Enhancement of Palmarumycin C12 and C13 Production in Liquid Culture of the Endophytic Fungus Berkleasmium sp. Dzf12 by Oligosaccharides from Its Host Plant Dioscorea zingiberensis

 and
and
Abstract
:Abbreviations
| OW | crude oligosaccharide prepared by acid hydrolysis of the water-extracted polysaccharide |
| OS | crude oligosaccharide prepared by acid hydrolysis of the sodium hydroxide-extracted polysaccharide |
| OA | crude oligosaccharide prepared by acid hydrolysis of the acid-extracted polysaccharide |
| WEP | water-extracted polysaccharide |
| SEP | sodium hydroxide-extracted polysaccharide |
| AEP | acid-extracted polysaccharide |
| TFA | trifluoroacetic acid |
1. Introduction
2. Results and Discussion
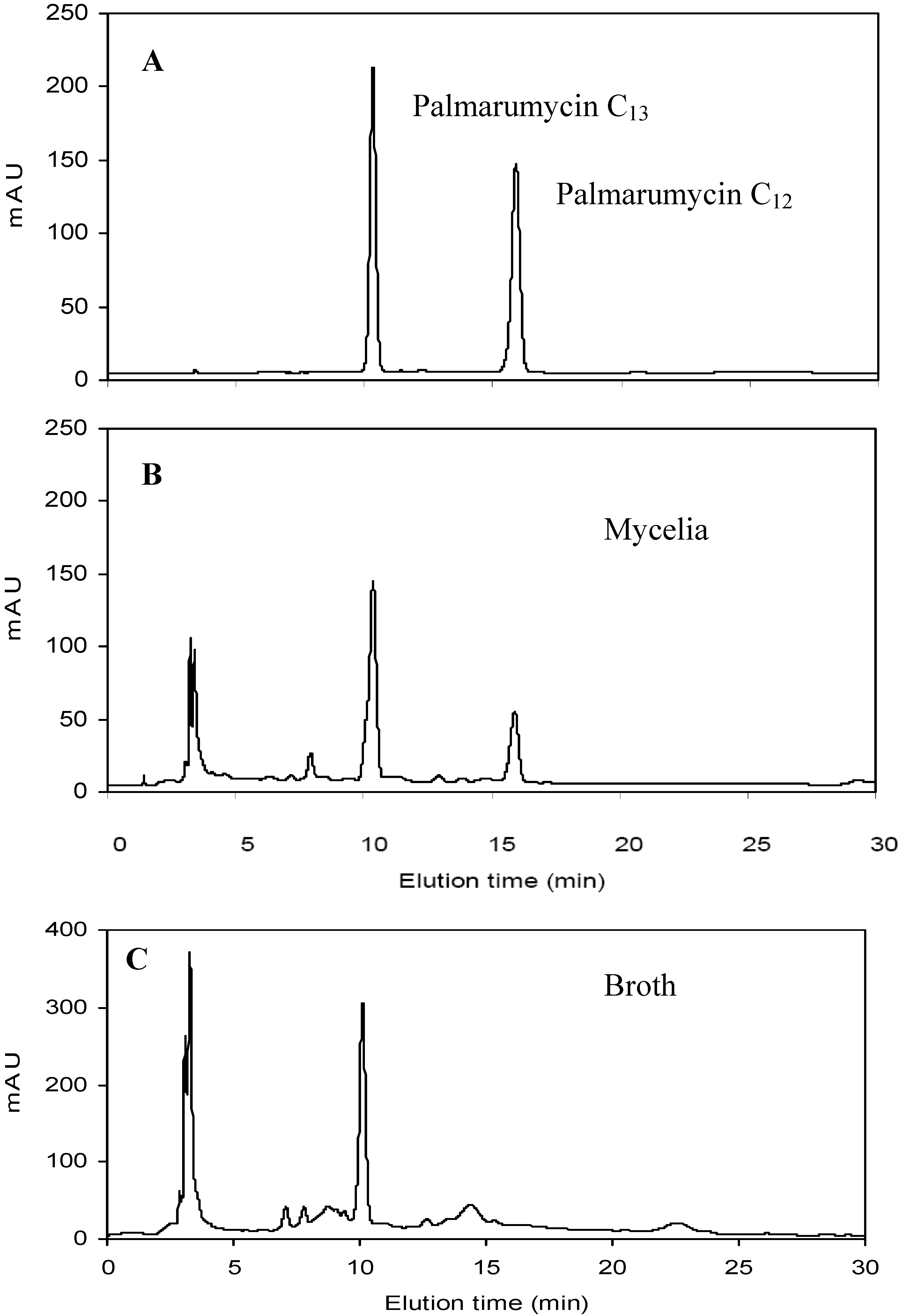
2.1. Effects of Oligosaccharides OW, OS and OA on Mycelia Growth and Palmarumycin Production

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oligo. Conc. (mg/L) | Mycelia biomass (g dw/L) | C12 content in mycelia (mg/g dw) | C13 content in mycelia (mg/g dw) | C13 yield in broth (mg/L) | C12 plus C13 yield (mg/L) | |
|---|---|---|---|---|---|---|
| CK | 0 | 6.84 ± 0.26 e | 1.31 ± 0.08 h | 3.24 ± 0.18 f | 108.28 ± 6.00 e | 139.45 ± 5.20 g |
| OW | 200 | 11.48 ± 0.26 b | 5.40 ± 0.24 c | 8.21 ± 0.21 b | 229.06 ± 6.62 b | 385.28 ± 8.32 b |
| 400 | 13.90 ± 0.86 a | 6.20 ± 0.31 b | 9.92 ± 0.18 a | 245.40 ± 4.69 a | 469.33 ± 9.43 a | |
| 600 | 14.06 ± 0.25 a | 4.95 ± 0.18 d | 6.50 ± 0.32 c | 193.78 ± 10.41c | 369.53 ± 16.80 b | |
