



Impact of Endogenous Lipids on Mechanical Properties of Wheat Gluten Fractions, Gliadin and Glutenin, under Small, Medium, and Large Deformations †
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Lipid Extraction
2.2.2. Extraction of Gluten Fractions and Sample Preparation
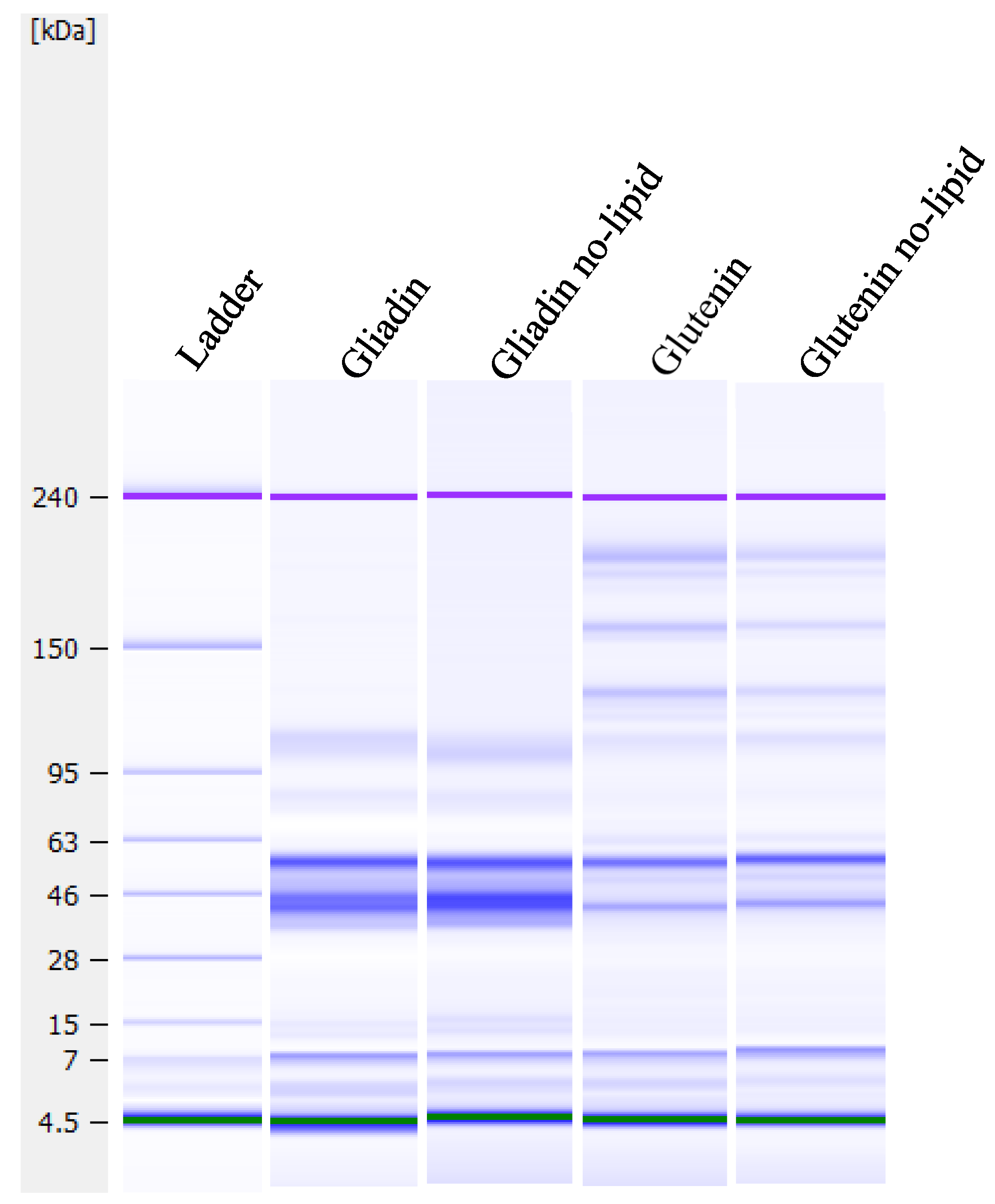
2.2.3. Microfluidic Sodium Dodecyl-Sulfate Polyacrylamide Gel Electrophoresis (SDS-PAGE)
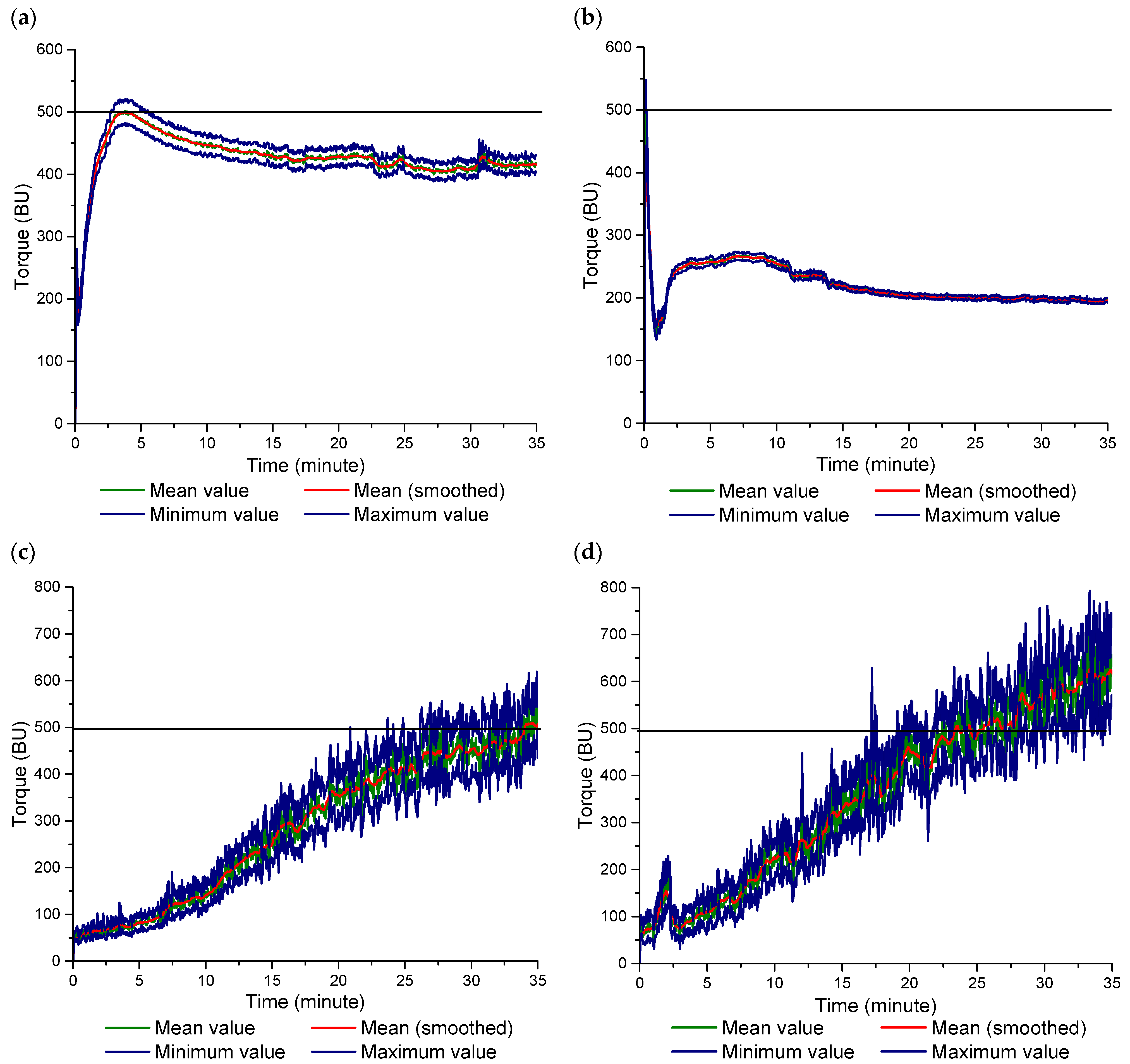
2.2.4. Farinograph Mixing
2.2.5. SAOS, MAOS, and LAOS Properties of Gluten Fractions with and without Endogenous Lipids
2.2.6. Statistical Analysis
3. Results and Discussion
3.1. Microfluidic SDS-PAGE
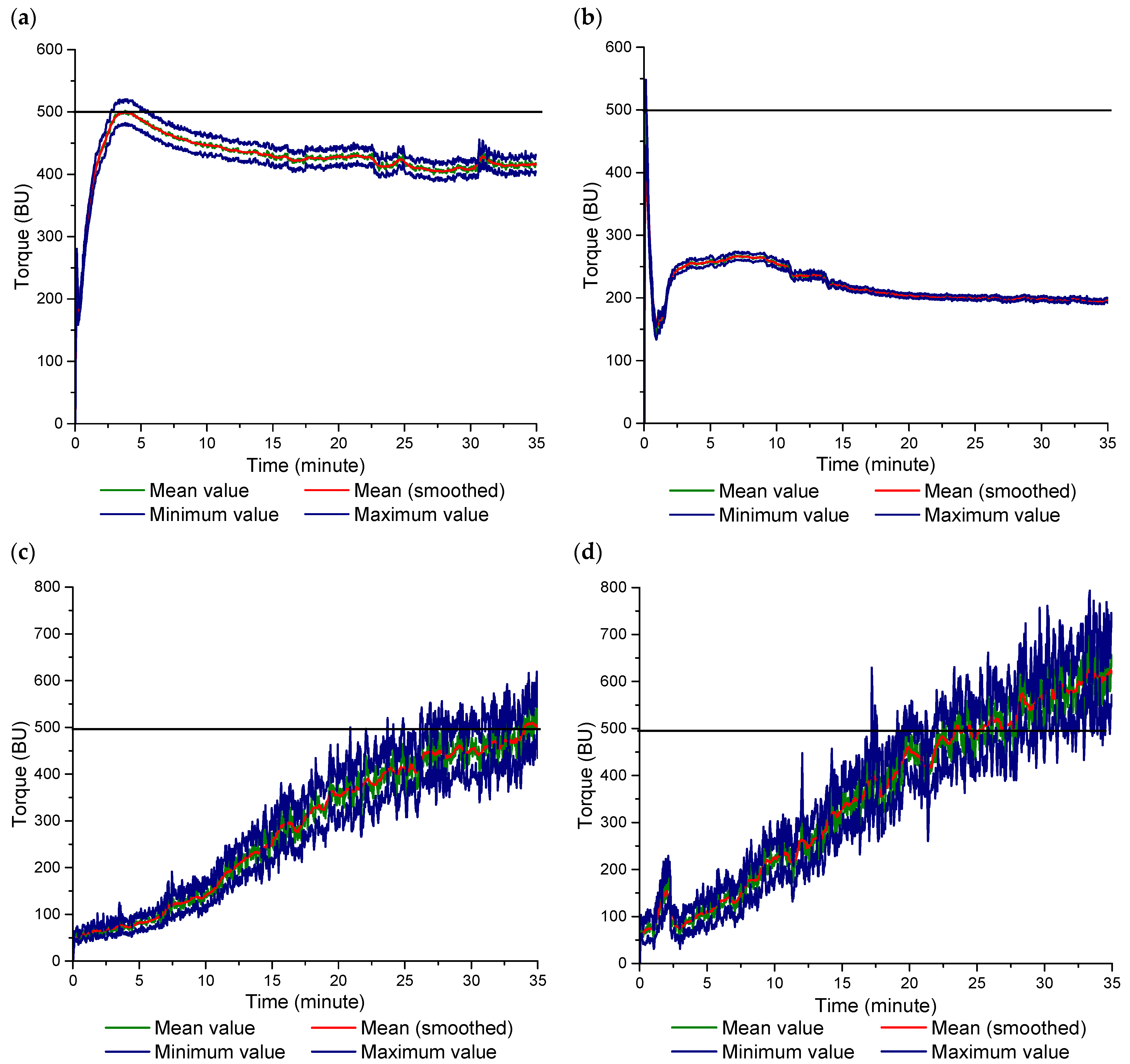
3.2. Farinograph Mixing
3.3. SAOS, MAOS, and LAOS Properties of Gluten Fractions with and without Endogenous Lipids
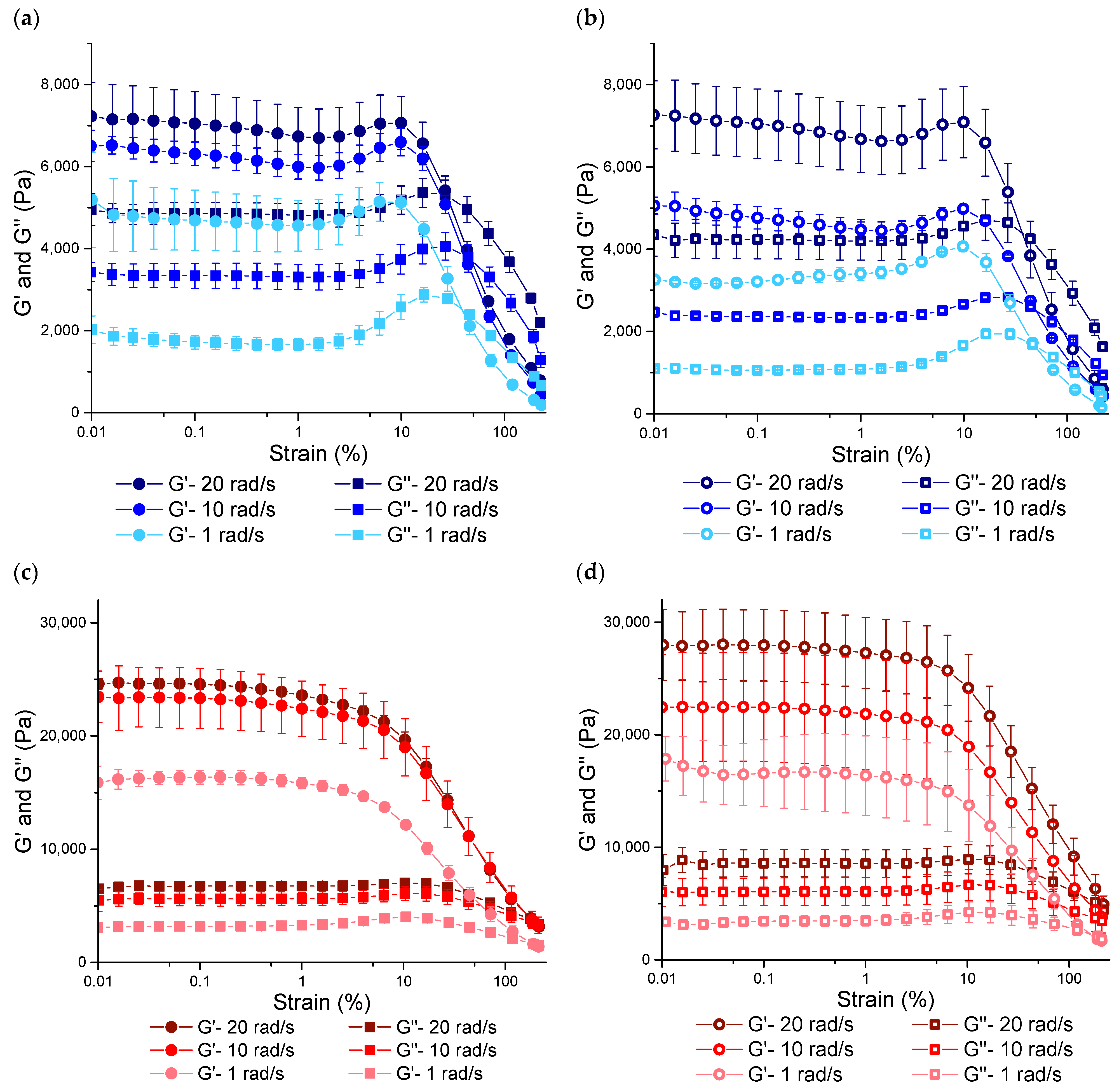
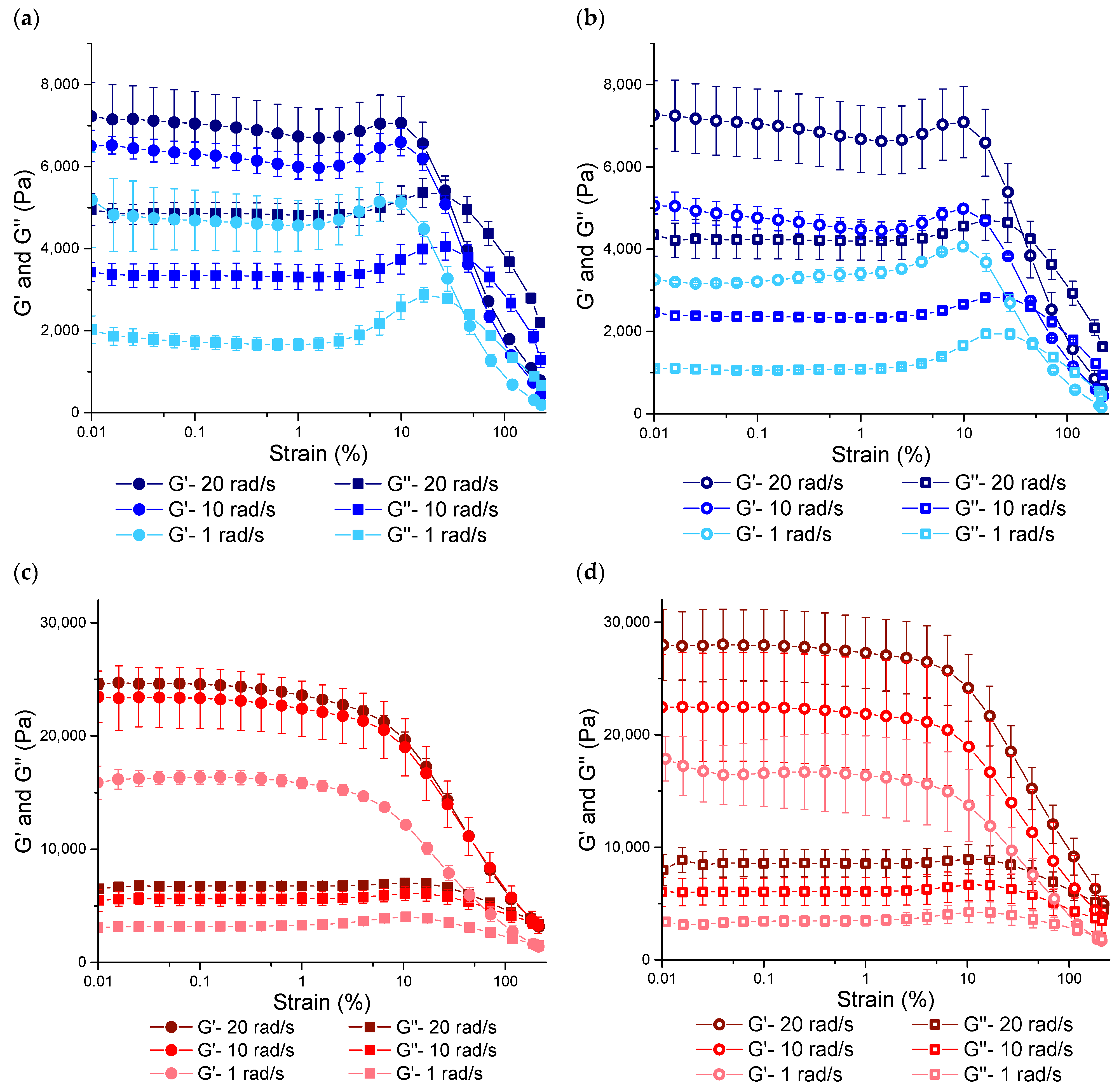
3.3.1. Rheological Properties under SAOS Deformations
3.3.2. Rheological Properties under MAOS Deformations
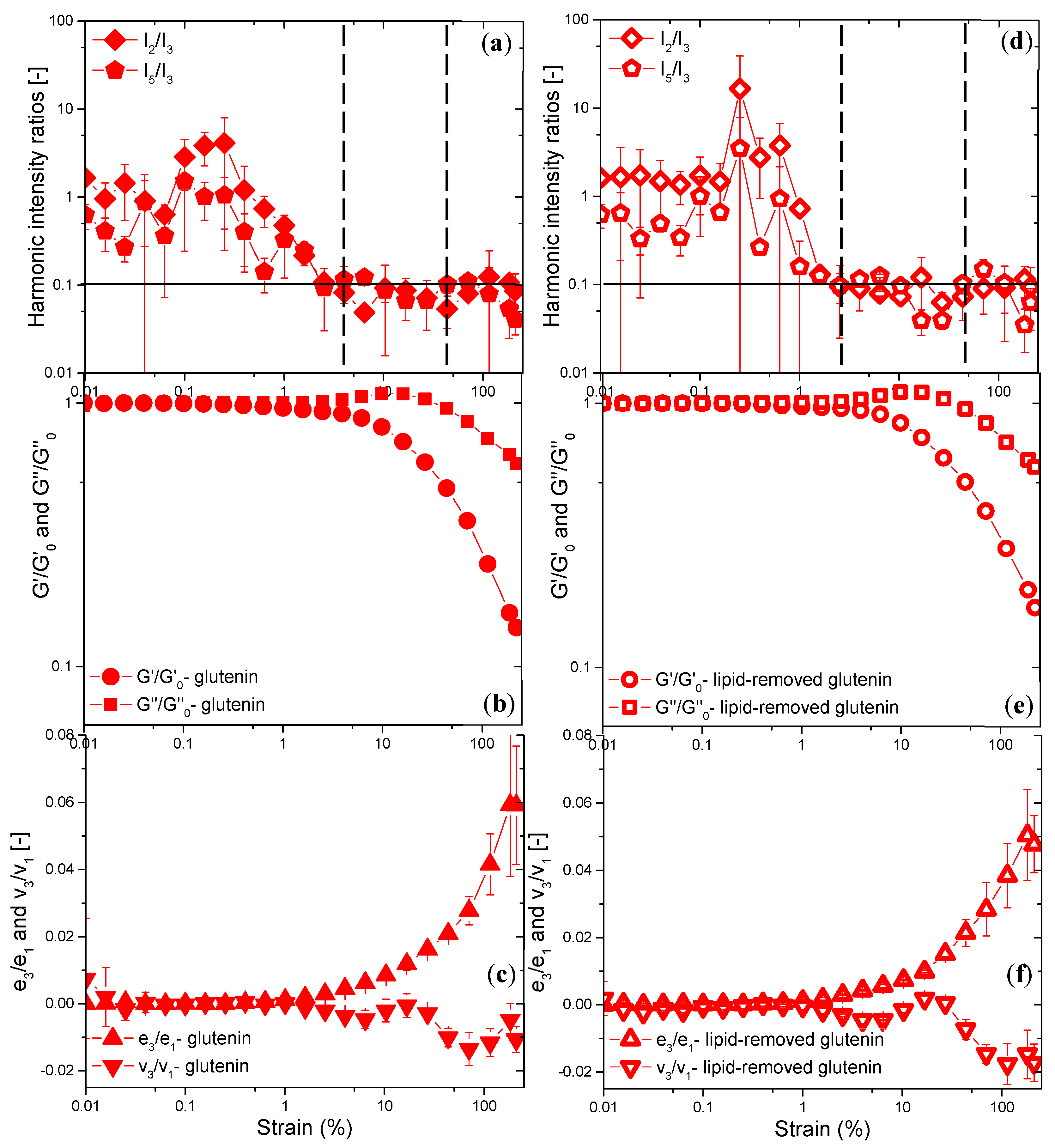
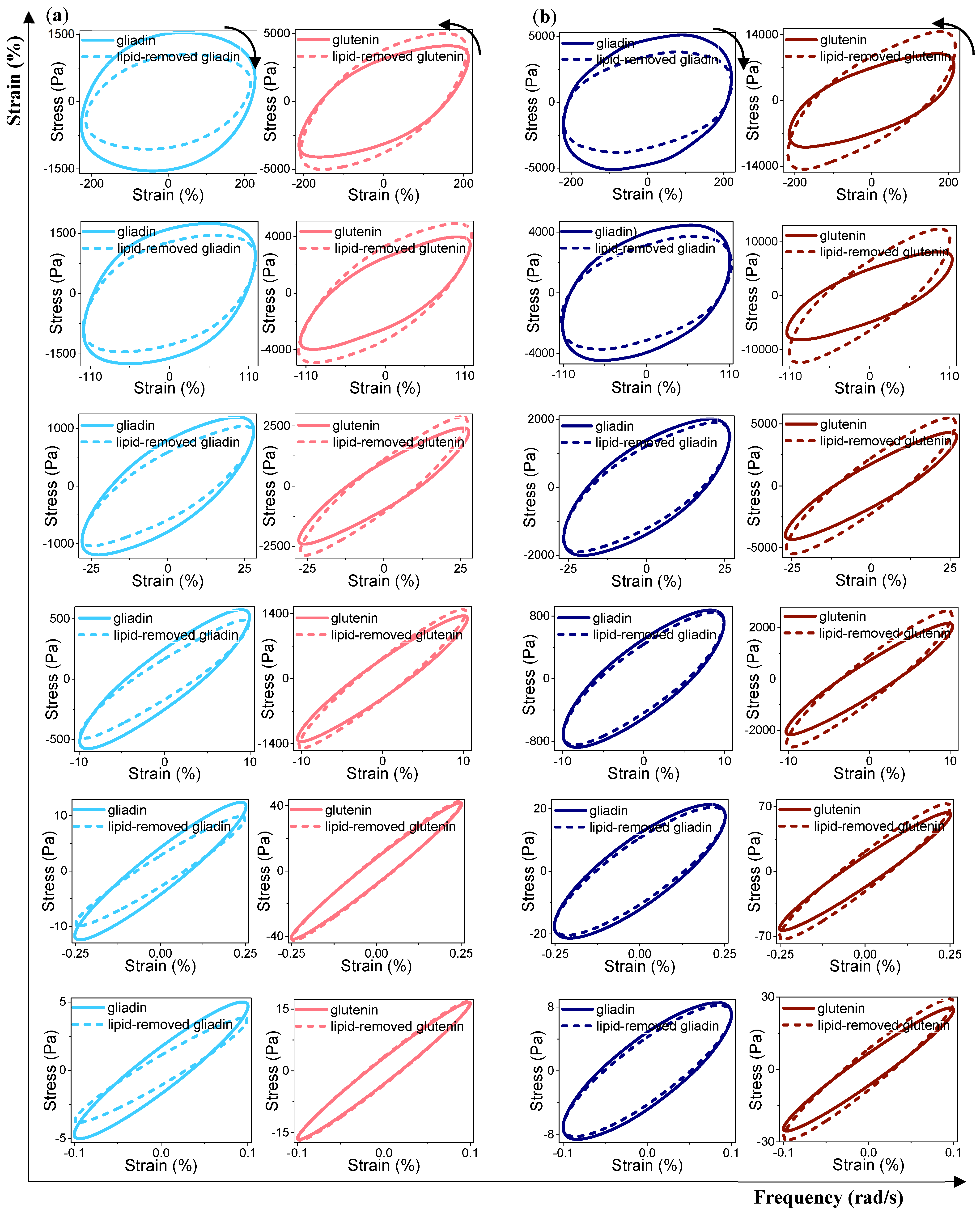
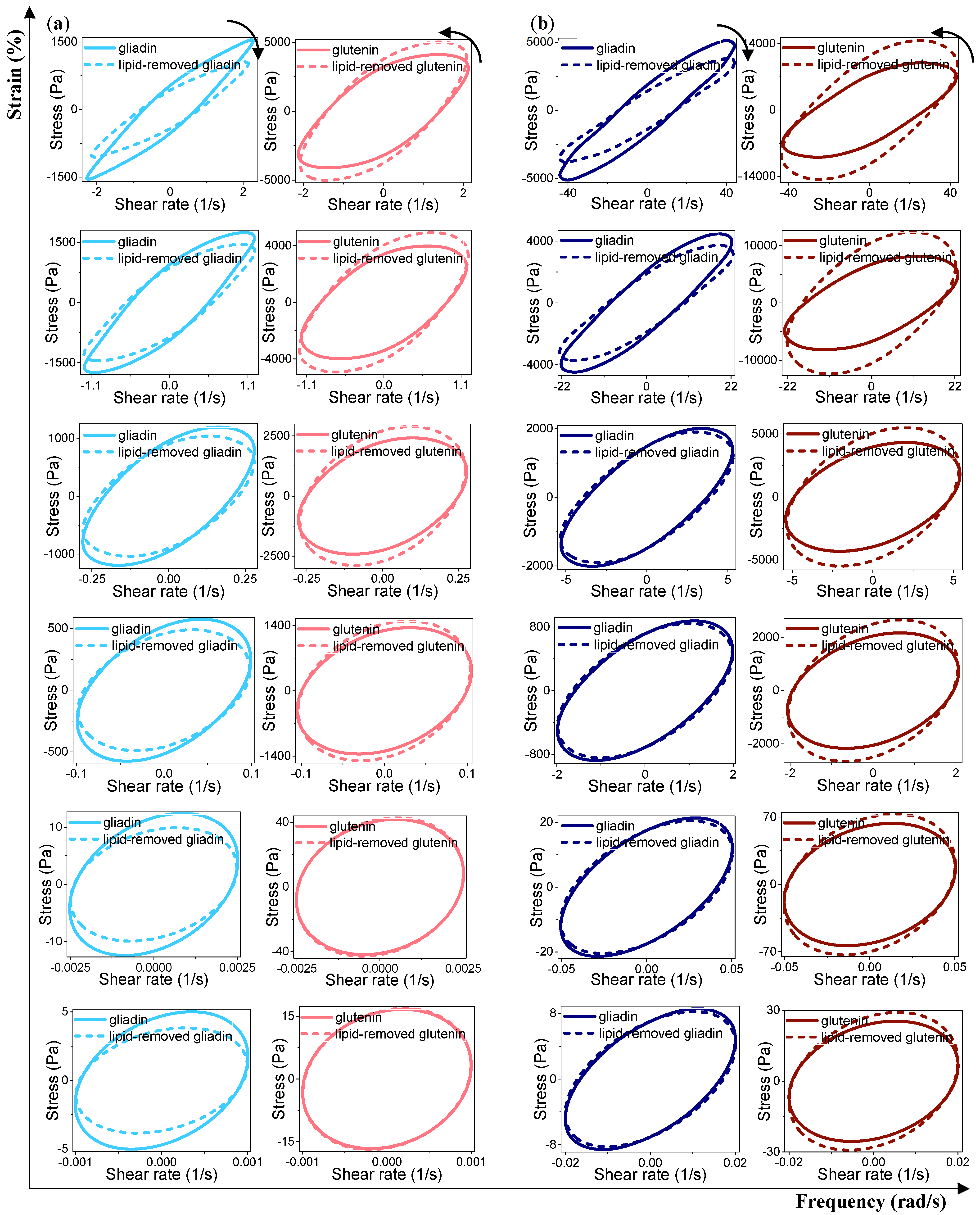
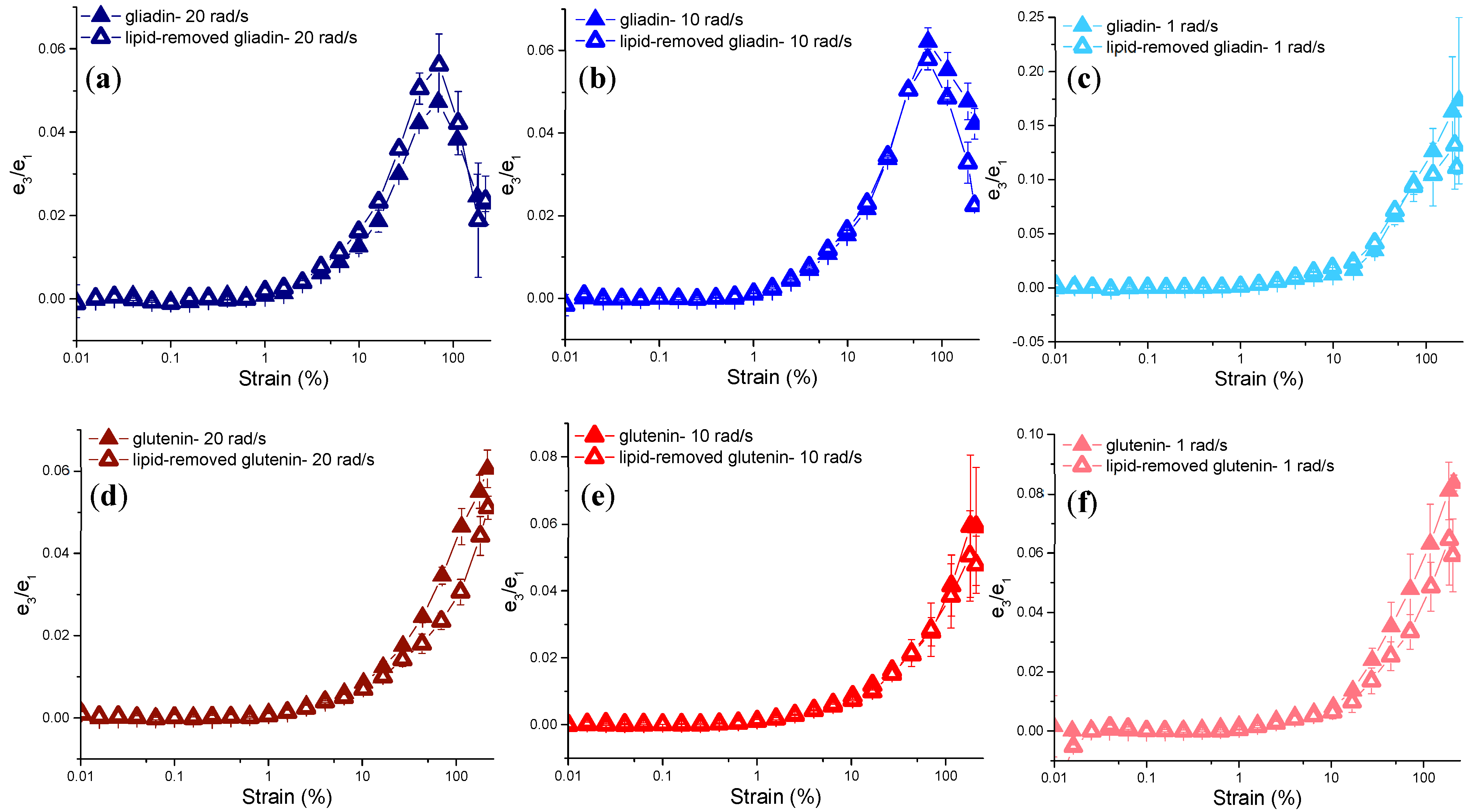
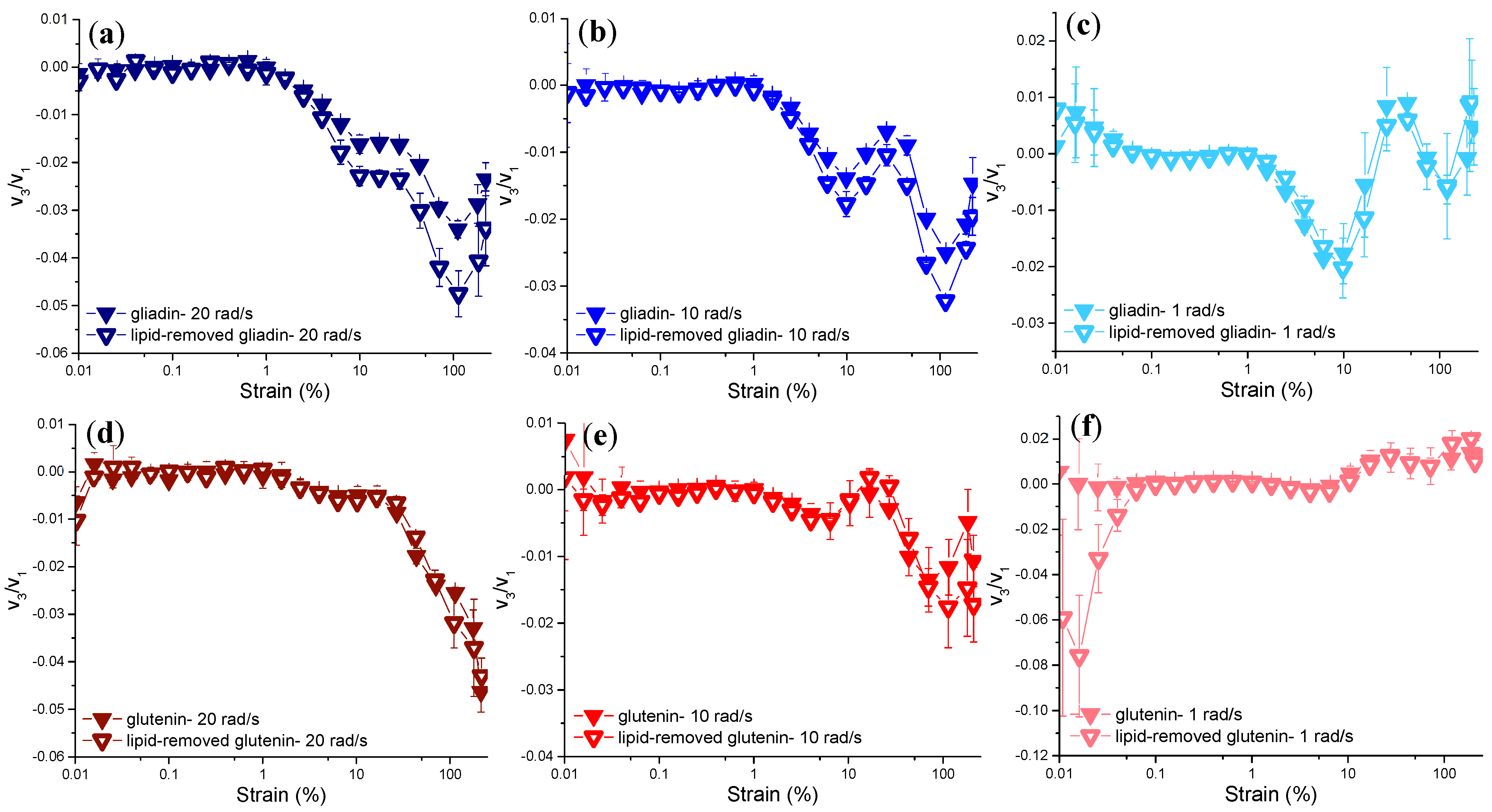
3.3.3. Rheological Properties under LAOS Deformations
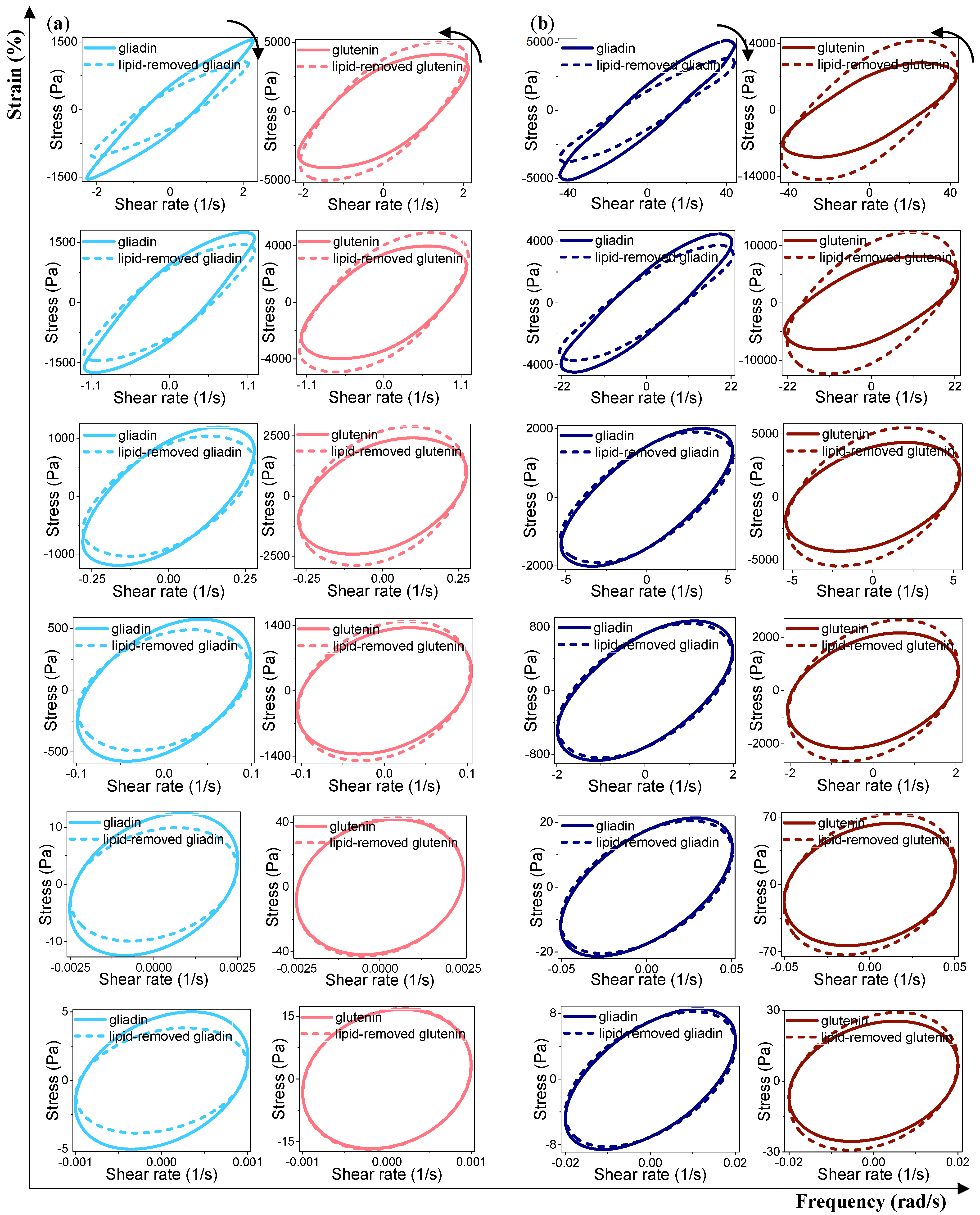
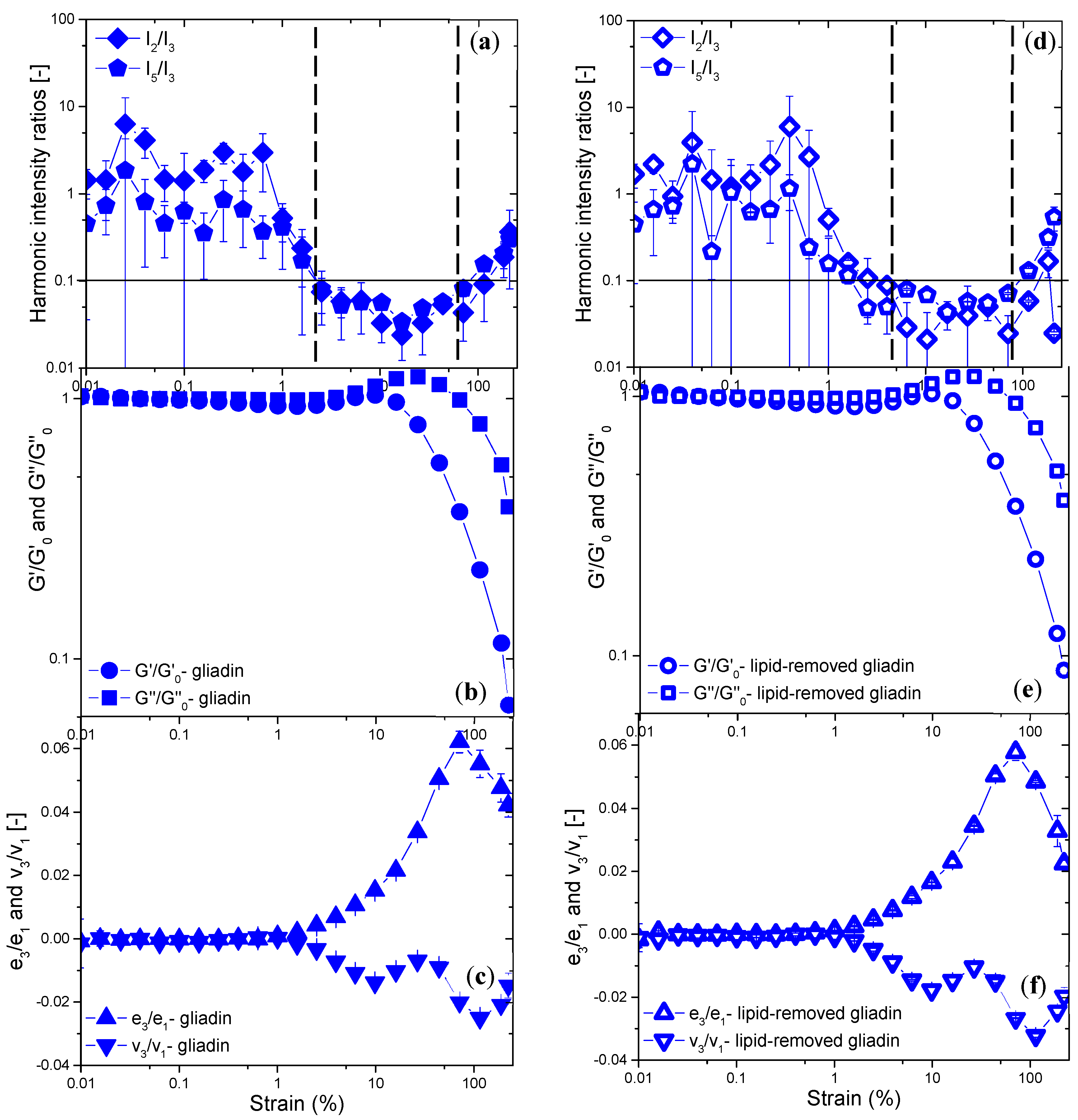
Analysis of the Elastic and Viscous Lissajous–Bowditch Curves
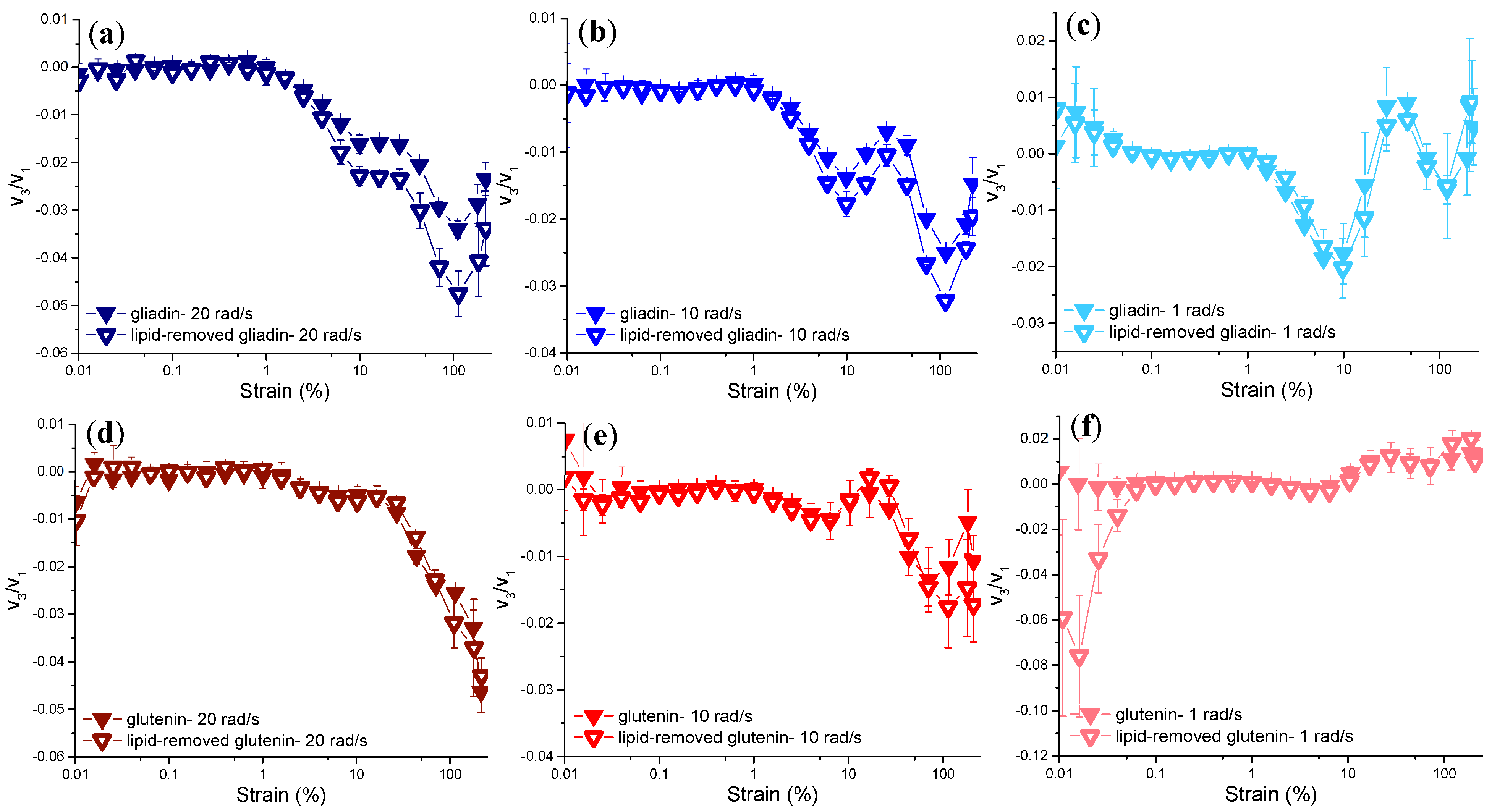
Evaluation of Non-Linear Behavior through the Chebyshev Coefficients
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Melis, S.; Delcour, J.A. Impact of wheat endogenous lipids on the quality of fresh bread: Key terms, concepts, and underlying mechanisms. Compr. Rev. Food Sci. Food Saf. 2020, 19, 3715–3754. [Google Scholar] [CrossRef]
