Submerged Fermentation of the Edible Mushroom Pleurotus ostreatus in a Batch Stirred Tank Bioreactor as a Promising Alternative for the Effective Production of Bioactive Metabolites

 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
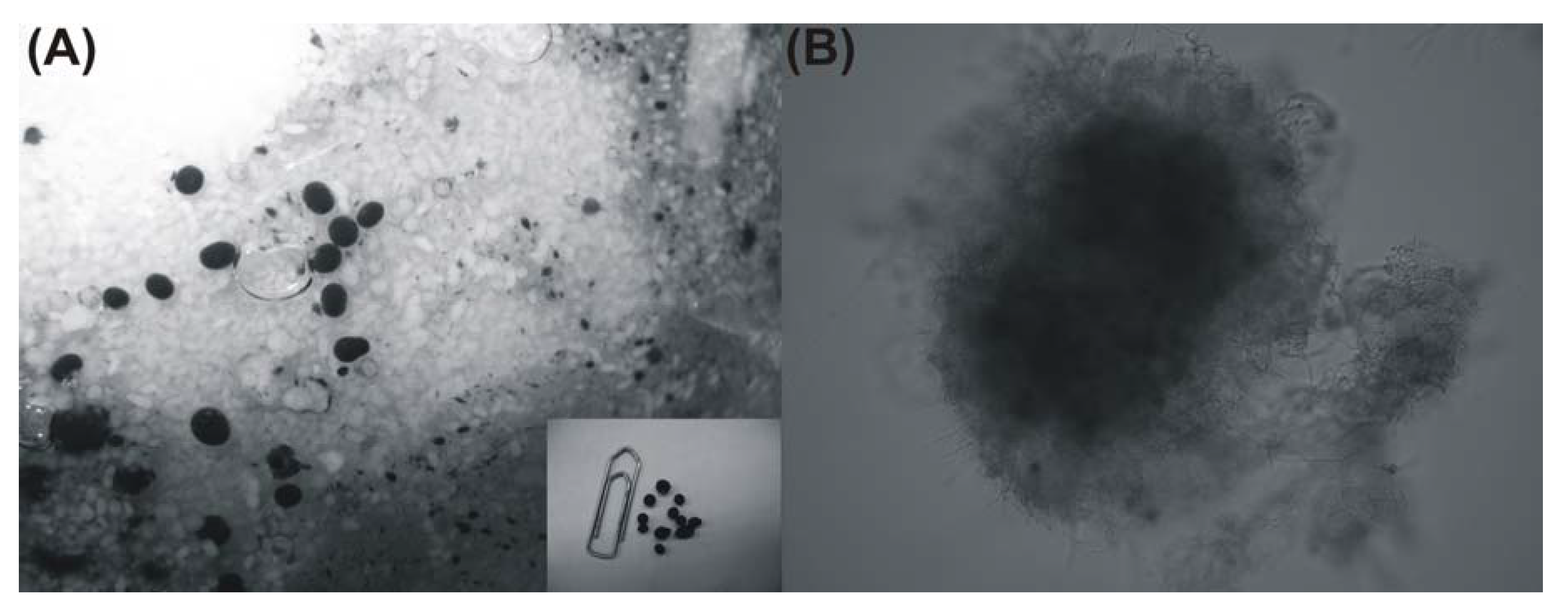
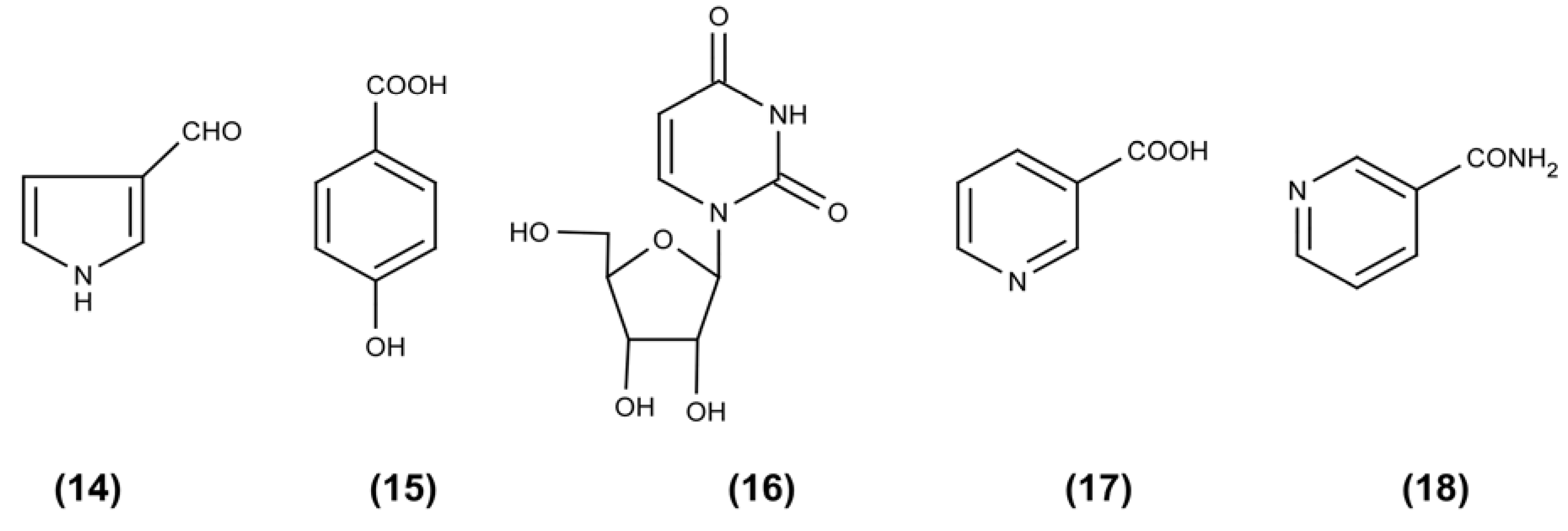
2. Results and Discussion



3. Experimental
3.1. General
3.2. Biological Material
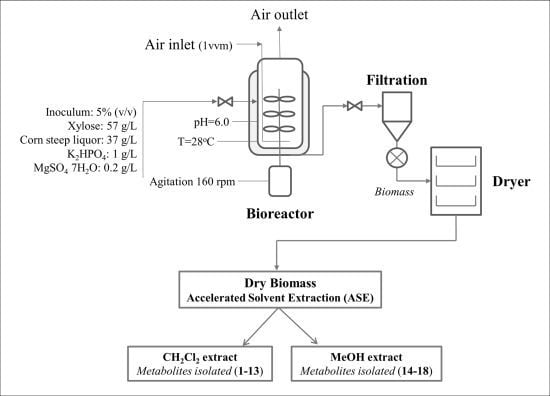
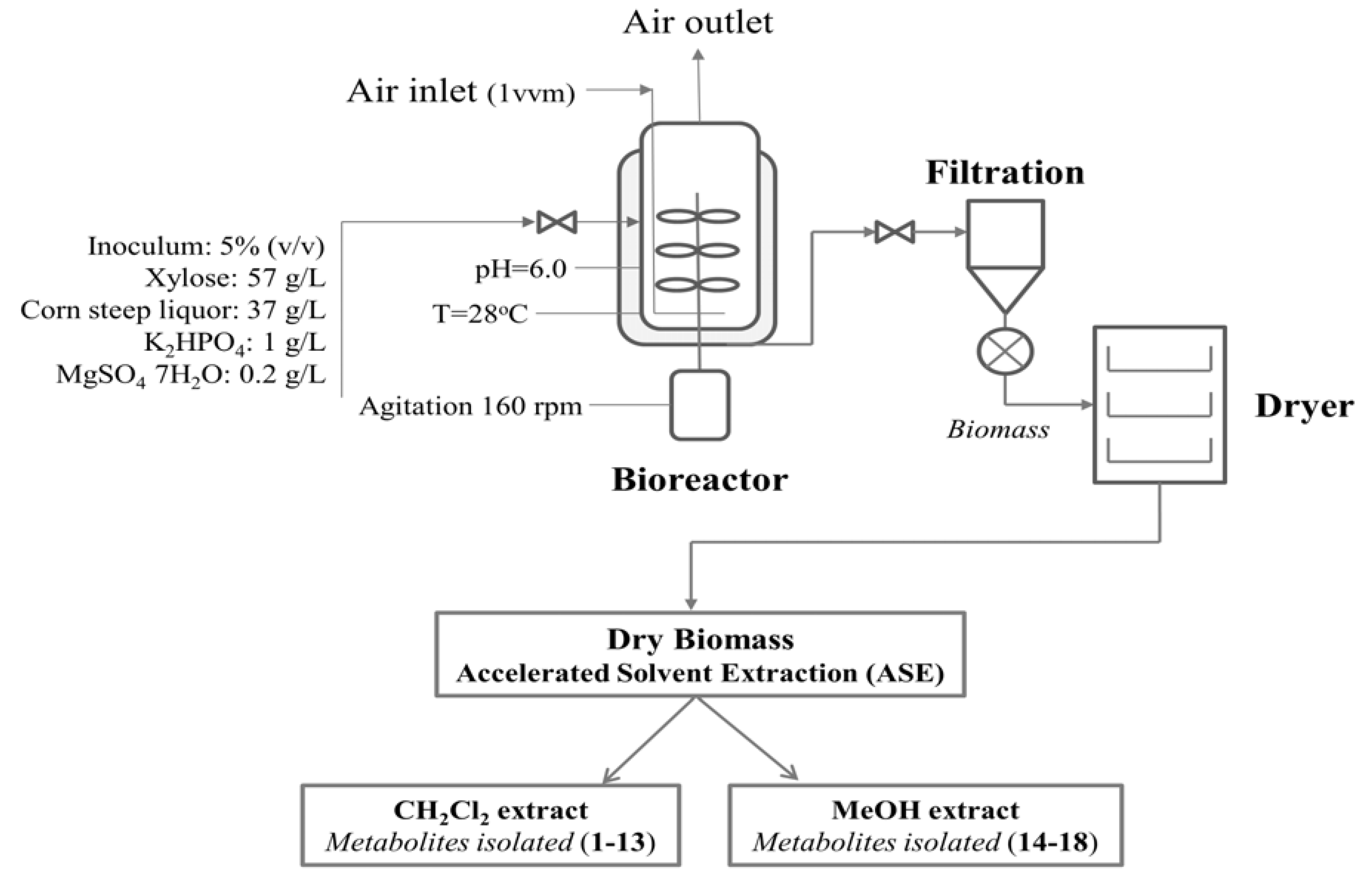
3.3. Media and Fermentation Conditions in 20-L Bioreactor

3.4. Extraction
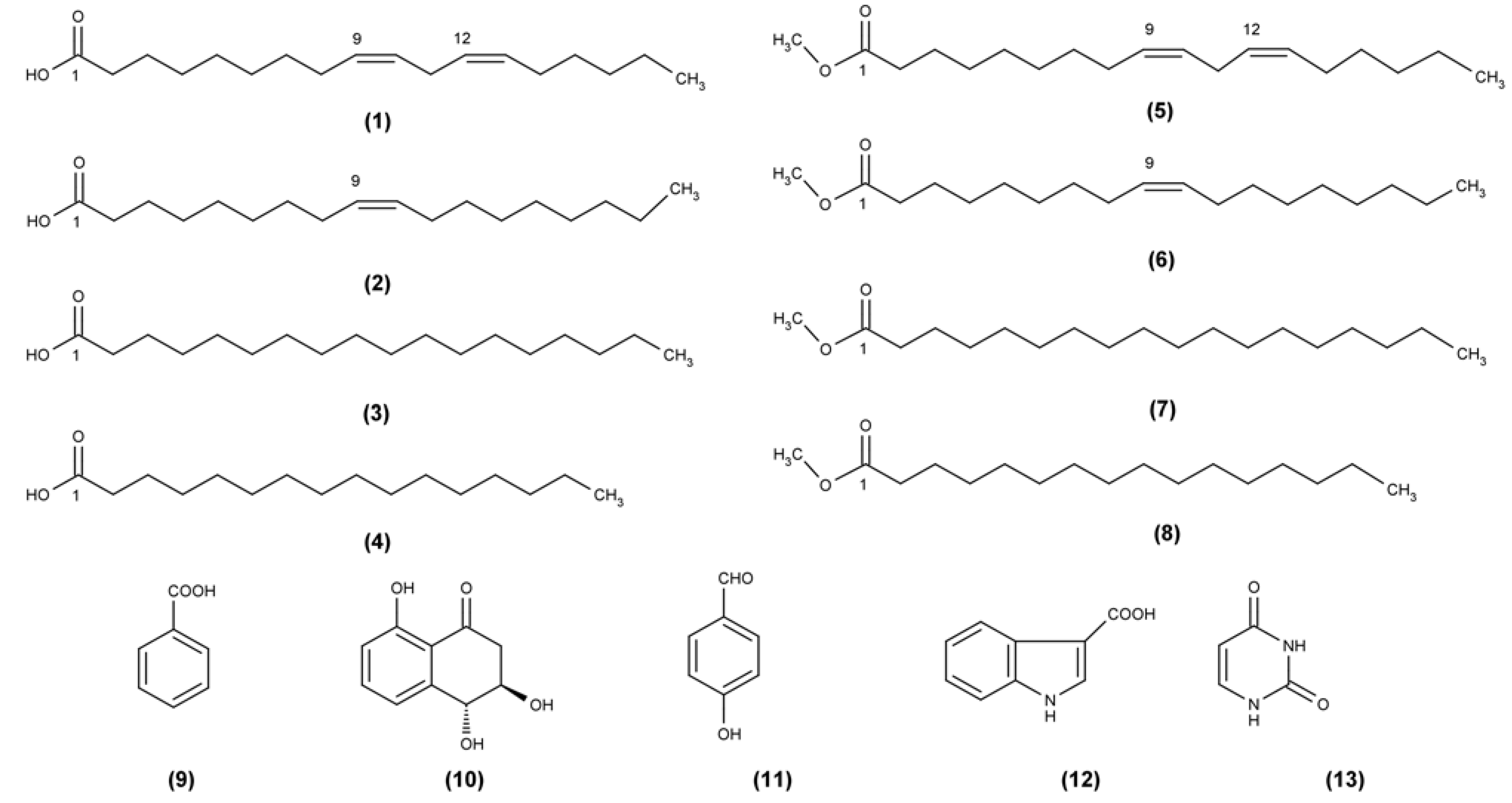
3.5. Isolation and Identification of Compounds
4. Conclusions
Acknowledgments
- Samples Availability: Samples of compounds 9-18 are available from the authors.
References and Notes
- Ying, J.Z.; Mao, X.L.; Ma, Q.M; Zong, Y.C.; Wen, H.A. Icons of Medicinal Fungi from China; Icons of Medicinal Fungi from China; Science Press: Beijing, China, 1987. [Google Scholar]
- Sullivan, R.; Smith, J.E.; Rowan, N.J. Medicinal Mushrooms and Cancer Therapy: Translating a traditional practice into Western medicine. Perspect. Biol. Med. 2006, 49, 159–170. [Google Scholar] [CrossRef]
- Barros, L.; Baptista, P.; Correia, D.M.; Morais, J.S.; Ferreira, I.C.F.R. Effects of conservation treatment and cooking on the chemical composition and antioxidant activity of Portuguese wild edible mushrooms. J. Agric. Food. Chem. 2007, 55, 4781–4788. [Google Scholar]
- Gregori, A.; Švagelj, M.; Pohleven, J. Cultivation Techniques and Medicinal Properties of Pleurotus spp. Food Technol. Biotech. 2007, 45, 238–249. [Google Scholar]
