Taiwanese Native Plants Inhibit Matrix Metalloproteinase-9 Activity after Ultraviolet B Irradiation

Abstract
:Introduction
Results and Discussion
Total phenolics contents in Taiwanese plants
Effect of Taiwanese Plants on cell viability on WS-1 cell

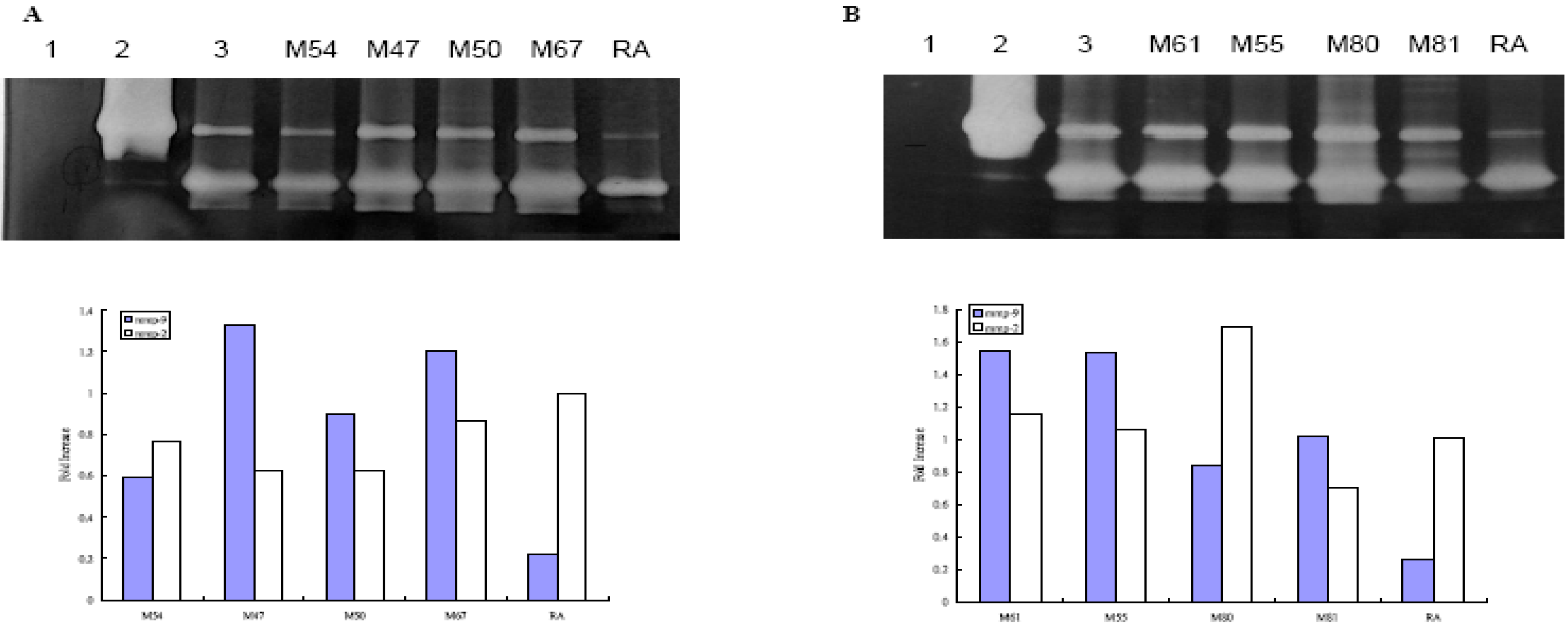{kind=link}
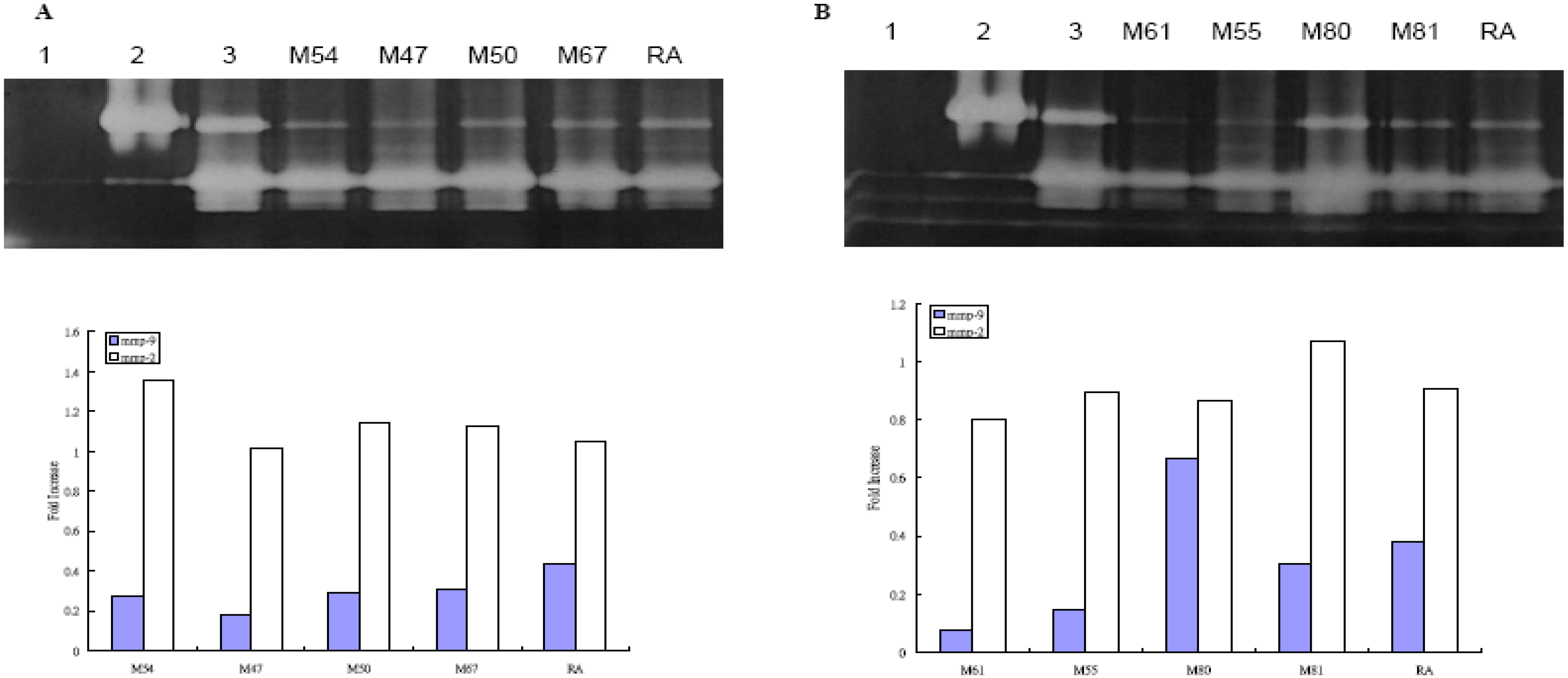{kind=link}
| Botanical name; Family | Voucher specimen | Yield (%) | Total phenolics (mg of GAE /g) |
|---|---|---|---|
| Alnus formosana (Burk.) Makino; Betulaceae | M54 | 10.2 | 15.6 |
| Diospyros discolor Willd.; Ebenaceae | M47 | 13.7 | 36.8 |
| Eriobotrya deflex (Hemsl.) Nakai; Rosaceae | M50 | 8.8 | 29.6 |
| Machilus japonica Sieb.&Zucc. var. kusanoi (Hayata) Liao; Lauraceae | M67 | 12.8 | 25.4 |
| Pyrrosia polydactylis (Hance) Ching; Polypodiaceae | M61 | 16.9 | 21.4 |
| Pyrus taiwanensis Iketani & Ohashi ; Rosaceae | M55 | 8.9 | 30.5 |
| Vitis adstricta Hance; Vitaceae | M80 | 9.2 | 4.8 |
| Vitis thunbergii Sieb. & Zucc; Vitaceae | M81 | 3.7 | 11.2 |
| Specimen | [μg/mL] | without UVB (%) mean ± SEM | [μg/mL] | with UVB (%) mean ± SEM |
|---|---|---|---|---|
| M54 | [100] | 90.45 ± 10.85 | [100] | 92.10 ± 2.20 |
| M47 | [100] | 98.10 ± 1.10 | [200] | 109.7 ± 19.24 |
| M50 | [100] | 103.9 ± 7.12 | [100] | 73.4 ± 23.4 |
| M67 | [100] | 117.3 ± 14.7 | [100] | 140.3 ± 26.3 |
| M61 | [100] | 94.4 ± 10.63 | [200] | 75.9 ± 13.5 |
| M55 | [10] | 94.43 ± 5.42 | [200] | 81.25 ± 8.65 |
| M80 | [100] | 128.1 ± 13.24 | [100] | 136.0 ± 6.20 |
| M81 | [100] | 130.8 ± 14.96 | [100] | 109.1 ± 33.95 |
| RA | [100] | 112.7 ± 6.03 | [100] | 122.7 ± 1.95 |
Effect of Taiwanese Plants on MMP-2 and 9 activities on WS-1 cell
Effect of Taiwanese Plants on MMP-2 and -9 activities after UVB irradiation on WS-1 cell


Conclusions
Experimental
Plant materials
Preparation of plant extracts
Determination of total phenolics
Cell culture
MTT assay for cell viability
MMP-9 zomography
Acknowledgments
References
