Molecular Cloning and Yeast Expression of Cinnamate 4-Hydroxylase from Ornithogalum saundersiae Baker


Abstract
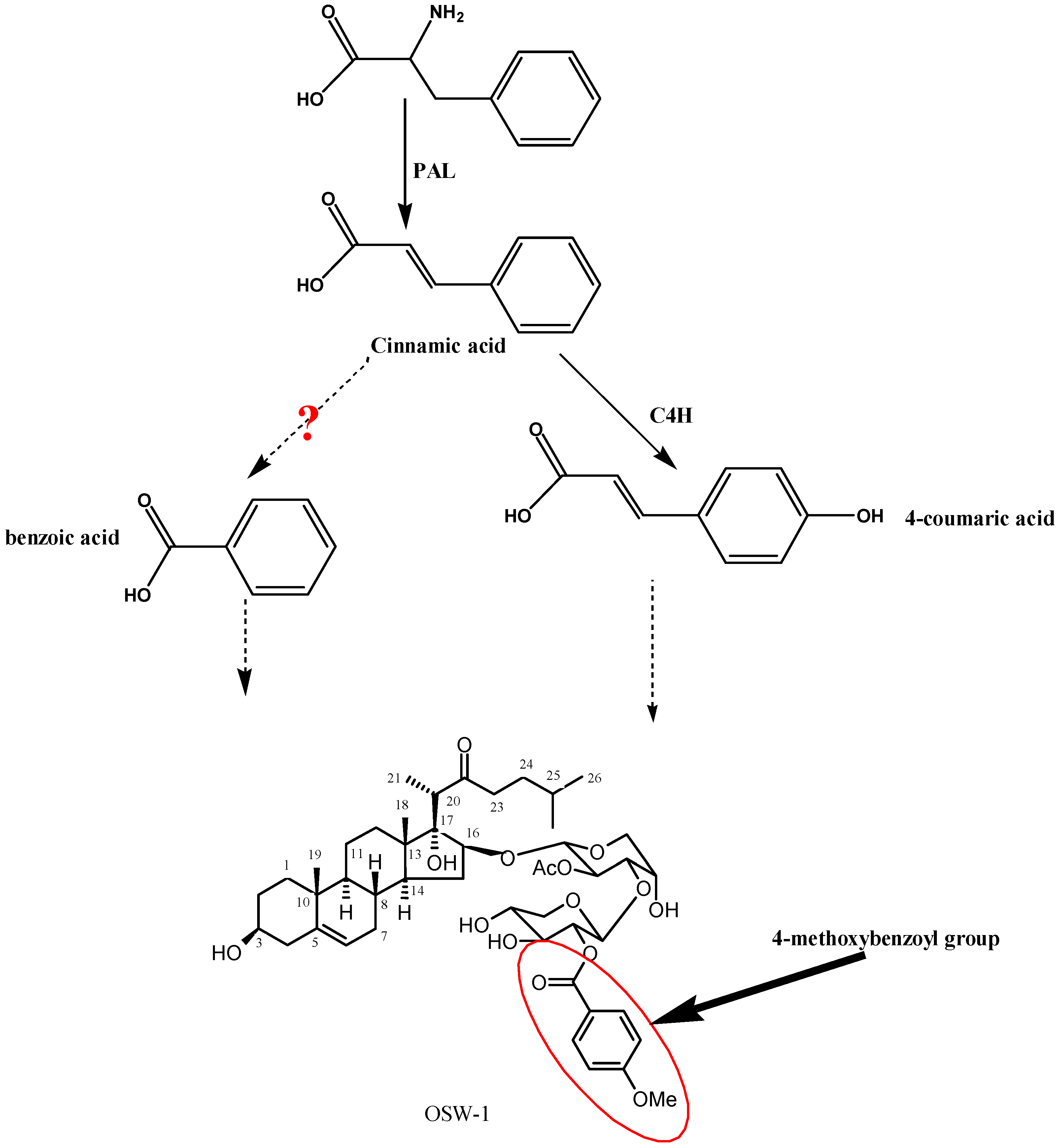
:1. Introduction

2. Results and Discussion
2.1. Transcriptome Analysis of OsaC4H Homology

2.2. Bioinfomational Analysis of Unigene 26946
2.3. Cloning and Analysis of Full-Length Gene Encoding OsaC4H

2.4. Functional Characterization of Recombinant OsaC4H
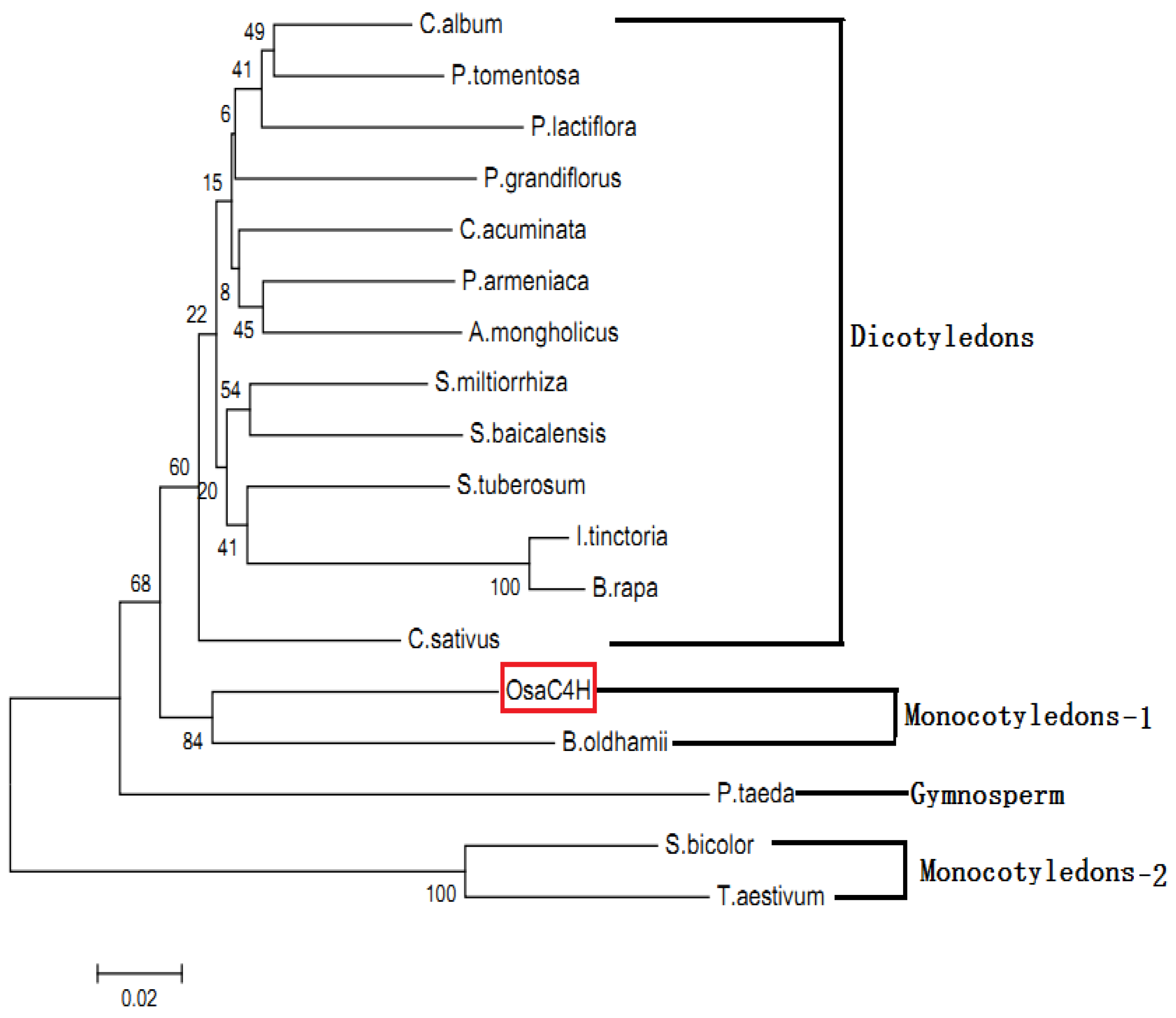
2.5. Molecular Evolution Analysis of OsaC4H
3. Experimental
3.1. Substrates, Chemicals and Enzymes
3.2. Strains and Plasmids

3.3. Plant Materials
3.4. Transcriptome Sequencing and Analysis
3.5. Bioinformatics Analyses
3.6. Generation of Full-length OSaC4H cDNA

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequences(5'-3') |
|---|---|
| Fcin450-1 | 5'- tcttcttcgc ccaagatatc aat -3' |
| Rcin450-1 | 5'- caagcggagc aatcaaaggg aaac -3' |
| Fcin450-2 | 5'- atggacctcc tcctcctaga g -3' |
| Rcin450-2 | 5'- ttagaacacc ctaggtttgg c -3' |
| FYeDP60cin450 | 5'- ctaaattacc ggatccatgg acctcctcct cctagag -3' |
| RYeDP60cin450 | 5'- gatcccccgc gaattcttag aacaccctag gtttggc -3' |
3.7. Expression and Characterization of OsaC4H in Yeast
3.8. Phylogenetic Analysis
4. Conclusions
Acknowledgements
Conflicts of Interest
References
- Mimaki, Y.; Kuroda, M.; Kameyama, A.; Sashida, Y.; Hirano, T.; Oka, K.; Maekawa, R.; Wada, T.; Sugita, K.; Beutler, J.A. Cholestane glycosides with potent cytostatic activities on various tumor cells from Ornithogalum saundersiae bulbs. Bioorg. Med. Chem. Lett. 1997, 7, 633–636. [Google Scholar] [CrossRef]
- Kubo, S.; Mimaki, Y.; Terao, M.; Sashida, Y.; Nikaido, T.; Ohmoto, T. Acylated cholestane glycosides from the bulbs of Ornithogalum saundersiae. Phytochemistry 1992, 31, 3969–3973. [Google Scholar] [CrossRef]
- Tang, Y.; Li, N.; Duan, J.A.; Tao, W. Structure, bioactivity, and chemical synthesis of OSW-1 and other steroidal glycosides in the genus Ornithogalum. Chem. Rev. 2013, 113, 5480–5514. [Google Scholar] [CrossRef]
