In Vitro Proliferation and Production of Cytokine and IgG by Human PBMCs Stimulated with Polysaccharide Extract from Plants Endemic to Gabon

Abstract
:1. Introduction
2. Results and Discussion
2.1. Plant Samples

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Angiosperm Phylogeny | Plants, Family and Names Used | Part and Names Used | Traditional Action | References | |
|---|---|---|---|---|---|
| Magnoliales | Uvaria klainei Pierre ex Engl. & Diels; Annonaceae | Leaves (UkL) | - | - | |
| Stems (UkS) | - | - | |||
| Ericales | Petersianthus macrocarpus (P. Beauv.) Liben; Lecythidaceae | Barks (PmB) | Antiseptic, abortive, hypotensive | [11,12] | |
| Sapindales | Trichoscypha addonii De wild; Anacardiaceae | Barks (TaB) | Dysentery, amenorrhea | [11] | |
| Fabales | Aphanocalyx microphyllus (Harms) Weiring | Fabaceae –Caesalpinioideae | Barks (AmB) | - | - |
| Librevillea klaineana (Pierre ex Harms) Hoyle | Barks (LkB) | Venereal disease | |||
| Neochevalierodendron stephanii A. Chev.) J. Léonard | Leaves (NsL) | Antibiotic | [11] | ||
| Scorodophloeus zenkeri Harms | Barks (SzB) | Spice; treats high blood pressure, respiratory disease | |||
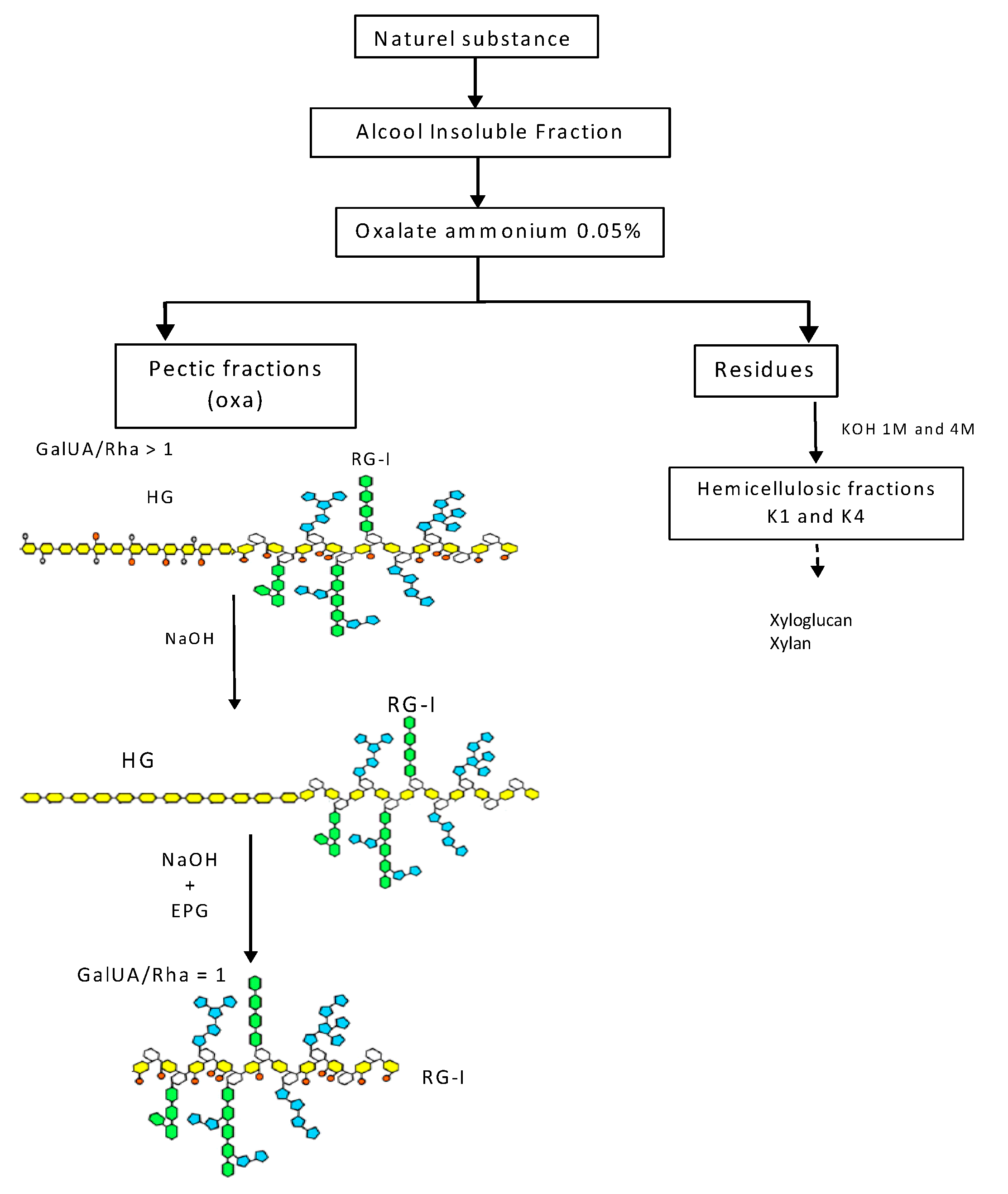
2.2. Isolation and Chemistry of Polysaccharides

2.3. Study Population
| Averages | Men | Women |
|---|---|---|
| Age (years) | 29.33 ± 3.1 | 27.67 ± 5.6 |
| Weights (Kg) | 74.17 ± 6.6 | 67.17 ± 10.2 |