- Pareyt, B.; Finnie, S.M.; Putseys, J.A.; Delcour, J.A. Lipids in bread making: Sources, interactions, and impact on bread quality. J. Cereal Sci. 2011, 54, 266–279. [Google Scholar] [CrossRef]
- McCann, T.H.; Small, D.M.; Batey, I.L.; Wrigley, C.W.; Day, L. Protein-lipid interactions in gluten elucidated using acetic-acid fractionation. Food Chem. 2009, 115, 105–112. [Google Scholar] [CrossRef]
- Yazar, G.; Kokini, J.L.; Smith, B. Effect of endogenous wheat gluten lipids on the non-linear rheological properties of the gluten network. Food Chem. 2022, 367, 130729. [Google Scholar] [CrossRef] [PubMed]
- Yazar, G.; Duvarci, O.; Tavman, S.; Kokini, J.L. LAOS behavior of the two major gluten fractions: Gliadin and glutenin. J. Cereal Sci. 2017, 77, 201–210. [Google Scholar] [CrossRef]
- Gerits, L.R.; Pareyt, B.; Delcour, J.A. Single run HPLC separation coupled to evaporative light scattering detection unravels wheat flour endogenous lipid redistribution during bread dough making. LWT-Food Sci. Technol. 2013, 53, 426–433. [Google Scholar] [CrossRef]
- Sroan, B.S.; Bean, S.R.; MacRitchie, F. Mechanism of gas cell stabilization in bread making. I. The primary gluten–starch matrix. J. Cereal Sci. 2009, 49, 32–40. [Google Scholar] [CrossRef]
- Békés, F.; Zawistowska, U.; Bushuk, W. Protein-lipids complexes in the gliadin fraction. Cereal Chem. 1983, 60, 371–378. [Google Scholar]
- Yazar, G.; Duvarci, O.; Tavman, S.; Kokini, J.L. Effect of mixing on LAOS properties of hard wheat flour dough. J. Food Eng. 2016, 190, 195–204. [Google Scholar] [CrossRef]