- Achnine, L.; Blancaflor, E.B.; Rasmussen, S.; Dixon, R.A. Colocalization of L-phenylalanine ammonia-lyase and cinnamate 4-hydroxylase for metabolic channeling in phenylpropanoid biosynthesis. Plant Cell 2004, 16, 3098–3109. [Google Scholar] [CrossRef]
- Kochs, G.; Grisebach, H. Phytoalexin synthesis in soybean: Purification and reconstitution of cytochrome P450 3,9-dihydroxypterocarpan 6a-hydroxylase and separation from cytochrome P450 cinnamate 4-hydroxylase. Arch. Biochem. Biophys. 1989, 273, 543–553. [Google Scholar] [CrossRef]
- Kochs, G.; Werck-Reichhart, D.; Grisebach, H. Further characterization of cytochrome P450 involved in phytoalexin synthesis in soybean: Cytochrome P450 cinnamate 4-hydroxylase and 3,9-dihydroxypterocarpan 6a-hydroxylase. Arch. Biochem. Biophys. 1992, 293, 187–194. [Google Scholar] [CrossRef]
- Singh, K.; Kumar, S.; Rani, A.; Gulati, A.; Ahuja, P.S. Phenylalanine ammonia-lyase (PAL) and cinnamate 4-hydroxylase (C4H) and catechins (flavan-3-ols) accumulation in tea. Funct Integr Genomics 2009, 9, 125–134. [Google Scholar] [CrossRef]
- Xu, H.; Park, N.I.; Li, X.; Kim, Y.K.; Lee, S.Y.; Park, S.U. Molecular cloning and characterization of phenylalanine ammonia-lyase, cinnamate 4-hydroxylase and genes involved in flavone biosynthesis in Scutellaria baicalensis. Bioresour. Technol. 2010, 101, 9715–9722. [Google Scholar] [CrossRef]
- Chen, H.; Jiang, H.; Morgan, J.A. Non-natural cinnamic acid derivatives as substrates of cinnamate 4-hydroxylase. Phytochemistry 2007, 68, 306–311. [Google Scholar] [CrossRef]
- Lu, S.; Zhou, Y.; Li, L.; Chiang, V.L. Distinct roles of cinnamate 4-hydroxylase genes in Populus. Plant Cell Physiol. 2006, 47, 905–914. [Google Scholar] [CrossRef]