| Heights (cm) | 171.5 ± 5.7 | 159.3 ± 3.4 |
| Temperature (°C) | 37.05 ± 0.3 | 37.1 ± 0.2 |
| Blood-pressure (mmHg) | 106.67/75 | 108.33/73.33 |
| Red corpuscle (mm3) | 4,323,333 ± 136,666.7 | 5,098,333 ± 301,666.7 |
| White corpuscle (mm3) | 3983.33 ± 783.3 | 6716.67 ± 1883.3 |
| Lymphocytes (mm3) | 1904.52 ± 314.1 | 2279.57 ± 699.4 |
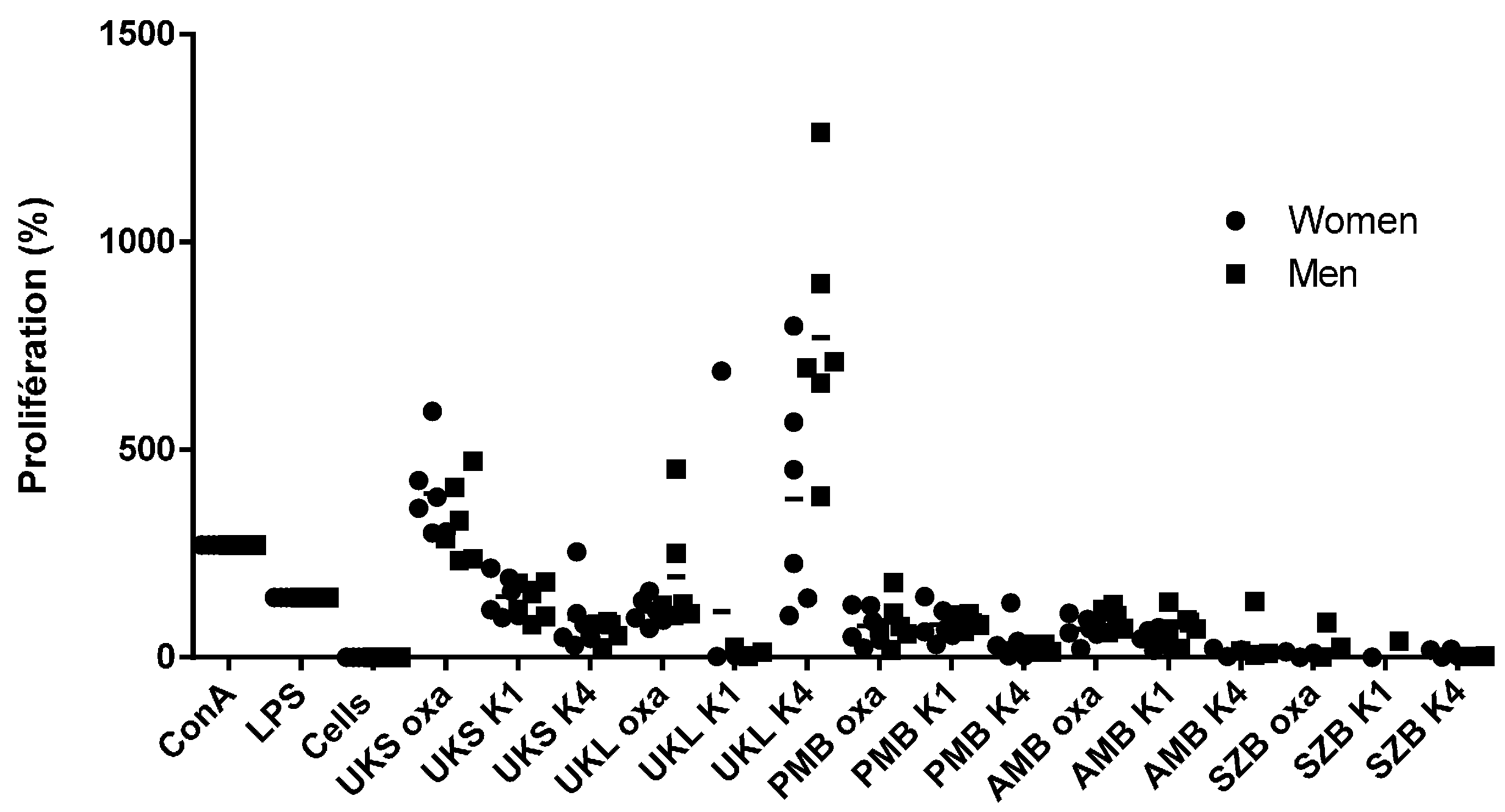
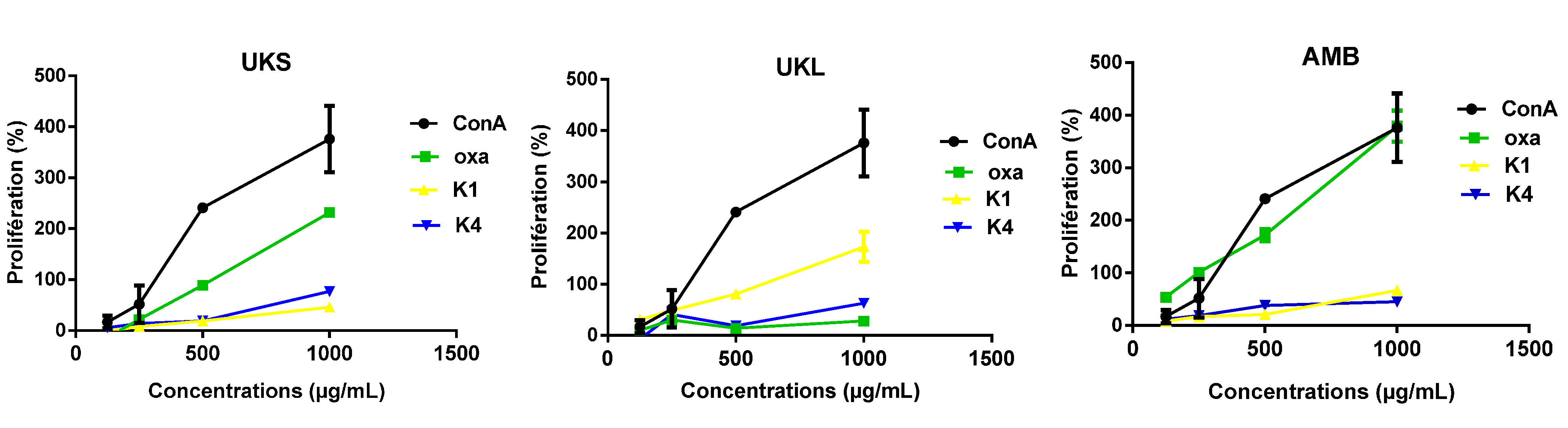
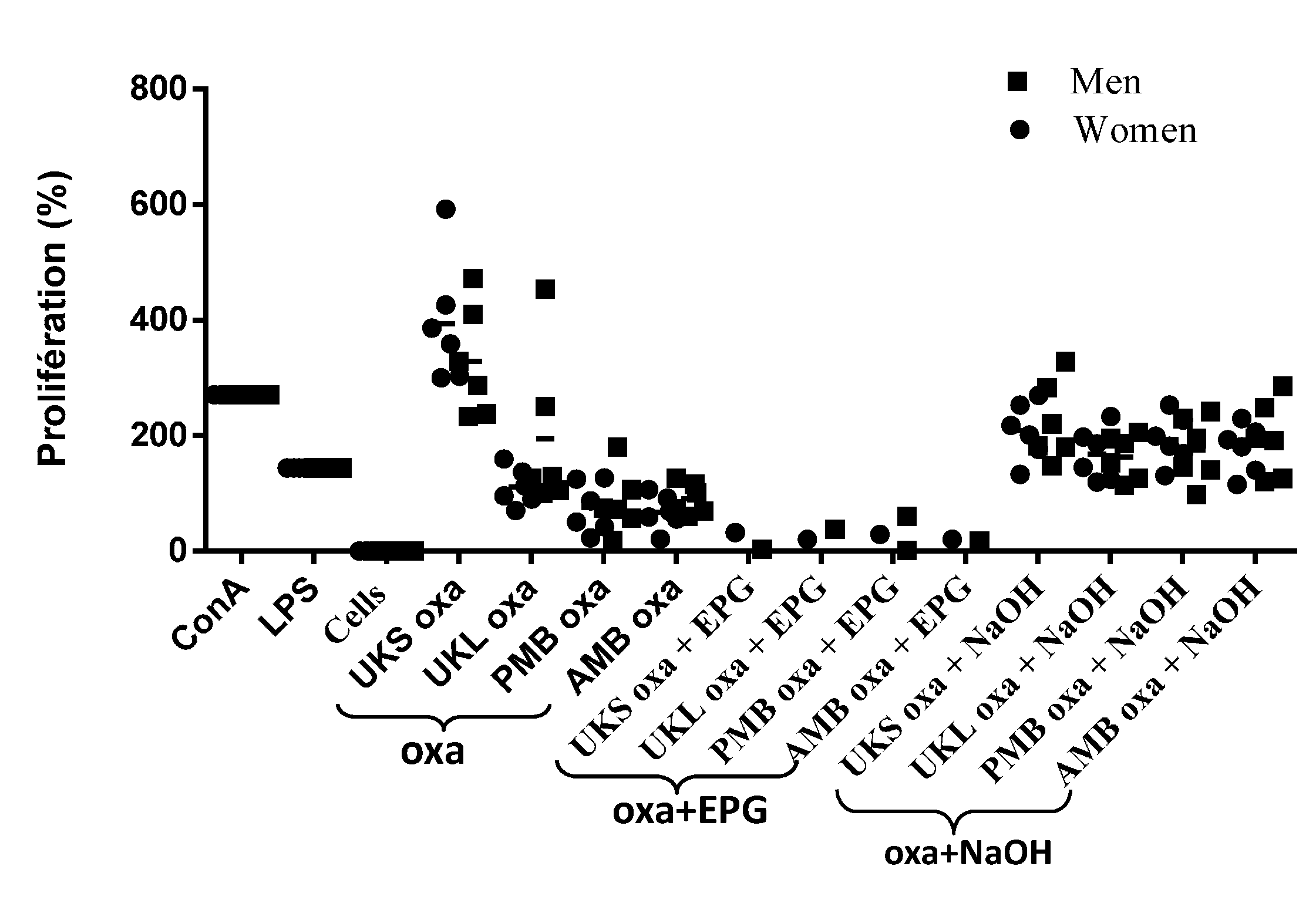
2.4. Analysis of PBMC Proliferation under Polysaccharides Stimulation


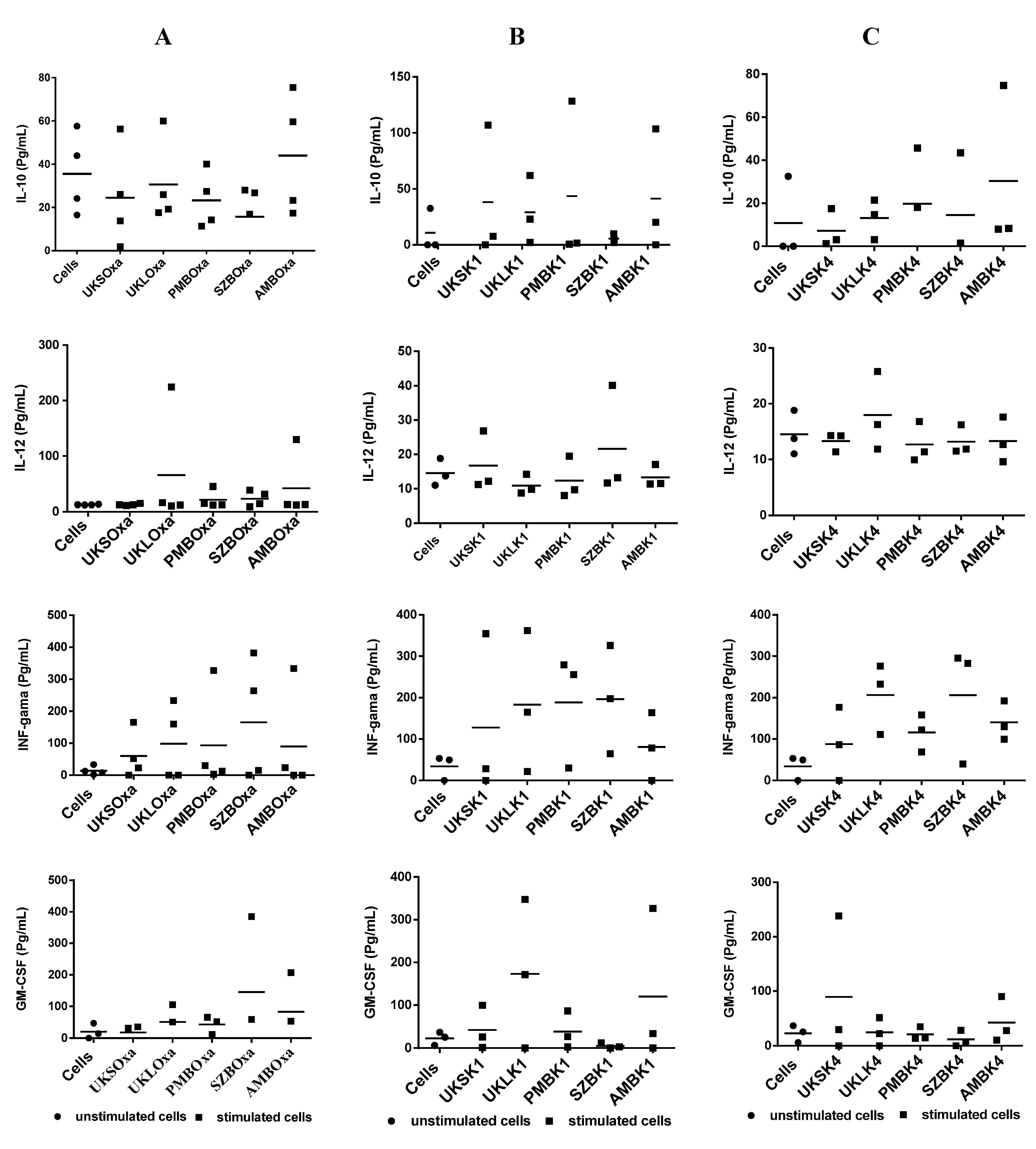
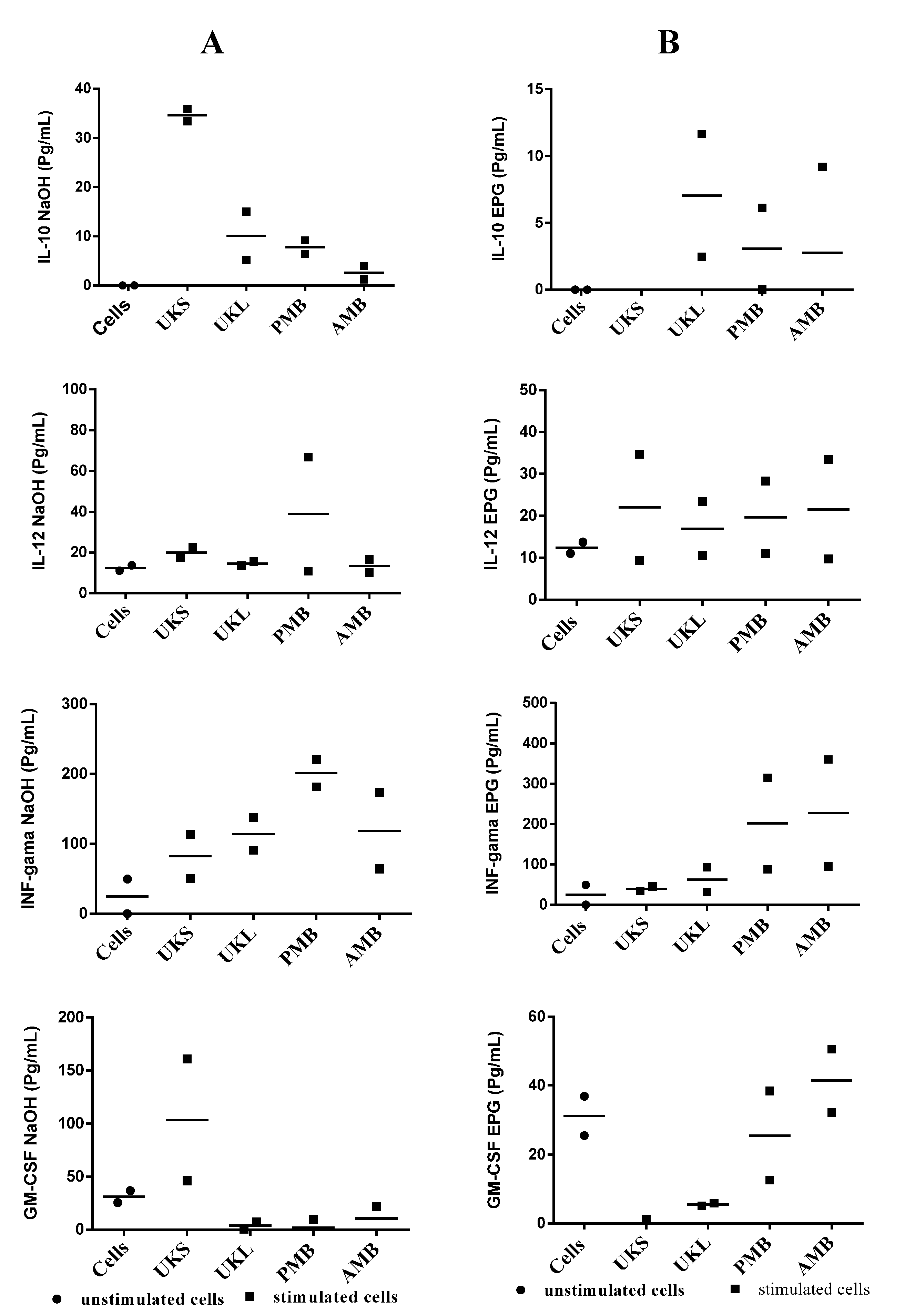
2.5. Analysis of Cytokine Synthesis

2.6. Effect of Chemical and Enzymatic Treatment on the Capacity of Immunostimulation by Polysaccharides


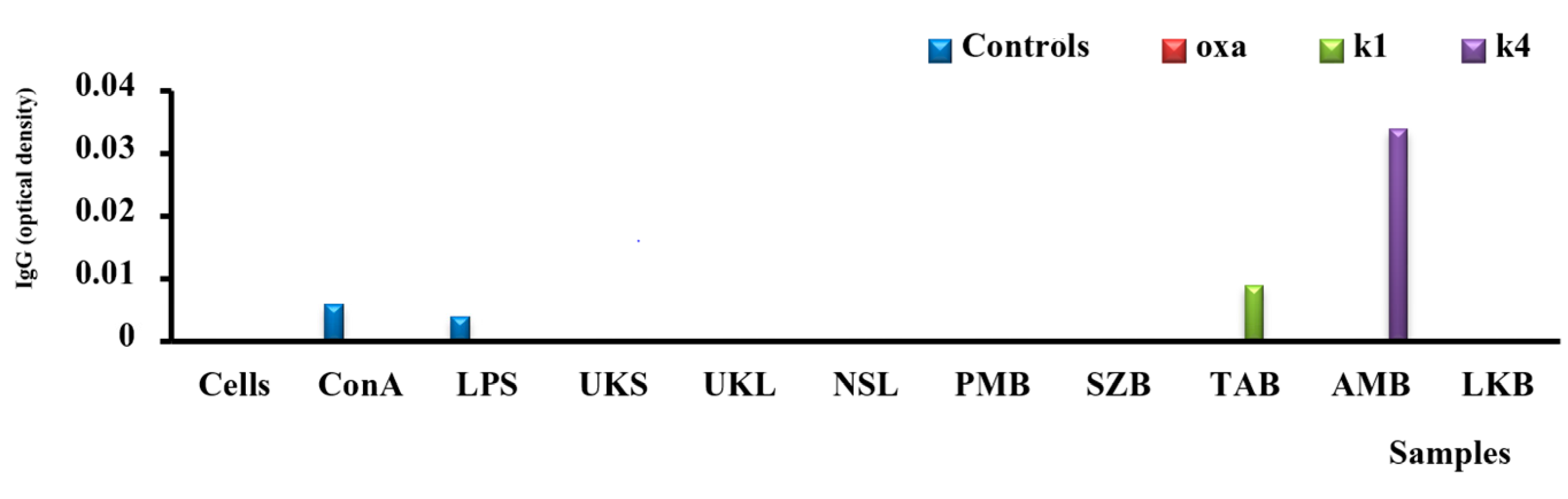
2.7. Analysis of IgG Secretion in Vitro

3. Experimental Section
3.1. Natural Samples
3.2. Human Samples
3.3. Polysaccharides from Cell Wall Extraction and Characterization
3.4. Analysis of PBMC Proliferation
3.5. Analysis of Cytokines Synthesis
3.6. Analysis of the Secretion of IgG
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Letonturier, P. Immunologie Générale,, 7th ed.; Masson: Paris, France, 2001; p. 180. [Google Scholar]
- Espinosa, E.; Chillet, P. Immunologie; Ellipses: Paris, France, 2006; p. 422. [Google Scholar]
- Moser, M.; Leo, O. Key concepts immunology. Vaccine 2010, 28, C2–C13. [Google Scholar] [CrossRef]
