Penostatin Derivatives, a Novel Kind of Protein Phosphatase 1B Inhibitors Isolated from Solid Cultures of the Entomogenous Fungus Isaria tenuipes

Abstract
:1. Introduction

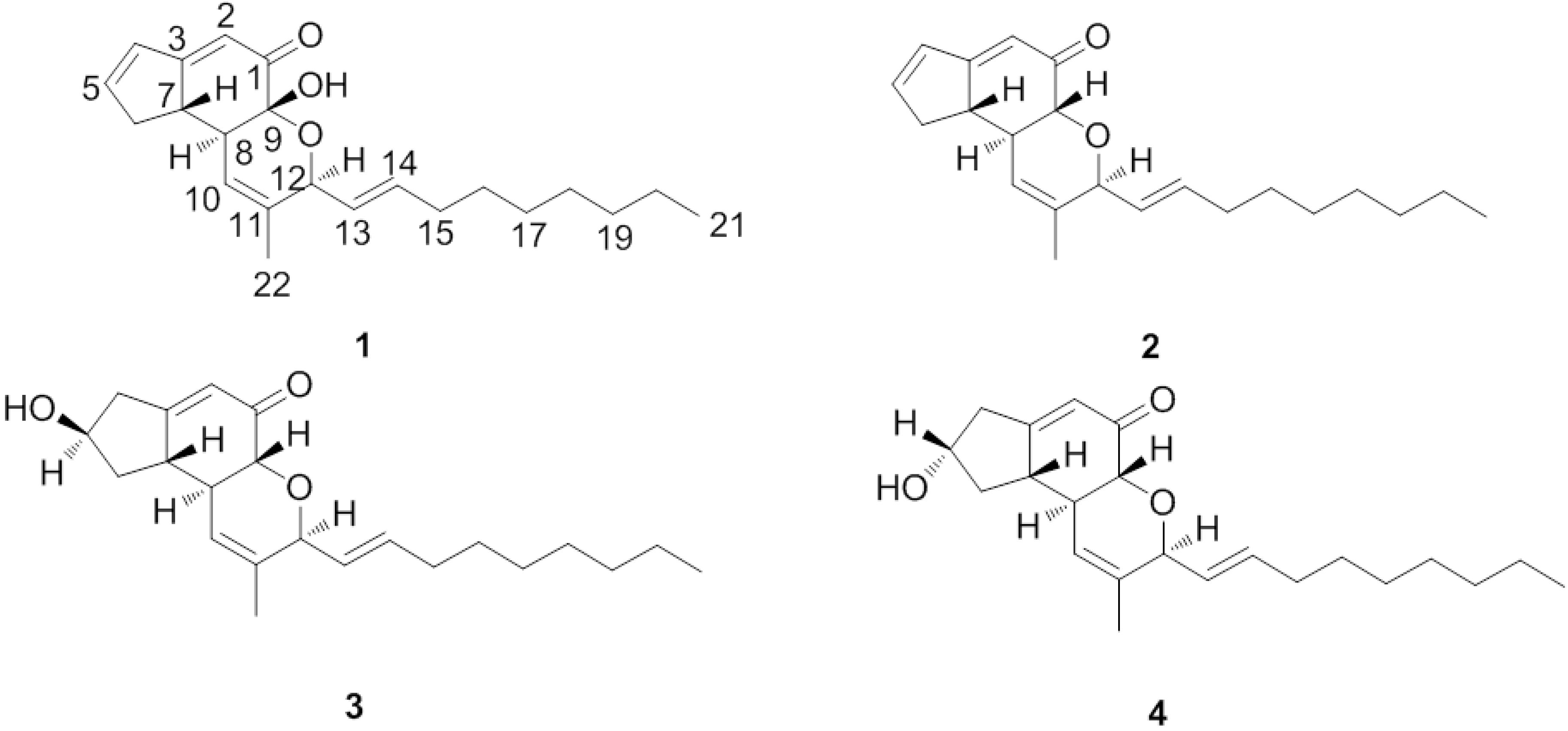
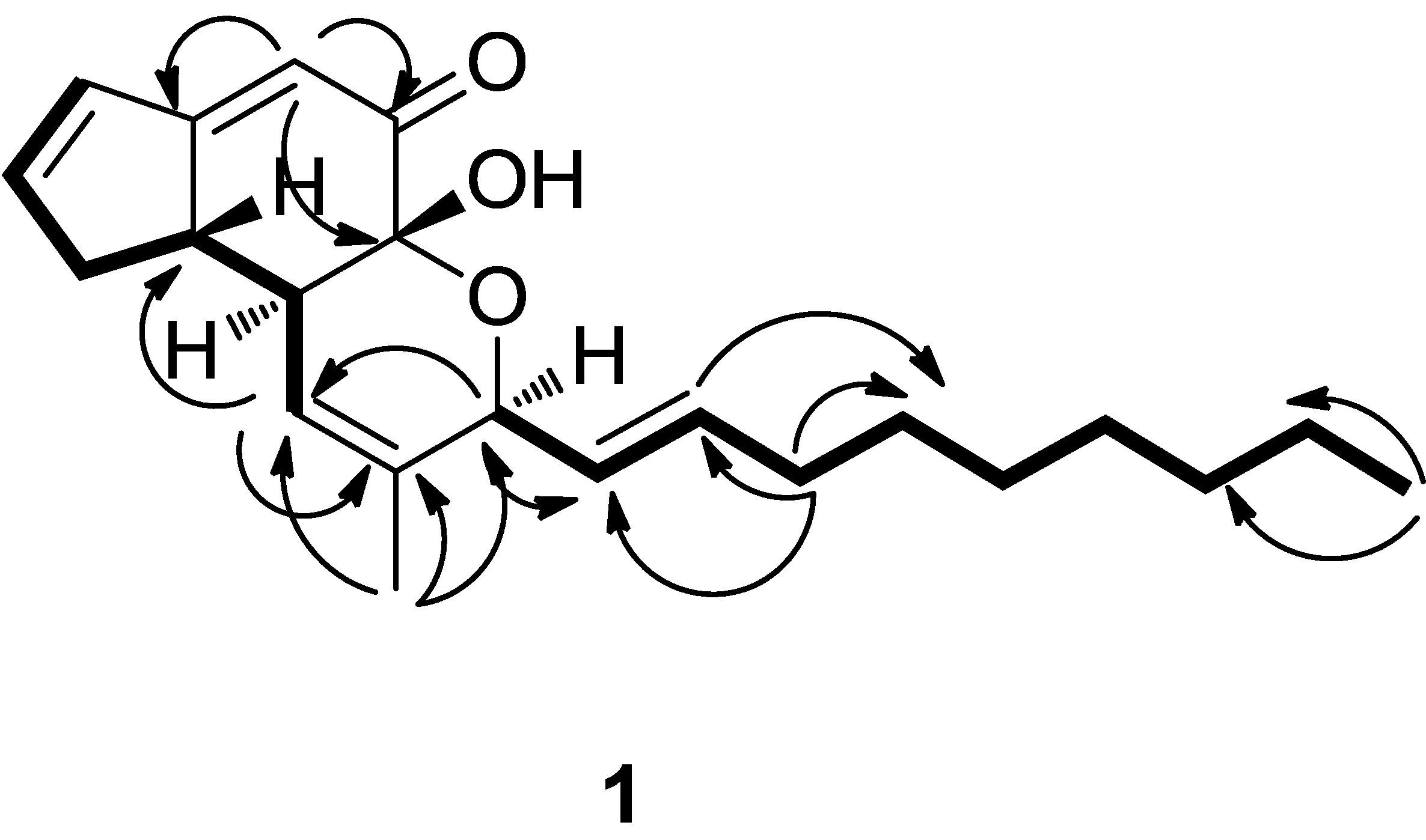
2. Results and Discussion
PTP1B Activities

{kind=link}
{kind=link}
| Position | 1 | Penostatin C (2) | ||
|---|---|---|---|---|
| δ(H) | δ(C) | δ(H) | δ(C) | |
| 1 | 194.7 | 197.8 | ||
| 2 | 5.95 (d, J = 2.4 Hz) | 113.8 | 5.87 (d, J = 2.4 Hz) | 115.7 |
| 3 | 174.6 | 173.8 | ||
| 4 | 6.54 (dt, J = 5.6; 2.3 Hz) | 131.4 | 6.52 (dt, J = 5.5; 2.4 Hz) | 131.6 |
| 5 | 6.87 (dt, J = 5.6; 2.3 Hz) | 150.9 | 6.82 (dt, J = 5.5; 2.3 Hz) | 149.4 |
| 6 | 2.90 (dddd, J = 16.9; 5.9; 1.5; 2.0 Hz) | 36.8 | 2.89 (dddd, J = 17.5; 6.5; 1.5; 2.0 Hz) | 36.2 |
| 2.52 (dddd, J = 16.9; 4.1; 1.5; 2.0 Hz) | 2.50 (dddd, J = 17.5; 4.2; 1.5; 2.0 Hz) | |||
| 7 | 2.99 (dddd, J = 10.8; 7; 3.7; 2.3 Hz) | 46.5 | 2.77 (dddd, J = 11.2; 6.8; 3.6; 2.5 Hz) | 45.7 |
| 8 | 2.28 (tq, J = 10.8; 1.9 Hz) | 44.9 | 2.49 (tq, J = 11.4; 2.2 Hz) | 44.9 |
| 9 | 91.9 | 4.44 (d, J = 11.1 Hz) | 74.9 | |
| 10 | 5.76 (m) | 119.7 | 5.67 (m) | 115.7 |
| 11 | 134.5 | 135.9 | ||
| 12 | 4.67 (d, J = 6.5 Hz) | 76.2 | 4.57 (d, J = 6.4 Hz) | 77.7 |
