
Capsaicin and TRPV1 Channels in the Cardiovascular System: The Role of Inflammation

 and
and 
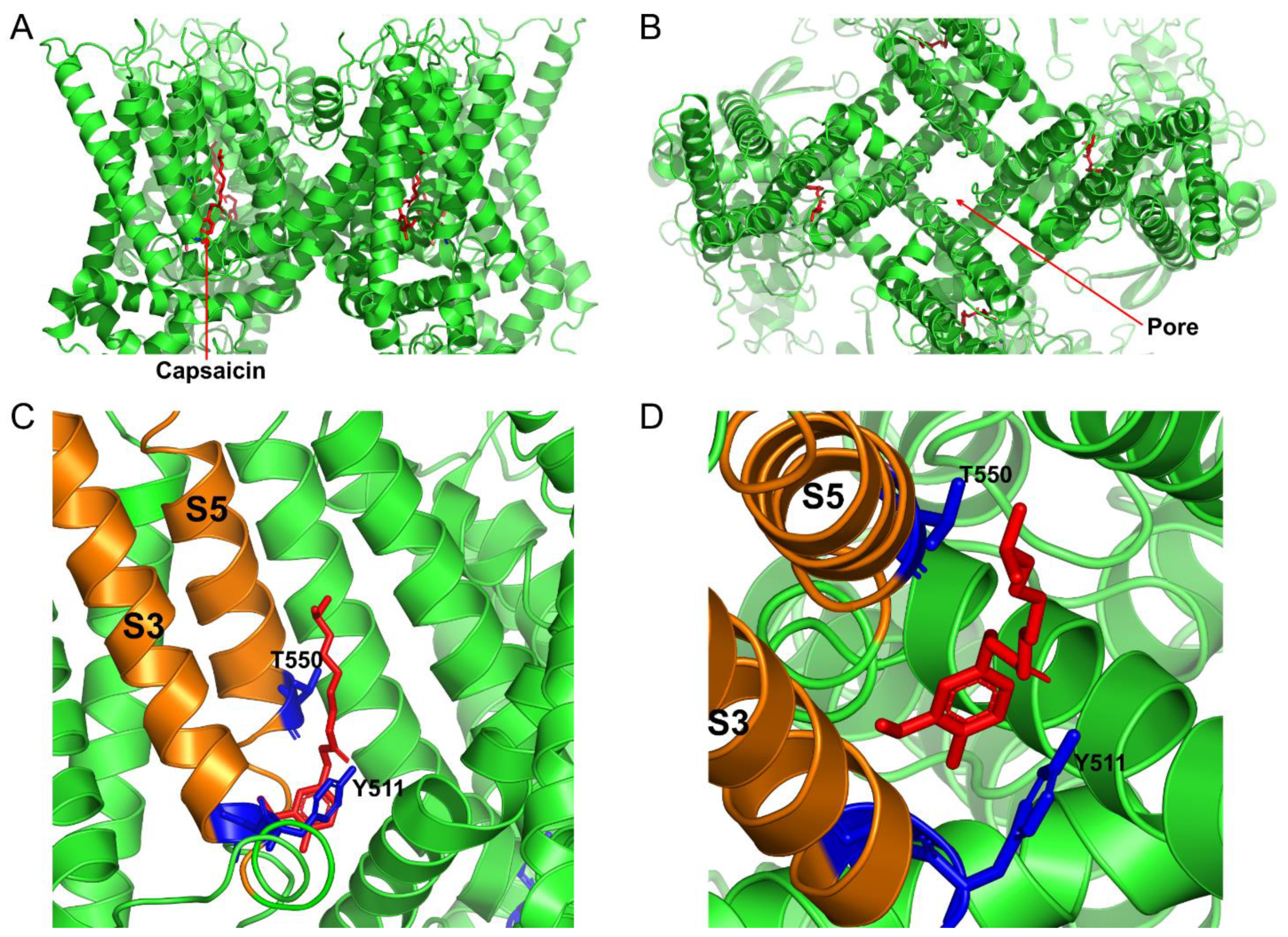{kind=link}
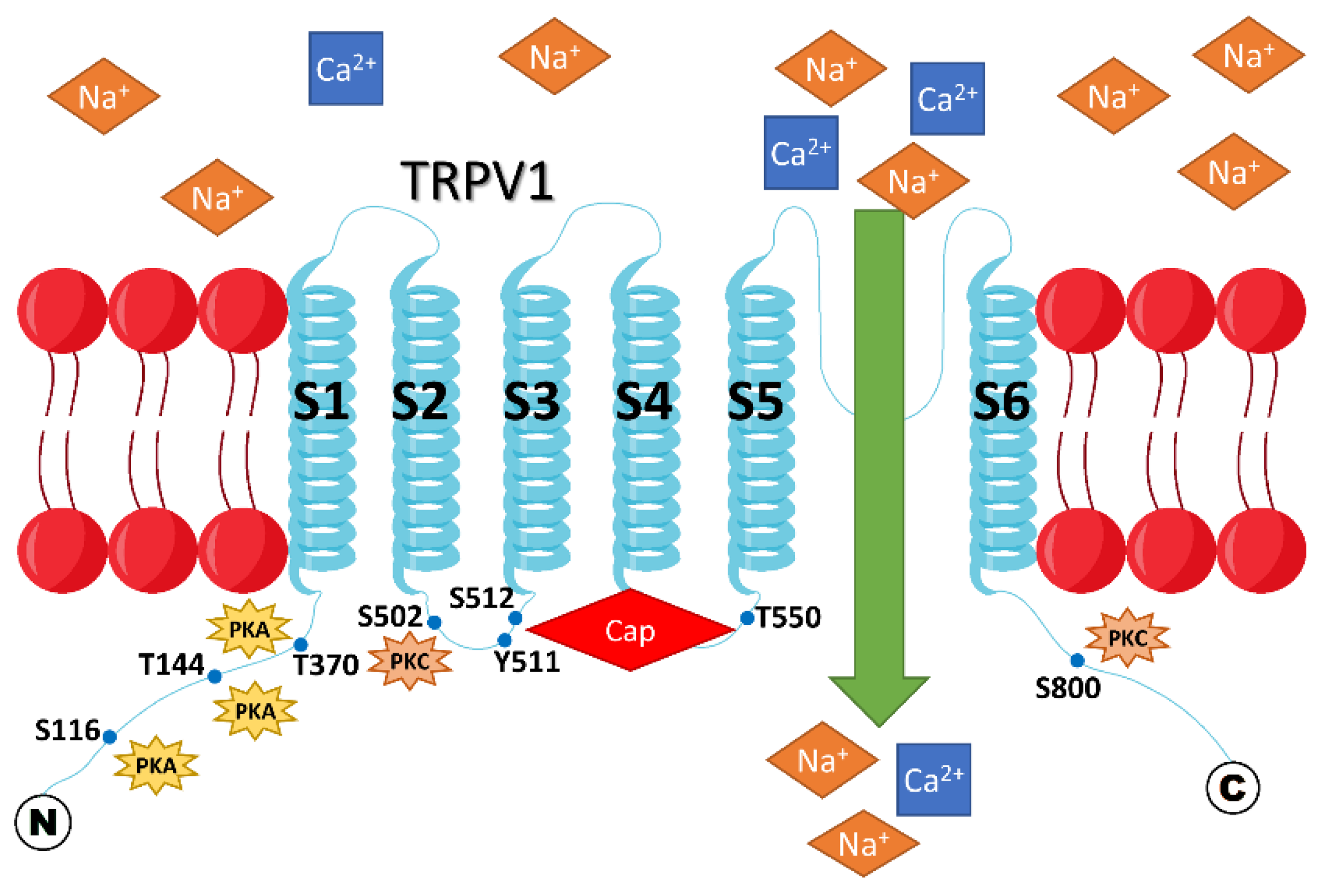{kind=link}
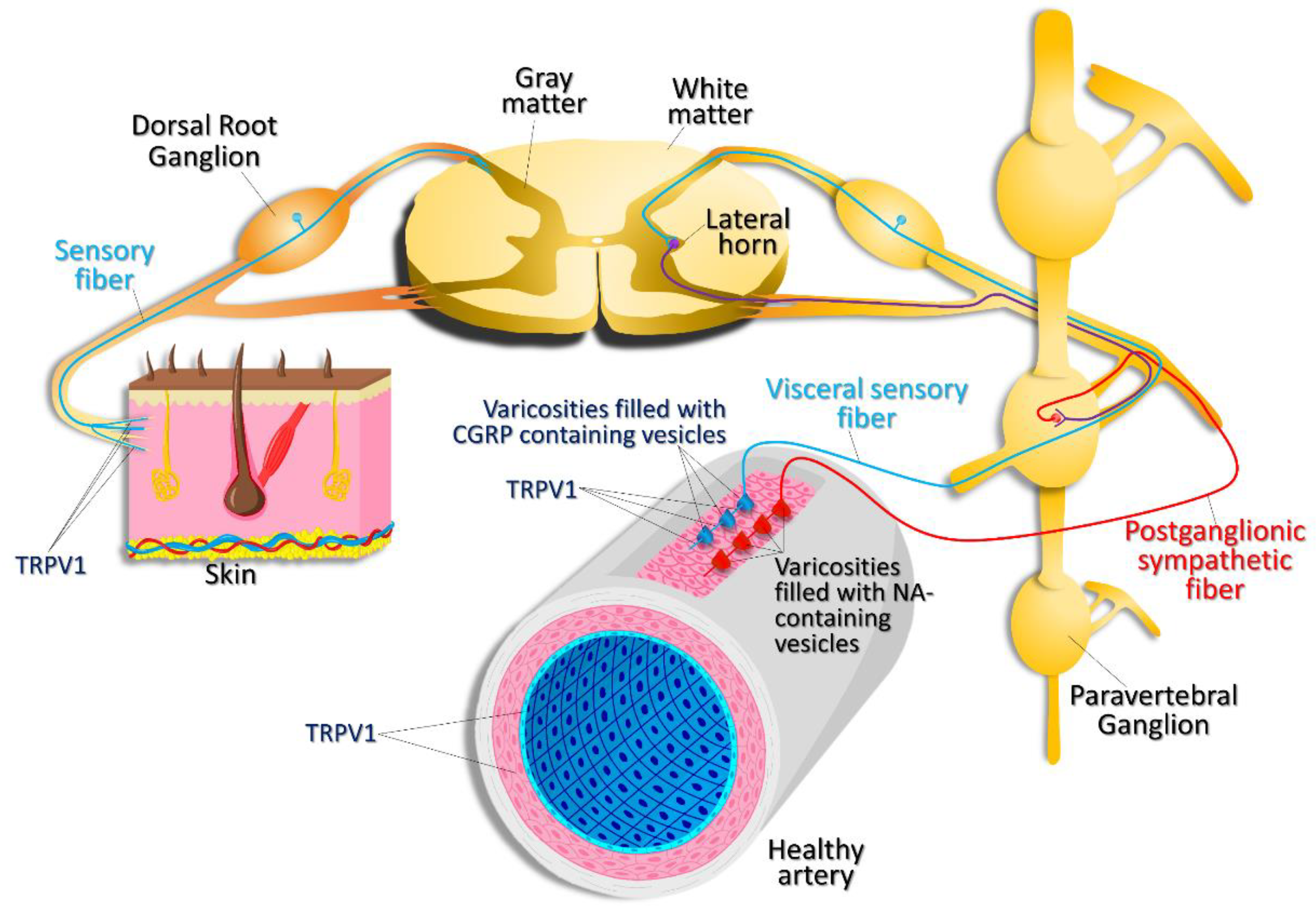{kind=link}
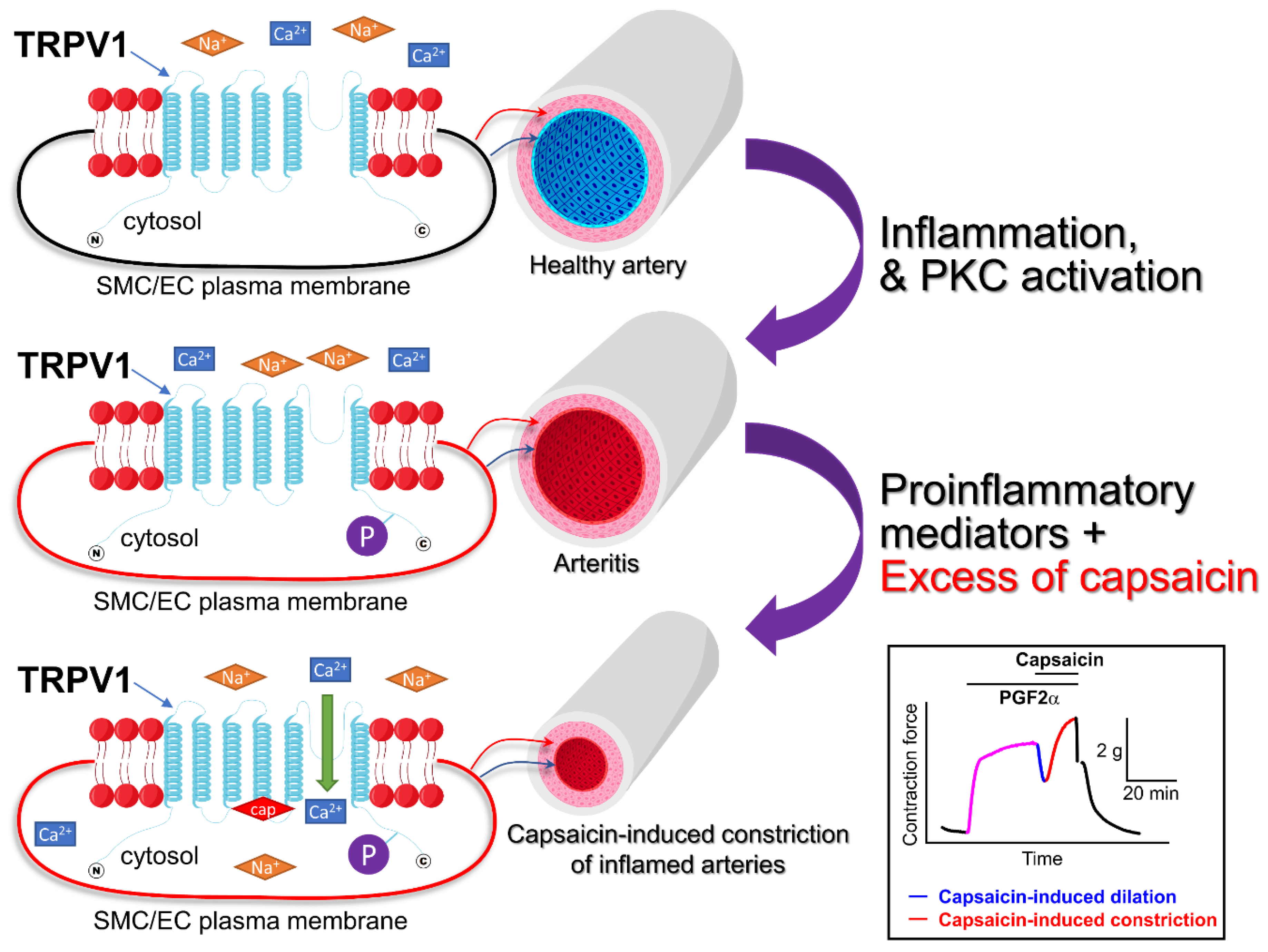{kind=link}
Abstract
:1. Introduction
2. The Beneficial Roles of Capsaicin and TRPV1
3. Vascular Effects of Capsaicin
3.1. Vasoconstrictory Effects of Capsaicin
3.2. Vasodilatory Effects of Capsaicin
3.3. Capsaicin-Induced TRPV1 Activation and Atherosclerosis
3.4. Capsaicin and Myocardial Infarction
3.5. Capsaicin-Induced TRPV1 Activation and Inflammation
3.6. Pharmacokinetics of Capsaicin
4. Clinical Implication of Capsaicin Overuse
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Govindarajan, V.S. Capsicum production, technology, chemistry, and quality. Part 1: History, botany, cultivation, and primary processing. Crit. Rev. Food Sci. Nutr. 1985, 22, 109–176. [Google Scholar] [CrossRef] [PubMed]
- Kraft, K.H.; Brown, C.H.; Nabhan, G.P.; Luedeling, E.; Luna Ruiz Jde, J.; Coppens d’Eeckenbrugge, G.; Hijmans, R.J.; Gepts, P. Multiple lines of evidence for the origin of domesticated chili pepper, Capsicum annuum, in Mexico. Proc. Natl. Acad. Sci. USA 2014, 111, 6165–6170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordt, S.E.; Julius, D. Molecular basis for species-specific sensitivity to “hot” chili peppers. Cell 2002, 108, 421–430. [Google Scholar] [CrossRef] [Green Version]
- Kwon, D.H.; Zhang, F.; Suo, Y.; Bouvette, J.; Borgnia, M.J.; Lee, S.Y. Heat-dependent opening of TRPV1 in the presence of capsaicin. Nat. Struct. Mol. Biol. 2021, 28, 554–563. [Google Scholar] [CrossRef]
- Cesare, P.; Moriondo, A.; Vellani, V.; McNaughton, P.A. Ion channels gated by heat. Proc. Natl. Acad. Sci. USA 1999, 96, 7658–7663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kollarik, M.; Undem, B.J. Mechanisms of acid-induced activation of airway afferent nerve fibres in guinea-pig. J. Physiol. 2002, 543, 591–600. [Google Scholar] [CrossRef]
- Jordt, S.E.; Tominaga, M.; Julius, D. Acid potentiation of the capsaicin receptor determined by a key extracellular site. Proc. Natl. Acad. Sci. USA 2000, 97, 8134–8139. [Google Scholar] [CrossRef] [Green Version]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 1997, 389, 816–824. [Google Scholar] [CrossRef]
- Caterina, M.J.; Leffler, A.; Malmberg, A.B.; Martin, W.J.; Trafton, J.; Petersen-Zeitz, K.R.; Koltzenburg, M.; Basbaum, A.I.; Julius, D. Impaired nociception and pain sensation in mice lacking the capsaicin receptor. Science 2000, 288, 306–313. [Google Scholar] [CrossRef]
- Kichko, T.I.; Reeh, P.W. Why cooling is beneficial: Non-linear temperature-dependency of stimulated iCGRP release from isolated rat skin. Pain 2004, 110, 215–219. [Google Scholar] [CrossRef]