- Sroan, B.S.; MacRitchie, F. Mechanism of gas cell stabilization in breadmaking. II. The secondary liquid lamellae. J. Cereal Sci. 2009, 49, 41–46. [Google Scholar] [CrossRef]
- Gerits, L.R.; Pareyt, B.; Delcour, J.A. A lipase based approach for studying the role of wheat lipids in breadmaking. Food Chem. 2014, 156, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Iwaki, S.; Hayakawa, K.; Fu, B.-X.; Otobe, C. Changes in hydrophobic interactions among gluten proteins during dough formation. Processes 2021, 9, 1244. [Google Scholar] [CrossRef]
- Yazar, G.; Kokini, J.L.; Smith, B. Comparison of mixing and non-linear viscoelastic properties of carob germ glutelins and wheat glutenin. Food Hydrocoll. 2023, 143, 108922. [Google Scholar] [CrossRef]
- Osborne, T.B. The Proteins of the Wheat Kernel (Monograph); Carnegie Institute, Washington: Washington, DC, USA, 1907. [Google Scholar]
- Smith, B.M.; Bean, S.R.; Schober, T.J.; Tilley, M.; Herald, T.J.; Aramouni, F.M. Composition and molecular weight distribution of carob germ protein fractions. J. Agric. Food Chem. 2010, 58, 7794–7800. [Google Scholar] [CrossRef] [PubMed]
- Agilent Technologies. Agilent Protein 230 Kit Quick Start Guide. Agilent Technologies, Inc. 2016. Available online: https://www.agilent.com/cs/library/usermanuals/public/Protein-230_Assay_QSG_RevC.pdf. (accessed on 20 March 2024).
- AACC International. Approved Methods of Analysis, 11th ed.; AACC International: St. Paul, MN, USA, 2010. [Google Scholar]
- Ewoldt, R.H.; Hosoi, A.E.; McKinley, G.H. New measures for characterizing nonlinear viscoelasticity in large amplitude oscillatory shear. J. Rheol. 2008, 52, 1427–1458. [Google Scholar] [CrossRef]