- Liu, C.H.; Lin, Q.X.; Gao, Y.; Ye, L.; Xing, Y.Y.; Xi, T. Characterization and antitumor activity of a polysaccharide from Strongylocentrotus nudus eggs. Carbohyd. Polym. 2007, 67, 313–318. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M.; Snader, K.M. The influence of natural products upon drug discovery. Nat. Prod. Rep. 2000, 17, 215–234. [Google Scholar]
- Berlin, C. Herbal medicine. Clin. Pediatr. (Phila) 2001, 40, 271–272. [Google Scholar] [CrossRef]
- Licciardi, P.V.; Underwood, J.R. Plant-derived medicines: A novel class of immunological adjuvants. Int. Immunopharmacol. 2011, 11, 390–398. [Google Scholar] [CrossRef]
- Diallo, D.; Paulsen, S.B.; Liljeba, T.H.A.; Michaelsen, T.E. Polysaccharides from the roots of Entada africana Guill. et Perr., Mimosaceae, with complement fixing activity. J. Pharmacol. 2001, 74, 159–171. [Google Scholar]
- Inngjerdingen, K.T.; Patel, T.R.; Chen, X.; Kenne, L.; Allen, S.; Morris, G.A.; Harding, S.E.; Matsumoto, T.; Diallo, D.; Yamada, H.; et al. Immunological and structural properties of a pectic polymer from Glinus. oppositifolius. Glycobiology 2007, 17, 1299–1310. [Google Scholar]
- Angone, S.A.; Nguema-Ona, E.; Driouich, A. La thérapie par les plantes en Afrique: Activités immunostimulantes des polysaccharides de la paroi végétale. Phytothérapie 2010, 8, 223–230. [Google Scholar] [CrossRef]