- Tokura, T.; Nakano, N.; Ito, T.; Matsuda, H.; Nagasako-Akazome, Y.; Kanda, T.; Ikeda, M.; Okumura, K.; Ogawa, H.; Nishiyama, C. Inhibitory effect of polyphenol-enriched apple extracts on mast cell degranulation in vitro targeting the binding between IgE and FcepsilonRI. Biosci. Biotechnol. Biochem. 2005, 69, 1974–1977. [Google Scholar] [CrossRef]
- McDougall, G.J.; Stewart, D. The inhibitory effects of berry polyphenols on digestive enzymes. Biofactors 2005, 23, 189–195. [Google Scholar] [CrossRef]
- Campbell, F.C.; Collett, G.P. Chemopreventive properties of curcumin. Future Oncol. 2005, 1, 405–414. [Google Scholar] [CrossRef]
- Anekonda, T.S. Resveratrol--a boon for treating Alzheimer's disease? Brain Res. Rev. 2006, 52, 316–326. [Google Scholar] [CrossRef]
- An, B.J.; Kwak, J.H.; Park, J.M.; Lee, J.Y.; Park, T.S.; Lee, J.T.; Son, J.H.; Jo, C.; Byun, M.W. Inhibition of enzyme activities and the antiwrinkle effect of polyphenol isolated from the persimmon leaf (Diospyros kaki folium) on human skin. Dermatol. Surg. 2005, 31, 848–854. [Google Scholar]
- Brenneisen, P.; Sies, H.; Scharffetter-Kochanek, K. Ultraviolet-B irradiation and matrix metalloproteinases: from induction via signaling to initial events. Ann. NY Acad. Sci. 2002, 973, 31–43. [Google Scholar] [CrossRef]
- Choi, C.P.; Kim, Y.I.; Lee, J.W.; Lee, M.H. The effect of narrowband ultraviolet B on the expression of matrix metalloproteinase-1, transforming growth factor-beta1 and type I collagen in human skin fibroblasts. Clin. Exp. Dermatol. 2007, 32, 180–185. [Google Scholar] [CrossRef]
- Chung, J.H.; Seo, J.Y.; Choi, H.R.; Lee, M.K.; Youn, C.S.; Rhie, G.; Cho, K.H.; Kim, K.H.; Park, K.C.; Eun, H.C. Modulation of skin collagen metabolism in aged and photoaged human skin in vivo. J. Invest. Dermatol. 2001, 117, 1218–1224. [Google Scholar] [CrossRef]
- Fisher, G.J.; Choi, H.C.; Bata-Csorgo, Z.; Shao, Y.; Datta, S.; Wang, Z.Q.; Kang, S.; Voorhees, J.J. Ultraviolet irradiation increases matrix metalloproteinase-8 protein in human skin in vivo. J. Invest. Dermatol. 2001, 117, 219–226. [Google Scholar] [CrossRef]
- Herrmann, G.; Wlaschek, M.; Lange, T.S.; Prenzel, K.; Goerz, G.; Scharffetter-Kochanek, K. UVA irradiation stimulates the synthesis of various matrix-metalloproteinases (MMPs) in cultured human fibroblasts. Exp. Dermatol. 1993, 2, 92–97. [Google Scholar] [CrossRef]