- Benitez-Angeles, M.; Morales-Lazaro, S.L.; Juarez-Gonzalez, E.; Rosenbaum, T. TRPV1: Structure, Endogenous Agonists, and Mechanisms. Int. J. Mol. Sci. 2020, 21, 3421. [Google Scholar] [CrossRef] [PubMed]
- Xin, H.; Tanaka, H.; Yamaguchi, M.; Takemori, S.; Nakamura, A.; Kohama, K. Vanilloid receptor expressed in the sarcoplasmic reticulum of rat skeletal muscle. Biochem. Biophys. Res. Commun. 2005, 332, 756–762. [Google Scholar] [CrossRef] [PubMed]
- Olah, Z.; Szabo, T.; Karai, L.; Hough, C.; Fields, R.D.; Caudle, R.M.; Blumberg, P.M.; Iadarola, M.J. Ligand-induced dynamic membrane changes and cell deletion conferred by vanilloid receptor 1. J. Biol. Chem. 2001, 276, 11021–11030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clapham, D.E. TRP channels as cellular sensors. Nature 2003, 426, 517–524. [Google Scholar] [CrossRef]
- Stander, S.; Moormann, C.; Schumacher, M.; Buddenkotte, J.; Artuc, M.; Shpacovitch, V.; Brzoska, T.; Lippert, U.; Henz, B.M.; Luger, T.A.; et al. Expression of vanilloid receptor subtype 1 in cutaneous sensory nerve fibers, mast cells, and epithelial cells of appendage structures. Exp. Dermatol. 2004, 13, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, H.; Sugimoto, T. VR1-immunoreactive primary sensory neurons in the rat trigeminal ganglion. Brain Res. 2001, 890, 184–188. [Google Scholar] [CrossRef]
- Chen, X.; Sun, W.; Gianaris, N.G.; Riley, A.M.; Cummins, T.R.; Fehrenbacher, J.C.; Obukhov, A.G. Furanocoumarins are a novel class of modulators for the transient receptor potential vanilloid type 1 (TRPV1) channel. J. Biol. Chem. 2014, 289, 9600–9610. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Ye, L.; Zhang, Q.; Wu, F.; Wang, L. The role of TRPV1 channels in atherosclerosis. Channels 2020, 14, 141–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurt, C.M.; Lu, Y.; Stary, C.M.; Piplani, H.; Small, B.A.; Urban, T.J.; Qvit, N.; Gross, G.J.; Mochly-Rosen, D.; Gross, E.R. Transient Receptor Potential Vanilloid 1 Regulates Mitochondrial Membrane Potential and Myocardial Reperfusion Injury. J. Am. Heart Assoc. 2016, 5, e003774. [Google Scholar] [CrossRef]
- Andrei, S.R.; Sinharoy, P.; Bratz, I.N.; Damron, D.S. TRPA1 is functionally co-expressed with TRPV1 in cardiac muscle: Co-localization at z-discs, costameres and intercalated discs. Channels 2016, 10, 395–409. [Google Scholar] [CrossRef] [PubMed]
- Hoebart, C.; Rojas-Galvan, N.S.; Ciotu, C.I.; Aykac, I.; Reissig, L.F.; Weninger, W.J.; Kiss, A.; Podesser, B.K.; Fischer, M.J.M.; Heber, S. No functional TRPA1 in cardiomyocytes. Acta Physiol. 2021, 232, e13659. [Google Scholar] [CrossRef] [PubMed]