- Walker, A.; Sillans, R. Les plantes utiles du Gabon. Pharm. Biol. 1961, 1, 27. [Google Scholar] [CrossRef]
- Ogundaini, A.O.; Ojewole, J.A.O.; Yisak, W. Preliminary phytochemical studies of Combretodendron macrocarpum (P. Beauv) keay with refence to its hypotensive principles. J. Ethnopharmacol. 1983, 9, 337–345. [Google Scholar]
- Mengome, L.E.; Voxeur, A.; Akué, J.P.; Lerouge, P. Screening of antioxidant activities of polysaccharides extracts from endemic plants in Gabon. Bioact. Carbohydr. Diet. Fibre 2014, 3, 77–88. [Google Scholar] [CrossRef]
- Na, S.Y.; Jung, W.J.; Kim, S.M.; Park, J.K.; Lee, S.M.; Kim, O.S.; Synytsya, A.; Park, Y. Purification, characterization and immunostimulating activity of water-soluble polysaccharide isolated from Capsosiphon fulvescens. Int. Immunopharmacol. 2010, 10, 364–370. [Google Scholar] [CrossRef]
- Yoon, Y.D.; Han, S.B.; Kang, J.S.; Lee, C.W.; Park, S.K.; Lee, H.S.; Kang, J.S.; Kim, H.M. Toll-like receptor 4-dependent activation of macrophages by polysaccharide isolated from the radix of Platycodon grandiflorum. Int. Immunopharmacol. 2003, 3, 1873–1882. [Google Scholar] [CrossRef]
- Yoon, Y.D.; Kang, J.S.; Han, S.B.; Park, S.K.; Lee, H.S.; Kang, J.S.; Kim, H.M. Activation of mitogen-activated protein kinases and AP-1 by polysaccharide isolated from the radix of Platycodon grandiflorum in RAW 264.7 cells. Int. Immunopharmacol. 2004, 4, 1477–1487. [Google Scholar] [CrossRef]
