Factors Affecting Polyphenol Biosynthesis in Wild and Field Grown St. John’s Wort (Hypericum perforatum L. Hypericaceae/Guttiferae)


Abstract
:Contents
- Introduction
- Genetic variability: chemotaxonomy, phenotypic variations and breeding within Hypericum genus; a focus on H. perforatum L.
- Ontogenetic stage and seasonal variation
- Agricultural practices: irrigation, soil, fertilization
- Biotic and abiotic stress
- Post-Harvest and stability
- Conclusive remarks
- References
1. Introduction
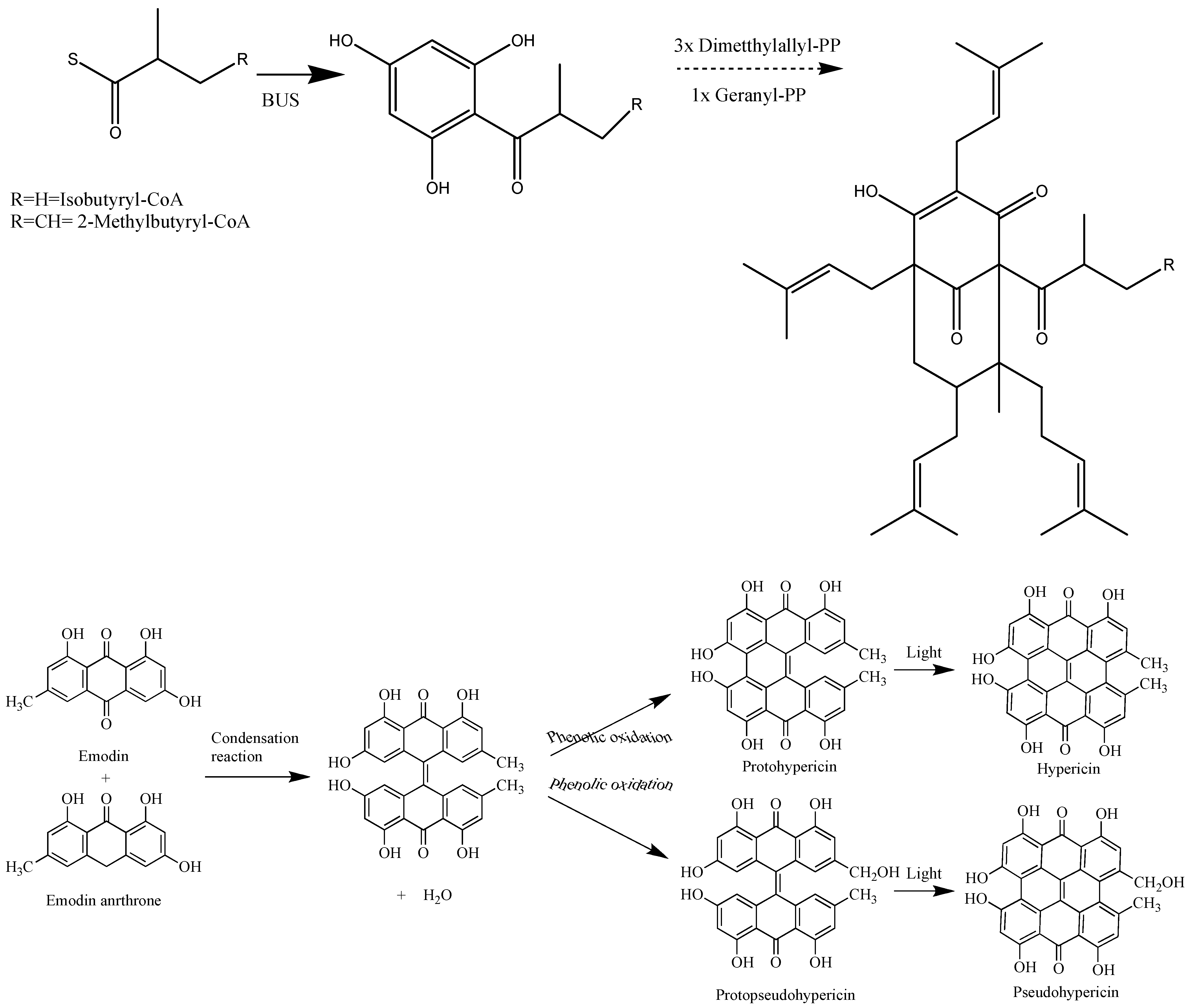

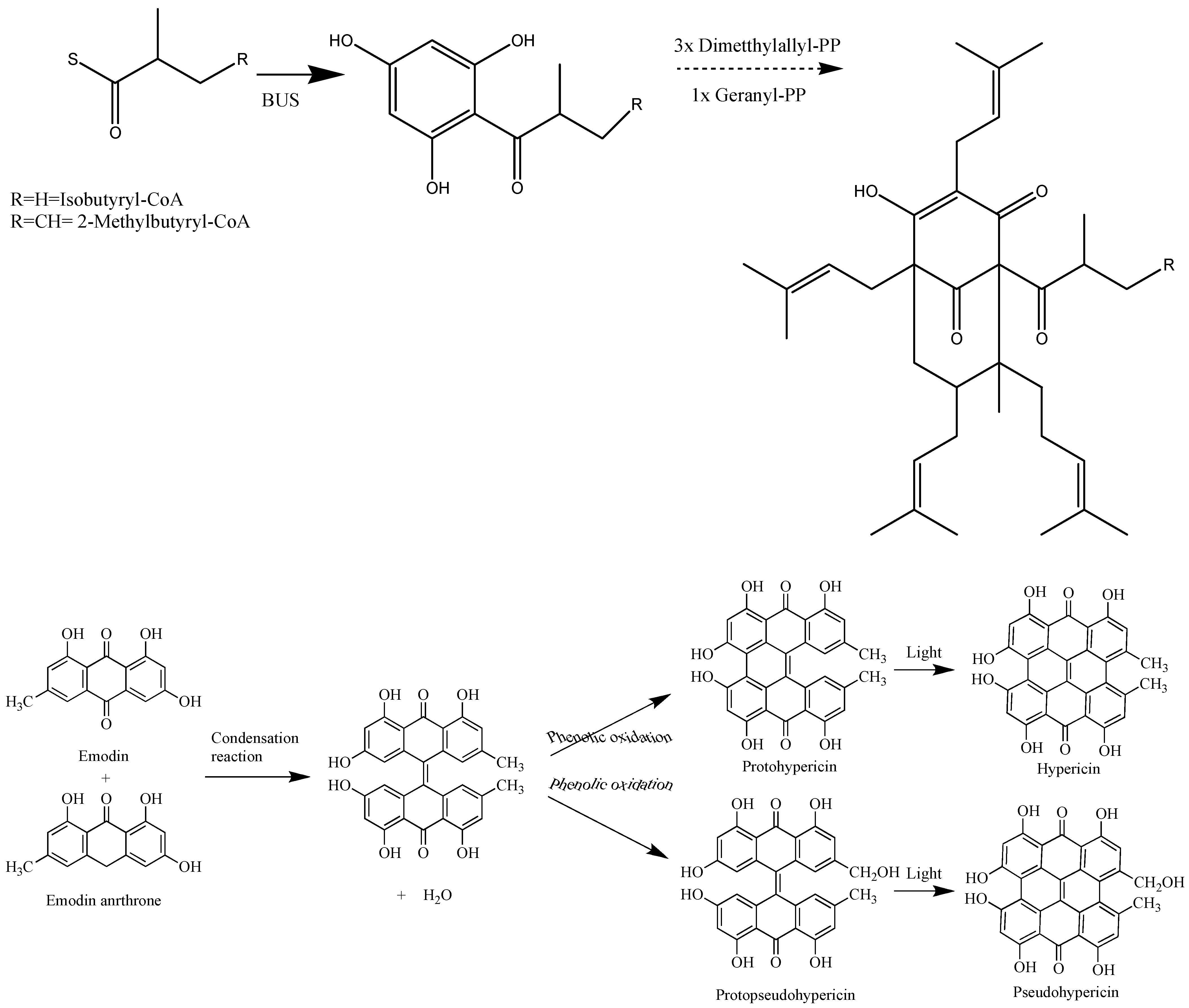{kind=link}
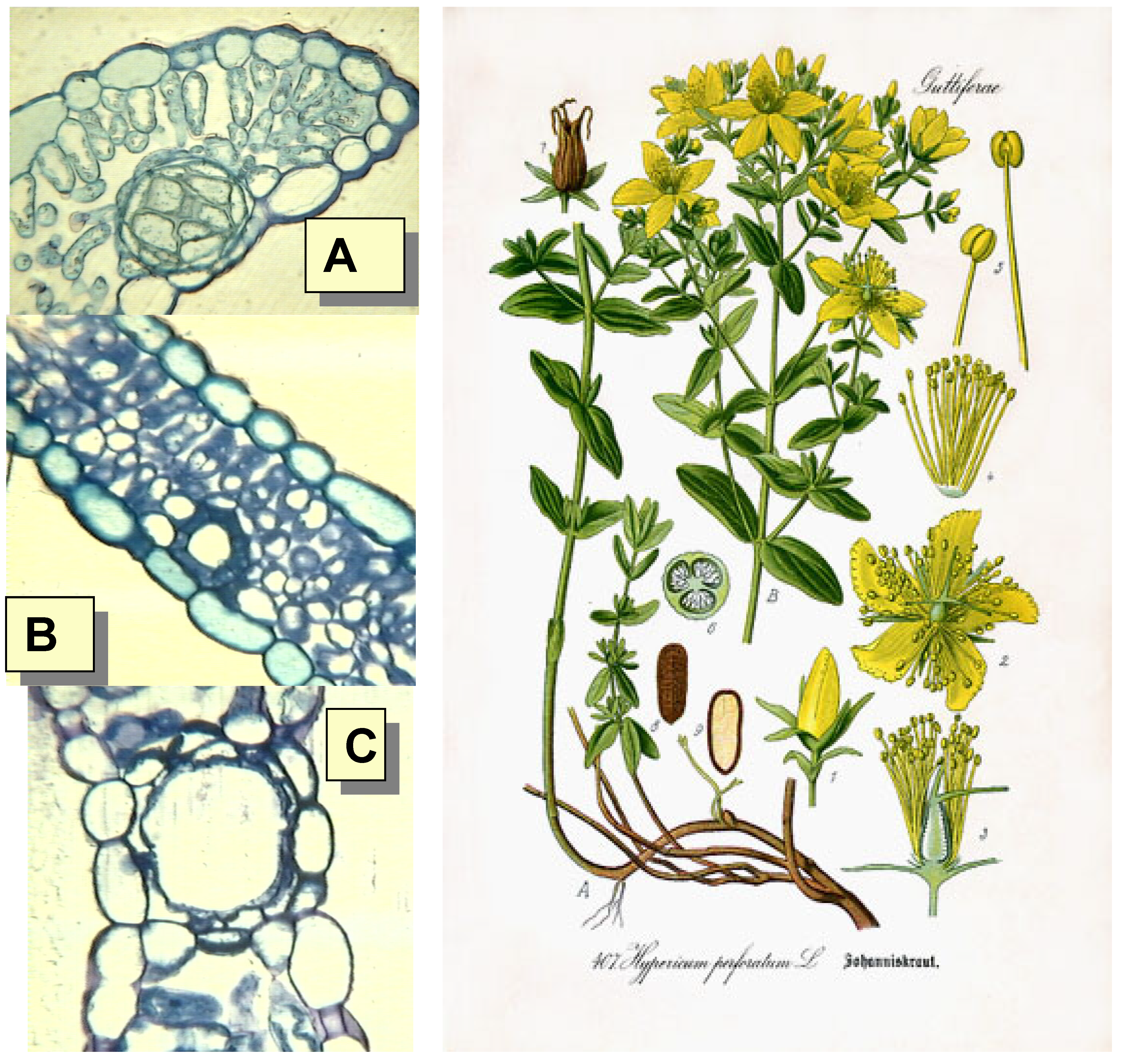{kind=link}
| Substance | Approximate amount (mg/g dry weight) |
|---|---|
| Naphtodianthrones | |
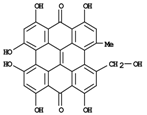
| Hypericin | 0.1-7a |
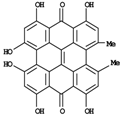 | |
| Pseudohypericin | 0.1-12a |
 | |
| Localization | aDark glands in leaf and petal margin; stamens |
| Acylphloroglucinols | |
| Hyperforin | 0.3-150b |
 | |
| Localization | bTranslucent glands in leaves, carpellar leaves and sepals |
| Flavonoids | |
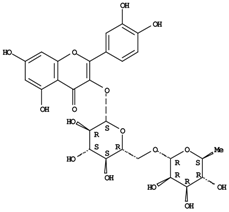
| Hyperoside | 1-25c |
 | |
| Rutin | 0-35c |
 | |
| Localization | cFloral dehiscent leaves: sepals, stamens, petals. Likely accumulation in vacuoles. |
| Biflavones | |
| Amentoflavone | 0-1.8d |
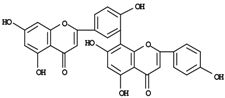 | |
| Biapigenin | 0.3-10.2d |
 | |
| Localization | dFloral deiscent leaves: sepals, stamens, petals. Likely accumulation in vacuoles. |

2. Genetic variability: chemotaxonomy, phenotypic variations and breeding within Hypericum genus; a focus on H. perforatum L.
2.1. Hypericum genus: chemodiversity
2.2. Hypericum perforatum: chemodiversity in the wild
| Origin/Cultivar | Amount1 | Method | Notes | Ref. | ||
|---|---|---|---|---|---|---|
| Hpf2 | Hyp3 | Pse4 | ||||
| North American | ||||||
| Wild - Canada, BC | nm5 | 0.33-0.63 | nm | HPLC | Aerial parts at full flowering | [52] |
| Wild - Canada, Nova Scotia | nm | 0.12-0.29 | nm | HPLC | Aerial parts at full flowering | [52] |
| Wild - Canada, Ontario | nm | 0.13-0.27 | nm | HPLC | Aerial parts at full flowering | [52] |
| Wild - Canada, Oregon | nm | 0.24-0.54 | nm | HPLC | Aerial parts at full flowering | [52] |
| Wild - USA, California | nm | 0.11-0.44 | 2.78-4.27 | HPLC | Aerial parts Higher values in flowers | [50,51] |
| Wild - USA, Montana | nm | 0.05-0.18 | 0.76-1.07 | HPLC | Aerial parts Higher values in flowers | [50,51] |
| Wild - USA, Oregon | nm | 0.37-3.87* | nm | UV6 | Aerial parts * Flowers only | [49] |
| Wild - USA, Oregon | nm | 0.60 mg/g | 2.86 mg/g | HPLC | Aerial parts Higher values in flowers | [50,51] |
| Cultivated – USA | *27.37 **14.01 | *1.46 **0.38 | *2.51 **0.71 | HPLC | *Flowers ** Leaves | [114] |
| South American | ||||||
| Cultivated – Brazil | nm | 3.83 | nm | HPLC | Leaves, stems, vegetative stage | [150] |
| Cultivated - Chile | nm | 1.1-1.28 | nm | UV | Aerial parts at full flowering | [171] |
| Cultivated - Chile | nm | 1.9-2.5 | nm | UV | Flowers | [53] |
| European and Middle East | ||||||
| Wild - Armenia | 150 | 0.23-0.05 | nm | HPLC | Low variability of samples collected from different sites. Unspecified aerial parts | [65] |
| Wild - Armenia | nm | 0.06-0.64 | 0.2-1.6 | HPLC | FlowersFrom 10 different regions | [66] |
| Wild - Bulgaria | nm | 1.25 | nm | UV | Unspecified aerial parts | [36] |
| Wild - Croatiassp. perforatum | nm | 0.3 | nm | HPLC | Unspecified aerial parts. Richer in flavonoids | [42] |
| Wild - Croatia,ssp. agustifolium | nm | 1 | nm | HPLC | Unspecified aerial parts. Devoid of rutin | [42] |
| Wild - Greece (Crete Island) | nm | *1.1 **0.25 | *1.3 **0.17 | HPLC | *Flowers **Leaves and petioles | [67] |
| Wild - India | 1.66-4.62 | 1.19-2.68 | 13.74-41.64 | HPLC | Aerial parts | [72] |
| Wild - Iran | 1.01 | nm | UV | Aerial parts af full flowering | [168] | |
| Wild - Italy | *12.4-21.1 **31.5-41.5 | *2.8-6.6 **0.6-2.9 | *5.2-10.8 **2.4-6.2 | HPLC | *Dried flowers only **Fruits. Samples from 5 different italian regions | [58] |
| Wild - Italy | 8.3-31.81 | 8.34-14.79 | nm | HPLC | Aeria parts at full flowering ssp. angustifolium and ssp. perforatum evidenced both genetical and ecological differences | [60] |
| Wild – Italy | *10.72-23.32 **14.55-24.26 | *0.35-0.52 **1.12-1.72 | nm | HPLC | * ssp. Perforatum ** ssp. Veronense Aerial parts at full flowering | [173] |
| Wild - Italy (Tuscany) | 0.84-4.82 | 0.02-0.11 | 0.03-0.33 | HPLC | Flowers only. Chemotype devoid of rutin | [57] |
| Wild - Italy (Tuscany) | 82.5-52 101.4-109.5 fw 7 | *0.03-0.2 | nm | HPLC | Flowering tops *Expressed as total naphtodianthrones | [151] |
| Wild - Russia (Udmurtia) | nm | 12 fw | nm | UV | Flower buds | [63] |
| Wild - Serbia | 3.55 | 0.17 | nm | HPLC | Aerial parts at full flowering | [64] |
