Phytochemicals of Apple Pomace as Prospect Bio-Fungicide Agents against Mycotoxigenic Fungal Species—In Vitro Experiments

 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
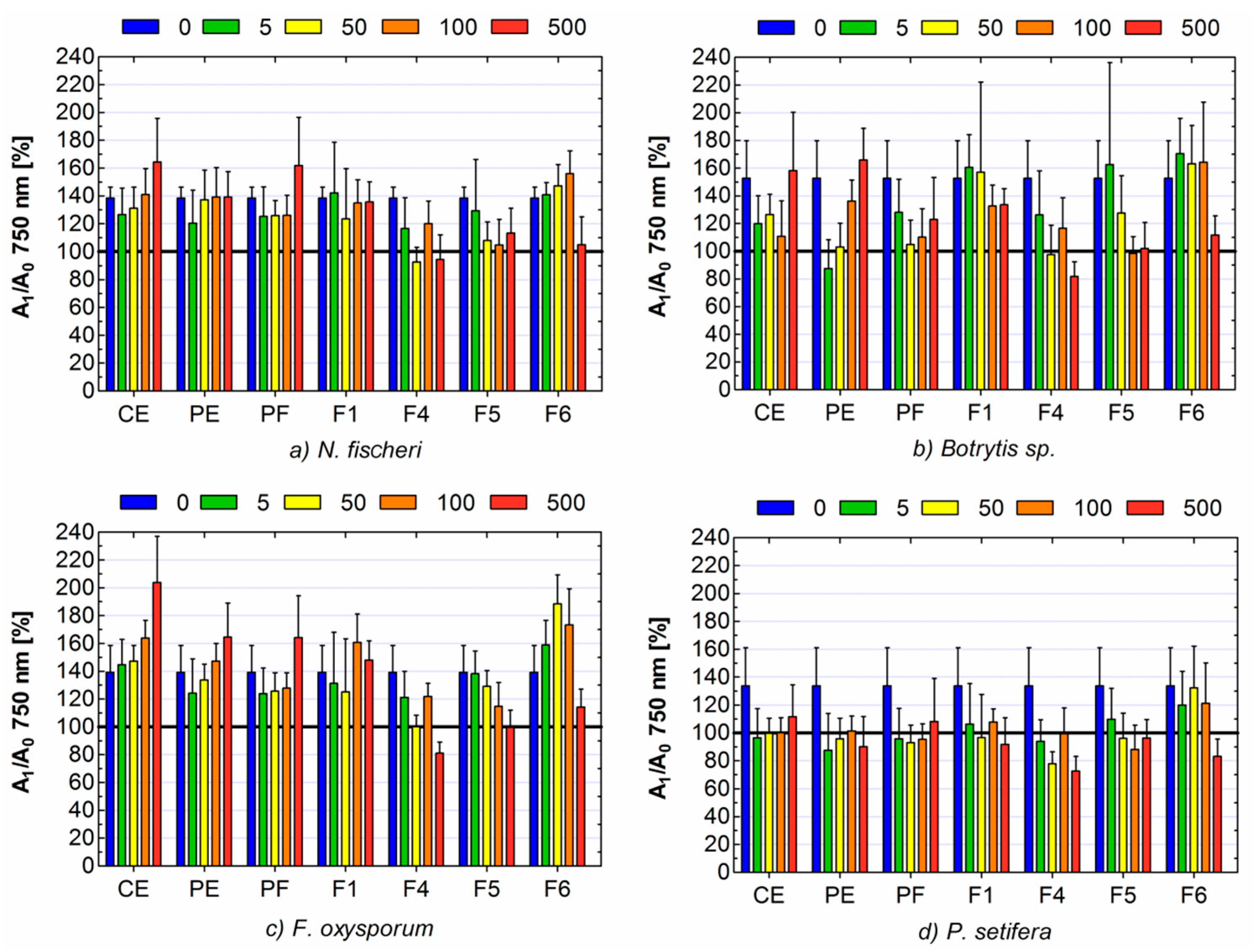
2. Results and Discussion
3. Materials and Methods
3.1. Materials
3.1.1. Apple Pomace
3.1.2. Fungal Strains and Culture Conditions
3.2. Extract Preparation and Fractionation
3.3. Phytochemical Analysis
3.3.1. Identification and Quantification of Individual Compounds in Crude, Purified Extracts and Its Fractions
3.3.2. Isolation Process of Pinnatifidanoside D
3.3.3. Nuclear Magnetic Resonance (NMR) Analysis
3.3.4. Antioxidant Activity
3.3.5. Antifungal Activity
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Santana-Méridas, O.; González-Coloma, A.; Sánchez-Vioque, R. Agricultural Residues as a Source of Bioactive Natural Products. Phytochem. Rev. 2012, 11, 447–466. [Google Scholar] [CrossRef]
- Heidorn, E.; Utvik, K.; Gengler, C.; Alati, K.; Collet, D.; Attivissimo, V.; Colantonio, M. Agriculture, Forestry and Fishery Statistics 2017 Edition; Forti, R., Ed.; European Commission: Luxemburg, Belgium, 2017. [Google Scholar] [CrossRef]
- Górnaś, P.; Mišina, I.; Olšteine, A.; Krasnova, I.; Pugajeva, I.; Lācis, G.; Siger, A.; Michalak, M.; Soliven, A.; Segliņa, A. Phenolic Compounds in Different Fruit Parts of Crab Apple: Dihydrochalcones as Promising Quality Markers of Industrial Apple Pomace By-Products. Ind. Crops Prod. 2015, 74, 607–612. [Google Scholar] [CrossRef]
- Waldbauer, K.; McKinnon, R.; Kopp, B. Apple Pomace as Potential Source of Natural Active Compounds. Planta Med. 2017, 83, 994–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhushan, S.; Kalia, K.; Sharma, M.; Singh, B.; Ahuja, P.S. Processing of Apple Pomace for Bioactive Molecules. Crit. Rev. Biotechnol. 2008, 28, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Rabaneda, F.; Jáuregui, O.; Lamuela-Raventós, R.M.; Viladomat, F.; Bastida, J.; Codina, C. Qualitative Analysis of Phenolic Compounds in Apple Pomace Using Liquid Chromatography Coupled to Mass Spectrometry in Tandem Mode. Rapid Commun. Mass Spectrom. 2004, 18, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Kołodziejczyk, K.; Sójka, M.; Abadias, M.; Viñas, I.; Guyot, S.; Baron, A. Polyphenol Composition, Antioxidant Capacity, and Antimicrobial Activity of the Extracts Obtained from Industrial Sour Cherry Pomace. Ind. Crops Prod. 2013, 51, 279–288. [Google Scholar] [CrossRef]