- Wieser, H. Chemistry of gluten proteins. Food Microbiol. 2007, 24, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Wieser, H.; Koehler, P.; Scherf, K.A. Chemistry of wheat gluten proteins: Qualitative composition. Cereal Chem. 2023, 100, 23–35. [Google Scholar] [CrossRef]
- Chaudhary, N.; Virdi, A.S.; Dangi, P.; Khatkar, B.S.; Mohanty, A.K.; Singh, N. Protein, thermal and functional properties of α-, γ- and ω-gliadins of wheat and their effect on bread making characteristics. Food Hydrocoll. 2022, 124, 107212. [Google Scholar] [CrossRef]
- Shewry, P.R.; Tatham, A.S.; Forde, J.; Kreis, M.; Miflin, B.J. The classification and nomenclature of wheat gluten proteins: A reassessment. J. Cereal Sci. 1986, 4, 97–106. [Google Scholar] [CrossRef]
- Wieser, H.; Bushuk, W.; MacRitchie, F. The polymeric glutenins. In Gliadin and Glutenin: The Unique Balance of Wheat Quality; Wrigley, C., Bekes, F., Bushuk, W., Eds.; AACC International: St. Paul, MN, USA, 2006; pp. 213–240. [Google Scholar]
- Bock, J.E. The Farinograph: Understanding Farinograph curves. In The Farinograph Handbook—Advances in Technology, Science, and Applications, 4th ed.; Bock, J.E., Don, C., Eds.; Woodhead Publishing: Sawston, UK, 2022; pp. 33–41. [Google Scholar]
- Van Bockstaele, F.; De Leyn, I.; Eeckhout, M.; Dewettinck, K. Rheological properties of wheat flour dough and the relationship with bread volume. I. Creep-recovery measurements. Cereal Chem. 2008, 85, 753–761. [Google Scholar] [CrossRef]
- Papantoniou, E.; Hammond, E.W.; Scriven, F.; Gordon, M.H.; Schofield, J.D. Effects of endogenous flour lipids on the quality of short-dough biscuit. J. Sci. Food Agric. 2004, 84, 1371–1380. [Google Scholar] [CrossRef]
- Pichot, R.; Watson, R.L.; Norton, I.T. Phospholipids at the interface: Current trends and challenges. Int. J. Mol. Sci. 2013, 14, 11767–11794. [Google Scholar] [CrossRef]