- Leonard, E.; Yan, Y.; Lim, K.H.; Koffas, M.A. Investigation of two distinct flavone synthases for plant-specific flavone biosynthesis in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2005, 71, 8241–8248. [Google Scholar] [CrossRef]
- Ro, D.K.; Douglas, C.J. Reconstitution of the entry point of plant phenylpropanoid metabolism in yeast (Saccharomyces cerevisiae): Implications for control of metabolic flux into the phenylpropanoid pathway. J. Biol. Chem. 2004, 279, 2600–2607. [Google Scholar]
- Shin, S.Y.; Jung, S.M.; Kim, M.D.; Han, N.S.; Seo, J.H. Production of resveratrol from tyrosine in metabolically engineered Saccharomyces cerevisiae. Enzyme Microb. Technol. 2011, 51, 211–216. [Google Scholar]
- Watts, K.T.; Lee, P.C.; Schmidt-Dannert, C. Exploring recombinant flavonoid biosynthesis in metabolically engineered Escherichia coli. Chembiochem 2004, 5, 500–507. [Google Scholar] [CrossRef]
- Baek, M.H.; Chung, B.Y.; Kim, J.H.; Kim, J.S.; Lee, S.S.; An, B.C.; Lee, I.J.; Kim, T.H. cDNA cloning and expression pattern of cinnamate-4-hydroxylase in the Korean black raspberry. BMB Rep. 2008, 41, 529–536. [Google Scholar] [CrossRef]
- Chen, A.H.; Chai, Y.R.; Li, J.N.; Chen, L. Molecular cloning of two genes encoding cinnamate 4-hydroxylase (C4H) from oilseed rape (Brassica napus). J. Biochem Mol. Biol. 2007, 40, 247–260. [Google Scholar] [CrossRef]
- Hotze, M.; Schroder, G.; Schroder, J. Cinnamate 4-hydroxylase from Catharanthus roseus, and a strategy for the functional expression of plant cytochrome P450 proteins as translational fusions with P450 reductase in Escherichia coli. FEBS Lett. 1995, 374, 345–350. [Google Scholar] [CrossRef]
- Ro, D.K.; Mah, N.; Ellis, B.E.; Douglas, C.J. Functional characterization and subcellular localization of poplar (Populus trichocarpa x Populus deltoides) cinnamate 4-hydroxylase. Plant Physiol. 2001, 126, 317–329. [Google Scholar] [CrossRef]