| 13 | 5.37 (dd, J = 6.5; 15.5 Hz) | 128.1 | 5.62 (dd, J = 6.6; 15.5 Hz) | 126.3 |
| 14 | 5.79 (dd, J = 15.5, 6.5 Hz) | 135.6 | 5.74 (dt, J = 15.5; 6.9 Hz) | 136.2 |
| 15 | 2.08 (m) | 31.8 | 2.09 (m) | 31.8 |
| 16 | 1.42 (m) | 28.7 | 1.31 (m) | 28.6 |
| 17 | 1.31 (brs) | 28.8 | 1.28 (brs) | 28.7 |
| 18 | 1.31 (brs) | 28.7 | 1.28 (brs) | 28.6 |
| 19 | 1.31 (brs) | 31.6 | 1.28 (brs) | 31.6 |
| 20 | 1.31 (brs) | 22.3 | 1.28 (brs) | 22.3 |
| 21 | 0.91(t, J = 6.8 Hz) | 12.9 | 0.88(t, J = 6.8 Hz) | 13.0 |
| 22 | 1.65 (s) | 18.2 | 1.59 (s) | 18.7 |

| Compound | IC50 (μM) | |
|---|---|---|
| PTP1B | LAR a | |
| 1 | 12.53 ± 0.9 | a |
| 2 | 0.37 ± 0.05 | 53.33 ± 4.06 |
| 3 | 15.87 ± 2.49 | a |
| 4 | 33.65 ± 7.33 | a |
| Sodium orthovanadate (positive control) | 0.65 ± 0.08 | 0.89 ± 0.02 |
3. Experimental
3.1. General
3.2. Fungal Material and Cultivation Conditions
3.3. Extraction and Isolation
3.4. Physico-Chemical Properties
 = 59.3 (c = 0.003, MeOH). IR (KBr) vmax: 3442, 2926, 2854, 1659, 1618 cm−1. UV (CHCl3) λmax (lg ε): 289 (3.45), 203 (3.35) nm. 1H- and 13C-NMR, see Table 1. Positive ion HR-ESI-MS [M+Na]+ m/z 365.20820 (calcd for C22H30O3Na, 365.20872). = 87.5 (c = 2.2, MeOH). IR (KBr) vmax: 3427, 2926, 2854, 1658, 1618 cm−1. UV (CHCl3) λmax (lg ε): 280 (3.39), 0.3422 (3.54) nm. 1H- and 13C-NMR, see