Rice Bran Feruloylated Oligosaccharides Activate Dendritic Cells via Toll-Like Receptor 2 and 4 Signaling



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
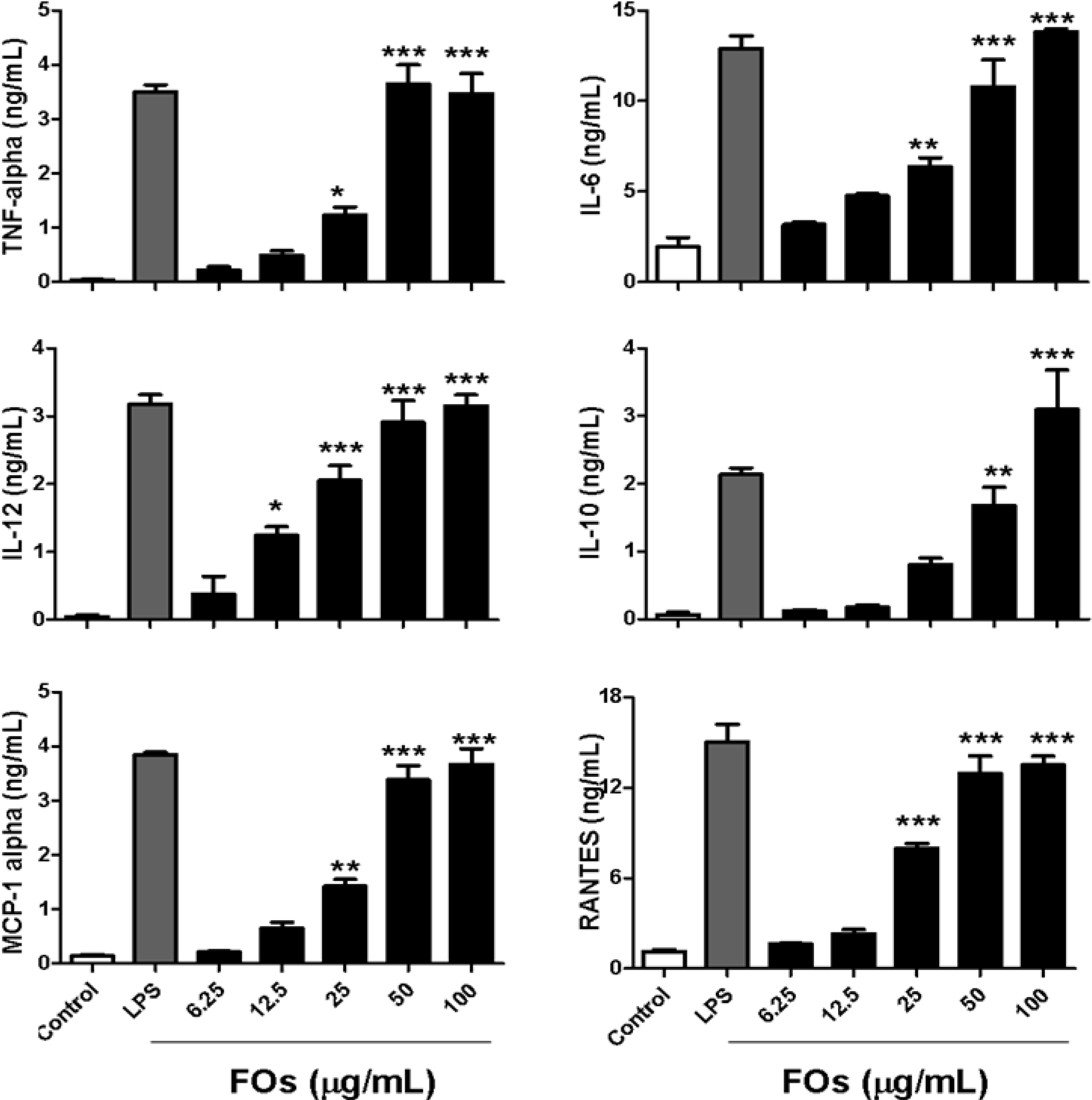
2.1. FOs Increase Cytokine and Chemokine Production by BMDCs
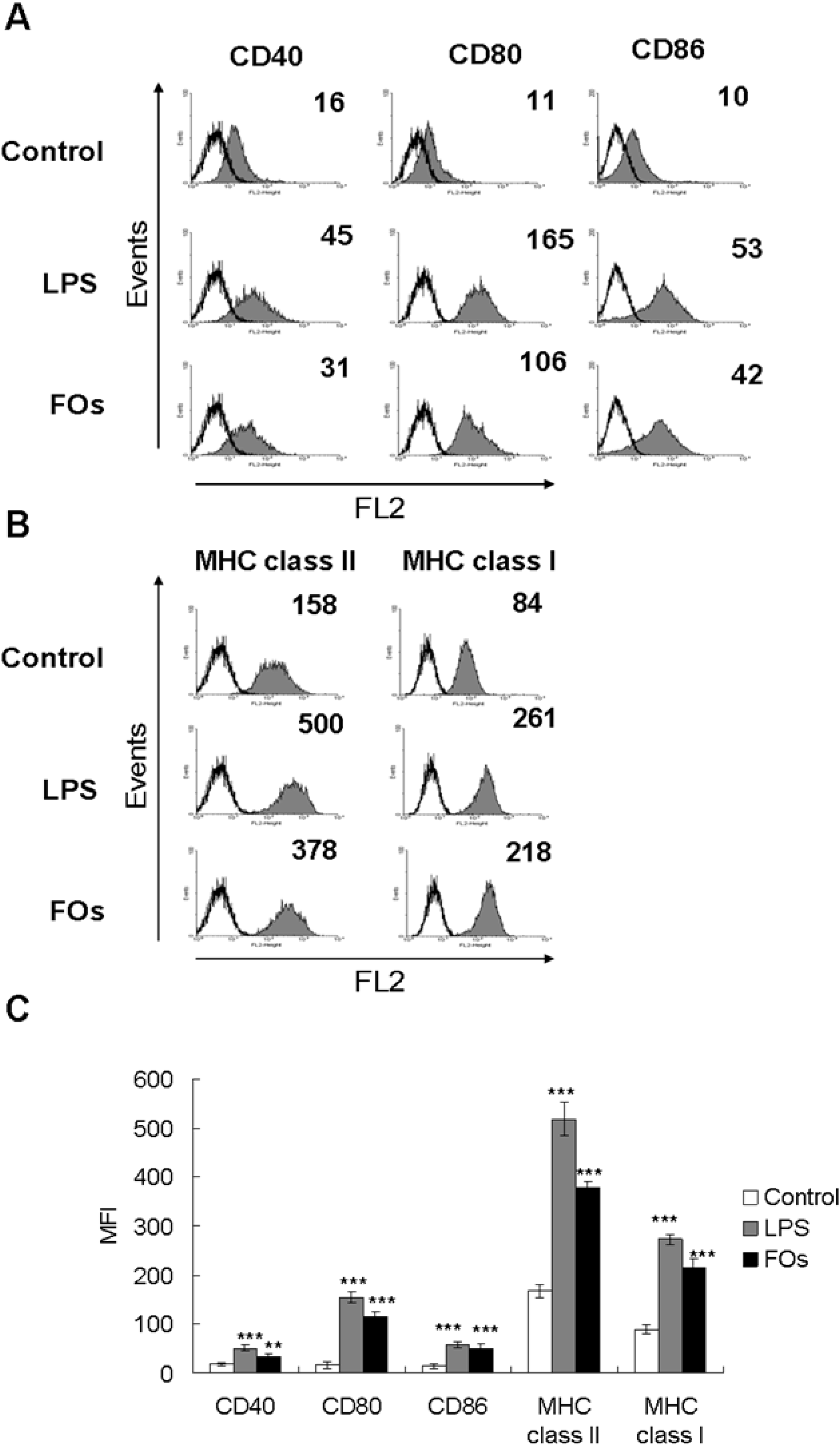
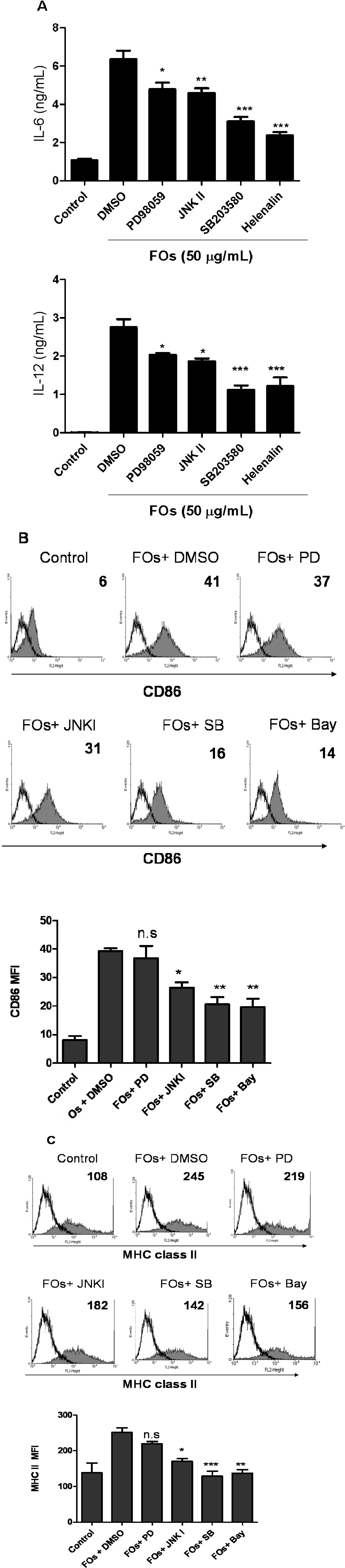
2.2. FOs Up-Regulate the Expression of Immunomodulatory Cell Surface Markers on BMDCs

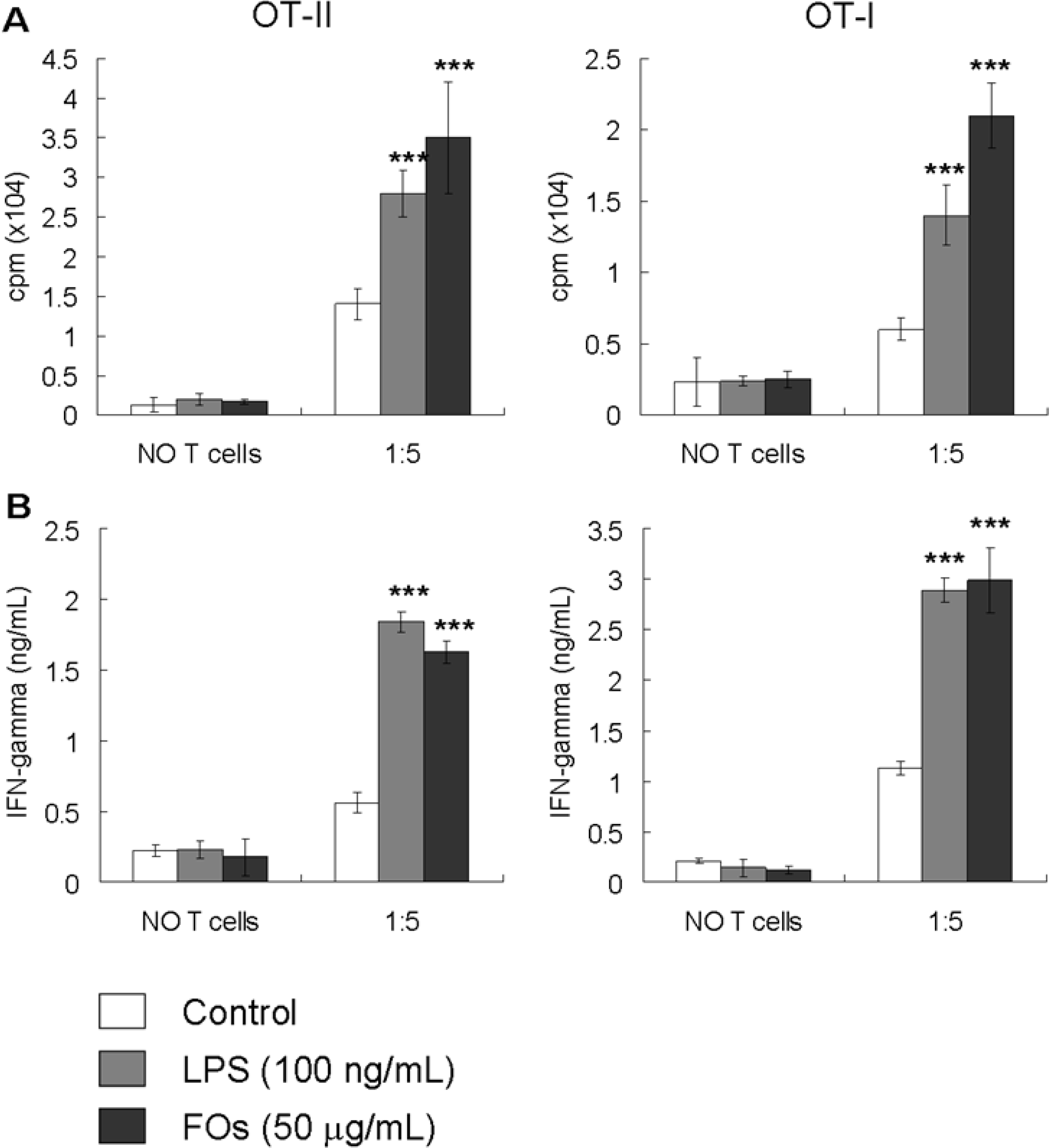
2.3. FOs Increase the Ability of BMDCs to Stimulate OVA-Specific T-cell Proliferation


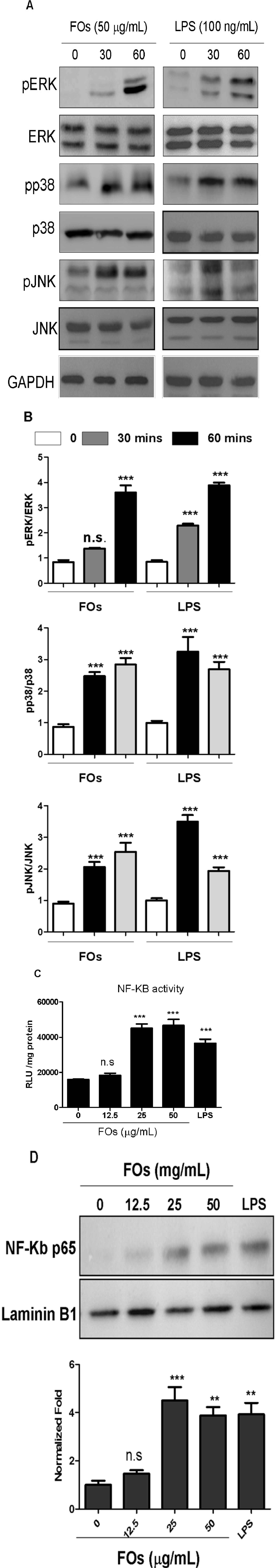
2.4. FOs is Involved with ERK, p38, JNK Kinases, and NF-κB p65 Activation


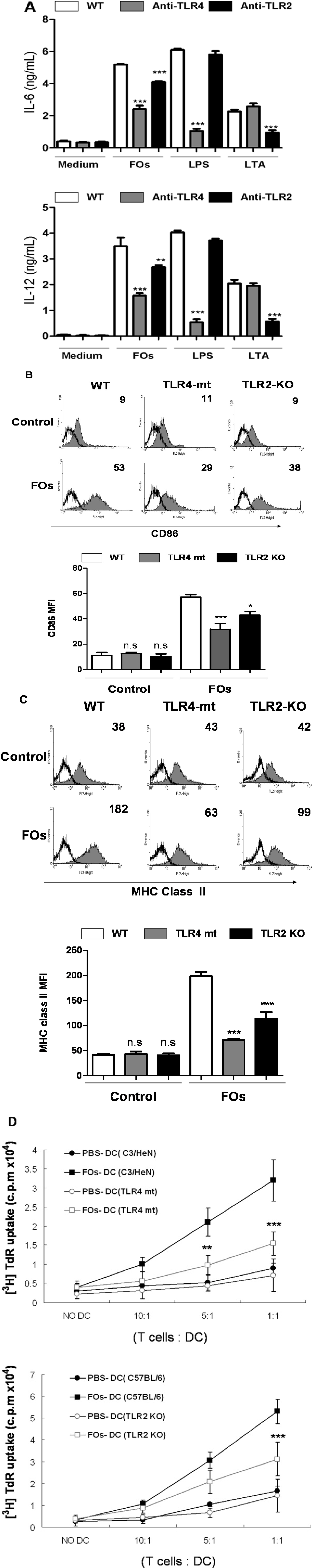
2.5. BMDCs from TLR 4/2-Deficient Mice do not Respond to FOs Stimulation

3. Discussion
4. Experimental
4.1. Mice and Cell Cultures
4.2. Feruloylated Oligosaccharides
4.3. In Vitro dcs Activation
4.4. Flow Cytometric Analysis of Surface Markers
4.5. Cytokine Detection
4.6. Neutralization Experiments
4.7. OVA-Specific T Cell Activation
4.8. Allogenic Mixed Lymphocyte Reaction
4.9. Activity Assay for MAPKs and NF-κB
4.10. Statistical Analysis
5. Conclusions
Abbreviations
| FOs | Feruloylated oligosaccharides |
| DCs | dendritic cells |
| MAPK | mitogen-activated protein kinase |
| TLR | Toll-like receptor |
| FA | ferulic acid |
| APCs | antigen-presenting cells |
| MHC | major histocompatibility complex |