- Monteiro, A.M.F.; Arêas, E.P.G.; Schröder, A.; Fa, N. Gliadin effect on fluctuation properties of phospholipid giant vesicles. Colloids Surf. B Biointerfaces 2004, 34, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Georget, D.M.; Belton, P.S. Effects of temperature and water content on the secondary structure of wheat gluten studied by FTIR spectroscopy. Biomacromolecules 2006, 7, 469–475. [Google Scholar] [CrossRef]
- Pézolet, M.; Bonenfant, S.; Dousseau, F.; Popineau, Y. Conformation of wheat gluten proteins: Comparison between functional and solution states as determined by infrared spectroscopy. FEBS Lett. 1992, 299, 247–250. [Google Scholar] [CrossRef]
- Wellner, N.; Belton, P.S.; Tatham, A.S. Fourier transform IR spectroscopic study of hydration-induced structure changes in the solid state of ω-gliadins. Biochem. J. 1996, 319, 741–747. [Google Scholar] [CrossRef]
- Mejia, C.D.; Mauer, L.J.; Hamaker, B.R. Similarities and differences in secondary structure of viscoelastic polymers of maize α-zein and wheat gluten proteins. J. Cereal Sci. 2007, 45, 353–359. [Google Scholar] [CrossRef]
- Wang, Y.; Belton, P.S.; Bridon, H.; Garanger, E.; Wellner, N.; Parker, M.L.; Grant, A.; Feillet, P.; Noel, T.R. Physicochemical studies of caroubin: A gluten-like protein. J. Agric. Food Chem. 2001, 49, 3414–3419. [Google Scholar] [CrossRef] [PubMed]
- Belton, P.S.; Colquhoun, I.J.; Grant, A.; Wellner, N.; Field, J.M.; Shewry, P.R.; Tatham, A.S. FTIR and NMR studies on the hydration of a high-Mr subunit of glutenin. Int. J. Biol. Macromol. 1995, 17, 74–80. [Google Scholar] [CrossRef]