- Caux, C.; Dezutter-Dambuyant, C.; Schmitt, D.; Bonchereau, J. GM-CSF ant TNF-α cooperate in the generation of dentritic Langerhans cells. Nature 1992, 360, 258–261. [Google Scholar] [CrossRef]
- Zhang, C.; Huang, K. Characteristic immunostimulation by MAP, a polysaccharide isolated from the mucus of the loach, Misgurnus. anguillicaudatus. Carbohydr. Polym. 2005, 59, 75–82. [Google Scholar] [CrossRef]
- Battaglia, M.; Stabilini, A.; Draghici, E.; Gregori, S.; Mocchetti, C.; Bonifacio, E.; Roncarolo, M.G. Rapamycin and interleukin-10 treatment induces T regulatory type 1 cells that mediate antigen-specific transplantation tolerance. Diabetes 2006, 55, 40–49. [Google Scholar] [CrossRef]
- Georgel, P.; Macquin, C.; Bahram, S. The heterogeneous allelic repertoire of human toll-like receptor (TLR) genes. PLoS One 2009, 4, e7803. [Google Scholar] [CrossRef]
- Priyankar, D.; Tapas, K.C. In vitro modulation of TH1 and TH2 cytokine expression by edible tuber of Dioscorea alata and study of correlation patterns of the cytokine expression. Food Sci. Hum. Wellness 2014, 3, 1–8. [Google Scholar] [CrossRef]
- Maxwell, E.G.; Belshaw, N.J.; Waldron, K.W.; Morris, V.J. Pectin—An emerging new bioactive food polysaccharide. Trends Food Sci. Technol. 2012, 24, 64–73. [Google Scholar] [CrossRef]
- Yang, R.-Y.; Hsu, D.K.; Liu, F.-T. Expression of galectin-3 modulates T-cell growth and apoptosis. Proc. Natl. Acad. Sci. USA 1996, 93, 6737–6742. [Google Scholar] [CrossRef]