- Yoshie, K.; Rajendran, P.S.; Massoud, L.; Mistry, J.; Swid, M.A.; Wu, X.; Sallam, T.; Zhang, R.; Goldhaber, J.I.; Salavatian, S.; et al. Cardiac TRPV1 afferent signaling promotes arrhythmogenic ventricular remodeling after myocardial infarction. JCI Insight 2020, 5, e124477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrade, F.; Rangel-Sandoval, C.; Rodriguez-Hernandez, A.; Lopez-Dyck, E.; Elizalde, A.; Virgen-Ortiz, A.; Bonales-Alatorre, E.; Valencia-Cruz, G.; Sanchez-Pastor, E. Capsaicin Causes Vasorelaxation of Rat Aorta through Blocking of L-type Ca2+ Channels and Activation of CB1 Receptors. Molecules 2020, 25, 3957. [Google Scholar] [CrossRef] [PubMed]
- Hiett, S.C.; Owen, M.K.; Li, W.; Chen, X.; Riley, A.; Noblet, J.; Flores, S.; Sturek, M.; Tune, J.D.; Obukhov, A.G. Mechanisms underlying capsaicin effects in canine coronary artery: Implications for coronary spasm. Cardiovasc. Res. 2014, 103, 607–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cesare, P.; McNaughton, P. A novel heat-activated current in nociceptive neurons and its sensitization by bradykinin. Proc. Natl. Acad. Sci. USA 1996, 93, 15435–15439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amadesi, S.; Nie, J.; Vergnolle, N.; Cottrell, G.S.; Grady, E.F.; Trevisani, M.; Manni, C.; Geppetti, P.; McRoberts, J.A.; Ennes, H.; et al. Protease-activated receptor 2 sensitizes the capsaicin receptor transient receptor potential vanilloid receptor 1 to induce hyperalgesia. J. Neurosci. 2004, 24, 4300–4312. [Google Scholar] [CrossRef] [PubMed]
- Prescott, E.D.; Julius, D. A modular PIP2 binding site as a determinant of capsaicin receptor sensitivity. Science 2003, 300, 1284–1288. [Google Scholar] [CrossRef] [PubMed]
- Cao, E.; Cordero-Morales, J.F.; Liu, B.; Qin, F.; Julius, D. TRPV1 channels are intrinsically heat sensitive and negatively regulated by phosphoinositide lipids. Neuron 2013, 77, 667–679. [Google Scholar] [CrossRef] [Green Version]
- Stein, A.T.; Ufret-Vincenty, C.A.; Hua, L.; Santana, L.F.; Gordon, S.E. Phosphoinositide 3-kinase binds to TRPV1 and mediates NGF-stimulated TRPV1 trafficking to the plasma membrane. J. Gen. Physiol. 2006, 128, 509–522. [Google Scholar] [CrossRef] [Green Version]
- Rohacs, T. Phosphoinositide regulation of TRPV1 revisited. Pflügers Arch. Eur. J. Physiol. 2015, 467, 1851–1869. [Google Scholar] [CrossRef] [Green Version]
- Bhave, G.; Zhu, W.; Wang, H.; Brasier, D.J.; Oxford, G.S.; Gereau, R.W. cAMP-dependent protein kinase regulates desensitization of the capsaicin receptor (VR1) by direct phosphorylation. Neuron 2002, 35, 721–731. [Google Scholar] [CrossRef] [Green Version]
- De Petrocellis, L.; Harrison, S.; Bisogno, T.; Tognetto, M.; Brandi, I.; Smith, G.D.; Creminon, C.; Davis, J.B.; Geppetti, P.; Di Marzo, V. The vanilloid receptor (VR1)-mediated effects of anandamide are potently enhanced by the cAMP-dependent protein kinase. J. Neurochem. 2001, 77, 1660–1663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Premkumar, L.S.; Ahern, G.P. Induction of vanilloid receptor channel activity by protein kinase C. Nature 2000, 408, 985–990. [Google Scholar] [CrossRef]