| Wild - Serbia | 1.52 | 0.2 | 0.5 | HPLC | Aerial parts at full flowering | [172] |
| Wild - Solvenia | 13.59 | 5.20 | 5.22 | HPLC | Aerial parts at full flowering | [55] |
| Wild - Turkey | nm | *0-**2.73 | nm | HPLC | * Stems ** Flowers | [100] |
| Wild - Turkey | nm | 0.44-2.82 | nm | HPLC | Aerial parts at full flowering | [62] |
| Wild - Turkey | nm | 2.15-2.46 | nm | UV | Aerial parts at full flowering | [93] |
| Wild - Turkey | nm | 0.28-4.46 | nm | UV | Aerial parts at full flowering | [61] |
| Cultivated - Denmark | 3 fw7 | nm | nm | HPLC | Leaves only | [92] |
| Cultivated - Germany | 2 | 0.1-5.2 | 0.1-16 | HPLC | Flowering tops | [6] |
| Cultivated - Germany cv. Hyperiflor | 12.3 | 0.8 | nm | HPLC | Aerial parts af full flowering | [136] |
| Cultivated - Germany cv. Hyperimed | 26.51 | 0.87 | nm | HPLC | Aerial parts at full flowering | [136] |
| Cultivated - Germany cv. Topaz | 12.81 | 1.11 | nm | HPLC | Aerial parts at full flowering | [136] |
| Cultivated - Hungary | nm | 0.10-2.648 | nm | HPLC | Aerial parts at full flowering Selected breeding lines. | [7] |
| Cultivated - Hungary cv Topas and selected breeding lines | nm | 0.64-3.88 | 3.18-11.88 | HPLC | Aerial parts at full flowering | [43] |
| Cultivated - HungaryHybrids selection | nm | 1.52-11.9 fw7 | nm | UV | Reproductive parts. | [68] |
| Cultivated - Italy cv. Zorzi | nm | 1.36 | nm | HPLC | Aerial parts at full flowering | [139] |
| Cultivated - Lithuania | nm | 0.23-1.24 | nm | HPLC | Flowering tops Seeds from 21 wild accessions | [54] |
| Cultivated - Poland | nm | 7-9 | nm | HPLC | Aerial parts at full flowering | [169] |
| Cultivated - Slovakia | nm | 1.59 | nm | UV | Aerial parts at full flowering 4th generation of selected ex-vitro plants from regenerated in vitro somaclones | [48] |
| Cultivated - Slovakia | *91.6-107.5 | **1.1-5.8 | ***0.7-6.9 | HPLC | *Isolated unripe fruits **Buds Fluctuation during ontogenesis Samples devoid of rutin | [94] |
| Cultivated - Switzerland | *0.9-6.4 | **0.9-6.1 | nm | HPLC | *Maximum in flowers with open petals **Maximum in closed buds | [170] |
| Cultivated - Switzerland | nm | 0.23-1.72 | 0.81-3.27 | HPLC | Aerial parts at full flowering Screening of 24 accessions | [69] |
| Cultivated -Switzerland | nm | 0.34-1.08 | 1.25-2.36 | HPLC | Aerial parts at full flowering cv Topas, Hyperimed, Elixir | [69] |
| Oceania | ||||||
| Wild - Australia | nm | 0.12-0.8 | nm | HPLC | Aerial parts at full flowering | [52] |
| Wild - Australia | nm | 1,02-4.84 | nm | UV | Leaves only | [71] |
| Wild - Australia | nm | *1.2-2.35 **2.97-5.02 | nm | UV | * Broadleaved ** Narrowleaved Aerial parts at full flowering | [41] |
| Greenhouse or controlled environment | ||||||
| Controlled Environment | *3.13-7.39 fw **8.46-13.91 fw | *0.03-0.09 fw **0.75-1.18 fw | *0.05-0.16 fw **0.98-1.47 fw | HPLC | *Shoots **Flower buds | [39] |
| Controlled Environment | 30 | 1.3 | 2.8 | HPLC | Leaves | [112] |
| Controlled Environment | nm | 18-38 | 42-115 | HPLC | Isolated flowers | [89] |
| Greenhouse Cultivated | *0.65-3.17 **1.97-9.00 | *0.57-3.93 **2.31-6.72 | *1.07-4.10 **1.13-3.97 | HPLC | * Flowering top ** Flowers. Seeds wild-collected in France and Switzerland. | [59] |
| Greenhouse Cultivated cv. Anthos | 5.09 | 0.09 | 0.11 | HPLC | Unspecified aerial parts | [65] |
| Greenhouse Cultivated cv. New Stem | 3.4 | 0.09 | 0.11 | HPLC | Unspecified aerial parts | [65] |
| Greenhouse Cultivated cv Topaz | 2.92 | 0.06 | 0.11 | HPLC | Unspecified aerial parts | [65] |
| Greenhouse Cultivated cv. Topaz | *1.89 **4.45 | *0.82-2.10 **1.31-5.09 | nm | HPLC | * Flowering top ** Flowers Seeds wild-collected in France and Switzerland. | [59] |
| Greenhouse Cultivated cv. Topaz | nm | *0.23-0.39 **0.29-0.59 | *0.44-0.89 **0.71-1.52 | HPLC | *5 cm tops ** Flowers only | [96] |
2.2.1. Chemodiversity – American populations
2.2.2. Chemodiversity – European and Middle East populations
2.2.3. Chemodiversity – Asiatic and Oceanian populations
2.2.4. Future developments
3. Phenolic content during ontogenetic stage and seasonal variation
3.1. Ontogenetic stage
3.2. Seasonal variation
3.3. Future developments
4. Agricultural practices ( irrigation, soil, fertilization)
4.1. Water availability
4.2. Altitude
4.3. Fertilization and nutrient supply
4.6. Future developments
5. Biotic and abiotic stress
5.1. Metal contamination
5.2. Pathogens and herbivores
5.4. Temperature
5.3. Future developments
6. Post-harvest and stability
6.1. Future developments
7. Conclusions
7.1. Focusing breeding and agronomic practices on phenolic metabolism
7.2. Standardized quantifications
7.3. Standardized production
7.4. Provision of more knowledge about hyperforin and other bioactive constituents
References