| OS | 100 | 8.69 ± 0.26 d | 2.42 ± 0.14 g | 4.21 ± 0.14 e | 169.23 ± 7.01 d | 226.90 ± 8.35 f |
| 200 | 9.24 ± 0.29 cd | 5.09 ± 0.19 cd | 6.88 ± 0.06 c | 232.16 ± 9.47 ab | 340.96 ± 14.79 c | |
| 400 | 9.55 ± 0.47 c | 8.06 ± 0.30 a | 5.22 ± 0.06 d | 179.05 ± 7.21 d | 305.83 ± 8.69 d | |
| OA | 200 | 8.80 ± 0.24 d | 2.63 ± 0.10 fg | 4.33 ± 0.16 e | 176.69 ± 8.76 d | 237.89 ± 9.33 f |
| 400 | 9.32 ± 0.37 cd | 3.44 ± 0.17 e | 6.29 ± 0.35 c | 241.72 ± 7.16 ab | 332.28 ± 5.92 c | |
| 600 | 9.82 ± 0.30 c | 2.93 ± 0.15 f | 5.15 ± 0.12 d | 205.61 ± 9.00 c | 283.56 ± 10.75 e | |

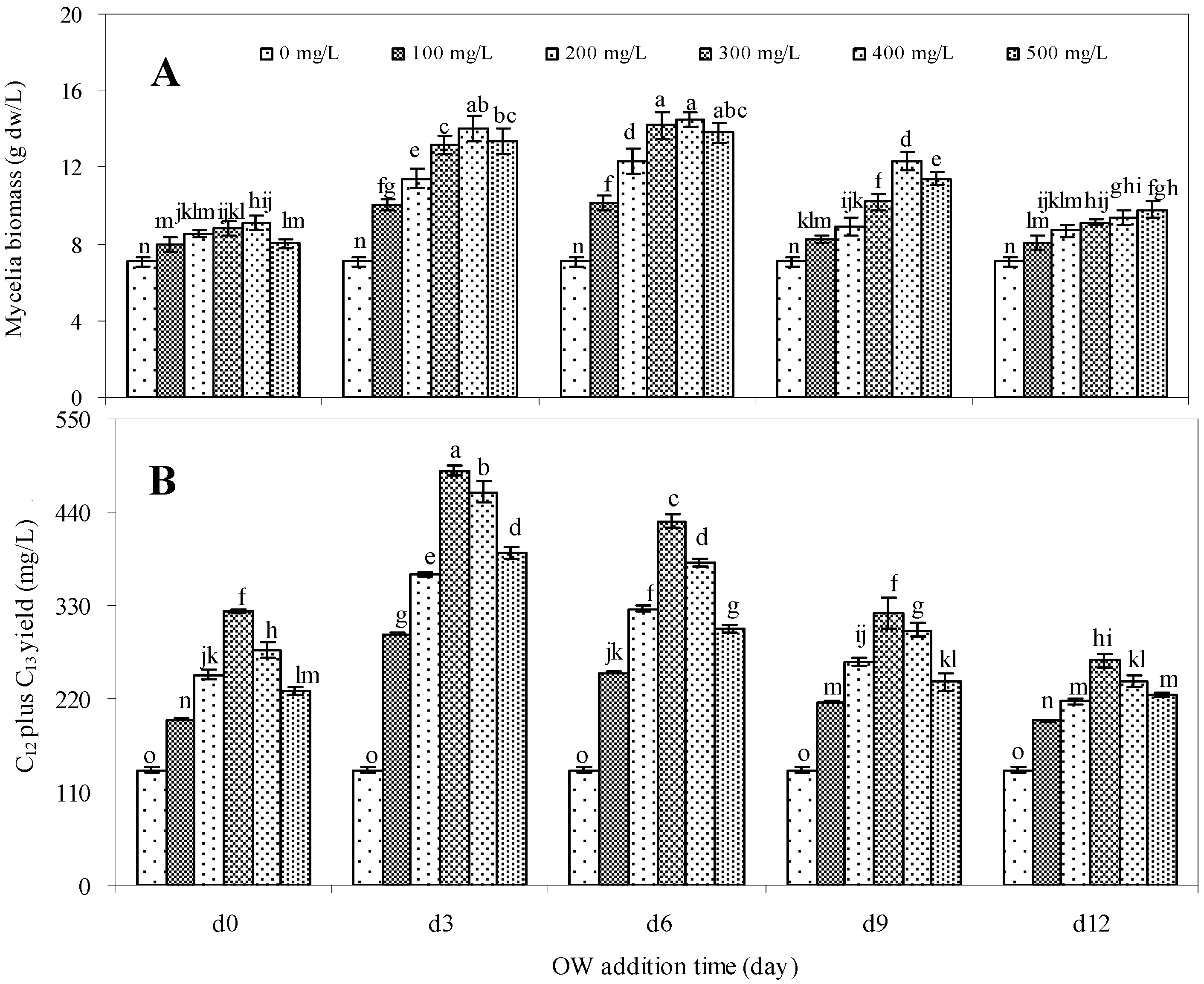
2.2. Effects of Oligosaccharide OW Addition Time on Mycelia Growth and Palmarumycin Production

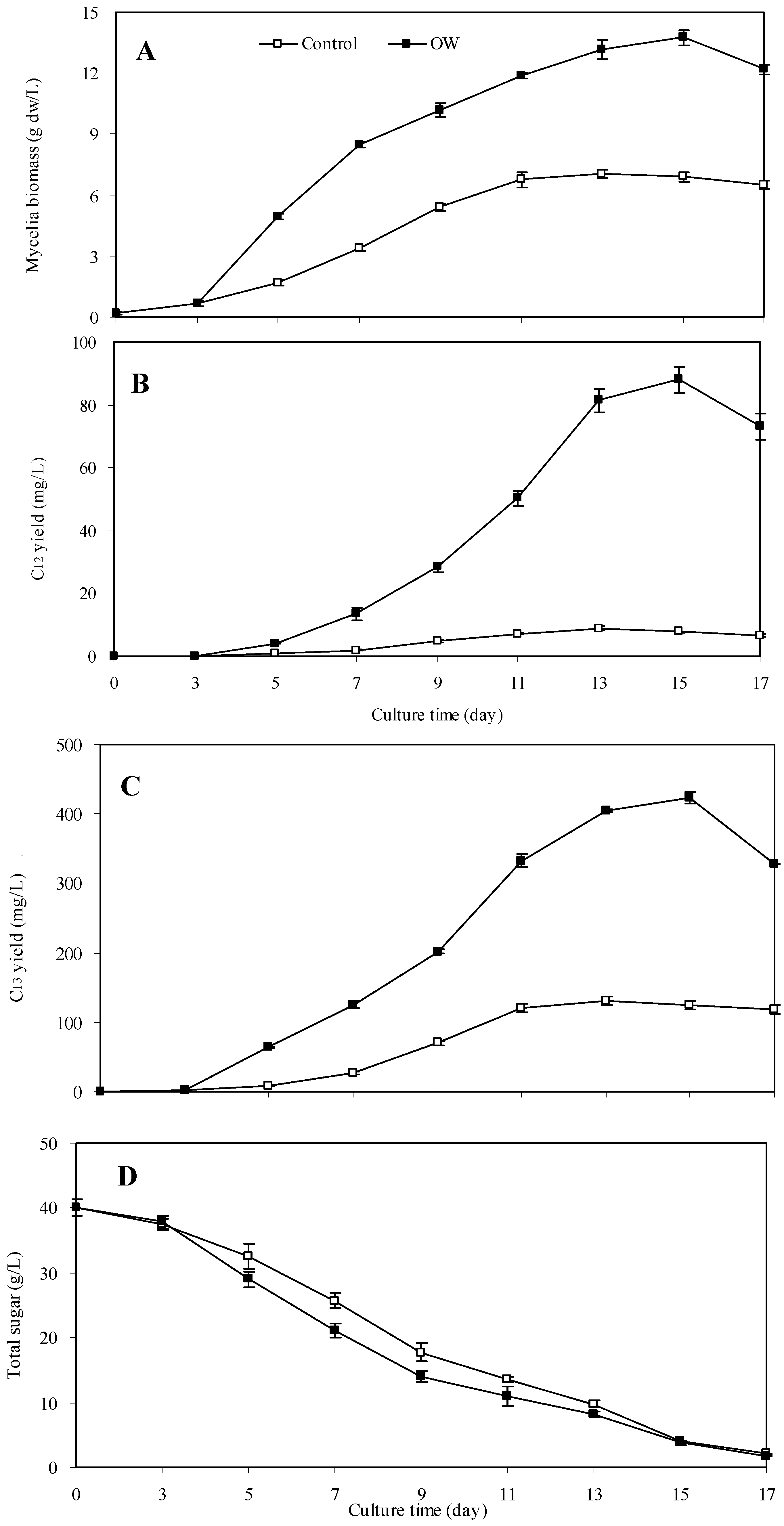
2.3. Kinetics of Mycelia Growth and Palmarumycin Accumulation after Treatment with Oligosaccharide OW

3. Experimental
3.1. Endophytic Fungus and Culture Conditions
3.2. Preparation of Oligosaccharides
3.3. Application of Oligosaccharides
3.4. Determination of Mycelia Biomass
3.5. Extraction and Quantification of Palmarumycins C12 and C13
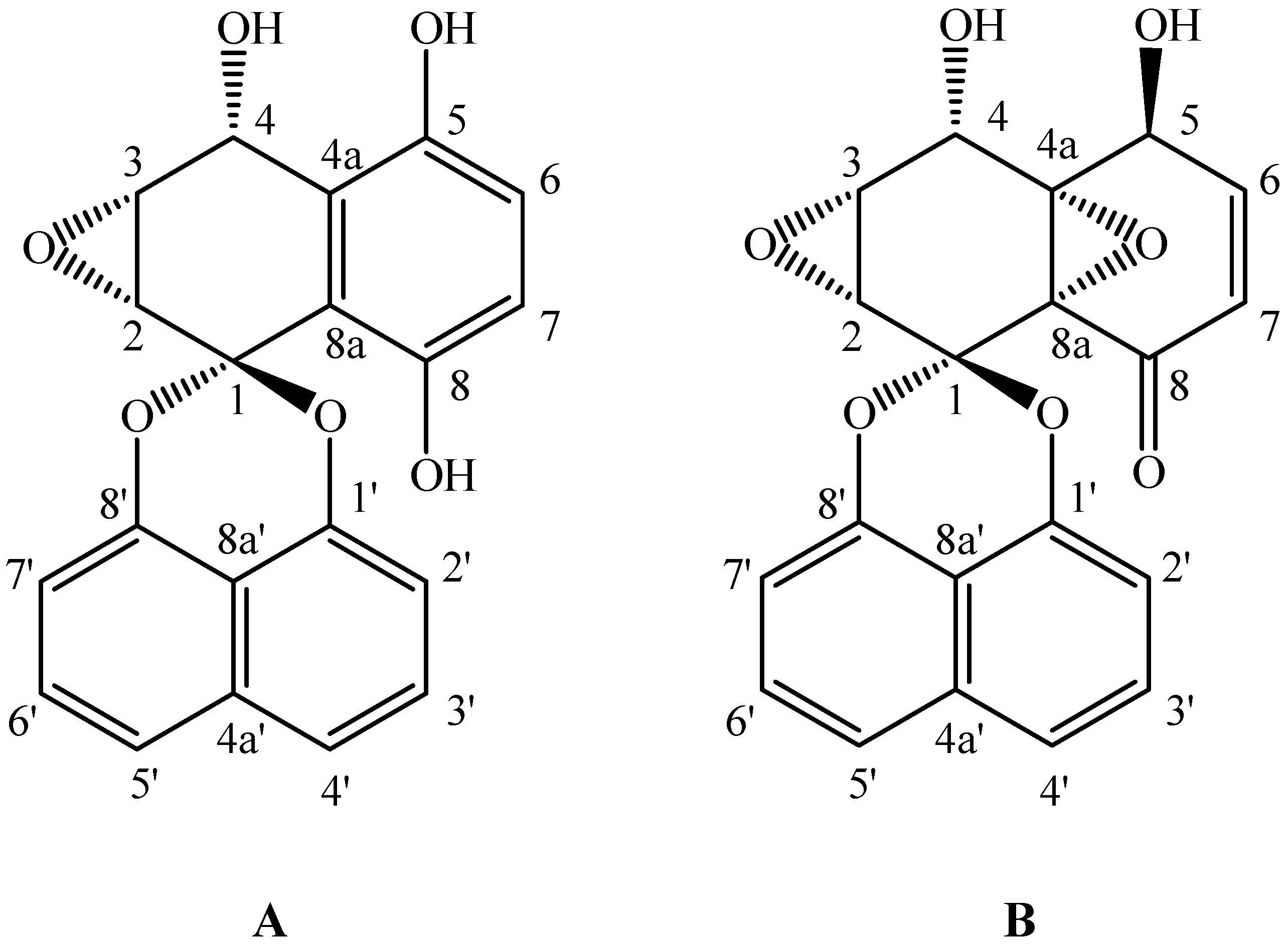
3.6. Physicochemical and Spectroscopic Data of Palmarumycins C12 and C13

3.7. Measurement of Total Sugar Content
3.8. Statistical Analysis
4. Conclusions
Acknowledgements
References and Notes
- Aly, A.H.; Debbab, A.; Kjer, J.; Proksch, P. Fungal endophytes from higher plants: A prolific source of phytochemicals and other bioactive natural products. Fungal Divers. 2010, 41, 1–16. [Google Scholar]