- Paclik, D.; Werner, L.; Guckelberger, O.; Wiedenmann, B.; Sturm, A. Galectin distinctively regulate central monocyte and macrophage function. Cell. Immunol. 2011, 271, 97–103. [Google Scholar] [CrossRef]
- Chan, G.C.-F.; Chan, W.K.; Sze, D.M.-Y. The effects of β-glucan on human immune and cancer cells. J. Hematol. Oncol. 2009, 2, 25. [Google Scholar] [CrossRef] [Green Version]
- Akramiené, D.; Kondrotas, A.; Didziapetriené, J.; Kévelaitis, E. Effects of β-glucans on the immune system. Medicina (Kaunas) 2007, 43, 597–606. [Google Scholar]
- Gronhaug, T.E.; Kiyohara, H.; Syveaass, A.; Diallo, D.; Yamada, H.; Paulsen, B.S. β-d-(1,4)-galactan-containing side chains in RG-I regions of pectic polysaccharides from Biophytum petersianum Klotzsch, contribute to expression of immunomodulating activity against intestinal Peyer’s patch cells and macrophages. Phytochemistry 2011, 72, 2139–2147. [Google Scholar] [CrossRef]
- Vos, A.P.; M’Rabet, L.; Stahl, B.; Boehm, G.; Garssen, J. Immune- modulatory effects and potential working mechanisms of orally applied nondigestible carbohydrates. Crit. Rev. Immunol. 2007, 27, 97–140. [Google Scholar] [CrossRef]