- Canter, P.H.; Thomas, H.; Ernst, E. Bringing medicinal plants into cultivation: opportunities and challenges for biotechnology. Trends Biotechnol. 2005, 23, 180–185. [Google Scholar] [CrossRef]
- Bombardelli, E.; Bombardelli, V. Twenty years’ experience in the botanical health food market. Fitoterapia 2005, 76, 495–507. [Google Scholar] [CrossRef]
- Camire, M.E.; Kantor, M.A. Dietary Supplements: nutritional and legal considerations. Food Technol. 1999, 53, 87–96. [Google Scholar]
- Grunwald, J. The world market for hypericum products. Nutraceuticals World 1999, 5/6, 22–25. [Google Scholar]
- Gaudin, M.; Simonnet, X.; Debrunner, F. Colletotrichum gloeosporioides as the cause of St. John’s wort (Hypericum perforatum) dieback in Switzerland and breeding for a tolerant variety. In Hypericum; Ernst, E., Ed.; Taylor&Francis: London, UK, 2003; pp. 23–42. [Google Scholar]
- Franke, R.; Schenk, R.; Bauermann, U. Variability in Hypericum perforatum L. breeding lines. Acta Hort. 1999, 502, 167–174. [Google Scholar]
- Pluhar, Z.S.; Bernàth, J.; Neumayer, E. Morphological, Production biological and chemical diversity of St. John’s Wort (Hypericum perforatum L.). Acta Hort. 2002, 576, 33–40. [Google Scholar]
- Hoelzl, J.; Petersen, M. Chemical constituents of Hypericum spp. In Hypericum; Ernst, E., Ed.; Taylor&Francis: London, UK, 2003; pp. 77–93. [Google Scholar]
- Murch, S.J.; Saxena, P.K. A melatonin-rich germplasm line of St. John's wort (Hypericum perforatum L.). J. Pineal Res. 2006, 41, 284–287. [Google Scholar] [CrossRef]
- Beerhues, L. Hyperforin. Phytochemistry 2006, 67, 2201–2207. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Verpoorte, R.; Suresh, B. Evaluation of in-vivo wound healing activity of Hypericum patulum (Family: Hypericaceae) leaf extract on different wound model in rats. J. Ethnopharmacol. 2000, 70, 315–321. [Google Scholar] [CrossRef]
- Reichling, J.; Weseler, A.; Saller, R. A current review of the antimicrobial activity of Hypericum perforatum L. Pharmacopsychiatry 2001, 34, 116–118. [Google Scholar] [CrossRef]
- Jacobson, J.M.; Feinman, L.; Liebes, L.; Ostrow, N.; Koslowski, V.; Tobia, A.; Cabana, B.E.; Lee, D.-H.; Spritzler, J.; Prince, A.M. Pharmacokinetics, safety, and antiviral effects of hypericin, a derivative of St. John's wort plant, in patients with chronic hepatitis C virus infection. Antimicrob. Agents 2001, 45, 517–524. [Google Scholar] [CrossRef]
- Schinazi, R.F.; Chu, C.K.; Babu, J.R.; Oswald, B.J.; Saalmann, V.; Cannon, D.L.; Eriksson, B.F.; Nasr, M. Anthraquinones as a new class of antiviral agents against human immunodeficiency virus. Antivir. Res. 1990, 13, 265–272. [Google Scholar] [CrossRef]
- Oblozinsky, M.; Bezakova, L.; Holkova, I.; Vanko, M.; Kartnig, T.; Psenak, M. Antilipoxigenase activity of compounds from Hypericum perforatum. Biologia. 2006, 61, 331–332. [Google Scholar] [CrossRef]
- Schempp, C.M.; Simon-Haarhaus, B.; Simon, J.C. Phototoxic and apoptosis-inducing capacity of pseudohypericin. Planta Med. 2002, 68, 171–173. [Google Scholar] [CrossRef]
- Stavropoulos, N.E.; Kim, A.; Nseyo, U.U.; Tsimaris, I.; Chung, T.D.; Miller, T.A.; Redlak, M.; Nseyo, U. O.; Skalkos, D. Hypericum perforatum L. extract - Novel photosensitizer against human bladder cancer cells. J. Photoch. Photobio. B Biology 2006, 84, 64–69. [Google Scholar] [CrossRef]
- Martarelli, D.; Martarelli, B.; Pediconi, D.; Nabissi, M. I.; Perfumi, M.; Pompei, P. Hypericum perforatum methanolic extract inhibits growth of human prostatic carcinoma cell line orthotopically implanted in nude mice. Cancer Lett. 2004, 210, 27–33. [Google Scholar] [CrossRef]
- Rezvani, A.H.; Overstreet, D.H.; Perfumi, M.; Massi, M. Plant derivatives in the treatment of alcohol dependency. Pharmacol. Biochem. Behav. 2003, 75, 593–606. [Google Scholar] [CrossRef]
- Kiesslich, T.; Krammer, B.; Plaetzer, K. Cellular mechanisms and prospective applications of hypericin in photodynamic therapy. Curr. Med. Chem. 2006, 13, 2189–2204. [Google Scholar] [CrossRef]
- Whitten, D.L.; Myers, S.P.; Hawrelak, J.A.; Wohlmuth, H. The effect of St John's wort extracts on CYP3A: a systematic review of prospective clinical trials. Brit. J. Clin. Pharmacol. 2006, 62, 512–526. [Google Scholar] [CrossRef]
- Schempp, C.M.; Windeck, T.; Hezel, S.; Simon, J.C. Topical treatment of atopic dermatitis with St. John's wort cream - a randomized, placebo controlled, double blind half-side comparison. Phytomedicine 2003, 10, 31–37. [Google Scholar] [CrossRef]
- Grundmann, O.; Kelber, O.; Butterweck, V. Effects of St. John's wort extract and single constituents on stress-induced hyperthermia in mice. Planta Med. 2006, 72, 1366–1371. [Google Scholar] [CrossRef]