Table 1. Positive ion HR-ESI-MS [M+Na]+ m/z 349.22453 (calcd for C22H30O2Na, 349.22445). It was identified as penostatin C by comparison of the spectral data with the literature [17,18]. = +56.8 (c = 2.2, MeOH). IR (KBr) vmax: 3425, 2924, 2853, 1675, 1633 cm−1. UV (CHCl3) λmax (lg ε): 253 (3.78), 202 (3.90) nm. 1H-NMR (CDCl3) δ 5.64 (1H, s, H-2), 2.42 (1H, d, J = 1.74 Hz, H-4α), 2.98 (1H, dd, J = 7.5 Hz, 18.5, H-4β), 4.43 ((1H, m, H-5), 2.53 (1H, ddd, J = 11.8,7.0,1.9 Hz, H-6α), 1.50 (1H, td, J = 12.5,4.8 Hz, H-6β), 2.45 (1H, m, H-7), 2.53 (1H, m, H-8), 4.08 (1H, d, J = 11.5 Hz, H-9), 5.92 (1H, m, H-10), 4.55 (1H, d, J = 6.5 Hz, H-12), 5.62 (1H, dd, J = 13.9,5.6 Hz, H-13), 5.74(1H, dt, J = 13.8,6.7 Hz, H-14), 2.12 (2H, m, H-15), 1.42 (2H, m, H-16), 1.31 (4H, brs, H-17, 18, 19, 20), 0.91 (3H, t, J = 6.8 Hz, H-21), 1.69 (3H, s, H-22). 13C-NMR (CDCl3) δ 197.1 (s, C-1), 121.5 (d, C-2), 172.3 (s, C- 3), 40.9 (t, C-4), 70.2 (d, C-5), 37.8 (t, C-6), 45.4 (d, C-7), 44.9 (d, C-8), 73.7 (d, C-9), 121.2 (d, C-10), 135.9 (s, C-11), 77.5 (d, C-12), 126.3 (d, C-13), 135.8 (d, C-14), 31.8 (t, C-15), 28.7 (t, C-16), 28.7 (t, C-17), 28.7 (t, C-18), 31.6 (t, C-19), 22.3 (t, C-20), 13.0 (q, C-21), 18.7 (q, C-22). Positive ion HR-ESI-MS [M+Na]+ m/z 367.23543 (calcd for C22H32O3Na, 