- Schulz, B.; Boyle, C.; Draeger, S.; Rommert, A.K.; Krohn, K. Endophytic fungi: A source of novel biologically active secondary metabolites. Mycol. Res. 2002, 106, 996–1004. [Google Scholar]
- Strobel, G.; Daisy, B.; Castillo, U.; Harper, J. Natural products from endophytic microorganisms. J. Nat. Prod. 2004, 67, 257–268. [Google Scholar]
- Kharwar, R.N.; Mishra, A.; Gond, S.K.; Stierle, A.; Stierle, D. Anticancer compounds derived from fungal endophytes: Their importance and future challenges. Nat. Prod. Rep. 2011, 28, 1208–1228. [Google Scholar]
- Zhao, J.; Shan, T.; Mou, Y.; Zhou, L. Plant-derived bioactive compounds produced by endophytic fungi. Mini-Rev. Med. Chem. 2011, 11, 159–168. [Google Scholar] [CrossRef]
- Cai, X.; Shan, T.; Li, P.; Huang, Y.; Xu, L.; Zhou, L.; Wang, M.; Jiang, W. Spirobisnaphthalenes from the endophytic fungus Dzf12 of Dioscorea zingiberensis and their antimicrobial activities. Nat. Prod. Commun. 2009, 4, 1469–1472. [Google Scholar]
- Zhou, L.; Zhao, J.; Shan, T.; Cai, X.; Peng, Y. Spirobisnaphthalenes from fungi and their biological activities. Mini-Rev. Med. Chem. 2010, 10, 977–989. [Google Scholar] [CrossRef]
- Cai, Y.S.; Guo, Y.W.; Krohn, K. Structure, bioactivities, biosynthetic relationships and chemical synthesis of the spirodioxynaphthalene. Nat. Prod. Rep. 2010, 27, 1840–1870. [Google Scholar]
- Krohn, K.; Michel, A.; Floerke, U.; Aust, H.-J.; Draeger, S.; Schulz, B. Palmarumycins C1-C16 from Coniothyrium sp.: Isolation, structure elucidation, and biological activity. Liebigs Ann. Chem. 1994, 11, 1099–1108. [Google Scholar]