- Tang, Y.Z.; Zhu, L.W.; Li, H.M.; Li, D.S. Submerged culture of mushrooms in bioreactors-challenges, current-state-of-the-art, and future. Food Technol. Biotech. 2007, 45, 221–229. [Google Scholar]
- Papaspyridi, L.-M.; Katapodis, P.; Gonou-Zagou, Z.; Kapsanaki-Gotsi, E.; Christakopoulos, P. Optimization of biomass production with enhanced glucan and dietary fibres content by Pleurotus ostreatus ATHUM 4438 under submerged culture. Biochem. Eng. J. 2010, 50, 131–138. [Google Scholar] [CrossRef]
- Sato, K.; Aoki, M.; Takagi, J.; Zimmermann, K.; Noyori, R. A practical Method for Alcohol oxidation with aqueous hydrogen peroxide under organic solvent-and halide-free conditions. Bull. Chem. Soc. Jpn. 1999, 72, 2287–2306. [Google Scholar] [CrossRef]
- Couché, E.; Fkyerat, A.; Tabacchi, R. Stereoselective Synthesis of cis- and trans-3,4-Dihydro-3,4,8-trihydroxynaphthalen-1(2H)-one. Helv. Chim. Acta 2009, 92, 903–917. [Google Scholar] [CrossRef]
- Tan, J.; Bednarek, P.; Liu, J.; Schneider, B.; Svatoš, A.; Hahlbrock, K. Universally occurring phenylpropanoid and species-specific indolic metabolites in infected and uninfected Aravidopsis thaliana roots and leaves. Phytochemistry 2004, 65, 691–699. [Google Scholar] [CrossRef]
- Bano, S.; Ahmad, V.U.; Perveen, S.; Bano, N.; Shameel, S.M. Marine Natural Products. II. Chemical Constituents of Red Alga Botryocladia leptopoda. Planta Med. 1987, 53, 117–118. [Google Scholar] [CrossRef]