367.23514). It was identified as penostatin A by comparison of the spectral data with the literature [17,18]. = −24.5 (c = 2.2, MeOH). IR (KBr) vmax: 3460, 2921, 1671, 1632, 1466 cm−1. UV (CHCl3) λmax (lg ε): 234 (3.43), 202 (3.45) nm. 1H-NMR (CDCl3) δ 5.65 (1H, s, H-2), 2.36 (1H, t, J = 1.68 Hz, H-4α), 2.87 (1H, m, H-4β), 4.50 (1H, m, H-5), 2.63 (1H, d, J = 19.8 Hz, H-6α), 1.57 (1H, ddd, J = 10.7, 9.2, 5.9 Hz, H-6β), 2.27 (1H, m, H-7), 2.85 (1H, m, H-8), 4.14 (1H, d, J = 11.6 Hz, H-9), 5.93 (1H, m, H-10), 4.57 (1H, d, J = 6.5 Hz, H-12), 5.62 (1H, dd, J = 5.5, 6.0 Hz, H-13), 5.74(1H, dt, J = 13.8, 6.7 Hz, H-14), 2.12 (2H, m, H-15), 1.42 (2H, m, H-16), 1.31 (4H, brs, H-17, 18, 19, 20), 0.91 (3H, t, J = 6.8 Hz, H-21), 1.69 (3H, s, H-22). 13C-NMR (CDCl3) δ 198.3 (s, C-1), 122.9 (d, C-2), 174.5 (s, C-3), 42.4 (t, C-4), 71.4 (d, C-5), 39.5 (t, C-6), 46.2 (d, C-7), 46.1 (d, C-8), 75.1 (d, C-9), 122.4 (d, C-10), 137.3 (s, C-11), 78.8 (d, C-12), 127.5 (d, C-13), 137.1 (d, C-14), 33.1 (t, C-15), 29.9 (t, C-16), 29.9 (t, C-17), 30.0 (t, C-18), 32.9 (t, C-19), 23.5 (t, C-20), 14.3 (q, C-21), 19.9 (C-22). Positive ion HR-ESI-MS [M+Na]+ m/z 367.23523 (calcd for C22H32O3Na, 367.23514). It was identified as penostatin B by comparison of the spectral data with the literature [17,18].