- Oh, J.H.; Kim, A.; Park, J.M.; Kim, S.H.; Chung, A.S. Ultraviolet B-induced matrix metalloproteinase-1 and -3 secretions are mediated via PTEN/Akt pathway in human dermal fibroblasts. J. Cell. Physiol. 2006, 209, 775–785. [Google Scholar] [CrossRef]
- Bolognia, J.L. Dermatologic and cosmetic concerns of the older woman. Clin. Geriatr. Med. 1993, 9, 209–229. [Google Scholar]
- Correia, H.S.; Batista, M.T.; Dinis, T.C. The activity of an extract and fraction of Agrimonia eupatoria L. against reactive species. Biofactors 2007, 29, 91–104. [Google Scholar] [CrossRef]
- Mandel, S.; Weinreb, O.; Amit, T.; Youdim, M.B. Cell signaling pathways in the neuroprotective actions of the green tea polyphenol (-)-epigallocatechin-3-gallate: implications for neurodegenerative diseases. J. Neurochem. 2004, 88, 1555–1569. [Google Scholar] [CrossRef]
- Nakazato, T.; Sagawa, M.; Yamato, K.; Xian, M.; Yamamoto, T.; Suematsu, M.; Ikeda, Y.; Kizaki, M. Myeloperoxidase is a key regulator of oxidative stress mediated apoptosis in myeloid leukemic cells. Clin. Cancer Res. 2007, 13, 5436–5445. [Google Scholar]
- Devipriya, N.; Sudheer, A.R.; Vishwanathan, P.; Menon, V.P. Modulatory potential of ellagic acid, a natural plant polyphenol on altered lipid profile and lipid peroxidation status during alcohol-induced toxicity: a pathohistological study. J. Biochem. Mol. Toxicol. 2008, 22, 101–112. [Google Scholar]
- Joubert, E.; Richards, E.S.; Merwe, J.D.; De Beer, D.; Manley, M.; Gelderblom, W.C. Effect of species variation and processing on phenolic composition and in vitro antioxidant activity of aqueous extracts of Cyclopia spp. (Honeybush Tea). J. Agric. Food Chem. 2008, 56, 954–963. [Google Scholar] [CrossRef]
- Yadav, A.S.; Bhatnagar, D. Modulatory effect of spice extracts on iron-induced lipid peroxidation in rat liver. Biofactors 2007, 29, 147–157. [Google Scholar] [CrossRef]