- Bode, H.B.; Walker, M.; Zeeck, A. Secondary metabolites by chemical screening. 42 Cladospirones B to I from Sphaeropsidales sp. F-24'707 by variation of culture conditions. Eur. J. Org. Chem. 2000, 18, 3185–3193. [Google Scholar]
- Schlingmann, G.; West, R.R.; Milne, L.; Pearce, C.J.; Carter, G.T. Diepoxins, novel fungal metabolites with antibiotic activity. Tetrahedron Lett. 1993, 34, 7225–7228. [Google Scholar]
- Chu, M.; Truumees, I.; Patel, M.G.; Gullo, V.P.; Blood, C.; King, I.; Pai, J.K.; Puar, M.S. A novel class of anti-tumor metabolites from the fungus Nattrassia mangiferae. Tetrahedron Lett. 1994, 35, 1343–1346. [Google Scholar]
- Thiergardt, R.; Hug, P.; Rihs, G.; Peter, H.H. Cladospirone bisepoxide-a novel fungal metabolite structure determination. Tetrahedron Lett. 1994, 35, 1043–1046. [Google Scholar]
- Thiergardt, R.; Rihs, G.; Hug, P.; Peter, H.H. Cladospirone bisepoxide: Definite structure assignment including absolute configuration and selective chemical transformations. Tetrahedron 1995, 51, 733–742. [Google Scholar]
- Petersen, F.; Moerker, T.; Vanzanella, F.; Peter, H.H. Production of cladosporine bisepoxide, a new fungal metabolite. J. Antibiot. 1994, 47, 1098–1103. [Google Scholar]
- Zhou, L.G.; Wu, J.Y. Development and application of medicinal plant tissue cultures for production of drugs and herbal medicinals in China. Nat. Prod. Rep. 2006, 23, 789–810. [Google Scholar]