- Mandadi, S.; Tominaga, T.; Numazaki, M.; Murayama, N.; Saito, N.; Armati, P.J.; Roufogalis, B.D.; Tominaga, M. Increased sensitivity of desensitized TRPV1 by PMA occurs through PKCepsilon-mediated phosphorylation at S800. Pain 2006, 123, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Jeske, N.A.; Patwardhan, A.M.; Henry, M.A.; Milam, S.B. Fibronectin stimulates TRPV1 translocation in primary sensory neurons. J. Neurochem. 2009, 108, 591–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, J.; Shin, J.S.; Lee, S.Y.; Hwang, S.W.; Koo, J.; Cho, H.; Oh, U. Phosphorylation of vanilloid receptor 1 by Ca2+/calmodulin-dependent kinase II regulates its vanilloid binding. J. Biol. Chem. 2004, 279, 7048–7054. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Duan, Y.; Riley, A.M.; Welch, M.A.; White, F.A.; Grant, M.B.; Obukhov, A.G. Long-term diabetic microenvironment augments the decay rate of capsaicin-induced currents in mouse dorsal root ganglion neurons. Molecules 2019, 24, 775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, R.R.; Samad, T.A.; Jin, S.X.; Schmoll, R.; Woolf, C.J. p38 MAPK activation by NGF in primary sensory neurons after inflammation increases TRPV1 levels and maintains heat hyperalgesia. Neuron 2002, 36, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Yoshioka, M.; Lim, K.; Kikuzato, S.; Kiyonaga, A.; Tanaka, H.; Shindo, M.; Suzuki, M. Effects of red-pepper diet on the energy metabolism in men. J. Nutr. Sci. Vitaminol. 1995, 41, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, M.; St-Pierre, S.; Drapeau, V.; Dionne, I.; Doucet, E.; Suzuki, M.; Tremblay, A. Effects of red pepper on appetite and energy intake. Br. J. Nutr. 1999, 82, 115–123. [Google Scholar] [CrossRef]
- Lacroix, J.S.; Buvelot, J.M.; Polla, B.S.; Lundberg, J.M. Improvement of symptoms of non-allergic chronic rhinitis by local treatment with capsaicin. Clin. Exp. Allergy 1991, 21, 595–600. [Google Scholar] [CrossRef]
- Hara, A.; Sakurada, T.; Sakurada, S.; Matsumura, H.; Kisara, K. Antinociceptive effects of neonatal capsaicin in rats with adjuvant arthritis. Naunyn. Schmiedebergs Arch. Pharmacol. 1984, 326, 248–253. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Zaror-Behrens, G.; Himms-Hagen, J. Capsaicin desensitization induces atrophy of brown adipose tissue in rats. Am. J. Physiol. 1990, 259, R324–R332. [Google Scholar] [CrossRef]
- Banerjee, S.; McCormack, S. Capsaicin for Acute or Chronic Non-Cancer Pain: A Review of Clinical Effectiveness, Safety, and Cost-Effectiveness; Canadian Agency for Drugs and Technologies in Health: Ottawa, ON, Canada, 2020.