| BMDCs | bone marrow-derived dendritic cells |
| ERK | extracellular signal-regulated kinase |
| JNK | c-JUN N-terminal kinase |
| NF-κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| TNF-alpha | tumor necrosis factor |
| IL | interleukin |
| TH | T helper cells |
| IFN-γ | interferon-γ |
| LPS | lipopolysaccharides |
| LTA | lipoteichoic acid |
| MFI | mean fluorescence intensity |
| OVA | ovalbumin |
| MCP-1 | monocyte chemotactic protein-1 |
| MIP-1β | macrophage inflammatory protein-1 beta |
| RANTES | regulated on activation; normal T cell expressed and secreted |
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interests
References
- Moore, J.; Cheng, Z.; Su, L.; Yu, L. Effects of solid-state enzymatic treatments on the Antioxidant properties of wheat bran. J. Agric. Food Chem. 2006, 54, 9032–9045. [Google Scholar] [CrossRef]
- Saulnier, L.; Vigouroux, J.; Thibault, J.F. Isolation and partial characterization of feruloylated oligosaccharides from maize bran. Carbohydr. Res. 1995, 272, 241–253. [Google Scholar] [CrossRef]
- Yuan, X.; Wang, J.; Yao, H.; Chen, F. Free radical-scavenging capacity and inhibitory activity on rat erythrocyte hemolysis of feruloyl oligosaccharides from wheat bran insoluble dietary fiber. LWT 2005, 38, 877–883. [Google Scholar] [CrossRef]
- Fang, H.Y.; Chen, Y.K.; Chen, H.H.; Lin, S.Y.; Fang, Y.T. Immunomodulatory effects of feruloylated oligosaccharides from rice bran. Food Chem. 2012, 134, 836–840. [Google Scholar] [CrossRef]