- Bednareka, E.; Dobrowolskia, J.C.; Dobrosz-Teperekc, K.; Kozerskia, L.; Lewandowskia, W.; Mazureka, A.P. Theoretical and experimental 1H, 13C, 15N, and 17O NMR chemical shifts for 5-halogenouracils. J. Mol. Struct. 2000, 554, 233–243. [Google Scholar] [CrossRef]
- Chattipakorn, N.; Settakorn, J.; Petsophonsakul, P.; Suwannahoi, P.; Mahakraukrouch, P.; Srichairatanakool, S.; Chattipakorn, S.C. Cardiac mortality is associated with low levels of omega-3 and omega-6 fatty acids in the heart of cadavers with a history of coronary heart disease. Nutr. Res. 2009, 29, 696–704. [Google Scholar] [CrossRef]
- Garrett, R.H.; Grisham, C.M. Principals of Biochemistry with a Human Focus; Brooks/Cole: Pacific Grove, CA, USA, 2001. [Google Scholar]
- Loppez-Huertas, E. Health effects of oleic acid and long chain omega-3 fatty acids (EPA and DHA) enriched milks. A review of intervention studies. Pharm. Res. 2010, 61, 200–207. [Google Scholar] [CrossRef]
- Matsuura, R.; Ukeda, H.; Sawamura, M. Tyrosinase inhibitory activity of citrus essential oils. J. Agric. Food Chem. 2006, 54, 2309–2313. [Google Scholar] [CrossRef]
- Mu, Y.M.; Yanase, T.; Nishi, Y.; Tanaka, A.; Saito, M.; Jin, C.H.; Mukasa, C.; Okabe, T.; Nomura, M.; Goto, K.; Nawata, H. Saturated FFAs, Palmitic Acid and Stearic Acid, Induce Apoptosis in Human Granulosa Cells. Endocrinology 2001, 142, 3590–3597. [Google Scholar] [CrossRef]
- Shushni, M.A.M.; Mentel, R.; Lindequist, U.; Jansen, R. Balticols A–F, new naphthalenone derivatives with antiviral activity from an Ascomycetous Fungus. Chem. Biodivers. 2009, 6, 127–137. [Google Scholar] [CrossRef]