- Laggner, H.; Schreier, S.; Hermann, M.; Exner, M.; Muehl, A.; Gmeiner, B.M.K.; Kapiotis, S. The main components of St John's Wort inhibit low-density lipoprotein atherogenic modification: a beneficial side effect of an OTC antidepressant drug? Free Radical Res. 2007, 41, 234–241. [Google Scholar] [CrossRef]
- Barnes, J.; Anderson, L.A.; Phillipson, J.D. St John's wort (Hypericum perforatum L.): a review of its chemistry, pharmacology and clinical properties. J. Pharm. Pharmacol. 2001, 53, 583–600. [Google Scholar] [CrossRef]
- Linde , K.; Berner, M.M; Kriston, L. St John's wort for major depression. Cochrane Database Syst. Rev. 2008, 4, 1–147. [Google Scholar]
- Medina, M.A.; Martinez-Poveda, B.; Amores-Sanchez, M.I.; Quesada, A.R. Hyperforin: More than an antidepressant bioactive compound? Life Sci. 2006, 79, 101–111. [Google Scholar]
- Nöldner, M.; Schötz, K. Rutin is essential for the antidepressant activity of Hypericum perforatum extracts in the forced swimming Test. Planta Med. 2002, 68, 577–580. [Google Scholar] [CrossRef]
- Jensen, A.G.; Hansen, S.H.; Nielsen, E.O. Adhyperforin as a contributor to the effect of Hypericum perforatum L. in biochemical models of antidepressant activity. Life Sci. 2001, 68, 1593–1605. [Google Scholar] [CrossRef]
- Baureithel, K.H.; Büter, K.B.; Engesser, A.; Burkard, W.; Schaffner, W. Inhibition of benzodiazepine binding in vitro by amentoflavone, a constituent of various species of Hypericum. Pharm. Acta Helv. 1997, 72, 153–157. [Google Scholar] [CrossRef]
- USP. Monograph (565) Botanical Extracts. Pharmacopoeial Forum 1999, 25, 7880–7883. [Google Scholar]
- Becker, H. Boosting the quality and potency of St. John's-wort. Agric. Res. Mag. 2000, 48, 12–13. [Google Scholar]
- Buter, B.; Orlacchio, C.; Soldati, A.; Berger, K. Significance of genetic and environmental aspects in the field cultivation of Hypericum perforatum. Planta Med. 1998, 64, 431–437. [Google Scholar] [CrossRef]
- Mathis, C.; Ourisson, G. Etude chemo-taxonomique du genre Hypericum. Repartition de l’hypericine. Phytochemistry 1964, 2, 157–171. [Google Scholar] [CrossRef]
- Crockett, S.L.; Schaneberg, B.; Khan, I.A. Phytochemical profiling of new and old world Hypericum (St. John’s wort) species. Phytochem. Anal. 2005, 16, 479–485. [Google Scholar] [CrossRef]
- Kitanov, M.K. Hypericin and pseudohypericin in some Hypericum species. Biochem. Syst. Ecol. 2001, 29, 171–178. [Google Scholar] [CrossRef]
- Radusiene, J.; Bagdonaite, E. Phenotypic variation in Hypericum perforatum L. and H. maculatum Crantz wild populations in Lithuania. J. Herb Spi. Med. Plants 2002, 9, 345–351. [Google Scholar] [CrossRef]
- Kosuth, J.; Koperdakova, J.; Tolonen, A.; Hohtola, A.; Cellarova, E. The content of hypericins and phloroglucinols in Hypericum perforatum L. seedlings at early stage of development. Plant Sci. 2003, 165, 515–521. [Google Scholar] [CrossRef]
- Couceiro, M.A.; Afreen, F.; Zobayed, S.M.A.; Kozai, T. Variation in concentrations of major bioactive compounds of St. John’s Wort: Effects of harvesting time, temperature and germplasm. Plant Sci. 2006, 170, 128–134. [Google Scholar] [CrossRef]
- Campbell, M.H.; Delfosse, E.S. Biology of Australian weeds. 13. Hypericum perforatum L. J. Austr. Inst. Agric. Sci. 1984, 50, 63–73. [Google Scholar]
- Southwell, I.A.; Campbell, M.H. Hypericin content variation in Hypericum perforatum in Australia. Phytochemistry 1990, 30, 475–478. [Google Scholar] [CrossRef]
- Males, Z.; Brantner, A.H.; Sovic, K.; Pilepic, H.K.; Plazibat, M. Comparative phytochemical and antimicrobial investigation on Hypericum perforatum L. subsp. perforatum and H. perforatum subsp. angustifolium (DC) Gaudin. Acta Pharm. 2006, 56, 359–367. [Google Scholar]
- Pluhar, Z.; Bernath, J.; Nemeth, E. Investigations on the intraspecific variability of Hypericum perforatum L. J. Herbs, Spi. Med. Plants 2002, 2/3, 83–88. [Google Scholar]
- Martonfi, P.; Repcak, M.; Mihokova, L. Hypericum maculatum Crantz subsp. maculatum x H. perforatum L. (Hypericaceae): corroboration of natural hybridisation in hypericum by secondary metabolite analysis. Folia Geobot. Phytotax. 1996, 31, 245–250. [Google Scholar] [CrossRef]
- Schulte, J.; Schaffner, W. Kreuzungsexperimente mit verschiedenen Arten der gattung Hypericum. Z. Arzn. Gew. Pfl. 1999, 4, 126–133. [Google Scholar]
- Cellarova, E.; Koperdakova, J.; Brutovska, R. Genetic background of hypericin production in St. John’s wort (Hypericum perforatum L.) somaclones and their progenies. Abstracts of world conference on medicinal and aromatic plants. Budapest. Hungary 2001, 101. [Google Scholar]
- Cellarova, E.; Brutovska, R.; Daxnerova, Z.; Brunakova, K.; Weigel, R.C. Correlation between hypericin content and the ploidy of somaclones of Hypericum perforatum L. Acta Biotechnol. 1997, 17, 83–90. [Google Scholar] [CrossRef]