- Shim, S.-H.; Jo, S.-J.; Kim, J.-C.; Choi, G.-J. Control Efficacy of Phloretin Isolated from Apple Fruits Against Several Plant Diseases. Plant Pathol. J. 2010, 26, 280–285. [Google Scholar] [CrossRef] [Green Version]
- Sanzani, S.M.; Schena, L.; Nigro, F.; de Girolamo, A.; Ippolito, A. Effect of Quercetin and Umbelliferone on the Transcript Level of Penicillium expansum Genes Involved in Patulin Biosynthesis. Eur. J. Plant Pathol. 2009, 125, 223–233. [Google Scholar] [CrossRef]
- Frąc, M.; Gryta, A.; Oszust, K.; Kotowicz, N. Fast and Accurate Microplate Method (Biolog MT2) for Detection of Fusarium Fungicides Resistance/Sensitivity. Front. Microbiol. 2016, 7, 1–16. [Google Scholar] [CrossRef]
- Becher, R.; Hettwer, U.; Karlovsky, P.; Deising, H.B.; Wirsel, S.G.R. Adaptation of Fusarium graminearum to Tebuconazole Yielded Descendants Diverging for Levels of Fitness, Fungicide Resistance, Virulence, and Mycotoxin Production. Phytopathology 2010, 100, 444–453. [Google Scholar] [CrossRef]
- Lourenço, R.M.D.C.; Melo, P.; da Silva, S.; de Almeida, A.B.A. Flavonoids as Antifungal Agents. In Antifungal Metabolites from Plants; Razzaghi-Abyaneh, M., Rai, M., Eds.; Springer: Berlin, Germany, 2013; pp. 1–469. [Google Scholar]
- Upasani, M.L.; Gurjar, G.S.; Kadoo, N.Y.; Gupta, V.S. Dynamics of Colonization and Expression of Pathogenicity Related Genes in Fusarium oxysporum f. sp. ciceri during Chickpea Vascular Wilt Disease Progression. PLoS ONE 2016, 11, e0156490. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-C.; Li, L.C.; Cai, B.; Cai, L.T.; Chen, X.J.; Yu, Z.H.; Zhang, C.Q. Metabolic Phenotype Characterization of Botrytis cinerea, the Causal Agent of Gray Mold. Front. Microbiol. 2018, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Oszust, K.; Panek, J.; Pertile, G.; Siczek, A.; Oleszek, M.; Frąc, M. Metabolic and Genetic Properties of Petriella setifera Precultured on Waste. Front. Microbiol. 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kwaśna, H.; Łakomy, P.; Łabędzki, A. Morphological characteristics and DNA sequence analysis of Petriella setifera and Oidiodendron setiferum from twigs of diseased oak. Acta Mycol. 2005, 40, 267–275. [Google Scholar] [CrossRef]
- Schwarze, F.W. Wood Decay Under the Microscope. Fungal Biol. Rev. 2007, 21, 133–170. [Google Scholar] [CrossRef]