- Hopkins, R.A.; Connolly, J.E. The specialized roles of immature and mature dendritic cells in antigen cross-presentation. Immunol. Res. 2012, 53, 91–107. [Google Scholar] [CrossRef]
- Steinman, R.M. Decisions about dendritic cells: Past, present, and future. Annu. Rev. Immunol. 2012, 30, 1–22. [Google Scholar] [CrossRef]
- Hivroz, C.; Chemin, K.; Tourret, M.; Bohineust, A. Crosstalk between T lymphocytes and dendritic cells. Crit. Rev. Immunol. 2012, 32, 139–155. [Google Scholar] [CrossRef]
- Lanzavecchia, A.; Sallusto, F. The instructive role of dendritic cells on T cell responses: Iineages, plasticity and kinetics. Curr. Opin. Immunol. 2001, 13, 291–298. [Google Scholar] [CrossRef]
- Hanke, N.; Alizadeh, D.; Katsanis, E.; Larmonier, N. Dendritic cell tumor killing activity and its potential applications in cancer immunotherapy. Crit. Rev. Immunol. 2013, 33, 1–21. [Google Scholar] [CrossRef]
- Hubbell, J.A.; Thomas, S.N.; Swartz, M.A. Materials engineering for immunomodulation. Nature 2009, 462, 449–460. [Google Scholar] [CrossRef]
- Sallusto, F.; Lanzavecchia, A. Efficient presentation of soluble antigen by cultured human dendritic cells is maintained by granulocyte/macrophage colony-stimulating factor plus interleukin 4 and downregulated by tumor necrosis factor alpha. J. Exp. Med. 1994, 179, 1109–1118. [Google Scholar] [CrossRef]