- Zaid, M.A.; Afaq, F.; Syed, D.N.; Dreher, M.; Mukhtar, H. Inhibition of UVB-mediated oxidative stress and markers of photoaging in immortalized HaCaT keratinocytes by pomegranate polyphenol extract POMx. Photochem. Photobiol. 2007, 83, 882–888. [Google Scholar] [CrossRef]
- Fisher, G.J.; Talwar, H.S.; Lin, J.; Voorhees, J. J. Molecular mechanisms of photoaging in human skin in vivo and their prevention by all-trans retinoic acid. Photochem. Photobiol. 1999, 69, 154–157. [Google Scholar] [CrossRef]
- Fisher, G.J.; Voorhees, J.J. Molecular mechanisms of photoaging and its prevention by retinoic acid: ultraviolet irradiation induces MAP kinase signal transduction cascades that induce Ap-1-regulated matrix metalloproteinases that degrade human skin in vivo. J. Investig. Dermatol. Symp. Proc. 1998, 3, 61–68. [Google Scholar]
- Fisher, G.J.; Talwar, H.S.; Lin, J.; Lin, P.; McPhillips, F.; Wang, Z.; Li, X.; Wan, Y.; Kang, S.; Voorhees, J.J. Retinoic acid inhibits induction of c-Jun protein by ultraviolet radiation that occurs subsequent to activation of mitogen-activated protein kinase pathways in human skin in vivo. J. Clin. Invest. 1998, 101, 1432–1440. [Google Scholar] [CrossRef]
- Hou, W.C.; Lin, R.D.; Cheng, K.T.; Hung, Y.T.; Cho, C.H.; Chen, C.H.; Hwang, S.Y.; Lee, M.H. Free radical-scavenging activity of Taiwanese native plants. Phytomedicine 2003, 10, 170–175. [Google Scholar]
- Rittie, L.; Fisher, G.J. UV-light-induced signal cascades and skin aging. Ageing Res. Rev. 2002, 1, 705–720. [Google Scholar] [CrossRef]
- Hase, T.; Shinta, K.; Murase, T.; Tokimitsu, I.; Hattori, M.; Takimoto, R.; Tsuboi, R.; Ogawa, H. Histological increase in inflammatory infiltrate in sun-exposed skin of female subjects: the possible involvement of matrix metalloproteinase-1 produced by inflammatory infiltrate on collagen degradation. Br. J. Dermatol. 2000, 142, 267–273. [Google Scholar] [CrossRef]
- Brenneisen, P.; Wenk, J.; Wlaschek, M.; Krieg, T.; Scharffetter-Kochanek, K. Activation of p70 ribosomal protein S6 kinase is an essential step in the DNA damage-dependent signaling pathway responsible for the ultraviolet B-mediated increase in interstitial collagenase (MMP-1) and stromelysin-1 (MMP-3) protein levels in human dermal fibroblasts. J. Biol. Chem. 2000, 275, 4336–4344. [Google Scholar]