- Wu, P.L.; Lin, F.W.; Wu, T.S.; Kuoh, C.S.; Lee, K.H.; Lee, S.J. Cytotoxic and anti-HIV principles from the rhizomes of Begonia nantoensis. Chem. Pharm. Bull. 2004, 52, 345–349. [Google Scholar] [CrossRef]
- Pedneault, K.; Angers, P.; Avis, T.; Gosselin, A.; Tweddel, R.J. Fatty acid profiles of polar and non-polar lipids of Pleurotus ostreatus and P. cornucopiae var. ‘ citrino-pileatus’ grown at different temperatures. Mycol. Res. 2007, 111, 1228–1234. [Google Scholar]
- Beltran-Garcia, M.J.; Estarron-Espinosa, M.; Ogura, T. Volatile compounds secreted by the Oyster Mushroom (Pleurotus ostreatus) and their antibacterial activities. J. Agric. Food Chem. 1997, 45, 4049–4052. [Google Scholar] [CrossRef]
- Basarab, G.S.; Steffens, J.J.; Wawrzak, Z.; Schwartz, R.S.; Lundqvist, T.; Jordan, D.B. Catalytic mechanism of scytalone dehydratase: Site-directed mutagenesis, kinetic isotope effects, and alternate substrates. Biochemistry 1999, 38, 6012–6024. [Google Scholar] [CrossRef]
- Henson, J.M.; Butler, M.J.; Day, A.W. The dark side of the mycelium: melanins of phytopathogenic fungi. Annu. Rev. Phytopathol. 1999, 37, 447–471. [Google Scholar] [CrossRef]
- Petersen, R.H.; Nicholl, D.B.G.; Hughes, K.W. Mating systems of some putative polypore-agaric relatives. Plant Syst. Evol. 1997, 201, 135–158. [Google Scholar]
- Selvakumar, P.; Rajasekar, Ζ.S.; Periasamy, Ζ.K.; Raaman, Ζ.N. Isolation and characterization of melanin pigment from Pleurotus cystidiosus (telomorph of Antromycopsis macrocarpa). World J. Microbiol. Biotechnol. 2008, 24, 2125–2131. [Google Scholar]