= 59.3 (c = 0.003, MeOH). IR (KBr) vmax: 3442, 2926, 2854, 1659, 1618 cm−1. UV (CHCl3) λmax (lg ε): 289 (3.45), 203 (3.35) nm. 1H- and 13C-NMR, see Table 1. Positive ion HR-ESI-MS [M+Na]+ m/z 365.20820 (calcd for C22H30O3Na, 365.20872). = 87.5 (c = 2.2, MeOH). IR (KBr) vmax: 3427, 2926, 2854, 1658, 1618 cm−1. UV (CHCl3) λmax (lg ε): 280 (3.39), 0.3422 (3.54) nm. 1H- and 13C-NMR, see Table 1. Positive ion HR-ESI-MS [M+Na]+ m/z 349.22453 (calcd for C22H30O2Na, 349.22445). It was identified as penostatin C by comparison of the spectral data with the literature [17,18]. = +56.8 (c = 2.2, MeOH). IR (KBr) vmax: 3425, 2924, 2853, 1675, 1633 cm−1. UV (CHCl3) λmax (lg ε): 253 (3.78), 202 (3.90) nm. 1H-NMR (CDCl3) δ 5.64 (1H, s, H-2), 2.42 (1H, d, J = 1.74 Hz, H-4α), 2.98 (1H, dd, J = 7.5 Hz, 18.5, H-4β), 4.43 ((1H, m, H-5), 2.53 (1H, ddd, J = 11.8,7.0,1.9 Hz, H-6α), 1.50 (1H, td, J = 12.5,4.8 Hz, H-6β), 2.45 (1H, m, H-7), 2.53 (1H, m, H-8), 4.08 (1H, d, J = 11.5 Hz, H-9), 5.92 (1H, m, H-10), 4.55 (1H, d, J = 6.5 Hz, H-12), 5.62 (1H, dd, J = 13.9,5.6 Hz, H-13), 5.74(1H, dt, J = 13.8,6.7 Hz, H-14), 2.12 (2H, m, H-15), 1.42 (2H, m, H-16), 1.31 (4H, brs, H-17, 18, 19, 20), 0.91 (3H, t, J = 6.8 Hz, H-21), 1.69 (3H, s, H-22). 13C-NMR (CDCl3) δ 197.1 (s, C-1), 121.5 (d, C-2), 172.3 (s, C- 3), 40.9 (t, C-4), 70.2 (d, C-5), 37.8 (t, C-6), 45.4 (d, C-7), 44.9 (d, C-8), 73.7 (d, C-9), 121.2 (d, C-10), 135.9 (s, C-11), 77.5 (d, C-12), 126.3 (d, C-13), 135.8 (d, C-14), 31.8 (t, C-15), 28.7 (t, C-16), 28.7 (t, C-17), 28.7 (t, C-18), 31.6 (t, C-19), 22.3 (t, C-20), 13.0 (q, C-21), 18.7 (q, C-22). Positive ion HR-ESI-MS [M+Na]+ m/z 367.23543 (calcd for C22H32O3Na, 367.23514). It was identified as penostatin A by comparison of the spectral data with the literature [17,18]. = −24.5 (c = 2.2, MeOH). IR (KBr) vmax: 3460, 2921, 1671, 1632, 1466 cm−1. UV (CHCl3) λmax (lg ε): 234 (3.43), 202 (3.45) nm. 1H-NMR (CDCl3) δ 5.65 (1H, s, H-2), 2.36 (1H, t, J = 1.68 Hz, H-4α), 2.87 (1H, m, H-4β), 4.50 (1H, m, H-5), 2.63 (1H, d, J = 19.8 Hz, H-6α), 1.57 (1H, ddd, J = 10.7, 9.2, 5.9 Hz, H-6β), 2.27 (1H, m, H-7), 2.85 (1H, m, H-8), 4.14 (1H, d, J = 11.6 Hz, H-9), 5.93 (1H, m, H-10), 4.57 (1H, d, J = 6.5 Hz, H-12), 5.62 (1H, dd, J = 5.5, 6.0 Hz, H-13), 5.74(1H, dt, J = 13.8, 6.7 Hz, H-14), 2.12 (2H, m, H-15), 1.42 (2H, m, H-16), 1.31 (4H, brs, H-17, 18, 19, 20), 0.91 (3H, t, J = 6.8 Hz, H-21), 1.69 (3H, s, H-22). 13C-NMR (CDCl3) δ 198.3 (s, C-1), 122.9 (d, C-2), 174.5 (s, C-3), 42.4 (t, C-4), 71.4 (d, C-5), 39.5 (t, C-6), 46.2 (d, C-7), 46.1 (d, C-8), 75.1 (d, C-9), 122.4 (d, C-10), 137.3 (s, C-11), 78.8 (d, C-12), 127.5 (d, C-13), 137.1 (d, C-14), 33.1 (t, C-15), 29.9 (t, C-16), 29.9 (t, C-17), 30.0 (t, C-18), 32.9 (t, C-19), 23.5 (t, C-20), 14.3 (q, C-21), 19.9 (C-22). Positive ion HR-ESI-MS [M+Na]+ m/z 367.23523 (calcd for C22H32O3Na, 367.23514). It was identified as penostatin B by comparison of the spectral data with the literature [17,18].3.5. Biological Evaluation