- Koperdakova, J.; Kosuth, J.; Cellarova, E. Variation in the content of hypericins in four generations of seed progeny of Hypericum perforatum somaclones. J. Plant Res. 2007, 120, 123–128. [Google Scholar] [CrossRef]
- Constantine, G.H.; Karchesy, J. Variations in hypericin concentrations in Hypericum perforatum L. and commercial products. Pharm. Biol. 1998, 36, 365–367. [Google Scholar] [CrossRef]
- Walker, L.; Sirvent, T.; Gibson, D.; Vance, N. Regional differences in hypericin and pseudohypericin concentrations and five morphological traits among Hypericum perforatum plants in the northwesten United States. Can. J. Bot. 2001, 79, 1248–1255. [Google Scholar] [CrossRef]
- Sirvent, T.M.; Walker, L.; Vance, N.; Gibson, D.M. Variation in hypericins from wild populations of Hypericum perforatum L. in the Pacific Northwest of the U.S.A. Econ. Bot. 2002, 56, 41–48. [Google Scholar] [CrossRef]
- Jensen, K.I.N.; Gaul, S.O.; Specht, E.G.; Doohan, D.J. Hypericin content of Nova Scotia biotypes of Hypericum perforatum L. Can. J Plant Sci. 1995, 75, 923–926. [Google Scholar] [CrossRef]
- Hevia, F.; Berti, M.; Wilckens, R.; Cifuentes, P. Quality and yield in St. John’s Wort (Hypericum perforatum L.) harvested in different phenological stages. Acta Agron. Hung. 2002, 50, 349–358. [Google Scholar] [CrossRef]
- Badgonaite, E.; Valdimaras, J.; Ivanauskas, L.; Labokas, J. Ex situ studies on chemical and morphological variability of Hypericum perforatum L. in Lithuania. Biologija 2007, 53, 63–70. [Google Scholar]
- Umek, A.; Kreft, S.; Kartnig, T.; Heydel, B. Quantitative phytochemical analyses of six Hypericum species growing in Slovenia. Planta Med. 1999, 65, 388–390. [Google Scholar] [CrossRef]
- Brantner, A.; Kartnig, T.; Ouehenberger, F. Vergleichende phytochemische Untersuchungen an Hypericum perforatum L. und Hypericum maculatum Crantz. Sci. Pharm. 1994, 62, 261–276. [Google Scholar]
- Martonfi, P.; Repcak, M.; Ciccarelli, D.; Garbari, F. Hypericum perforatum L. – Chemotype without rutin from Italy. Biochem. Syst. Ecol. 2001, 29, 659–661. [Google Scholar] [CrossRef]
- Pietta, P.; Gardana, C.; Pietta, A. omparative evaluation of St. John’s Wort from different italian regions. Farmaco 2001, 56, 491–496. [Google Scholar] [CrossRef]
- Poutaraud, A.; Girardin, P. Agronomic and chemical characterization of 39 Hypericum perforatum accessions between 1998 and 2000. Plant Breeding 2004, 123, 480–484. [Google Scholar] [CrossRef]
- Maggi, F.; Ferretti, G.; Pocceschi, N.; Menghini, L.; Ricciutelli, M. Morphological, histochemical and phytochemical investigation of the genus Hypericum of the Central Italy. Fitoterapia 2004, 75, 702–711. [Google Scholar] [CrossRef]
- Cirak, C.; Ayan, A.K.; Kevseroglu, K.; Ozen, T. Variation of hypericin in St. John’s wort (Hypericum perforatum) from wild populations of northern Turkey. Acta Bot. Hung. 2006, 48, 55–64. [Google Scholar] [CrossRef]
- Cirak, C.; Radusiene, J.; Karabuk, B.S.; Janulis, V. Variation of bioactive substances and morphological traits in Hypericum perforatum populations from Northern Turkey. Biochem. Syst. Ecol. 2007, 35, 403–409. [Google Scholar] [CrossRef]
- Kireeva, T.B.; Sharanov, U.L.; Letchamo, W. Biochemical and Eco-physiological studies on Hypericum spp. In Perspectives on new crops and new uses; Janick, J., Ed.; ASHS Press: Alexandria, VA, USA, 1999; pp. 467–468. [Google Scholar]
- Smelcerovic, A.; Spiteller, M. Phytochemical analysis of nine Hypericum L. species from Serbia and Macedonia. Pharmazie 2006, 61, 251–252. [Google Scholar]
- Kirakosyan, A.; Gibson, D.M.; Sirvent, T. A comparative study of Hypericum perforatum plants as sources of hypericins and hyperforins. J. Herbs Spi. Med. Plants 2003, 10, 73–88. [Google Scholar] [CrossRef]
- Melikian, E.; Boroyan, R.; Karaguezian, A.; Charchoglian, A.; Gabrielian, E.; Panossian, A. Hypericin content in St. John’s Wort (Hypericum perforatum L.) growing in Armenia. Pharm. Pharmacol. Lett. 1998, 3, 101–102. [Google Scholar]
- Xenophontos, M.; Stavropoulos, E.; Avramakis, E.; Navakoudis, E.; Dornemann, D.; Kotzabasis, K. Influence of the developmental stage on the (proto)hypericin and (proto)pseudohypericin levels of Hypericum plants from Crete. Planta Med. 2007, 73, 1309–1315. [Google Scholar] [CrossRef]
- Stefanovits-Banyai, E.; Nagy, A.; Sardi, E.; Vegvari, A; Pluhar, Z. Morphological, chemical and biochemical characterization of St. John's Wort (Hypericum perforatum L.) hybrids. Acta Biol. Szegedensis 2002, 46, 259–261. [Google Scholar]
- Gaudin, M.; Simonnet, X.; Debruner, N.; Ryser, A. Breeding for a Hypericum perforatum L. variety both productive and Colletotrichum gloeosporoides (Penz.) tolerant. J. Herbs Spi. Med. Plants 2002, 9, 107–120. [Google Scholar] [CrossRef]
- Richter, O. St. John’s Wort (Hypericum perforatum). Richter’s herbal seed company at http://www.richters.com accessed in may 2008.