- Gutiérrez, A.; Caramelo, L.; Prieto, A.; Martìnez, M.J.; Martìnez, A.T. Anisaldehyde production and aryl-alcohol oxidase and dehydrogenase activities in ligninolytic fungi of the genus Pleurotus. Appl. Environ. Microbiol. 1994, 60, 1783–1788. [Google Scholar]
- Jumpathong, J.; Peberby, J.; Fujii, I.; Lumyong, S. Chemical investigation of novel ascomycetes using PCR based screening approaches. World J. Microbiol. Biotechnol. 2011. [Google Scholar] [CrossRef]
- Mantle, P.G. The role of tryptophan as a biosynthetic precursor of indole-diterpenoid fungal metabolites: Continuing a debate. Phytochemistry 2009, 70, 7–10. [Google Scholar] [CrossRef]
- Purkarthhofer, T.; Gruber, K.; Fechter, M.; Grienge, H. Asymetric biocatalytic hydrocyanation of pyrrole carboxaldehydes. Tetrahedron 2005, 61, 7661–7668. [Google Scholar] [CrossRef]
- Wang, Z.J.; Li, G.M.; Nie, B.M.; Lu, Y.; Yin, M. Neuroprotective effect of the stearic acid against oxidative stress via phosphatidylinositol 3-kinase pathway. Chem.-Biol. Interact. 2006, 160, 80–87. [Google Scholar]
- Kupche, É.L.; Mikstais, U.Ya. Effect of the solvent on the 13C NMR spectra of pyrimidine nucleosides. Chem. Heterocycl. Compd. 1980, 16, 1184–1186. [Google Scholar] [CrossRef]
- Connolly, G.P.; Duley, J.A. Uridine and its nucleotides: Biological actions, therapeutic potentials. Trends Pharm. Sci. 1999, 20, 218–225. [Google Scholar] [CrossRef]
- Lourenço, M.C.S.; de Souza, M.V.N.; Pinheiro, A.C.; de Ferreira, M.L.; Gonçalves, R.S.B.; Noqueira, T.C.M.; Peralta, M.A. Evaluation of anti-tuberular activity of nicotinic acid and isoniazid analogues. ARCIVOC 2007, xv, 181–191. [Google Scholar]