- Pertile, G.; Panek, J.; Oszust, K.; Siczek, A.; Frąc, M. Intraspecific Functional and Genetic Diversity of Petriella setifera. PeerJ 2018, 6, e4420. [Google Scholar] [CrossRef] [PubMed]
- Frąc, M.; Jezierska-Tys, S.; Yaguchi, T. Occurrence, Detection, and Molecular and Metabolic Characterization of Heat-Resistant Fungi in Soils and Plants and Their Risk to Human Health. Adv. Agron. 2015, 132, 161–204. [Google Scholar] [CrossRef]
- Sanzani, S.M.; Girolamo, A.; Schena, L.; Solfrizzo, M.; Ippolito, A.; Visconti, A. Control of Penicillium expansum and Patulin Accumulation on Apples by Quercetin and Umbelliferone. Eur. Food Res. Technol. 2009, 228, 381–389. [Google Scholar] [CrossRef]
- Steinkellner, S.; Mammerler, R. Effect of Flavonoids on the Development of Fusarium oxysporum f. sp. lycopersici. J. Plant Interact. 2007, 2, 17–23. [Google Scholar] [CrossRef]
- Naseer, R.; Sultana, B.; Khan, M.Z.; Naseer, D.; Nigam, P. Utilization of Waste Fruit-Peels to Inhibit Aflatoxins Synthesis by Aspergillus flavus: A Biotreatment of Rice for Safer Storage. Bioresour. Technol. 2014, 172, 423–428. [Google Scholar] [CrossRef]
- Parvez, M.M.; Tomita-Yokotani, K.; Fujii, Y.; Konishi, T.; Iwashina, T. Effects of Quercetin and Its Seven Derivatives on the Growth of Arabidopsis thaliana and Neurospora crassa. Biochem. Syst. Ecol. 2004, 32, 631–635. [Google Scholar] [CrossRef]
- Dick, A.J.; Redden, P.R.; DeMarco, A.C.; Lidster, P.D.; Grindley, T.B. Flavonoid Glycosides of Spartan Apple Peel. J. Agric. Food Chem. 1987, 35, 529–531. [Google Scholar] [CrossRef]
- Lu, Y.; Foo, L.Y. Identification and Quantification of Major Polyphenols in Apple Pomace. Food Chem. 1997, 59, 187–194. [Google Scholar] [CrossRef]
- Vasantha Rupasinghe, H.P.; Kean, C. Polyphenol Concentrations in Apple Processing By-Products Determined Using Electrospray Ionization Mass Spectrometry. Can. J. Plant Sci. 2008, 88, 759–762. [Google Scholar] [CrossRef]
- Li, L.Z.; Gao, P.Y.; Song, S.J.; Yuan, Y.Q.; Liu, C.T.; Huang, X.X.; Liu, Q.B. Monoterpenes and Flavones from the Leaves of Crataegus pinnatifida with Anticoagulant Activities. J. Funct. Foods 2015, 12, 237–245. [Google Scholar] [CrossRef]
- Oleszek, W.; Lee, C.Y.; Jaworski, A.W.; Price, K.R. Identification of Some Phenolic Compounds in Apples. J. Agric. Food Chem. 1988, 36, 430–432. [Google Scholar] [CrossRef]