- Lawrence, T.; Gilroy, D.W.; Colville-Nash, P.R.; Willoughby, D.A. Possible new role for NF-κB in the resolution of inflammation. Nat. Med. 2001, 7, 1291–1297. [Google Scholar] [CrossRef]
- Zhu, N.; Cui, J.; Qiao, C.; Li, Y; Ma, Y.; Zhang, J.; Shen, B. cAMP modulates macrophage development by suppressing M-CSF-induced MAPKs activation. Cell. Mol. Immunol. 2008, 5, 153–157. [Google Scholar] [CrossRef]
- Kim, G.Y.; Han, M.G.; Song, Y.S.; Shin, B.C.; Shin, Y.I.; Lee, H.J.; Moon, D.O.; Lee, C.M.; Kwak, J.Y.; Bae, Y.S.; et al. Proteoglycan isolated from Phellinus linteus induces Toll-like receptors 2- and 4-mediated maturation of murine dendritic cells via activation of ERK, p38, and NF-kappaB. Biol. Pharm. Bull. 2004, 27, 1656–1662. [Google Scholar] [CrossRef]
- Kim, H.S.; Kim, J.Y.; Kang, J.S.; Kim, H.M.; Kim, Y.O.; Hong, I.P.; Lee, M.K.; Hong, J.T.; Kim, Y.; Han, S.B. Cordlan polysaccharide isolated from mushroom Cordyceps militaris induces dendritic cell maturation through toll-like receptor 4 signalings. Food Chem. Toxicol. 2010, 48, 1926–1933. [Google Scholar] [CrossRef]
- Lu, H.; Yang, Y.; Gad, E.; Wenner, C.A.; Chang, A.; Larson, E.R.; Dang, Y.; Martzen, M.; Standish, L.J.; Disis, M.L. Polysaccharide krestin is a novel TLR2 agonist that mediates inhibition of tumor growth via stimulation of CD8 T cells and NK cells. Clin. Canc. Res. 2011, 17, 67–76. [Google Scholar] [CrossRef]