- Mahmood, Z.A.; Ahmed, S.W.; Azhar, I.; Sualeh, M.; Baig, M.T.; Zaha, S. Bioactive alkaloids produced by fungi. Updates on alkaloids from the species of the genera Boletus, Fusarium and Psilocybe. Pak. J. Pharm. Sci. 2010, 23, 349–357. [Google Scholar]
- Soni, M.G.; Carabin, I.G.; Burdock, G.A. Safety Assessment of esters of p-hydroxybenzoic (parabens). Food Chem. Toxicol. 2005, 43, 985–1015. [Google Scholar] [CrossRef]
- Li, L.Y.; Diang, Y.; Groth, I.; Menzel, K.D.; Peschel, G.; Voigt, K.; Deng, Z.W.; Sattler, I.; Lin, W.H. Pyrrole and indole alkaloids from an endophytic Fusarium incarnatum (HKI00504) isolated from the mangrove plant Aegiceras corniculatum. J. Asian Nat. Prod. Res. 2008, 10, 775–780. [Google Scholar]
- Tsuchinari, M.; Shimanuki, K.; Hiramatsu, F.; Murayama, T.; Koseki, T.; Shiono, Y. Fusapyridons A and B, Novel Pyridone Alkaloids from an Endophytic Fungus, Fusarium sp. YG-45. Z. Naturforsch 2007, 62, 1203–1207. [Google Scholar]
- Lapadatescu, C.; Giniès, C.; le Quèrè, J.-L.; Bonarme, P. Novel Sceme For Biosynthesis of Aryl Metabolites from L-Phenylanine in the Fungus Bjerkandera adusta. Appl. Environ. Microbiol. 2000, 66, 1517–1522. [Google Scholar] [CrossRef]
- Cağlarirmak, N. The nutrients of exotic mushrooms (Lentinula edodes and Pleurotus species) and an estimated approach to the volatile compounds. Food Chem. 2007, 105, 1188–1194. [Google Scholar] [CrossRef]