- Fevzioglu, M.; Ozturk, O.K.; Hamaker, B.R.; Campanella, O.H. Quantitative approach to study secondary structure of proteins by FT-IR spectroscopy, using a model wheat gluten system. Int. J. Biol. Macromol. 2020, 164, 2753–2760. [Google Scholar] [CrossRef]
- Dewan, A.; Khatkar, B.S.; Bangar, S.P.; Chaudhary, V.; Lorenzo, J.M. Infrared spectroscopy and microstructural assessment of dough with varying wheat gluten fractions. Food Anal. Methods 2022, 15, 2821–2829. [Google Scholar] [CrossRef]
- Yazar, G.; Caglar Duvarci, O.; Yildirim Erturk, M.; Kokini, J.L. LAOS (large amplitude oscillatory shear) applications for semisolid foods. In Rheology of Semisolid Foods; Food Engineering Series; Joyner, H., Ed.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 97–131. [Google Scholar]
- Barak, S.; Mudgil, D.; Khatkar, B.S. Influence of gliadin and glutenin fractions on rheological, pasting, and textural properties of dough. Int. J. Food Prop. 2014, 17, 1428–1438. [Google Scholar] [CrossRef]
- Song, H.Y.; Hyun, K. First-harmonic intrinsic nonlinearity of model polymer solutions in medium amplitude oscillatory shear (MAOS). Korea-Aust. Rheol. J. 2019, 31, 1–13. [Google Scholar] [CrossRef]
- Erturk, M.Y.; Minh, A.N.; Kokini, J.L. Advances in large amplitude oscillatory shear rheology of food materials. Front. Food Sci. Technol. 2023, 3, 1130165. [Google Scholar] [CrossRef]
- Ewoldt, R.; Bharadwaj, N.A. Low-dimensional intrinsic material functions for nonlinear viscoelasticity. Rheologica Acta 2013, 52, 201–219. [Google Scholar] [CrossRef]
- Singh, P.K.; Soulages, J.M.; Ewoldt, R. Frequency-sweep medium amplitude oscillatory shear (MAOS). J. Rheol. 2018, 62, 277–293. [Google Scholar] [CrossRef]