- Walker, A.A. Note on the Polyphenol Content of Ripening Apples. N. Z. J. Sci. 1963, 6, 492–494. [Google Scholar]
- Ingle, M.; Hyde, J. The Effect of Bruising on Discoloration and Concentration of Phenolic Compounds in Apple Tissue. Proc. Am. Soc. Hortic. Sci. 1968, 93, 738–745. [Google Scholar]
- Vamos-Vigyazo, L.; Gajzago, I. Studies on the Enzymatic Browning and the Polyphenol–polyphenol Oxidase Complex of Apple Cultivars. Confructa 1976, 21, 24–31. [Google Scholar]
- Burda, S.; Oleszek, W.; Lee, C.Y. Phenolic Compounds and Their Changes in Apples during Maturation and Cold Storage. J. Agric. Food Chem. 1990, 38, 945–948. [Google Scholar] [CrossRef]
- Gadkari, P.V.; Kadimi, U.S.; Balaraman, M. Catechin Concentrates of Garden Tea Leaves (Camellia Sinensis L.): Extraction/Isolation and Evaluation of Chemical Composition. J. Sci. Food Agric. 2014, 94, 2921–2928. [Google Scholar] [CrossRef] [PubMed]
- Heim, K.E.; Tagliaferro, A.R.; Bobilya, D.J. Flavonoid Antioxidants: Chemistry, Metabolism and Structure-Activity Relationships. J. Nutr. Biochem. 2002, 13, 572–584. [Google Scholar] [CrossRef]
- Brett, A.M.O.; Ghica, M.-E. Electrochemical Oxidation of Quercetin. Electroanalysis 2003, 15, 1745–1750. [Google Scholar] [CrossRef] [Green Version]
- Pinzari, F.; Ceci, A.; Abu-Samra, N.; Canfora, L.; Maggi, O.; Persiani, A. Phenotype MicroArrayTM System in the Study of Fungal Functional Diversity and Catabolic Versatility. Res. Microbiol. 2016, 167, 710–722. [Google Scholar] [CrossRef]
- Noveroske, R.L.; Kui, J.; Williams, E.B. Oxidation of Phloridzin and Phloretin Related to Resistance of Malus to Venturia Inaequalis. Phytopathology 1964, 54, 92–97. [Google Scholar]
- Gessler, C.; Patocchi, A.; Sansavini, S.; Tartarini, S.; Gianfranceschi, L. Venturia Inaequalis Resistance in Apple. CRC. Crit. Rev. Plant Sci. 2006, 25, 473–503. [Google Scholar] [CrossRef]
- Gosch, C.; Halbwirth, H.; Stich, K. Phloridzin: Biosynthesis, Distribution and Physiological Relevance in Plants. Phytochemistry 2010, 71, 838–843. [Google Scholar] [CrossRef] [PubMed]