- Southwell, I.A.; Bourke, C.A. Seasonal variation in hypericin content in Hypericum perforatum L. Phytochemistry 2001, 56, 437–441. [Google Scholar] [CrossRef]
- Verma, V.; Smelcerovic, A.; Zuehlke, S.; Hussain, M.A.; Ahmad, S.M.; Ziebach, T.; Qazi, G.N.; Spiteller, M. Phenolic constituents and genetic profile of Hypericum perforatum L. from India. Biochem. Syst. Ecol. 2008, 36, 210–206. [Google Scholar]
- Arnholdt-Schmitt, B. Characterization of Hypericum perforatum from various accessions by RAPD fingerprinting. J. Herb Spi. Med Plants 2002, 9, 163–169. [Google Scholar] [CrossRef]
- Arnholdt-Schmitt, B. RAPD analysis: a method to investigate aspects of the reproductive biology of Hypericum perforatum L. Theor. Appl. Genet. 2000, 100, 906–911. [Google Scholar] [CrossRef]
- Barcaccia, G.; Arzenton, F.; Sharbe, l.T.F.; Varotto, S.; Parrini, P.; Lucchin, M. Genetic diversity and reproductive biology in ecotypes of the facultative apomict Hypericum perforatum L. Heredity 2006, 96, 322–334. [Google Scholar] [CrossRef]
- Aziz, N.; Sauve, R.J.; Long, D.; Cherry, M. Genetic and phytochemical diversity assessment among eleven Hypericum accessions via AFLP and HPLC analyses. J. Herb Spi. Med Plants 2006, 12, 97–105. [Google Scholar] [CrossRef]
- Haluskova, J.; Cellarova, E. RFLP analysis of Hypericum perforatum L. somaclones and their progenies. Euphytica 1997, 95, 229–235. [Google Scholar] [CrossRef]
- Cellarova, E. Culture and biotechnology of Hypericum. In Hypericum; Ernst, E., Ed.; Taylor&Francis: London, UK, 2003; pp. 65–75. [Google Scholar]
- Crockett, S.L.; Douglas, A.W.; Scheffler, B.E.; Khan, I.A. Genetic profiling of Hypericum (St John’s Wort) species by nuclear ribosomial ITS sequence analysis. Planta Med. 2004, 70, 929–935. [Google Scholar] [CrossRef]
- Smelcerovic, A.; Vijeshwar, V.; Spiteller, M.; Syed Mudasir, A.; Satish, C.P.; Ghulam, N.Q. Phytochemical analysis and genetic characterization of six Hypericum species from Serbia. Phytochemistry 2006, 67, 171–177. [Google Scholar] [CrossRef]
- Urbanova, M.; Kosuth, J.; Cellarova, E. Genetic and biochemical analysis of Hypericum perforatum L. plants regerenated after cryopreservation. Plant Cell Rep. 2006, 25, 140–147. [Google Scholar] [CrossRef]
- Pawelczak, A.; Osinska, E.; Geszprych, A.; Weglarz, Z. Effect of propagation method on plant development and content of biologically active compounds in the herb of St. John's Wort (Hypericum perforatum). Acta Hort. 2006, 725, 461–466. [Google Scholar]
- Franklin, G.; Oliveira, M.; Dias, A.C.P. Production of transgenic Hypericum perforatum plants via particle bombarded-mediated transformation of novel organogenic cell suspension cultures. Plant Sci. 2007, 172, 1193–1203. [Google Scholar] [CrossRef] [Green Version]
- Kirakosan, A.; Sirvent, T.M.; Gibson, D.; Kaufman, P.B. The production of hypericins and hyperforin by in vitro cultures of Hypericum perforatum. Biotech. Appl. Biochem. 2004, 39, 71–81. [Google Scholar] [CrossRef]
- Onelli, E.; Rivetta, A.; Giorgi, A.; Bignami, M.; Cocucci, M.; Patrignani, G. Ultrastructural studies on the developing secretory nodules of Hypericum perforatum. Flora 2002, 197, 92–102. [Google Scholar] [CrossRef]
- Curtis, J.D.; Lersten, N.R. Internal secretory structures of Hypericum (Clusiaceae): H. perforatum L. and H. balnearicum L. New Phytol. 1990, 114, 571–580. [Google Scholar] [CrossRef]
- Kornfeld, A.; Kaufman, P.B.; Lu, C.R.; Gibson, D.M.; Bolling, S.F.; Warber, S.L.; Chang, S.C.; Kirakosyan, A. The production of hypericins in two selected Hypericum perforatum shoot cultures is related to differences in black gland structure. Plant Physiol. Biochem. 2007, 45, 24–32. [Google Scholar] [CrossRef]
- Guillet, G.; Podeszfinsky, C.; Regnault-Roger, C.; Arnason, J.T.; Philogène, B.J.R. Behavioral and biochemical adaptations of generalist and specialist herbivorous insects feeding on Hypericum perforatum (Guttiferae). Environ. Entomol. 2000, 29, 135–139. [Google Scholar] [CrossRef]
- Zobayed, S.M.A.; Afreen, F.; Goto, E.; Kozai, T. Plant-environment interactions: Accumulation of hypericin in dark glands of Hypericum perforatum. Ann. Bot. 2006, 98, 793–804. [Google Scholar] [CrossRef]
- Esau, K. Anatomy of Seed Plants, 2nd ed.; John Wiley and Sons: New York, UK, 1977. [Google Scholar]
- Gadzovska, S.; Maury, S.; Delaunay, A.; Spasenoski, M.; Joseph, C.; Hagège, D. Jasmonic acid elicitation of Hypericum perforatum L. cell suspensions and effects on the production of phenylpropanoids and naphtodianthrones. Plant Cell. Tiss. Organ Cult. 2007, 89, 1–13. [Google Scholar] [CrossRef]
- Soelberg, J.; Jorgensen, L.B.; Jager, A.K. Hyperforin accumulates in the translucent glands of Hypericum perforatum. Ann. Bot. 2007, 99, 1097–1100. [Google Scholar] [CrossRef]
- Cirak, C.; Saglam, B.; Ayan, A.K.; Kevseroglu, K. Morphogenetic and diurnal variation of hypericin in some Hypericum species from Turkey during the course of ontogenesis. Biochem. Syst. Ecol. 2006, 34, 1–13. [Google Scholar] [CrossRef]
- Tekel’ova, D.; Repcak, M.; Zemkova, E.; Toth, J. Quantitative changes in dianthrones, hyperforin and flavonoids content in the flower ontogenesis of Hypericum perforatum. Planta Med. 2000, 66, 778–780. [Google Scholar] [CrossRef]
- Fico, G.; Vitalini, S.; Colombo, N.; Tome, F. Hypericum perforatum L.; H. maculatum Crantz, H. calycinum L. and H. pulchrum L.: phytochemical and morphological studies. Nat. Prod. Comm. 2006, 1, 1129–1132. [Google Scholar]
- Seidler-Lozykowska, K.; Dabrowska, J.; Zygmunt, B. Content of active substances in herb of St. John’s Wort (Hypericum perforatum L.) var. Topaz in different vegetation phases. Herba Pol. 1999, 45, 169–172. [Google Scholar]
- Pasqua, G.; Avato, P.; Monacelli, B.; Santamaria, A.; Argentieri, M. Metabolites in cell suspension cultures, calli, and in vitro regenerated organs of Hypericum perforatum cv. Topas. Plant Sci. 2003, 165, 977–982. [Google Scholar] [CrossRef]
- Repcak, M.; Martonfi, P. The localization of secondary substances in Hypericum perforatum flowers. Biologia (Bratislava), Botany 1997, 52, 91–94. [Google Scholar]
- Maisenbacher, P.; Kovar, K.A. Analysis and stability of hyperici oleum. Planta Med. 1992, 58, 351–354. [Google Scholar] [CrossRef]
- Cirak, C.; Radusiene, J.; Karabuk, B.; Janulis, V.; Ivanauskas, L. Variation of bioactive compounds in Hypericum perforatum growing in Turkey during its phenological cycle. J. Integr. Plant Biol. 2007, 49, 615–620. [Google Scholar] [CrossRef]
- Seidler-Lozykowska, K. Secondary metabolites content in Hypericum sp. in different stages and plant parts. In Hypericum; Ernst, E., Ed.; Taylor&Francis: London, 2003; pp. 100–105. [Google Scholar]
- Cirak, C.; Radusiene, J.; Janulis, V.; Ivanauskas, L. Secondary metabolites in Hypericum perforatum: variation among plant parts and phenological stages. Bot. Helv. 2007, 117, 29–36. [Google Scholar] [CrossRef]
- Kartnig, T.; Heydel, B.; Lasser, B. Johanniskraut aus Schweizer Arzneipflanzenkultur. Agrarforshung 1996, 4, 299–304. [Google Scholar]
- Cicarelli, D.; Andreucci, A.; Pagni, A.M. The “black nodules” of Hypericum perforatum L. subsp. perforatum: morphological, anatomical and histochemical studies during the course of ontogenesis. Israel J. Plant Sci. 2001, 49, 33–40. [Google Scholar]
- Robson, N.K.B. Studies in the genus Hypericum L. (Guttiferae) 2. Characters of the genus. Bull. Brit. Museum 1981, 8, 55–226. [Google Scholar]
- Bomme, U. Produktionstechnologie von Johanniskraur (Hypericum perforatum L.). Z. Arzn. Gew. Pfl. 1997, 2, 127–134. [Google Scholar]
- Hannig, H.J.; Plescher, A.; Vollrath, G. Erfahrungen beim grossflachigen anbau von Johanniskraut – Anfordenungen an die industrielle Verwentung. Herba Germanica 1995, 3, 96–103. [Google Scholar]
