunal overturns across the latest Silurian Transgression Bioevent. In Proceedings of the 3rd IGCP 591 Annual Meeting, Lund, Sweden, 9–19 June 2013; Lindskog, A., Mehlqvist, K., Eds.; Lund University: Lund, Sweden, 2013; pp. 175–176. [Google Scholar]
- Howard, B.L.; Sharman, G.R.; Crowley, J.L.; Wersan, E.R. The leaky chronometer: Evidence for systematic cryptic Pb loss in laser ablation U-Pb dating of zircon relative to CA-TIMS. Terra Nova 2025, 37, 19–25. [Google Scholar] [CrossRef]
- Mehlqvist, K.; Larsson, K.; Vajda, V. Linking upper Silurian terrestrial and marine successions—Palynological study from Skåne, Sweden. Rev. Palaeobot. Palynol. 2014, 202, 1–14. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brookfield, M.E.; Catlos, E.J.; Garza, H.K. Reconciling Divergent Ages for the Oldest Recorded Air-Breathing Land Animal, the Millipede, Pneumodesmus newmani Wilson & Anderson, 2004: A Review of the Geology and Ages of the Basal Old Red Sandstone Stonehaven Group (Silurian–Early Devonian), Aberdeenshire, Scotland. Foss. Stud. 2025, 3, 6. https://doi.org/10.3390/fossils3020006
Brookfield ME, Catlos EJ, Garza HK. Reconciling Divergent Ages for the Oldest Recorded Air-Breathing Land Animal, the Millipede, Pneumodesmus newmani Wilson & Anderson, 2004: A Review of the Geology and Ages of the Basal Old Red Sandstone Stonehaven Group (Silurian–Early Devonian), Aberdeenshire, Scotland. Fossil Studies. 2025; 3(2):6. https://doi.org/10.3390/fossils3020006
Chicago/Turabian StyleBrookfield, Michael E., Elizabeth J. Catlos, and Hector K. Garza. 2025. "Reconciling Divergent Ages for the Oldest Recorded Air-Breathing Land Animal, the Millipede, Pneumodesmus newmani Wilson & Anderson, 2004: A Review of the Geology and Ages of the Basal Old Red Sandstone Stonehaven Group (Silurian–Early Devonian), Aberdeenshire, Scotland" Fossil Studies 3, no. 2: 6. https://doi.org/10.3390/fossils3020006
APA StyleBrookfield, M. E., Catlos, E. J., & Garza, H. K. (2025). Reconciling Divergent Ages for the Oldest Recorded Air-Breathing Land Animal, the Millipede, Pneumodesmus newmani Wilson & Anderson, 2004: A Review of the Geology and Ages of the Basal Old Red Sandstone Stonehaven Group (Silurian–Early Devonian), Aberdeenshire, Scotland. Fossil Studies, 3(2), 6. https://doi.org/10.3390/fossils3020006