- Baldisserotto, A.; Malisardi, G.; Scalambra, E.; Andreotti, E.; Romagnoli, C.; Vicentini, C.B.; Manfredini, S.; Vertuani, S. Synthesis, Antioxidant and Antimicrobial Activity of a New Phloridzin Derivative for Dermo-Cosmetic Applications. Molecules 2012, 17, 13275–13289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Lee, S.J.; Park, S.; Jeong, S.W.; Kim, C.Y.; Jin, J.S.; Jeong, E.D.; Kwak, Y.S.; Kim, S.T.; Bae, D.W.; et al. Determination of Flavonoid Level Variation in Onion (Allium Cepa L.) Infected by Fusarium oxysporum Using Liquid Chromatography–tandem Mass Spectrometry. Food Chem. 2012, 133, 1653–1657. [Google Scholar] [CrossRef]
- Das, S.; Rosazza, J.P. Microbial and Enzymatic Transformations of Flavonoids. J. Nat. Prod. 2006, 69, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, G.B.; Smagghe, G.; Wittevrongel, J.; Huynh, N.T.; Van Camp, J.; Raes, K. Metabolism of Quercetin and Naringenin by Food-Grade Fungal Inoculum, Rhizopus azygosporus Yuan et Jong (ATCC 48108). J. Agric. Food Chem. 2016, 64, 9263–9267. [Google Scholar] [CrossRef] [PubMed]
- Mierziak, J.; Kostyn, K.; Kulma, A. Flavonoids as Important Molecules of Plant Interactions with the Environment. Molecules 2014, 19, 16240–16265. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, L.; Atanasova-Penichon, V.; Chéreau, S.; Richard-Forget, F. Metabolomics to Decipher the Chemical Defense of Cereals against Fusarium graminearum and Deoxynivalenol Accumulation. Int. J. Mol. Sci. 2015, 16, 24839–24872. [Google Scholar] [CrossRef] [PubMed]
- Oszust, K.; Pawlik, A.; Siczek, A.; Janusz, G.; Gryta, A.; Bilińska-Wielgus, N.; Frąc, M. Efficient Cellulases Production by Trichoderma atroviride G79/11 in Submerged Culture Based on Soy Flour-Cellulose-Lactose. BioResources 2017, 12, 8468–8489. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J.L. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. A Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Oleszek, M.; Krzemińska, I. Enhancement of biogas production by co-digestion of maize silage with common goldenrod rich in biologically active compounds. BioResources 2017, 12, 704–714. [Google Scholar] [CrossRef]
- Kozachok, S.; Pecio, Ł.; Kolodziejczyk-Czepas, J.; Marchyshyn, S.; Nowak, P.; Mołdoch, J.; Oleszek, W. γ-Pyrone Compounds: Flavonoids and Maltol Glucoside Derivatives from Herniaria glabra L. Collected in the Ternopil Region of the Ukraine. Phytochemistry 2018, 152, 213–222. [Google Scholar] [CrossRef]
- Żuchowski, J.; Pecio, Ł.; Reszczyńska, E.; Stochmal, A. New Phenolic Compounds from the Roots of Lentil (Lens culinaris). Helv. Chim. Acta 2016, 99, 674–680; [Google Scholar] [CrossRef]