- Watanabe, H.; Shimizu, T.; Nishihira, J.; Abe, R.; Nakayama, T.; Taniguchi, M.; Sabe, H.; Ishibashi, T.; Shimizu, H. Ultraviolet A-induced production of matrix metalloproteinase-1 is mediated by macrophage migration inhibitory factor (MIF) in human dermal fibroblasts. J. Biol. Chem. 2004, 279, 1676–1683. [Google Scholar] [CrossRef]
- Thibodeau, A. Metalloproteinase inhibitors. Cosmet. & Toilet. 2000, 115, 75–82. [Google Scholar]
- Kujala, T.S.; Loponen, J.M.; Klika, K.D.; Pihlaja, K. Phenolics and betacyanins in red beetroot (Beta vulgaris) root: distribution and effect of cold storage on the content of total phenolics and three individual compounds. J. Agric. Food Chem. 2000, 48, 5338–5342. [Google Scholar]
- Windsor, J.; Poulsen, A.H.; Yamada, S.; Lyons, G.; Birkedal-Hansen, B.; Stetler-Stevenson, W.; Birkedal-Hansen, H. Matrix metalloproteinases. In Current Protocols in Cell Biology; Bonifacino, J.S., Dasso, M., Harford, J.B., Lippincott-Schwartz, J., Yamada, K.M., Eds.; John Wiley & Sons, Inc.: New York, USA, 2002; Chapter 10; pp. 10.8.1–10.8.23. [Google Scholar]
- Samples Availability: Available from the authors.
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lee, Y.-L.; Lee, M.-H.; Chang, H.-J.; Huang, P.-Y.; Huang, I.-J.; Cheng, K.-T.; Leu, S.-J. Taiwanese Native Plants Inhibit Matrix Metalloproteinase-9 Activity after Ultraviolet B Irradiation. Molecules 2009, 14, 1062-1071. https://doi.org/10.3390/molecules14031062
Lee Y-L, Lee M-H, Chang H-J, Huang P-Y, Huang I-J, Cheng K-T, Leu S-J. Taiwanese Native Plants Inhibit Matrix Metalloproteinase-9 Activity after Ultraviolet B Irradiation. Molecules. 2009; 14(3):1062-1071. https://doi.org/10.3390/molecules14031062
Chicago/Turabian StyleLee, Yueh-Lun, Mei-Hsien Lee, Hsiu-Ju Chang, Po-Yuan Huang, I-Jen Huang, Kur-Ta Cheng, and Sy-Jye Leu. 2009. "Taiwanese Native Plants Inhibit Matrix Metalloproteinase-9 Activity after Ultraviolet B Irradiation" Molecules 14, no. 3: 1062-1071. https://doi.org/10.3390/molecules14031062