- Hyun, K.; Kim, S.H.; Ahn, K.H.; Lee, S.J. Large amplitude oscillatory shear as a way to classify the complex fluids. J. Non-Newton. Fluid Mech. 2002, 107, 51–65. [Google Scholar] [CrossRef]
- Ewoldt, R.H.; Clasen, C.; Hosoi, A.E.; McKinley, G.H. Rheological fingerprinting of gastropod pedal mucus and synthetic complex fluids for biomimicking adhesive locomotion. Soft Matter 2007, 3, 634–643. [Google Scholar] [CrossRef] [PubMed]
- Macias-Rodriguez, B.A.; Ewoldt, R.H.; Marangoni, A.G. Nonlinear viscoelasticity of fat crystal networks. Rheol. Acta 2018, 57, 251–266. [Google Scholar] [CrossRef]
- Kokelaar, J.J.; van Vliet, T.; Prins, A. Strain hardening properties and extensibility of flour and gluten doughs in relation to breadmaking performance. J. Cereal Sci. 1996, 24, 199–214. [Google Scholar] [CrossRef]
- Menjivar, J.A. Fundamental aspects of dough rheology. In Dough Rheology and Baked Product Texture; Faridi, H., Faubion, J.M., Eds.; AVI Publishing: New York, NY, USA, 1990; pp. 1–29. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SAOS, MAOS, and LAOS Strains (%) | tanδ | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 20 rad/s | 10 rad/s | 1 rad/s | ||||||||||
| Gliadin | Lipid-Removed Gliadin | Glutenin | Lipid-Removed Glutenin | Gliadin | Lipid-Removed Gliadin | Glutenin | Lipid-Removed Glutenin | Gliadin | Lipid-Removed Gliadin | Glutenin | Lipid-Removed Glutenin | |
| 0.1 | 0.69 ± 0.07 a | 0.60 ± 0.00 a | 0.27 ± 0.00 c | 0.30 ± 0.00 d | 0.52 ± 0.03 a | 0.49 ± 0.02 a | 0.24 ± 0.00 c | 0.27 ± 0.01 d | 0.36 ± 0.02 a | 0.33 ± 0.01 a | 0.19 ± 0.00 c | 0.20 ± 0.01 c |
| 0.25 | 0.70 ± 0.07 a | 0.60 ± 0.00 a | 0.27 ± 0.00 c | 0.30 ± 0.00 d | 0.53 ± 0.03 a | 0.50 ± 0.02 a | 0.24 ± 0.00 c | 0.27 ± 0.01 d | 0.36 ± 0.02 a | 0.32 ± 0.01 a | 0.19 ± 0.00 c | 0.20 ± 0.01 c |