- Schneider, P.; Misiek, M.; Hoffmeister, D. In vivo and in vitro production options for fungal secondary metabolites. Mol. Pharmaceut. 2008, 5, 234–242. [Google Scholar] [CrossRef]
- Zhao, J.; Zhou, L.; Wu, J. Effects of biotic and abiotic elicitors on cell growth and tanshinone accumulation in Salvia miltiorrhiza cell cultures. Appl. Microbiol. Biotechnol. 2010, 87, 137–144. [Google Scholar] [CrossRef]
- Zhang, R.; Li, P.; Xu, L.; Chen, Y.; Sui, P.; Zhou, L.; Li, J. Enhancement of diosgenin production in Dioscorea zingiberensis cell culture by oligosaccharide elicitor from its endophytic fungus Fusarium oxysporum Dzf17. Nat. Prod. Commun. 2009, 4, 1459–1462. [Google Scholar]
- Li, P.; Mou, Y.; Shan, T.; Xu, J.; Li, Y.; Lu, S.; Zhou, L. Effects of polysaccharide elicitors from endophytic Fusarium oxysporium Dzf17 on growth and diosgenin production in cell suspension culture of Dioscorea zingiberensis. Molecules 2011, 16, 9003–9016. [Google Scholar]
- Zhao, J.; Zheng, B.; Li, Y.; Shan, T.; Mou, Y.; Lu, S.; Li, P.; Zhou, L. Enhancement of diepoxin ζ production by yeast extract and its fractions in liquid culture of Berkleasmium-like endophytic fungus Dzf12 from Dioscorea zingiberensis. Molecules 2011, 16, 847–856. [Google Scholar] [CrossRef]
- Ariyo, B.; Tamerler, C.; Bucke, C.; Keshavarz, T. Enhanced penicillin production by oligosaccharides from batch cultures of Penicillium chrysogenum in stirred-tank reactors. FEMS Microbiol. Lett. 1998, 166, 165–170. [Google Scholar] [CrossRef]