- Trinchieri, G. Interleukin-12: A cytokine at the interface of inflammation and immunity. Adv. Immunol. 1998, 70, 83–243. [Google Scholar] [CrossRef]
- Schroder, K.; Hertzog, P.J.; Ravasi, T.; Hume, D.A. Interferon-gamma: An overview of signals, mechanisms and functions. J. Leukoc. Biol. 2004, 75, 163–189. [Google Scholar]
- Sozzani, S.; Sallusto, F.; Luini, W.; Zhou, D.; Piemonti, L.; Allavena, P.; van Damme, J.; Valitutti, S.; Lanzavecchia, A.; Mantovani, A. Migration of dendritic cells in response to formyl peptides, C5a, and a distinct set of chemokines. J. Immunol. 1995, 155, 3292–3295. [Google Scholar]
- Collart, M.A.; Baeuerle, P.; Vassalli, P. Regulation of tumor necrosis factor alpha transcription in macrophages: Involvement of four kappa B-like motifs and of constitutive and inducible forms of NF-kappa B. Cell. Mol. Biol. 1990, 10, 1498–1506. [Google Scholar]
- Libermann, T.A.; Baltimore, D. Activation of interleukin-6 gene expression through the NF-kappa B transcription factor. Cell. Mol. Biol. 1990, 10, 2327–2334. [Google Scholar]
- Murphy, T.L.; Cleveland, M.G.; Kulesza, P.; Magram, J.; Murphy, K.M. Regulation of interleukin 12 p40 expression through an NF-kappa B half-site. Cell. Mol. Biol. 1995, 15, 5258–5267. [Google Scholar]