| 10 | 0.73 ± 0.05 a | 0.64 ± 0.01 a | 0.35 ± 0.01 c | 0.36 ± 0.00 c | 0.56 ± 0.03 a | 0.53 ± 0.00 a | 0.32 ± 0.01 c | 0.35 ± 0.02 c | 0.50 ± 0.04 a | 0.40 ± 0.00 a | 0.33 ± 0.01 c | 0.30 ± 0.00 c |
| 25 | 0.98 ± 0.05 a | 0.86 ± 0.02 b | 0.46 ± 0.01 c | 0.45 ± 0.01 c | 0.79 ± 0.03 a | 0.73 ± 0.00 a | 0.41 ± 0.01 c | 0.45 ± 0.04 c | 0.85 ± 0.08 a | 0.71 ± 0.00 a | 0.44 ± 0.04 c | 0.40 ± 0.01 c |
| 110 | 2.05 ± 0.09 a | 1.89 ± 0.21 a | 0.79 ± 0.01 c | 0.66 ± 0.05 d | 1.90 ± 0.02 a | 1.56 ± 0.02 b | 0.72 ± 0.05 c | 0.71 ± 0.11 c | 1.99 ± 0.21 a | 1.72 ± 0.07 a | 0.79 ± 0.13 c | 0.84 ± 0.09 c |
| 200 | 2.79 ± 0.14 a | 2.83 ± 0.60 a | 1.07 ± 0.04 c | 0.90 ± 0.07 d | 3.05 ± 0.31 a | 2.20 ± 0.13 b | 1.01 ± 0.11 c | 0.96 ± 0.14 c | 3.50 ± 0.45 a | 2.91 ± 0.56 a | 1.07 ± 0.18 c | 1.05 ± 0.07 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yazar, G.; Kokini, J.L.; Smith, B. Impact of Endogenous Lipids on Mechanical Properties of Wheat Gluten Fractions, Gliadin and Glutenin, under Small, Medium, and Large Deformations. Lipidology 2024, 1, 30-51. https://doi.org/10.3390/lipidology1010004
Yazar G, Kokini JL, Smith B. Impact of Endogenous Lipids on Mechanical Properties of Wheat Gluten Fractions, Gliadin and Glutenin, under Small, Medium, and Large Deformations. Lipidology. 2024; 1(1):30-51. https://doi.org/10.3390/lipidology1010004
Chicago/Turabian StyleYazar, Gamze, Jozef L. Kokini, and Brennan Smith. 2024. "Impact of Endogenous Lipids on Mechanical Properties of Wheat Gluten Fractions, Gliadin and Glutenin, under Small, Medium, and Large Deformations" Lipidology 1, no. 1: 30-51. https://doi.org/10.3390/lipidology1010004