- Kanagasabapathy, G.; Malek, S.N.A.; Kuppusamy, U.R.; Vikineswary, S. Chemical Composition and Antioxidant Properties of Extracts of Fresh Fruitbodies of Pleurotus sajor-caju (Fr.) Singer. J. Agric. Food Chem. 2011, 59, 2618–2626. [Google Scholar]
- Kundakovic, T.; Fokialakis, N.; Kovacevic, N.; Chinou, I. Essential oil composition of Achillea lingulata and A. umbellate. umbellate. Flavour Fragr.J. 2007, 22, 184–187. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Papaspyridi, L.-M.; Aligiannis, N.; Topakas, E.; Christakopoulos, P.; Skaltsounis, A.-L.; Fokialakis, N. Submerged Fermentation of the Edible Mushroom Pleurotus ostreatus in a Batch Stirred Tank Bioreactor as a Promising Alternative for the Effective Production of Bioactive Metabolites. Molecules 2012, 17, 2714-2724. https://doi.org/10.3390/molecules17032714
Papaspyridi L-M, Aligiannis N, Topakas E, Christakopoulos P, Skaltsounis A-L, Fokialakis N. Submerged Fermentation of the Edible Mushroom Pleurotus ostreatus in a Batch Stirred Tank Bioreactor as a Promising Alternative for the Effective Production of Bioactive Metabolites. Molecules. 2012; 17(3):2714-2724. https://doi.org/10.3390/molecules17032714
Chicago/Turabian StylePapaspyridi, Lefki-Maria, Nektarios Aligiannis, Evangelos Topakas, Paul Christakopoulos, Alexandros-Leandros Skaltsounis, and Nikolas Fokialakis. 2012. "Submerged Fermentation of the Edible Mushroom Pleurotus ostreatus in a Batch Stirred Tank Bioreactor as a Promising Alternative for the Effective Production of Bioactive Metabolites" Molecules 17, no. 3: 2714-2724. https://doi.org/10.3390/molecules17032714