- Hinz, M.; Löser, P.; Mathas, S.; Krappmann, D.; Dörken, B.; Scheidereit, C. Constitutive NF-êB maintains high expression of a characteristic gene network, including CD40, CD86, and a set of antiapoptotic genes in Hodgkin/Reed-Sternberg cells. Blood 2001, 97, 2798–2807. [Google Scholar] [CrossRef]
- Rescigno, M.; Martino, M.; Sutherland, C.L.; Gold, M.R.; Ricciardi-Castagnoli, P. Dendritic cell survival and maturation are regulated by different signaling pathways. J. Exp. Med. 1998, 188, 2175–2180. [Google Scholar] [CrossRef]
- Bode, K.A.; Schmitz, F.; Vargas, L.; Heeg, K.; Dalpke, A.H. Kinetic of RelA activation controls magnitude of TLR-mediated IL-12 p40 induction. J. Immunol. 2009, 182, 2176–2184. [Google Scholar] [CrossRef]
- Jones, B.W.; Heldwein, K.A.; Means, T.K.; Saukkonen, J.J.; Fenton, M.J. Differential roles of Toll-like receptors in the elicitation of proinflammatory responses by macrophages. Ann. Rheum. Dis. 2001, 60, 6–12. [Google Scholar] [CrossRef]
- Ozinsky, A.; Underhill, D.M.; Fontenot, J.D.; Hajjar, A.M.; Smith, K.D.; Wilson, C.B.; Schroeder, L.; Aderem, A. The repertoire for pattern recognition of pathogens by the innate immune system is defined by cooperation between toll-like receptors. Proc. Natl. Acad. Sci. USA 2000, 97, 13766–13771. [Google Scholar]
- Ninomiya-Tsuji, J.; Kishimoto, K.; Hiyama, A.; Inoue, J.; Cao, Z.; Matsumoto, K. The kinase TAK1 can activate the NIK-I kappaB as well as the MAP kinase cascade in the IL-1 signalling pathway. Nature 1999, 398, 252–256. [Google Scholar] [CrossRef]