- Tamerler, C.; Ariyo, B.; Bucke, C.; Keshavarz, T. Effect of mannan and alginate oligosaccharides on production in bioreactors of penicillin G and its biosynthetic intermediates. Ann. Microbiol. 2001, 51, 53–60. [Google Scholar]
- Ariyo, B.; Bucke, C.; Keshaverz, T. Alginate oligosaccharides as enhancers of penicillin production in cultures of Penicillium chrysogenum. Biotechnol. Bioeng. 1997, 53, 17–20. [Google Scholar] [CrossRef]
- Radman, R.; Bucke, C.; Keshavarz, T. Elicitor effects on Penicillium chrysogenum morphology in submerged cultures. Biotechnol. Appl. Biochem. 2004, 40, 229–233. [Google Scholar] [CrossRef]
- Liu, G.; Casqueiro, J.; Gutierrez, S.; Kosalkova, K.; Castillo, N.I.; Martin, J.F. Elicitation of penicillin biosynthesis by alginate in Penicillium chrysogenum, exerted on pcbAB, pcbC, and penDE genes at the transcriptional level. J. Microbiol. Biotechnol. 2001, 11, 812–818. [Google Scholar]
- Li, Y.; Li, P.; Mou, Y.; Zhao, J.; Shan, T.; Ding, C.; Zhou, L. Enhancement of diepoxin ζ production in liquid culture of endophytic fungus Berkleasmium sp. Dzf12 by polysaccharides from its host plant Dioscorea zingiberensis. World J. Microbiol. Biotechnol. 2012, 28. [Google Scholar] [CrossRef]
- Zhu, L.W.; Tang, Y.J. Significance of protein elicitor isolated from Tuber melanosporum on the production of ganoderic acid and Ganoderma polysaccharides during the fermentation of Ganoderma lucidum. Bioproc. Biosyst. Eng. 2010, 33, 999–1005. [Google Scholar] [CrossRef]