- Elmas, C.; Gezer, C. Capsaicin and Its Effects on Body Weight. J. Am. Coll. Nutr. 2021, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, J.E.; Parish, L.C.; Rapaport, M.; Rosenbaum, M.M.; Roenigk, H.H., Jr. Effects of topically applied capsaicin on moderate and severe psoriasis vulgaris. J. Am. Acad. Dermatol. 1986, 15, 504–507. [Google Scholar] [CrossRef]
- Final report on the safety assessment of capsicum annuum extract, capsicum annuum fruit extract, capsicum annuum resin, capsicum annuum fruit powder, capsicum frutescens fruit, capsicum frutescens fruit extract, capsicum frutescens resin, and capsaicin. Int. J. Toxicol. 2007, 26 (Suppl. S1), 3–106. [CrossRef] [PubMed]
- Randhawa, P.K.; Jaggi, A.S. TRPV1 channels in cardiovascular system: A double edged sword? Int. J. Cardiol. 2017, 228, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Basith, S.; Cui, M.; Hong, S.; Choi, S. Harnessing the Therapeutic Potential of Capsaicin and Its Analogues in Pain and Other Diseases. Molecules 2016, 21, 966. [Google Scholar] [CrossRef] [Green Version]
- Jancso, G.; Kiraly, E.; Jancso-Gabor, A. Pharmacologically induced selective degeneration of chemosensitive primary sensory neurones. Nature 1977, 270, 741–743. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.L.; Yan Liu, D.; Ma, L.Q.; Luo, Z.D.; Cao, T.B.; Zhong, J.; Yan, Z.C.; Wang, L.J.; Zhao, Z.G.; Zhu, S.J.; et al. Activation of transient receptor potential vanilloid type-1 channel prevents adipogenesis and obesity. Circ. Res. 2007, 100, 1063–1070. [Google Scholar] [CrossRef] [Green Version]
- Kang, J.H.; Goto, T.; Han, I.S.; Kawada, T.; Kim, Y.M.; Yu, R. Dietary capsaicin reduces obesity-induced insulin resistance and hepatic steatosis in obese mice fed a high-fat diet. Obesity 2010, 18, 780–787. [Google Scholar] [CrossRef]
- Avraham, Y.; Zolotarev, O.; Grigoriadis, N.C.; Poutahidis, T.; Magen, I.; Vorobiav, L.; Zimmer, A.; Ilan, Y.; Mechoulam, R.; Berry, E.M. Cannabinoids and capsaicin improve liver function following thioacetamide-induced acute injury in mice. Am. J. Gastroenterol. 2008, 103, 3047–3056. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wang, D.; Huang, J.; Hu, Y.; Xu, Y. Application of capsaicin as a potential new therapeutic drug in human cancers. J. Clin. Pharm. Ther. 2020, 45, 16–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, J.K.; Han, K.Y.; Kim, E.C.; Kim, Y.M.; Lee, S.W.; Kim, O.H.; Kim, K.W.; Gho, Y.S.; Kwon, Y.G. Capsaicin inhibits in vitro and in vivo angiogenesis. Cancer Res. 2004, 64, 644–651. [Google Scholar] [CrossRef] [Green Version]
- Amantini, C.; Ballarini, P.; Caprodossi, S.; Nabissi, M.; Morelli, M.B.; Lucciarini, R.; Cardarelli, M.A.; Mammana, G.; Santoni, G. Triggering of transient receptor potential vanilloid type 1 (TRPV1) by capsaicin induces Fas/CD95-mediated apoptosis of urothelial cancer cells in an ATM-dependent manner. Carcinogenesis 2009, 30, 1320–1329. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Xiao, C.; Jiang, W.; Yang, W.; Qin, Q.; Tan, Q.; Lian, B.; Liang, Z.; Wei, C. Capsaicin Inhibits Proliferation and Induces Apoptosis in Breast Cancer by Down-Regulating FBI-1-Mediated NF-kappaB Pathway. Drug Des. Devel. Ther. 2021, 15, 125–140. [Google Scholar] [CrossRef] [PubMed]
- Toth, B.; Gannett, P. Carcinogenicity of lifelong administration of capsaicin of hot pepper in mice. In Vivo 1992, 6, 59–63. [Google Scholar] [PubMed]
- Notani, P.N.; Jayant, K. Role of diet in upper aerodigestive tract cancers. Nutr. Cancer 1987, 10, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Buiatti, E.; Palli, D.; Decarli, A.; Amadori, D.; Avellini, C.; Bianchi, S.; Biserni, R.; Cipriani, F.; Cocco, P.; Giacosa, A.; et al. A case-control study of gastric cancer and diet in Italy. Int. J. Cancer 1989, 44, 611–616. [Google Scholar] [CrossRef]
- Lopez-Carrillo, L.; Hernandez Avila, M.; Dubrow, R. Chili pepper consumption and gastric cancer in Mexico: A case-control study. Am. J. Epidemiol. 1994, 139, 263–271. [Google Scholar] [CrossRef]
- Bayliss, W.M. On the origin from the spinal cord of the vaso-dilator fibres of the hind-limb, and on the nature of these fibres. J. Physiol. 1901, 26, 173–209. [Google Scholar] [CrossRef] [PubMed]
- Jancso, N.; Jancso-Gabor, A.; Szolcsanyi, J. Direct evidence for neurogenic inflammation and its prevention by denervation and by pretreatment with capsaicin. Br. J. Pharmacol. Chemother. 1967, 31, 138–151. [Google Scholar] [CrossRef] [Green Version]
- Jessell, T.M.; Iversen, L.L.; Cuello, A.C. Capsaicin-induced depletion of substance P from primary sensory neurones. Brain Res. 1978, 152, 183–188. [Google Scholar] [CrossRef]
- Zhong, B.; Ma, S.; Wang, D.H. Ablation of TRPV1 Elevates Nocturnal Blood Pressure in Western Diet-fed Mice. Curr. Hypertens. Rev. 2019, 15, 144–153. [Google Scholar] [CrossRef]
- Phan, T.X.; Ton, H.T.; Gulyas, H.; Porszasz, R.; Toth, A.; Russo, R.; Kay, M.W.; Sahibzada, N.; Ahern, G.P. TRPV1 expressed throughout the arterial circulation regulates vasoconstriction and blood pressure. J. Physiol. 2020, 598, 5639–5659. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Hou, J.; Du, J.; Feng, J.; Yang, Y.; Shen, Y.; Chen, S.; Feng, J.; Yang, D.; Li, D.; et al. Potential Protective Mechanism in the Cardiac Microvascular Injury. Hypertension 2018, 72, 116–127. [Google Scholar] [CrossRef]
- Ives, S.J.; Park, S.Y.; Kwon, O.S.; Gifford, J.R.; Andtbacka, R.H.I.; Hyngstrom, J.R.; Richardson, R.S. TRPV1 channels in human skeletal muscle feed arteries: Implications for vascular function. Exp. Physiol. 2017, 102, 1245–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donnerer, J.; Lembeck, F. Analysis of the effects of intravenously injected capsaicin in the rat. Naunyn. Schmiedebergs Arch. Pharmacol. 1982, 320, 54–57. [Google Scholar] [CrossRef]
- Pleschka, K.; Diebold, K.; Hardt, R.; Meyer, J.U. Responsiveness of microcirculation and local cold vasodilation to capsaicin in the intact and chronically denervated canine tongue. Acta Physiol. Hung. 1987, 69, 367–373. [Google Scholar]
- Manzini, S.; Perretti, F. Vascular effects of capsaicin in isolated perfused rat mesenteric bed. Eur. J. Pharmacol. 1988, 148, 153–159. [Google Scholar] [CrossRef]
- Russell, F.A.; King, R.; Smillie, S.J.; Kodji, X.; Brain, S.D. Calcitonin gene-related peptide: Physiology and pathophysiology. Physiol. Rev. 2014, 94, 1099–1142. [Google Scholar] [CrossRef] [Green Version]
- Kee, Z.; Kodji, X.; Brain, S.D. The Role of Calcitonin Gene Related Peptide (CGRP) in Neurogenic Vasodilation and Its Cardioprotective Effects. Front. Physiol. 2018, 9, 1249. [Google Scholar] [CrossRef] [Green Version]
- Pitetti, K.H.; Cole, D.J.; Ordway, G.A. Activation of gastric afferent fibers increases coronary arterial resistance in anesthetized dogs. J. Auton. Nerv. Syst. 1988, 23, 25–34. [Google Scholar] [CrossRef]