- Oleszek, M.; Kozachok, S. Antioxidant Activity of Plant Extracts and Their Effect on Methane Fermentation in Bioreactors. Int. Agrophysics 2018, 32, 395–401. [Google Scholar] [CrossRef]


{kind=link}
| Rt (min) | Compound | MW (g mol−1) | % w/w (Relative) | |||||
|---|---|---|---|---|---|---|---|---|
| 1 CE | PE | F1 | F4 | F5 | F6 | |||
| 3.77 | Pinnatifidanoside D (vomifoliol-9-O-[β-D-Xyl(1→6)-β-D-Glc]) | 518 | 0.11 (14) | 1.23 (16) | 5.9 (100) | - | - | - |
| 6.29 | Hyperoside (Q-3-O-β-D-Gal) | 464 | 0.16 (21) | 1.55 (20) | - | - | 33.4 (43) | 1.91 (3) |
| 6.33 | Rutin (Q-3-O-α-L-Rha(1→6)-β-D-Glc) | 610 | * | * | - | 2.3(5) | - | - |
| 6.64 | Isoquercetin (Q-3-O-β-D-Glc) | 464 | 0.02 (3) | 0.25 (3) | - | - | 5.2 (7) | - |
| 7.20 | Reynoutrin (Q-3-O-β-D-Xyl) | 434 | 0.05 (6) | 0.52 (7) | - | - | - | 20.17 (32) |
| 7.53 | Q-3-O-pentosyl | 434 | - | 0.05 (1) | - | - | - | 1.82 (3) |
| 7.91 | Avicularin (Q-3-O-α-L-Ara) | 434 | 0.1 (13) | 1.02 (13) | - | - | 3.5 (5) | 34.10 (53) |
| 8.19 | Q-3-O-pentosyl | 434 | 0.02 (2) | 0.16 (2) | - | - | - | 5.86 (9) |
| 8.47 | Quercitrin (Q-3-O-α-L-Rha) | 448 | 0.15 (19) | 1.58 (20) | - | - | 34.9 (45) | - |
| 10.31 | Phloridzin (phloretin-2′-O-β-D-Glc) | 436 | 0.17 (22) | 1.29 (17) | - | 44.7 (95) | - | - |
| 12.53 | Quercetin | 302 | * | 0.09 (1) | - | - | - | - |
| Total, % w/w | 0.77 | 7.75 | 5.9 | 47.00 | 77.04 | 63.86 | ||
| Sample | Reducing Power EC50 (μg mL−1) | Radical-Scavenging Activity IC50 (μg mL−1) |
|---|---|---|
| CE | >1500 | >1500 |
| PE | 298.33 ± 5.84 | 444.65 ± 10.57 |
| PF | >1500 | >1500 |
| F1 | 460.60 ± 28.84 | 1117.21 ± 59.10 |
| F4 | 137.38 ± 1.61 | 188.54 ± 7.95 |
| F5 | 100.83 ± 1.62 | 105.92 ± 1.23 |
| F6 | 93.94 ± 2.68 | 107.22 ± 1.77 |
| Ascorbic acid | 27.82 ± 0.07 | 73.61 ± 6.35 |
| Tested Sample | Concentration (µL mL−1) | A490/A750 | |||
|---|---|---|---|---|---|
| N. fischeri | F. oxysporum | Botrytis sp. | P. setifera | ||
| Crude extract | 0 | 0.93 | 0.96 | 0.89 | 0.89 |
| 5 | 0.97 | 1.05 | 1.09 | 0.91 | |
| 50 | 1.00 | 1.07 | 1.10 | 0.80 | |
| 100 | 0.99 | 1.07 | 1.08 | 0.91 | |
| 500 | 1.04 | 1.15 | 1.14 | 0.74 | |
| Purified extract | 0 | 0.93 | 0.96 | 0.89 | 0.89 |
| 5 | 1.02 | 1.07 | 1.06 | 1.07 | |
| 50 | 0.99 | 1.03 | 1.06 | 0.92 | |
| 100 | 0.98 | 1.01 | 1.11 | 0.71 | |
| 500 | 0.95 | 1.01 | 1.03 | 0.52 | |
| Polar fraction of the extract | 0 | 0.93 | 0.96 | 0.89 | 0.89 |
| 5 | 0.96 | 1.02 | 1.02 | 1.01 | |