- Li, P.; Mao, Z.; Lou, J.; Li, Y.; Mou, Y.; Lu, S.; Peng, Y. Enhancement of diosgenin in Dioscorea zingiberensis cell cultures by oiligosaccharides from its endophytic fungus Fusarium oxysporum Dzf17. Molecules 2011, 16, 10631–10644. [Google Scholar] [CrossRef]
- Murphy, T.; Parra, R.; Radman, R.; Roy, I.; Harrop, A.; Dixon, K.; Keshavarz, T. Novel application of oligosaccharides as elicitors for the enhancement of bacitracin A production in cultures of Bacillus licheniformis. Enzyme Microb. Technol. 2007, 40, 1518–1523. [Google Scholar] [CrossRef]
- Zhang, R.; Li, P.; Zhao, J.; Yin, C.; Zhou, L. Endophytic fungi from Dioscorea zingiberensis and their effects on the growth and diosgenin production of the host plant cultures. Nat. Prod. Res. Dev. 2010, 22, 11–15. [Google Scholar]
- Zhao, J.; Li, Y.; Shan, T.; Mou, Y.; Zhou, L. Enhancement of diepoxin ζ production with in situ resin adsorption in mycelial liquid culture of the endophytic fungus Berkleasmium sp. Dzf12 from Dioscorea zingiberensis. World J. Microbiol. Biotechnol. 2011, 27, 2753–2758. [Google Scholar] [CrossRef]
- Barrett, A.G.M.; Blaney, F.; Campbell, A.D.; Hamprecht, D.; Meyer, T.; White, A.J.P.; Witty, D.; Williams, D.J. Unified route to the palmarumycin and preussomerin natural products. Enantioselective synthesis of (-)-preussomerin G. J. Org. Chem. 2002, 67, 2735–2750. [Google Scholar]
- Schlingmann, G.; Matile, S.; Berova, N.; Nakanishi, K.; Carter, G.T. Absolute stereochemistry of the diepoxins. Tetrahedron 1996, 52, 435–446. [Google Scholar]