- Netea, M.G.; van Deuren, M.; Kullberg, B.J.; Cavaillon, J.-M.; van der Meer, J.W.M. Does the shape of lipid A determine the interaction of LPS with Toll-like receptors? Trends Immunol. 2002, 23, 135–139. [Google Scholar]
- Takeda, K.; Kaisho, T.; Akira, S. Toll-like receptors. Annu. Rev. Immunol. 2003, 21, 335–376. [Google Scholar] [CrossRef]
- Weinstein, S.L.; June, C.H.; DeFranco, A.L. Lipopolysaccharide-induced protein tyrosine phosphorylation in human macrophages is mediated by CD14. J. Immunol. 1993, 151, 3829–3838. [Google Scholar]
- Chen, M.H.; Li, W.S.; Lue, Y.S.; Chu, C.L; Pan, I.H.; Ko, C.H.; Chen, D.Y.; Lin, C.H.; Lin, S.H.; Chang, C.P.; et al. Clitocybe nuda activates dendritic cells and acts as a DNA vaccine adjuvant. Evid. Base Compl. Alternative Med. 2013, 2013, 761454. [Google Scholar]
- Li, K.Y.; Lai, P.; Lu, S.; Fang, Y.T.; Chen, H.H. Optimization of acid hydrolysis conditions for feruloylated oligosaccharides from rice bran through response surface methodolgy. J. Agric. Food Chem. 2008, 56, 8975–8978. [Google Scholar] [CrossRef]
- Lin, C.C.; Chu, C.L.; Ng, C.S.; Lin, C.Y.; Chen, D.Y.; Pan, I.H.; Huang, K.J. Immunomodulation of phloretin by impairing dendritic cell activation and function. Food Funct. 2014. [Google Scholar] [CrossRef]
- Asea, A.; Kraeft, S.K.; Kurt-Jones, E.A.; Stevenson, M.A.; Chen, L.B.; Finberg, R.W.; Koo, G.C.; Calderwood, S.K. HSP70 stimulates cytokine production through a CD14-dependant pathway, demonstrating itsdual role as a chaperone and cytokine. Nat. Med. 2000, 6, 435–442. [Google Scholar] [CrossRef]