- Vetvicka, V.; Thornton, B.P.; Ross, G.D. Soluble β-glucan polysaccharide Binding to the lectin site of neutrophil or natural killer cell complement receptor type 3 (CD11b/CD18) generates a primed state of the receptor capable of mediating cytotoxicity of IC3b-opsonized target cells. J. Clin. Investig. 1996, 98, 50–61. [Google Scholar] [CrossRef]
- Peter, J.R.; Elizabeth, L.; Adams, T.O.-S.; Andres, J.G.; Matthew, P.; Goldman, B.E.; Lockhart, L.A.; Barker, K.F.; Breuel, W.K.; DePonti, J.H.; et al. Oral delivery and gastrointestinal absorption of soluble glucans stimulate increased resistance to infectious challenge. J. Pharmacol. Exp. Ther. 2005, 314, 1079–1086. [Google Scholar]
- Schif, M.A.; Kerperien, J.; Bastiaans, J.; Szklany, K.; Meerding, J.; Horfman, G.; Boon, L.; van Wijk, F.; Garssen, J.; van’t Land, B. Alteration in regulatory T-cells induced by specific oligosaccharides improve vaccine responsiveness in mice. PLoS One 2013, 8, e75148. [Google Scholar] [CrossRef]
- Aubréville, A. Legumineuses caesalpinoïdées. In Flore du Gabon; Muséum National d’histoire Naturelle: Paris, France, 1968; Volume 15, pp. 1–362. [Google Scholar]
- Saint-Aubin de, G. La Forêt du Gabon; Cirad: Paris, France, 1963; pp. 1–208. [Google Scholar]
- Ray, B.; Loutier-Bourhis, C.; Condamine, E.; Driouich, A.; Lerouge, P. Structural investigation of hemicellulosic polysaccharides from Argania spinosa: Characterisation of novel xyloglucan motif. Carbohydr. Res. 2004, 339, 201–208. [Google Scholar] [CrossRef]