- Martin, S.E.; Pilkington, D.M.; Longhurst, J.C. Coronary vascular responses to chemical stimulation of abdominal visceral organs. Am. J. Physiol. 1989, 256, H735–H744. [Google Scholar] [CrossRef]
- Salonen, R.O.; Webber, S.E.; Widdicombe, J.G. Effects of neuropeptides and capsaicin on the canine tracheal vasculature in vivo. Br. J. Pharmacol. 1988, 95, 1262–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edvinsson, L.; Jansen, I.; Kingman, T.A.; McCulloch, J. Cerebrovascular responses to capsaicin in vitro and in situ. Br. J. Pharmacol. 1990, 100, 312–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scotland, R.S.; Chauhan, S.; Davis, C.; De Felipe, C.; Hunt, S.; Kabir, J.; Kotsonis, P.; Oh, U.; Ahluwalia, A. Vanilloid receptor TRPV1, sensory C-fibers, and vascular autoregulation: A novel mechanism involved in myogenic constriction. Circ. Res. 2004, 95, 1027–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lizanecz, E.; Bagi, Z.; Pasztor, E.T.; Papp, Z.; Edes, I.; Kedei, N.; Blumberg, P.M.; Toth, A. Phosphorylation-dependent desensitization by anandamide of vanilloid receptor-1 (TRPV1) function in rat skeletal muscle arterioles and in Chinese hamster ovary cells expressing TRPV1. Mol. Pharmacol. 2006, 69, 1015–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czikora, A.; Lizanecz, E.; Bako, P.; Rutkai, I.; Ruzsnavszky, F.; Magyar, J.; Porszasz, R.; Kark, T.; Facsko, A.; Papp, Z.; et al. Structure-activity relationships of vanilloid receptor agonists for arteriolar TRPV1. Br. J. Pharmacol. 2012, 165, 1801–1812. [Google Scholar] [CrossRef] [Green Version]
- Korishettar, A.M.; Nishijima, Y.; Wang, Z.; Xie, Y.; Fang, J.; Wilcox, D.A.; Zhang, D.X. Endothelin-1 potentiates TRPV1-mediated vasoconstriction of human adipose arterioles in a protein kinase C-dependent manner. Br. J. Pharmacol. 2021, 178, 709–725. [Google Scholar] [CrossRef]
- Jancso-Gabor, A.; Szolcsanyi, J.; Jancso, N. Irreversible impairment of thermoregulation induced by capsaicin and similar pungent substances in rats and guinea-pigs. J. Physiol. 1970, 206, 495–507. [Google Scholar] [CrossRef] [PubMed]
- Ohlen, A.; Lindbom, L.; Staines, W.; Hokfelt, T.; Cuello, A.C.; Fischer, J.A.; Hedqvist, P. Substance P and calcitonin gene-related peptide: Immunohistochemical localisation and microvascular effects in rabbit skeletal muscle. Naunyn. Schmiedebergs Arch. Pharmacol. 1987, 336, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Stjarne, P.; Lundblad, L.; Lundberg, J.M. Mechanical stimulation and capsaicin evoked vasodilation by parasympathetic reflex mechanisms in the pig nasal mucosa. Acta Otolaryngol. 1993, 113, 649–654. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Lozano-Cuenca, J.; Villalon, C.M.; de Vries, R.; Garrelds, I.M.; Avezaat, C.J.; van Kats, J.P.; Saxena, P.R.; MaassenVanDenBrink, A. Pharmacological characterisation of capsaicin-induced relaxations in human and porcine isolated arteries. Naunyn. Schmiedebergs Arch. Pharmacol. 2007, 375, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Gazzieri, D.; Trevisani, M.; Tarantini, F.; Bechi, P.; Masotti, G.; Gensini, G.F.; Castellani, S.; Marchionni, N.; Geppetti, P.; Harrison, S. Ethanol dilates coronary arteries and increases coronary flow via transient receptor potential vanilloid 1 and calcitonin gene-related peptide. Cardiovasc. Res. 2006, 70, 589–599. [Google Scholar] [CrossRef]
- Luo, D.; Zhang, Y.W.; Peng, W.J.; Peng, J.; Chen, Q.Q.; Li, D.; Deng, H.W.; Li, Y.J. Transient receptor potential vanilloid 1-mediated expression and secretion of endothelial cell-derived calcitonin gene-related peptide. Regul. Pept. 2008, 150, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Bratz, I.N.; Dick, G.M.; Tune, J.D.; Edwards, J.M.; Neeb, Z.P.; Dincer, U.D.; Sturek, M. Impaired capsaicin-induced relaxation of coronary arteries in a porcine model of the metabolic syndrome. Am. J. Physiol. Heart. Circ. Physiol. 2008, 294, H2489–H2496. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Luo, Z.; Ma, S.; Wong, W.T.; Ma, L.; Zhong, J.; He, H.; Zhao, Z.; Cao, T.; Yan, Z.; et al. Activation of TRPV1 by dietary capsaicin improves endothelium-dependent vasorelaxation and prevents hypertension. Cell Metab. 2010, 12, 130–141. [Google Scholar] [CrossRef] [Green Version]
- Lembeck, F.; Donnerer, J. Postocclusive cutaneous vasodilatation mediated by substance P. Naunyn. Schmiedebergs Arch. Pharmacol. 1981, 316, 165–171. [Google Scholar] [CrossRef]
- Davignon, J.; Ganz, P. Role of endothelial dysfunction in atherosclerosis. Circulation 2004, 109, III27–III32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Chen, X.; Riley, A.M.; Hiett, S.C.; Temm, C.J.; Beli, E.; Long, X.; Chakraborty, S.; Alloosh, M.; White, F.A.; et al. Long-term spironolactone treatment reduces coronary TRPC expression, vasoconstriction, and atherosclerosis in metabolic syndrome pigs. Basic Res. Cardiol. 2017, 112, 54. [Google Scholar] [CrossRef] [PubMed]
- Tabas, I.; Garcia-Cardena, G.; Owens, G.K. Recent insights into the cellular biology of atherosclerosis. J. Cell Biol. 2015, 209, 13–22. [Google Scholar] [CrossRef]
- Kunieda, K.; Someya, A.; Horie, S.; Ajioka, H.; Murayama, T. Lafutidine-induced increase in intracellular Ca2+ concentrations in PC12 and endothelial cells. J. Pharmacol. Sci. 2005, 97, 67–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.X.; Wang, J.; Wang, C.; Liu, J.; Shi, L.P.; Xu, M.; Wang, C. Functional expression of transient receptor potential vanilloid-related channels in chronically hypoxic human pulmonary arterial smooth muscle cells. J. Membr. Biol. 2008, 223, 151–159. [Google Scholar] [CrossRef]
- Harris, D.W.; Buckley, C.; Dunn, E.C.; Almeyda, J.; Rustin, M.H. Briquet’s syndrome—An unusual case of urticaria. J. R. Soc. Med. 1991, 84, 167. [Google Scholar] [CrossRef]
- Negulesco, J.A.; Noel, S.A.; Newman, H.A.; Naber, E.C.; Bhat, H.B.; Witiak, D.T. Effects of pure capsaicinoids (capsaicin and dihydrocapsaicin) on plasma lipid and lipoprotein concentrations of turkey poults. Atherosclerosis 1987, 64, 85–90. [Google Scholar] [CrossRef]
- Ahuja, K.D.; Kunde, D.A.; Ball, M.J.; Geraghty, D.P. Effects of capsaicin, dihydrocapsaicin, and curcumin on copper-induced oxidation of human serum lipids. J. Agric. Food Chem. 2006, 54, 6436–6439. [Google Scholar] [CrossRef]
- Harper, A.G.; Brownlow, S.L.; Sage, S.O. A role for TRPV1 in agonist-evoked activation of human platelets. J. Thromb. Haemost. 2009, 7, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Xiong, S.; Wang, P.; Ma, L.; Gao, P.; Gong, L.; Li, L.; Li, Q.; Sun, F.; Zhou, X.; He, H.; et al. Ameliorating Endothelial Mitochondrial Dysfunction Restores Coronary Function via Transient Receptor Potential Vanilloid 1-Mediated Protein Kinase A/Uncoupling Protein 2 Pathway. Hypertension 2016, 67, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Sogut, O.; Kaya, H.; Gokdemir, M.T.; Sezen, Y. Acute myocardial infarction and coronary vasospasm associated with the ingestion of cayenne pepper pills in a 25-year-old male. Int. J. Emerg. Med. 2012, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Akcay, A.B.; Ozcan, T.; Seyis, S.; Acele, A. Coronary vasospasm and acute myocardial infarction induced by a topical capsaicin patch. Turk. Kardiyol. Dern. Ars. 2009, 37, 497–500. [Google Scholar] [PubMed]