- Urban, P.; Werck-Reichhart, D.; Teutsch, H.G.; Durst, F.; Regnier, S.; Kazmaier, M; Pompon, D. Characterization of recombinant plant cinnamate 4-hydroxylase produced in yeast. Kinetic and spectral properties of the major plant P450 of the phenylpropanoid pathway. Eur. J. Biochem. 1994, 222, 843–850. [Google Scholar] [CrossRef]
- Pierrel, M.A.; Batard, Y.; Kazmaier, M.; Mignotte-Vieux, C.; Durst, F.; Werck-Reichhart, D. Catalytic properties of the plant cytochrome P450 CYP73 expressed in yeast. Substrate specificity of a cinnamate hydroxylase. Eur. J. Biochem. 1994, 224, 835–844. [Google Scholar] [CrossRef]
- Werck-Reichhart, D.; Batard, Y.; Kochs, G.; Lesot, A; Durst, F. Monospecific polyclonal antibodies directed against purified cinnamate 4-hydroxylase from Helianthus tuberosus. Immunopurification, immunoquantitation, and interspecies cross-reactivity. Plant Physiol. 1993, 102, 1291–1298. [Google Scholar]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. NCBI BLAST: A better web interface. Nucleic Acids Res. 2008, 36, D5–D16. [Google Scholar] [CrossRef]
- Ye, J.; McGinnis, S.; Madden, T.L. BLAST: Improvements for better sequence analysis. Nucleic Acids Res. 2006, 34, W6–W9. [Google Scholar] [CrossRef]
- McGinnis, S.; Madden, T.L. BLAST: At the core of a powerful and diverse set of sequence analysis tools. Nucleic Acids Res. 2004, 32, W20–25. [Google Scholar] [CrossRef]
- Achamlale, S.; Rezzonico, B.; Grignon-Dubois, M. Evaluation of Zostera detritus as a potential new source of zosteric acid. J. Appl. Phycol. 2009, 21, 347–352. [Google Scholar] [CrossRef]
- Veliky, I.A.; Martin, S.M. A fermenter for plant cell suspension cultures. Can. J. Microbiol. 1970, 16, 223–226. [Google Scholar] [CrossRef]
- Wheeler, D.L.; Church, D.M.; Federhen, S.; Lash, A.E.; Madden, T.L.; Pontius, J.U.; Schuler, G.D.; Schriml, L.M.; Sequeira, E.; Tatusova, T.A.; et al. Database resources of the National Center for Biotechnology. Nucleic Acids Res. 2003, 31, 28–33. [Google Scholar] [CrossRef]