- Meral, E.; Johannes, F.; Anthony, A.; Iris, S.; Maria, P.; Benjamin, M.; Peter, G.K. Effect of IL-15 on IgG versus IgE antibody-secreting cells in vitro. J. Immunol. Methods 2012, 375, 7–13. [Google Scholar] [CrossRef]
- De Grandmont, M.J.; Racine, C.; Roy, A.; Lemieux, R.; Néron, S. Intravenous immunoglobulins induce the in vitro differentiation of human B lymphocytes and the secretion of IgG. Blood 2003, 101, 8. [Google Scholar] [CrossRef]
- Sample Availability: Samples of polysacharides UKL, UKS, PmB, TaB, AmB, LKb, NsL, SzB are available from L.E.M. and P.L.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mengome, L.E.; Voxeur, A.; Akue, J.P.; Lerouge, P. In Vitro Proliferation and Production of Cytokine and IgG by Human PBMCs Stimulated with Polysaccharide Extract from Plants Endemic to Gabon. Molecules 2014, 19, 18543-18557. https://doi.org/10.3390/molecules191118543
Mengome LE, Voxeur A, Akue JP, Lerouge P. In Vitro Proliferation and Production of Cytokine and IgG by Human PBMCs Stimulated with Polysaccharide Extract from Plants Endemic to Gabon. Molecules. 2014; 19(11):18543-18557. https://doi.org/10.3390/molecules191118543
Chicago/Turabian StyleMengome, Line Edwige, Aline Voxeur, Jean Paul Akue, and Patrice Lerouge. 2014. "In Vitro Proliferation and Production of Cytokine and IgG by Human PBMCs Stimulated with Polysaccharide Extract from Plants Endemic to Gabon" Molecules 19, no. 11: 18543-18557. https://doi.org/10.3390/molecules191118543