- Bausinger, H.; Lipsker, D.Z.; Iylan, U.; Manié, S.; Briand, J.P.; Cazenave, J.P.; Muller, S.; Haeuw, J.F.; Ravanat, C.; de la Salle, H.; et al. Endotoxin-free heat-shock protein fails to induce APC activation. Eur. J. Immunol. 2002, 32, 3708–3713. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds rice bran feruloylated oligosaccharides are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lin, C.C.; Chen, H.H.; Chen, Y.K.; Chang, H.C.; Lin, P.Y.; Pan, I.-H.; Chen, D.-Y.; Chen, C.M.; Lin, S.Y. Rice Bran Feruloylated Oligosaccharides Activate Dendritic Cells via Toll-Like Receptor 2 and 4 Signaling. Molecules 2014, 19, 5325-5347. https://doi.org/10.3390/molecules19045325
Lin CC, Chen HH, Chen YK, Chang HC, Lin PY, Pan I-H, Chen D-Y, Chen CM, Lin SY. Rice Bran Feruloylated Oligosaccharides Activate Dendritic Cells via Toll-Like Receptor 2 and 4 Signaling. Molecules. 2014; 19(4):5325-5347. https://doi.org/10.3390/molecules19045325
Chicago/Turabian StyleLin, Chi Chen, Hua Han Chen, Yu Kuo Chen, Hung Chia Chang, Ping Yi Lin, I-Hong Pan, Der-Yuan Chen, Chuan Mu Chen, and Su Yi Lin. 2014. "Rice Bran Feruloylated Oligosaccharides Activate Dendritic Cells via Toll-Like Receptor 2 and 4 Signaling" Molecules 19, no. 4: 5325-5347. https://doi.org/10.3390/molecules19045325