- Ayan, A.K.; Cirak, C.; Yanar, O. Variations in total phenolics during ontogenetic, morphogenetic and diurnal cycles in Hypericum species from Turkey. J. Plant Biol. 2006, 49, 432–439. [Google Scholar] [CrossRef]
- Falk, H.; Schwarzberger, W. On the nature of “soluble” hypericin in Hypericum species. Monatsh. Chem. 1992, 123, 731–739. [Google Scholar] [CrossRef]
- Poutaraud, A.; Di Gregorio, F.; Chan Fook Tin, V.; Girardin, P. Effect of light on hypericin contents in fresh flowering top parts and in an extract of St. John’s Wort (Hypericum perforatum). Planta Med. 2001, 67, 254–259. [Google Scholar] [CrossRef]
- Fox, L.R.; Riberiro, S.P.; Brown, V.K.; Masters, G.J.; Clarke, I.P. Direct and indirect effects of climate change on St. John’s Wort, Hypericum perforatum L. (Hypericaceae). Oecologia 1999, 120, 113–122. [Google Scholar] [CrossRef]
- Zobayed, S.M.A.; Afreen, F.; Kozai, T. Phytochemical and physiological changes in the leaves of St. John’s Wort plants under a water stress condition. Environ. Exp. Bot. 2007, 59, 109–116. [Google Scholar] [CrossRef]
- Zobayed, S.M.A.; Afreen, F.; Kozai, T. Temperature stress alter the phytosynthetic efficiency and secondary metabolite concentrations in St. John’s wort. Plant Physiol. Biochem. 2005, 43, 977–984. [Google Scholar] [CrossRef]
- Grey, D.E.; Pallardy, S.G.; Garrett, H.E.; Rottinghaus, G.E. Effect of acute drought stress and time of harvest on phytochemistry and dry weight of St. John’s wort leaves and flowers. Planta Med. 2003, 69, 1024–1030. [Google Scholar] [CrossRef]
- Xenophontos, M.; Stavropoulos, I.; Avramakis, E.; Navakoudis, E.; Dörnemann, D.; Kotzabasis, K. Influence of the habitat altitude on the (proto)hypericin and (proto)pseudohypericin levels of Hypericum plants from Crete. Planta Med. 2008, 74, 1496–1503. [Google Scholar] [CrossRef]
- Briskin, D.P.; Leroy, A.; Gawienowski, M. Influence of nitrogen on the production of hypericins by St. John’s Wort. Plant. Physiol. Biochem. 2000, 38, 413–420. [Google Scholar] [CrossRef]
- Azizi, M.; Omidbaigi, R. Effect of NP supply on herb yield, hypericin content and cadmium accumulation of St.John’s wort (Hypericum perforatum L.). Acta Hort. 2002, 576, 267–271. [Google Scholar]
- Dias, A.C.P.; Majid, A.; Reza, O. Effect of nitrogen and phosporous fertilisation in the cultivation of Hypericum perforatum L. variety Topas. In Short lecture at the International Congress and 49th annual meeting of the Society of Medicinal Plant Research, Erlangen, Germany; 2001. [Google Scholar]
- Briskin, D.P.; Gawienowski, M. Differential effects of light and nitrogen on production on hypericins and leaf glands in Hypericum perforatum. Plant. Physiol. Biochem. 2001, 39, 1075–1081. [Google Scholar] [CrossRef]
- Mosaleeyanon, K.; Zobayed, S.M.A.; Afreen, F.; Kozai, T. Relationships between net photosynthetic rate and secondary metabolite contents in St. John’s wort. Plant Sci. 2005, 169, 523–531. [Google Scholar] [CrossRef]
- Busse, W. The significance of quality for efficacy and safety of herbal medicinal products. Drug Inf. J. 2000, 34, 15–23. [Google Scholar]
- De Pasquale, R.; Rapisarda, A.; Germanò, M.P.; Ragusa, S.; Kirjavainen, S.; Galati, E.M. Effects of cadmium on growth and pharmacologically active constituents of the medicinal plant Coriandrum sativum L. Water, Air, Soil Poll. 1995, 84, 147–157. [Google Scholar] [CrossRef]
- Radanovic, D.; Antic-Mladenovic, S.; Jakovljevic, M. Influence of some soil characteristics on heavy metal content in Hypericum perforatum L. and Achillea millefolium L. Acta Hort. 2002, 576, 295–301. [Google Scholar]
- Ayan, A.K.; Kizilkaya, R.; Cirak, C.; Kevseroglu, K. Heavy metal contents of St. John's wort (Hypericum perforatum L.) growing in northern Turkey. J. Plant Sci. 2006, 1, 182–186. [Google Scholar]
- Chizzola, R.; Lukas, B. Variability of the cadmium content in Hypericum species collected in eastern Austria. Water, Air Soil Poll. 2005, 170, 331–343. [Google Scholar] [CrossRef]
- Chizzola, R.; Michitsch, H.; Franz, C. Monitoring of metallic micronutrients and heavy metals in herbs, Spi. and medicinal plants from Austria. Eur. Food Res. Technol. 2003, 216, 407–411. [Google Scholar]
- Murch, S.J.; Haq, K.; Vasantha Rupasinghe, H.P.; Saxena, P.K. Nickel contamination affects growth and secondary metabolite composition of St. John’s wort (Hypericum perforatum L.). Environ. Exp. Bot. 2003, 49, 251–257. [Google Scholar] [CrossRef]
- Tirillini, B.; Ricci, A.; Pintore, G.; Chessa, M.; Sighinolfi, S. Induction of hypericins in Hypericum perforatum in response to chromium. Fitoterapia 2006, 77, 164–170. [Google Scholar] [CrossRef]
- Baroni Fornasiero, R. Fluorides effects on Hypericum perforatum plants: first field observations. Plant Sci. 2003, 165, 507–513. [Google Scholar] [CrossRef]
- VanEtten, H.D.; Mansfield, J.W.; Bailey, J.A.; Farmer, E.E. Two Classes of Plant Antibiotics: Phytoalexins versus "Phytoanticipins". Plant Cell. 1994, 6, 1191–1192. [Google Scholar]
- Sirvent, T.M.; Krasnoff, S.B.; Gibson, D.M. Induction of hypericins and hyperforins in Hypericum perforatum in response to damage by herbivores. J. Chem. Ecol. 2003, 29, 2667–2681. [Google Scholar] [CrossRef]
- Sirvent, T.; Gibson, D. Induction of hypericins and hyperforin in Hypericum perforatum L. in response to biotic and chemical elicitors. Physiol. Mol. Plant Pathol. 2002, 60, 311–320. [Google Scholar]
- Conceiçao, L.F.R.; Ferreres, F.; Tavares, R.M.; Dias, A.C.P. Induction of phenolic compounds in Hypericum perforatum L. cells by Colletotrichum gloeosporioides elicitation. Phytochemistry 2006, 67, 149–155. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.J.; Dong, J.F.; Zhu, M.Y. Nitric oxide mediates the fungal elicitor induces hypericin production on Hypericum perforatum cell suspension cultures through a jasminic acid dependent signal pathway. Plant Physiol. 2005, 139, 991–998. [Google Scholar] [CrossRef]
- Kegler, H. A Virus causing vein yellowing and necrotic leaf spots of ST. John’s Wort (Hypericum perforatum L.). In Hypericum; Ernst, E., Ed.; Taylor&Francis: London, 2003; pp. 43–47. [Google Scholar]
- Pank, F. The influence of chemical weed control on quality characters of medicinal and aromatic plants. Herba Hung. 1990, 29, 51–58. [Google Scholar]
- Cirak, C.; Aksoy, H.M.; Ayan, A.K.; Saglam, B.; Kevseroglu, K. Enhanced hypericin production in Hypericum perforatum and Hypericum pruinatum in response to inoculation with two fungal pathogens. Plant Protect. Sci. 2005, 41, 109–114. [Google Scholar]
- Heine, H.; Eger, H. Sortenprüfungen mit Johanniskraut-Versuchsergebnisse 1998 und 1999. 10. Bernburger Winterseminar zu Fragen der Arznei-und Gewürzpflanzen-produktion. Kurzfassung der Referate und Poster Teilnehmerliste 2000, 38. [Google Scholar]
- Bruni, R.; Pellati, F.; Bellardi, M.G.; Benvenuti, S.; Paltrinieri, S.; Bertaccini, A.; Bianchi, A. Herbal drug quality and phytochemical composition of Hypericum perforatum L. affected by ash yellows phytoplasma infection. J. Agric. Food Chem. 2004, 52, 964–968. [Google Scholar]
- McLusky, S.; Bennett, M.; Neale, M.; Lewis, M.; Gaskin, P.; Mansfield, J. Cell wall alteration and localized accumulation of feruloyl-3’-metoxytyramine in onion epodermis at sites of attempted penetration by Botrytis allii are associated with actin polarisation, peroxydase activity and suppresssion of flavonoid biosynthesis. Plant J. 1999, 17, 523–534. [Google Scholar] [CrossRef]
- Nishimura, T.; Zobayed, S.M.A.; Kozai, T.; Goto, E. Effect of light quality of blue and red fluorescent lamps on growth of St. John’s wort (Hypericum perforatum L.). Shokubutsu Kankyo Kogaku 2006, 18, 225–229. [Google Scholar]
- Ciccarelli, D.; Andreaucci, C.A.; Pagni, A.M. Translucent glands and secretory canals in Hypericum perforatum L. (Hypericaceae): morpohological, anatomical and histochemical studies during the course of ontogenesis. Ann. Bot. 2001, 88, 637–644. [Google Scholar] [CrossRef]