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hunter, T. Signaling—2000 and beyond. Cell 2000, 100, 113–127. [Google Scholar] [CrossRef]
- Manning, G.; Whyte, D.B.; Martinez, R.; Hunter, T.; Sudarsanam, S. The protein kinase complement of the human genome. Science 2002, 298, 1912–1934. [Google Scholar] [CrossRef]
- Forghieri, M.; Laggner, C.; Paoli, P.; Langer, T.; Manao, G.; Camici, G.; Bondioli, L.; Prati, F.; Costantino, L. Synthesis, activity and molecular modeling of a new series of chromones as low molecular weight protein tyrosine phosphatase inhibitors. Bioorg. Med. Chem. 2009, 17, 2658–2672. [Google Scholar] [CrossRef]
- Andersen, J.N.; Mortensen, O.H.; Peters, G.H.; Drake, P.G.; Iversen, L.F.; Olsen, O.H.; Jansen, P.G.; Andersen, H.S.; Tonks, N.K.; Moller, N.P. Structural and evolutionary relationships among protein tyrosine phosphatase domains. Mol. Cell. Biol. 2001, 21, 7117–7136. [Google Scholar] [CrossRef]
- Van Huijsduijnen, R.H.; Sauer, W.H.B.; Bombrun, A.; Swinnen, D. Prospects for inhibitors of protein tyrosine phosphatase 1B as antidiabetic drugs. J. Med. Chem. 2004, 47, 4142–4146. [Google Scholar] [CrossRef]
- Barr, A.J. Protein tyrosine phosphatases as drug targets: Strategies and challenges of inhibitor development. Future Med. Chem. 2010, 2, 1563–1576. [Google Scholar] [CrossRef]
- Liu, G. Protein tyrosine phosphatase 1B inhibition: Opportunities and challenges. Curr. Med. Chem. 2003, 10, 1407–1421. [Google Scholar] [CrossRef]
- Hajek, A.E.; Leger, R.J., St. Interactions between fungal pathogens and insect hosts. Rev. Entomol. 1994, 39, 293–322. [Google Scholar] [CrossRef]
- Isaka, M.; Kittakoop, P.; Kirtikara, K.; Hywel-Jones, N.L.; Thebtaranonth, Y. Bioactive substances from insect pathogenic fungi. Acc. Chem. Res. 2005, 38, 813–823. [Google Scholar] [CrossRef]
- Namatame, I.; Tomoda, H.; Tabata, N.; Si, S.; Ōmura, S. Structure elucidation of fungal beauveriolide III, a novel inhibitor of lipid droplet formation in mouse macrophages. J. Antibiot. 1999, 52, 7–12. [Google Scholar] [CrossRef]
- Takahashi, S.; Uchida, K.; Kakinuma, N.; Hashimoto, R.; Yanagisawa, T.; Nakagawa, A. The structures of pyridovericin and pyridomacrolidin, new metabolites from the entomopathogenic fungus, Beauveria Bassiana. J. Antibiot. 1998, 51, 1051–1054. [Google Scholar] [CrossRef]
- Fujita, T.; Inoue, K.; Yamamoto, S.; Ikumoto, T.; Sasaki, S.; Toyama, R.; Chiba, K.; Hoshino, Y.; Okumoto, T. Fungal metabolites: Part 11. A potent immunosuppressive activity found in Isaria sinclairii metabolite. J. Antibiot. 1994, 47, 208–215. [Google Scholar] [CrossRef]
- Strader, C.R.; Pearce, C.J.; Oberlies, N.H. Fingolimod (FTY720): A recently approved multiple sclerosis drug based on a fungal secondary metabolite. J. Nat. Prod. 2011, 74, 900–907. [Google Scholar] [CrossRef]