- Sato, Y.; Takaya, A.; Yamamoto, T. Meta-analytic approach to the accurate prediction of secreted virulence effectors in gram-negative bacteria. BMC Bioinforma. 2011, 12, 442. [Google Scholar] [CrossRef]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef]
- Punta, M.; Coggill, P.C.; Eberhardt, R.Y.; Mistry, J.; Tate, J.; Boursnell, C.; Pang, N.; Forslund, K.; Ceric, G.; Clements, J.; et al. The Pfam protein families database. Nucleic Acids Res. 2012, 40, D290–D301. [Google Scholar] [CrossRef]
- Moller, S.; Croning, M.D.; Apweiler, R. Evaluation of methods for the prediction of membrane spanning regions. Bioinformatics (Oxford) 2001, 17, 646–653. [Google Scholar] [CrossRef]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Emanuelsson, O.; Nielsen, H.; Brunak, S.; von Heijne, G. Predicting subcellular localization of proteins based on their N-terminal amino acid sequence. J. Mol. Biol. 2000, 300, 1005–1016. [Google Scholar] [CrossRef]
- Gietz, R.D.; Schiestl, R.H. Quick and easy yeast transformation using the LiAc/SS carrier DNA/PEG method. Nat. Protoc. 2007, 2, 35–37. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the OsaC4H gene and related plasmids are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kong, J.-Q.; Lu, D.; Wang, Z.-B. Molecular Cloning and Yeast Expression of Cinnamate 4-Hydroxylase from Ornithogalum saundersiae Baker. Molecules 2014, 19, 1608-1621. https://doi.org/10.3390/molecules19021608
Kong J-Q, Lu D, Wang Z-B. Molecular Cloning and Yeast Expression of Cinnamate 4-Hydroxylase from Ornithogalum saundersiae Baker. Molecules. 2014; 19(2):1608-1621. https://doi.org/10.3390/molecules19021608
Chicago/Turabian StyleKong, Jian-Qiang, Di Lu, and Zhi-Biao Wang. 2014. "Molecular Cloning and Yeast Expression of Cinnamate 4-Hydroxylase from Ornithogalum saundersiae Baker" Molecules 19, no. 2: 1608-1621. https://doi.org/10.3390/molecules19021608