- Ang, C.W.Y.; Hu, L.; Heinze, T.M.; Cui, Y.; Hreeman, J.P.; Kozak, K.; Luo, W. Instability of St. John’s Wort (Hypericum perforatum L.) and degradation of hyperforin in aqueous solutions and functional beverage. J. Agric. Food Chem. 2004, 52, 6156–6164. [Google Scholar]
- Bilia, A.R.; Bergonzi, M.C.; Morgenni, F.; Mazzi, G.; Vincieri, F.F. Evaluation of chemical stability of St. John’s wort commercial extract and some preparations. Int. J. Pharm. 2001, 213, 199–208. [Google Scholar] [CrossRef]
- Orth, H.C.J.; Schmidt, P.C. Stability and stabilization of hyperforin. Pharm. Ind. 2000, 62, 60–63. [Google Scholar]
- Cantwell, M.I.; Reid, M.S. Postharvest physiology and handling of fresh culinary herbs. J. Herbs, Spi. Med. Plants 1993, 1, 93–127. [Google Scholar] [CrossRef]
- Gaedcke, F. Herstellung, Qualitat, Analytik und Anwendung von Johanniskraut-Extracten. Zeitschr. Arznei Gewurtzpfl. 1997, 2, 63–72. [Google Scholar]
- Kramer, W.; Wiartalla, R. Bestimmung von Naphtodianthronen (Gesamthypericin) in Johanniskraut (Hypericum perforatum L.). Pharm. Zeit. Wissensch. 1992, 137, 202–207. [Google Scholar]
- Bottcher, H.; Gunther, I.; Kabelitz, L. Physiological postharvest responses of common Saint-John’s wort herbs (Hypericum perforatum L.). Postharvest Biol. Tech. 2003, 29, 342–350. [Google Scholar]
- Boeno-Diniz, A.C.; Astarita, L.V.; Romanato-Santarem, E. Alteracao dos metabolitos secundarios em plantas de Hypericum perforatum L. (Hypericaceae) submetidas a secagem e ao congelamento. Acta Bot. Bras. 2007, 21, 443–450. [Google Scholar]
- Bergonzi, M.C.; Bilia, A.R.; Gallori, S.; Guerrini, D.; Vincieri, F.F. Variability in the constituents of Hypericum perforatum L. and some commercial extracts. Drug Dev. Ind. Pharm. 2001, 27, 491–497. [Google Scholar] [CrossRef]
- Tanko, H.; Carrier, D.J.; Duan, L.; Clausen, E. Pre- and post-harvest processing of medicinal plants. Plant Gen. Res. 2005, 3, 304–313. [Google Scholar] [CrossRef]
- Wang, Z.; Lin, S.; Hu, M. Contents of hypericin and pseudohypericin in five commercial products of St John’s Wort (Hypericum perforatum). J. Sci. Food Agric. 2004, 80, 1402–1406. [Google Scholar]
- Kalpana, J.; Preeti, C.; Dnyaneshwar, W.; Bhushan, P. Molecular markers in herbal drug technology. Curr. Sci. 2004, 87, 159–165. [Google Scholar]
- Steck, N.; Messmer, M.; Schaffner, W.; Berger-Bueter, K. Molecular matkers as a tool to verify sexual and apomictic off-spring of intraspecific crosses in Hypericum perforatum. Planta Med. 2001, 67, 384–385. [Google Scholar] [CrossRef]
- Roos, G.; Roseler, C.; Buter, K.B.; Simmen, U. Classification and correlation of St. John’s wort extracts by nuclear magnetic resonance spectroscopy, multivariate data analysis and pharmacological activity. Planta Med. 2004, 70, 771–777. [Google Scholar] [CrossRef]
- Rasmussen, B.; Cloarec, O.; Tang, H.; Staerk, D.; Jaroszewski, J.W. Multivariate analysis of integrated and full-resolution 1H-NMR spectral data from complex pharmaceutical preparations: St. John’s Wort. Planta Med. 2006, 72, 556–563. [Google Scholar] [CrossRef]
- Biffignandi, P.M.; Bilia, A.R. The growing knowledge of St. John’s Wort (Hypericum perforatum L.) drug interactions and their clinical significance. Curr. Ther. Res. 2000, 61, 389–394. [Google Scholar] [CrossRef]
- Caccia, S. Main Active Components of St. John’s Wort (Hypericum perforatum) Extracts: Current Analytical Procedures for Pharmacokinetics and Concentration-Response Studies. Curr. Pharm. Anal. 2006, 2, 59–68. [Google Scholar] [CrossRef]
- Zobayed, S.M.A.; Saxena, P.K. Production of St. John’s wort plants under controlled environment for maximizing biomass and secondary metabolites. In Vitro Cell. Dev. Biol. 2004, 40, 108–114. [Google Scholar] [CrossRef]
- Murch, S.J.; Rupasinghe, H.P.V.; Saxena, P.K. An in vitro and hydroponic growing system for hypericin, pseudohypericin and hyperforin production of St. John’s wort. Planta Med. 2002, 68, 1108–1112. [Google Scholar] [CrossRef]
- Zobayed, S.M.A.; Saxena, P.K. In vitro-grown roots: a superior explant for prolific shoot regeneration of St. John’s wort (Hypericum perforatum L. cv “New Stem”) in a temporary immersion bioreactor. Plant Sci. 2003, 165, 463–470. [Google Scholar] [CrossRef]
- Zobayed, S.M.A.; Murch, S.J.; Rupasinghe, H.P.V.; Saxena, P.K. Elevated carbon supplyaltered hypericin and hyperforin contents of St. John’s wort (Hypericum perforatum) grown in bioreactors. Plant Cell, Tiss. Org. Cult. 2003, 75, 143–149. [Google Scholar] [CrossRef]
- Mueller, S.C.; Majcher-Peszynska, J.; Uehleke, B.; Klammt, S.; Mundkowski, R.G.; Miekisch, W.; Sievers, H.; Bauer, S.; Frank, B.; Kundt, G.; Drewelow, B. The extent of induction of CYP3A by St. John's wort varies among products and is linked to hyperforin dose. Eur. J. Clin. Pharmacol. 2006, 62, 29–36. [Google Scholar]
- Gutmann, H.; Poller, B.; Buter, K.B.; Pfrunder, A.; Schaffner, W.; Drewe, J. Hypericum perforatum: which constituents may induce intestinal MDR1 and CYP3A4 mRNA expression? Planta Med. 2006, 72, 685–690. [Google Scholar] [CrossRef]
- Laakman, G.; Schule, C.; Baghai, T.; Kieser, M. St. John’s Wort in mild to moderate depression: the relevance of hyperforin fot the clinical efficacy. Pharmacopsychiatry 1998, 31, 54–59. [Google Scholar] [CrossRef]
- Bais, H.P.; Vepachedu, R.; Lawrence, C.B.; Stermitz, F.R.; Vivanco, J.M. Molecular and biochemical characterization of an enzyme responsible for the formation of hypericin in St. John’s Wort (Hypericum perforatum L.). J. Biol. Chem. 2003, 278, 32413–32422. [Google Scholar]
- Mahmoudi, M.; Morteza-Semnani, K.; Saeedi, M.; Javanmardi, A. Hypericins content, anti-inflammatory and analgesic activity of iranian Hypericum perforatum. Ind. J. Biol. Biotech. 2006, 3, 91–97. [Google Scholar]
- Gruszczyk, M.; Kieltyka, A. Yields and raw material quality of Hypericum perforatum L. and Solidago virga-aurea L. from one-year and two-year plantations. Herba Polonica 2005, 51, 5–9. [Google Scholar]
- Buter, K.B.; Buter, B. Ontogenetic variation regarding hypericin and hyperforin levels in four accessions of Hypericum perforatum L. J. Herb Spi. Med. Plants 2002, 9, 95–100. [Google Scholar] [CrossRef]
- Berti, M.; Hevia, F.; Wilckens, R.; Joublan, J.P.; Serri, H.; Allende, J. Fertilizacion nitrogenada del cultivo de hierba de San Juan (Hypericum perforatum L.) en Chillan, Provincia de Nuble, Chile. Cien. Investig. Agr. 2000, 27, 107–116. [Google Scholar]
- Smelcerovic, A.; Spiteller, M.; Zuehlke, S. Comparison of methods for the exhaustive extraction of hypericins, flavonoids and hyperforin from Hypericum perforatum L. J. Agric. Food Chem. 2006, 54, 2750–2753. [Google Scholar] [CrossRef]
- Sagratini, G.; Ricciutelli, M.; Vittori, S.; Ozturk, N.; Ozturk, Y.; Maggi, F. Phytochemical and antioxidant analysis of eight Hypericum taxa from Central Italy. Fitoterapia 2008, 72, 210–213. [Google Scholar]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bruni, R.; Sacchetti, G. Factors Affecting Polyphenol Biosynthesis in Wild and Field Grown St. John’s Wort (Hypericum perforatum L. Hypericaceae/Guttiferae). Molecules 2009, 14, 682-725. https://doi.org/10.3390/molecules14020682
Bruni R, Sacchetti G. Factors Affecting Polyphenol Biosynthesis in Wild and Field Grown St. John’s Wort (Hypericum perforatum L. Hypericaceae/Guttiferae). Molecules. 2009; 14(2):682-725. https://doi.org/10.3390/molecules14020682
Chicago/Turabian StyleBruni, Renato, and Gianni Sacchetti. 2009. "Factors Affecting Polyphenol Biosynthesis in Wild and Field Grown St. John’s Wort (Hypericum perforatum L. Hypericaceae/Guttiferae)" Molecules 14, no. 2: 682-725. https://doi.org/10.3390/molecules14020682