- Isaka, M.; Palasarn, S.; Tobwor, P.; Boonruangprapa, T.; Tasanathai, K. Bioactive anthraquinone dimers from the leafhopper pathogenic fungus Torrubiella sp. BCC 28517. J. Antibiot. 2012, 11, 571–574. [Google Scholar]
- Isaka, M.; Chinthanom, P.; Sappan, M.; Chanthaket, R.; Luangsa-ard, J.J.; Prabpai, S.; Kongsaeree, P. Lanostane and hopane triterpenes from the entomopathogenic fungus Hypocrella sp. BCC 14524. J. Nat. Prod. 2011, 74, 2143–2150. [Google Scholar] [CrossRef]
- Zhu, J.S.; Halpern, G.M.; Jones, K.J. The scientific rediscovery of an ancient chinese herbal medicine: Cordyceps sinensis: Part I. J. Altern Complement. Med. 1998, 4, 289–303. [Google Scholar] [CrossRef]
- Takahashi, C.; Numata, A.; Yamada, T.; Minoura, K.; Enomoto, S.; Konishi, K.; Nakai, M.; Matsuda, C.; Nomoto, K. Penostatins, novel cytotoxic metabolites from a Penicillium species separated from a green alga. Tetrahedron Lett. 1996, 37, 655–658. [Google Scholar] [CrossRef]
- Iwamoto, C.; Minoura, K.; Oka, T.; Ohta, T.; Hagishita, S.; Numata, A. Absolute stereostructures of novel cytotoxic metabolites, penostatins A-E, from a Penicillium species separated from an Enteromorpha alga. Tetrahedron 1999, 55, 14353–14368. [Google Scholar] [CrossRef]
- Scott, L.M.; Lawrence, H.R.; Sebti, S.M.; Lawrence, N.J.; Wu, J. Targeting protein tyrosine phosphatases for anticancer drug discovery. Curr. Pharm. Des. 2010, 16, 1843–1862. [Google Scholar] [CrossRef]
- Bleasdale, J.E.; Ogg, D.; Palazuk, B.J.; Jacob, C.S.; Swanson, M.L.; Wang, X.Y.; Thompson, D.P.; Conradi, R.A.; Mathews, W.R.; Laborde, A.L.; et al. Small molecule peptidomimetics containing a novel phosphotyrosine bioisostere inhibit protein tyrosine phosphatase 1B and augment insulin action. Biochemistry 2001, 40, 5642–5654. [Google Scholar] [CrossRef]
- Baykov, A.A.; Evtushenko, O.A.; Avaeva, S.M. A malachite green procedure for orthophosphate determination and its use in alkaline phosphatase-based enzyme immunoassay. Anal. Biochem. 1988, 171, 266–270. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chen, Y.-P.; Yang, C.-G.; Wei, P.-Y.; Li, L.; Luo, D.-Q.; Zheng, Z.-H.; Lu, X.-H. Penostatin Derivatives, a Novel Kind of Protein Phosphatase 1B Inhibitors Isolated from Solid Cultures of the Entomogenous Fungus Isaria tenuipes. Molecules 2014, 19, 1663-1671. https://doi.org/10.3390/molecules19021663
Chen Y-P, Yang C-G, Wei P-Y, Li L, Luo D-Q, Zheng Z-H, Lu X-H. Penostatin Derivatives, a Novel Kind of Protein Phosphatase 1B Inhibitors Isolated from Solid Cultures of the Entomogenous Fungus Isaria tenuipes. Molecules. 2014; 19(2):1663-1671. https://doi.org/10.3390/molecules19021663
Chicago/Turabian StyleChen, Yu-Peng, Chun-Gui Yang, Pei-Yao Wei, Lin Li, Du-Qiang Luo, Zhi-Hui Zheng, and Xin-Hua Lu. 2014. "Penostatin Derivatives, a Novel Kind of Protein Phosphatase 1B Inhibitors Isolated from Solid Cultures of the Entomogenous Fungus Isaria tenuipes" Molecules 19, no. 2: 1663-1671. https://doi.org/10.3390/molecules19021663