- Sayin, M.R.; Karabag, T.; Dogan, S.M.; Akpinar, I.; Aydin, M. A case of acute myocardial infarction due to the use of cayenne pepper pills. Wien. Klin. Wochenschr. 2012, 124, 285–287. [Google Scholar] [CrossRef] [PubMed]
- Imamura, M.; Smith, N.C.; Garbarg, M.; Levi, R. Histamine H3-receptor-mediated inhibition of calcitonin gene-related peptide release from cardiac C fibers. A regulatory negative-feedback loop. Circ. Res. 1996, 78, 863–869. [Google Scholar] [CrossRef] [PubMed]
- Tsujikawa, K.; Yayama, K.; Hayashi, T.; Matsushita, H.; Yamaguchi, T.; Shigeno, T.; Ogitani, Y.; Hirayama, M.; Kato, T.; Fukada, S.; et al. Hypertension and dysregulated proinflammatory cytokine production in receptor activity-modifying protein 1-deficient mice. Proc. Natl. Acad. Sci. USA 2007, 104, 16702–16707. [Google Scholar] [CrossRef] [Green Version]
- Mair, J.; Lechleitner, P.; Langle, T.; Wiedermann, C.; Dienstl, F.; Saria, A. Plasma CGRP in acute myocardial infarction. Lancet 1990, 335, 168. [Google Scholar] [CrossRef]
- Tang, Z.L.; Dai, W.; Li, Y.J.; Deng, H.W. Involvement of capsaicin-sensitive sensory nerves in early and delayed cardioprotection induced by a brief ischaemia of the small intestine. Naunyn Schmiedebergs Arch Pharmacol 1999, 359, 243–247. [Google Scholar] [CrossRef]
- Kallner, G. Release and effects of calcitonin gene-related peptide in myocardial ischaemia. Scand. Cardiovasc. J. Suppl. 1998, 49, 1–36. [Google Scholar] [PubMed]
- Wahrborg, P.; Eliasson, T.; Edvardsson, N.; Ekman, R.; Mannheimer, C.; Hedner, T. Acute ischemic chest pain is not associated with increased calcitonin gene-related peptide (CGRP) levels in peripheral plasma nor in the coronary circulation. Scand. Cardiovasc. J. 1999, 33, 295–299. [Google Scholar] [CrossRef]
- Fragasso, G.; Palloshi, A.; Piatti, P.M.; Monti, L.; Rossetti, E.; Setola, E.; Montano, C.; Bassanelli, G.; Calori, G.; Margonato, A. Nitric-oxide mediated effects of transdermal capsaicin patches on the ischemic threshold in patients with stable coronary disease. J. Cardiovasc. Pharmacol. 2004, 44, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Roessler, A.E.; Lynch, T.L.t.; Haar, L.; Mallick, F.; Lui, Y.; Tranter, M.; Ren, M.H.; Xie, W.R.; Fan, G.C.; et al. Cardioprotection via the skin: Nociceptor-induced conditioning against cardiac MI in the NIC of time. Am. J. Physiol. Heart Circ. Physiol. 2019, 316, H543–H553. [Google Scholar] [CrossRef] [PubMed]
- Zhong, B.; Ma, S.; Wang, D.H. Protective Effects of TRPV1 Activation Against Cardiac Ischemia/ Reperfusion Injury is Blunted by Diet-Induced Obesity. Cardiovasc. Hematol. Disord Drug Targets 2020, 20, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.L.; Guo, Z.; Wang, L.L.; Wu, J. Degeneration of capsaicin sensitive sensory nerves enhances myocardial injury in acute myocardial infarction in rats. Int. J. Cardiol. 2012, 160, 41–47. [Google Scholar] [CrossRef]
- Gu, Y.; Li, G.; Huang, L.M. Inflammation induces Epac-protein kinase C alpha and epsilon signaling in TRPV1-mediated hyperalgesia. Pain 2018, 159, 2383–2393. [Google Scholar] [CrossRef]
- Joseph, J.; Qu, L.; Wang, S.; Kim, M.; Bennett, D.; Ro, J.; Caterina, M.J.; Chung, M.K. Phosphorylation of TRPV1 S801 Contributes to Modality-Specific Hyperalgesia in Mice. J. Neurosci. 2019, 39, 9954–9966. [Google Scholar] [CrossRef] [PubMed]
- Silverman, H.A.; Chen, A.; Kravatz, N.L.; Chavan, S.S.; Chang, E.H. Involvement of Neural Transient Receptor Potential Channels in Peripheral Inflammation. Front. Immunol. 2020, 11, 590261. [Google Scholar] [CrossRef] [PubMed]
- Gamse, R.; Lackner, D.; Gamse, G.; Leeman, S.E. Effect of capsaicin pretreatment on capsaicin-evoked release of immunoreactive somatostatin and substance P from primary sensory neurons. Naunyn. Schmiedebergs Arch. Pharmacol. 1981, 316, 38–41. [Google Scholar] [CrossRef]
- Long, J.B.; Rigamonti, D.D.; Dosaka, K.; Kraimer, J.M.; Martinez-Arizala, A. Somatostatin causes vasoconstriction, reduces blood flow and increases vascular permeability in the rat central nervous system. J. Pharmacol. Exp. Ther. 1992, 260, 1425–1432. [Google Scholar] [PubMed]
- Helyes, Z.; Pinter, E.; Sandor, K.; Elekes, K.; Banvolgyi, A.; Keszthelyi, D.; Szoke, E.; Toth, D.M.; Sandor, Z.; Kereskai, L.; et al. Impaired defense mechanism against inflammation, hyperalgesia, and airway hyperreactivity in somatostatin 4 receptor gene-deleted mice. Proc. Natl. Acad. Sci. USA 2009, 106, 13088–13093. [Google Scholar] [CrossRef] [Green Version]
- Szolcsanyi, J.; Helyes, Z.; Oroszi, G.; Nemeth, J.; Pinter, E. Release of somatostatin and its role in the mediation of the anti-inflammatory effect induced by antidromic stimulation of sensory fibres of rat sciatic nerve. Br. J. Pharmacol. 1998, 123, 936–942. [Google Scholar] [CrossRef] [Green Version]
- Sattler, K.; El-Battrawy, I.; Cyganek, L.; Lang, S.; Lan, H.; Li, X.; Zhao, Z.; Utikal, J.; Wieland, T.; Borggrefe, M.; et al. TRPV1 activation and internalization is part of the LPS-induced inflammation in human iPSC-derived cardiomyocytes. Sci. Rep. 2021, 11, 14689. [Google Scholar] [CrossRef]
- Wang, Y.; Cui, L.; Xu, H.; Liu, S.; Zhu, F.; Yan, F.; Shen, S.; Zhu, M. TRPV1 agonism inhibits endothelial cell inflammation via activation of eNOS/NO pathway. Atherosclerosis 2017, 260, 13–19. [Google Scholar] [CrossRef]
- Zhao, J.F.; Ching, L.C.; Kou, Y.R.; Lin, S.J.; Wei, J.; Shyue, S.K.; Lee, T.S. Activation of TRPV1 prevents OxLDL-induced lipid accumulation and TNF-alpha-induced inflammation in macrophages: Role of liver X receptor alpha. Mediators Inflamm. 2013, 2013, 925171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Choi, C.S.; Bae, I.H.; Choi, J.K.; Park, Y.H.; Park, M. A novel, topical, nonsteroidal, TRPV1 antagonist, PAC-14028 cream improves skin barrier function and exerts anti-inflammatory action through modulating epidermal differentiation markers and suppressing Th2 cytokines in atopic dermatitis. J. Dermatol. Sci. 2018, 91, 184–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Feng, C.; He, H.; He, J.; Wang, J.; Li, X.; Wang, S.; Li, W.; Hou, J.; Liu, T.; et al. Sensitization of TRPV1 receptors by TNF-alpha orchestrates the development of vincristine-induced pain. Oncol. Lett. 2018, 15, 5013–5019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ninomiya, Y.; Tanuma, S.I.; Tsukimoto, M. Differences in the effects of four TRPV1 channel antagonists on lipopolysaccharide-induced cytokine production and COX-2 expression in murine macrophages. Biochem. Biophys. Res. Commun. 2017, 484, 668–674. [Google Scholar] [CrossRef]