| 50 | 1.00 | 1.06 | 1.03 | 0.94 | |
| 100 | 0.99 | 1.05 | 1.03 | 0.90 | |
| 500 | 1.04 | 1.10 | 1.04 | 0.94 | |
| Fraction 1 | 0 | 0.93 | 0.96 | 0.89 | 0.89 |
| 5 | 0.96 | 1.00 | 0.98 | 0.67 | |
| 50 | 0.98 | 1.02 | 1.01 | 0.59 | |
| 100 | 0.97 | 1.01 | 1.02 | 0.75 | |
| 500 | 0.94 | 0.97 | 0.95 | 0.59 | |
| Fraction 4 | 0 | 0.93 | 0.96 | 0.89 | 0.89 |
| 5 | 0.95 | 0.97 | 1.04 | 0.67 | |
| 50 | 0.93 | 1.04 | 0.97 | 0.78 | |
| 100 | 0.98 | 1.13 | 1.01 | 0.96 | |
| 500 | 1.03 | 1.44 | 1.20 | 1.26 | |
| Fraction 5 | 0 | 0.93 | 0.96 | 0.89 | 0.89 |
| 5 | 0.93 | 0.95 | 0.95 | 0.58 | |
| 50 | 0.93 | 0.94 | 0.93 | 0.62 | |
| 100 | 0.93 | 0.95 | 0.94 | 0.75 | |
| 500 | 1.01 | 1.02 | 1.02 | 0.89 | |
| Fraction 6 | 0 | 0.93 | 0.96 | 0.89 | 0.89 |
| 5 | 1.01 | 1.07 | 1.09 | 0.69 | |
| 50 | 1.04 | 1.13 | 1.11 | 0.85 | |
| 100 | 1.02 | 1.12 | 1.05 | 0.79 | |
| 500 | 1.00 | 1.06 | 1.10 | 0.82 | |
 —A490/A750 < 0.75,
—A490/A750 < 0.75,  —0.75 ≤ A490/A750 ≤ 0.95,
—0.75 ≤ A490/A750 ≤ 0.95,  —0.95 ≤ A490/A750 < 1.05,
—0.95 ≤ A490/A750 < 1.05,  —1.05 ≤ A490/A750 ≤ 1.10,
—1.05 ≤ A490/A750 ≤ 1.10,  —1.10 < A490/A750 ≤ 1.20,
—1.10 < A490/A750 ≤ 1.20,  —1.20 < A490/A750.
—1.20 < A490/A750.© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oleszek, M.; Pecio, Ł.; Kozachok, S.; Lachowska-Filipiuk, Ż.; Oszust, K.; Frąc, M. Phytochemicals of Apple Pomace as Prospect Bio-Fungicide Agents against Mycotoxigenic Fungal Species—In Vitro Experiments. Toxins 2019, 11, 361. https://doi.org/10.3390/toxins11060361
Oleszek M, Pecio Ł, Kozachok S, Lachowska-Filipiuk Ż, Oszust K, Frąc M. Phytochemicals of Apple Pomace as Prospect Bio-Fungicide Agents against Mycotoxigenic Fungal Species—In Vitro Experiments. Toxins. 2019; 11(6):361. https://doi.org/10.3390/toxins11060361
Chicago/Turabian StyleOleszek, Marta, Łukasz Pecio, Solomiia Kozachok, Żaneta Lachowska-Filipiuk, Karolina Oszust, and Magdalena Frąc. 2019. "Phytochemicals of Apple Pomace as Prospect Bio-Fungicide Agents against Mycotoxigenic Fungal Species—In Vitro Experiments" Toxins 11, no. 6: 361. https://doi.org/10.3390/toxins11060361
APA StyleOleszek, M., Pecio, Ł., Kozachok, S., Lachowska-Filipiuk, Ż., Oszust, K., & Frąc, M. (2019). Phytochemicals of Apple Pomace as Prospect Bio-Fungicide Agents against Mycotoxigenic Fungal Species—In Vitro Experiments. Toxins, 11(6), 361. https://doi.org/10.3390/toxins11060361