- Zhu, L.W.; Zhong, J.J.; Tang, Y.J. Significance of fungal elicitors on the production of ganoderic acid and Ganoderma polysaccharides by the submerged culture of medicinal mushroom Ganoderma lucidum. Process Biochem. 2008, 43, 1359–1370. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the oligosaccharides are available from the authors.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, Y.; Shan, T.; Mou, Y.; Li, P.; Zhao, J.; Zhao, W.; Peng, Y.; Zhou, L.; Ding, C. Enhancement of Palmarumycin C12 and C13 Production in Liquid Culture of the Endophytic Fungus Berkleasmium sp. Dzf12 by Oligosaccharides from Its Host Plant Dioscorea zingiberensis. Molecules 2012, 17, 3761-3773. https://doi.org/10.3390/molecules17043761
Li Y, Shan T, Mou Y, Li P, Zhao J, Zhao W, Peng Y, Zhou L, Ding C. Enhancement of Palmarumycin C12 and C13 Production in Liquid Culture of the Endophytic Fungus Berkleasmium sp. Dzf12 by Oligosaccharides from Its Host Plant Dioscorea zingiberensis. Molecules. 2012; 17(4):3761-3773. https://doi.org/10.3390/molecules17043761
Chicago/Turabian StyleLi, Yan, Tijiang Shan, Yan Mou, Peiqin Li, Jianglin Zhao, Wensheng Zhao, Youliang Peng, Ligang Zhou, and Chunbang Ding. 2012. "Enhancement of Palmarumycin C12 and C13 Production in Liquid Culture of the Endophytic Fungus Berkleasmium sp. Dzf12 by Oligosaccharides from Its Host Plant Dioscorea zingiberensis" Molecules 17, no. 4: 3761-3773. https://doi.org/10.3390/molecules17043761