- Wang, Y.; Fu, X.; Huang, L.; Wang, X.; Lu, Z.; Zhu, F.; Xiao, Z. Increased Asics Expression via the Camkii-CREB Pathway in a Novel Mouse Model of Trigeminal Pain. Cell Physiol. Biochem. 2018, 46, 568–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rollyson, W.D.; Stover, C.A.; Brown, K.C.; Perry, H.E.; Stevenson, C.D.; McNees, C.A.; Ball, J.G.; Valentovic, M.A.; Dasgupta, P. Bioavailability of capsaicin and its implications for drug delivery. J. Control. Release. 2014, 196, 96–105. [Google Scholar] [CrossRef] [Green Version]
- Alothman, Z.A.; Wabaidur, S.M.; Khan, M.R.; Abdel Ghafar, A.; Habila, M.A.; Ahmed, Y.B. Determination of capsaicinoids in Capsicum species using ultra performance liquid chromatography-mass spectrometry. J. Sep. Sci. 2012, 35, 2892–2896. [Google Scholar] [CrossRef] [PubMed]
- Jarret, R.L.; Baldwin, E.; Perkins, B.; Bushway, R.; Guthrie, K. Diversity of Fruit Quality Characteristics in Capsicum frutescens. HortScience 2007, 42, 16–19. [Google Scholar] [CrossRef] [Green Version]
- Chaiyasit, K.; Khovidhunkit, W.; Wittayalertpanya, S. Pharmacokinetic and the effect of capsaicin in Capsicum frutescens on decreasing plasma glucose level. J. Med. Assoc. Thai. 2009, 92, 108–113. [Google Scholar]
- Babbar, S.; Marier, J.F.; Mouksassi, M.S.; Beliveau, M.; Vanhove, G.F.; Chanda, S.; Bley, K. Pharmacokinetic analysis of capsaicin after topical administration of a high-concentration capsaicin patch to patients with peripheral neuropathic pain. Ther. Drug Monit. 2009, 31, 502–510. [Google Scholar] [CrossRef] [PubMed]
- Steffee, C.H.; Lantz, P.E.; Flannagan, L.M.; Thompson, R.L.; Jason, D.R. Oleoresin capsicum (pepper) spray and "in-custody deaths". Am. J. Forensic. Med. Pathol. 1995, 16, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Lechner, A.; Alderson, T.; Gautam, S.; Flaker, G. Ventricular fibrillation due to coronary spasm after pepper spray. Pacing Clin. Electrophysiol. 2021, 44, 548–551. [Google Scholar] [CrossRef]
- Szolcsanyi, J.; Oroszi, G.; Nemeth, J.; Szilvassy, Z.; Blasig, I.E.; Tosaki, A. Functional and biochemical evidence for capsaicin-induced neural endothelin release in isolated working rat heart. Eur. J. Pharmacol. 2001, 419, 215–221. [Google Scholar] [CrossRef]
- Kark, T.; Bagi, Z.; Lizanecz, E.; Pasztor, E.T.; Erdei, N.; Czikora, A.; Papp, Z.; Edes, I.; Porszasz, R.; Toth, A. Tissue-specific regulation of microvascular diameter: Opposite functional roles of neuronal and smooth muscle located vanilloid receptor-1. Mol. Pharmacol. 2008, 73, 1405–1412. [Google Scholar] [CrossRef] [PubMed]
- Akcay, M.; Gedikli, O.; Yuksel, S. An unusual side effect of weight loss pills in a young man; acute myocardial infarction due to cayenne pepper pills. Anatol. J. Cardiol. 2017, 18, 310–311. [Google Scholar] [CrossRef]
- Rodney, E.; Parente, T.; Vasavada, B.C.; Sacchi, T.J. Life-threatening ventricular arrhythmias in patients with silent myocardial ischemia due to coronary-artery spasm. N. Engl. J. Med. 1992, 327, 956, author reply 957. [Google Scholar]
- Pedersen, F.; Pietersen, A.; Sandoe, E. Silent myocardial ischaemia and life threatening ventricular arrhythmias. Ann. Clin. Res. 1988, 20, 404–409. [Google Scholar]
- Myerburg, R.J.; Kessler, K.M.; Mallon, S.M.; Cox, M.M.; deMarchena, E.; Interian, A., Jr.; Castellanos, A. Life-threatening ventricular arrhythmias in patients with silent myocardial ischemia due to coronary-artery spasm. N. Engl. J. Med. 1992, 326, 1451–1455. [Google Scholar] [CrossRef]
- Mustafa, S.; Ismael, H.N. Ethanol potentiates heat response in the carotid artery via TRPV1. Life Sci. 2017, 188, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Van Gerven, L.; Steelant, B.; Cools, L.; Callebaut, I.; Backaert, W.; de Hoon, J.; Ampe, E.; Talavera, K.; Hellings, P.W. Low-dose capsaicin (0.01 mM) nasal spray is equally effective as the current standard treatment for idiopathic rhinitis: A randomized, double-blind, placebo-controlled trial. J. Allergy Clin. Immunol. 2021, 147, 397–400. [Google Scholar] [CrossRef]
- Simpson, D.M.; Robinson-Papp, J.; Van, J.; Stoker, M.; Jacobs, H.; Snijder, R.J.; Schregardus, D.S.; Long, S.K.; Lambourg, B.; Katz, N. Capsaicin 8% Patch in Painful Diabetic Peripheral Neuropathy: A Randomized, Double-Blind, Placebo-Controlled Study. J. Pain 2017, 18, 42–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, S.; Simpson, D.M.; Moyle, G.; Brew, B.J.; Schifitto, G.; Larbalestier, N.; Orkin, C.; Fisher, M.; Vanhove, G.F.; Tobias, J.K. NGX-4010, a capsaicin 8% patch, for the treatment of painful HIV-associated distal sensory polyneuropathy: Integrated analysis of two phase III, randomized, controlled trials. AIDS Res. Ther. 2013, 10, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mudau, M.; Genis, A.; Lochner, A.; Strijdom, H. Endothelial dysfunction: The early predictor of atherosclerosis. Cardiovasc. J. Afr. 2012, 23, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Jensen, H.A.; Mehta, J.L. Endothelial cell dysfunction as a novel therapeutic target in atherosclerosis. Expert. Rev. Cardiovasc. Ther. 2016, 14, 1021–1033. [Google Scholar] [CrossRef] [PubMed]
- Gimbrone, M.A., Jr.; Garcia-Cardena, G. Endothelial Cell Dysfunction and the Pathobiology of Atherosclerosis. Circ. Res. 2016, 118, 620–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansson, G.K. Inflammatory mechanisms in atherosclerosis. J. Thromb. Haemost. 2009, 7 (Suppl. 1), 328–331. [Google Scholar] [CrossRef]
- Berg, A.H.; Scherer, P.E. Adipose tissue, inflammation, and cardiovascular disease. Circ. Res. 2005, 96, 939–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Munjuluri, S.; Wilkerson, D.A.; Sooch, G.; Chen, X.; White, F.A.; Obukhov, A.G. Capsaicin and TRPV1 Channels in the Cardiovascular System: The Role of Inflammation. Cells 2022, 11, 18. https://doi.org/10.3390/cells11010018
Munjuluri S, Wilkerson DA, Sooch G, Chen X, White FA, Obukhov AG. Capsaicin and TRPV1 Channels in the Cardiovascular System: The Role of Inflammation. Cells. 2022; 11(1):18. https://doi.org/10.3390/cells11010018
Chicago/Turabian StyleMunjuluri, Sreepadaarchana, Dru A. Wilkerson, Gagandeep Sooch, Xingjuan Chen, Fletcher A. White, and Alexander G. Obukhov. 2022. "Capsaicin and TRPV1 Channels in the Cardiovascular System: The Role of Inflammation" Cells 11, no. 1: 18. https://doi.org/10.3390/cells11010018
APA StyleMunjuluri, S., Wilkerson, D. A., Sooch, G., Chen, X., White, F. A., & Obukhov, A. G. (2022). Capsaicin and TRPV1 Channels in the Cardiovascular System: The Role of Inflammation. Cells, 11(1), 18. https://doi.org/10.3390/cells11010018