Mulberry Fruit Extract Protects Pancreatic β-Cells against Hydrogen Peroxide-Induced Apoptosis via Antioxidative Activity

 and
and 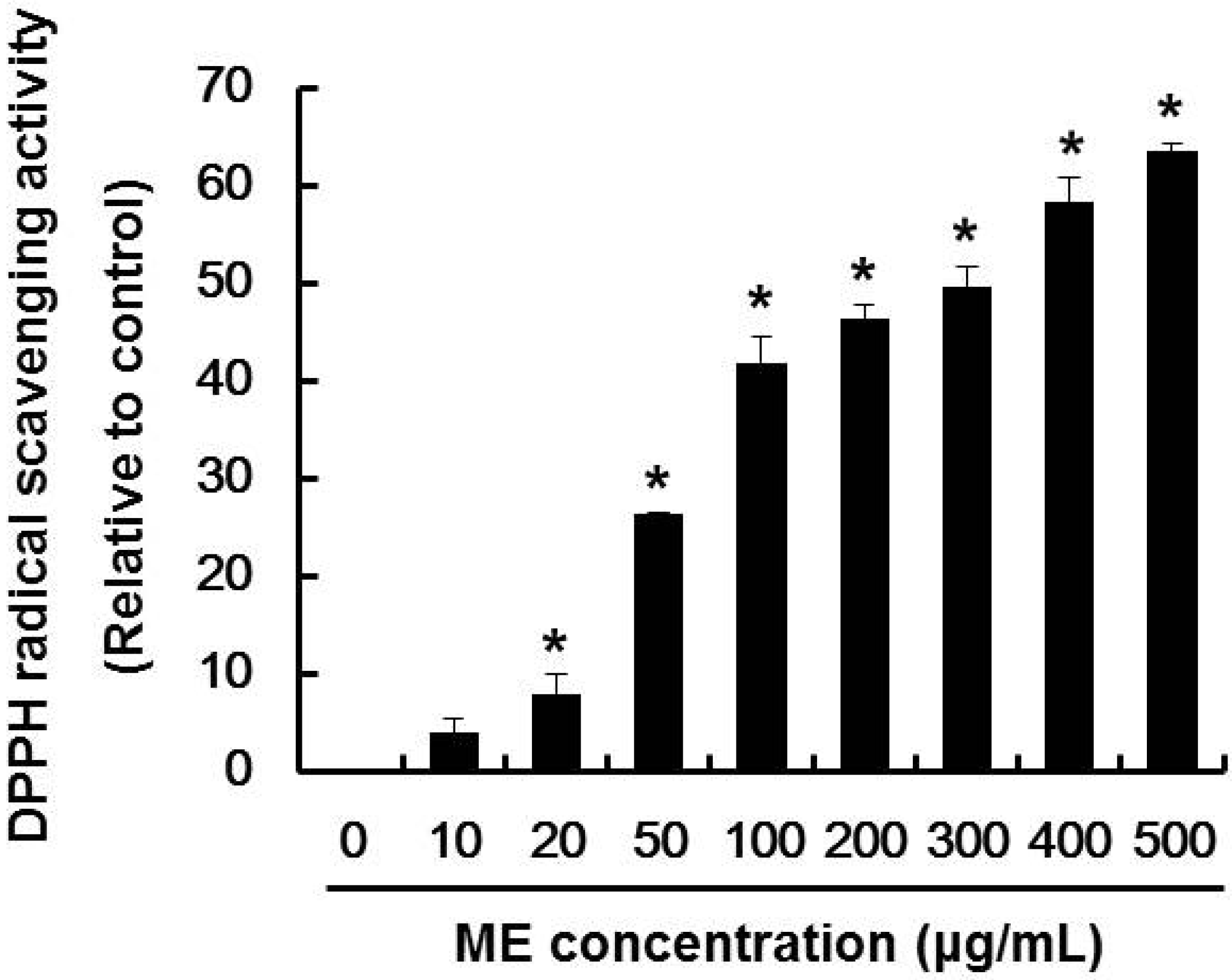{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. DPPH Radical Scavenging Activity of ME

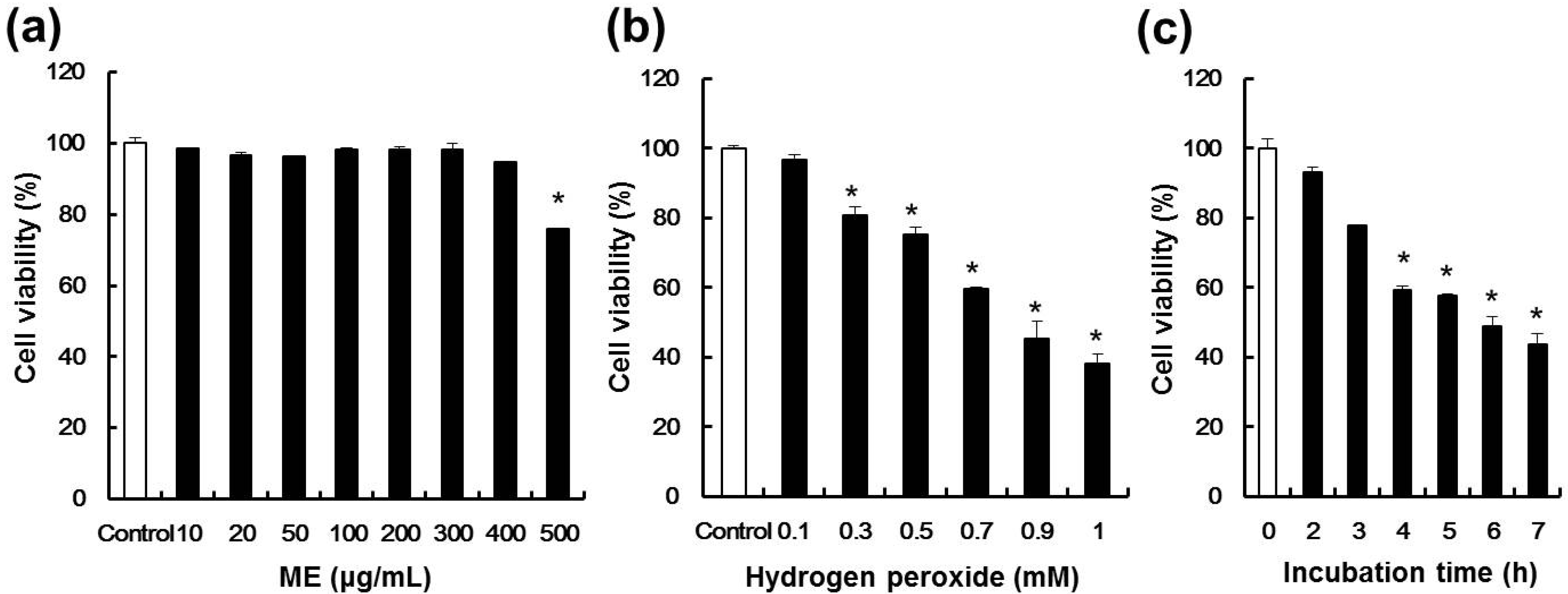
2.2. Effect of ME and Hydrogen Peroxide on Cell Viability of Pancreatic β-cells

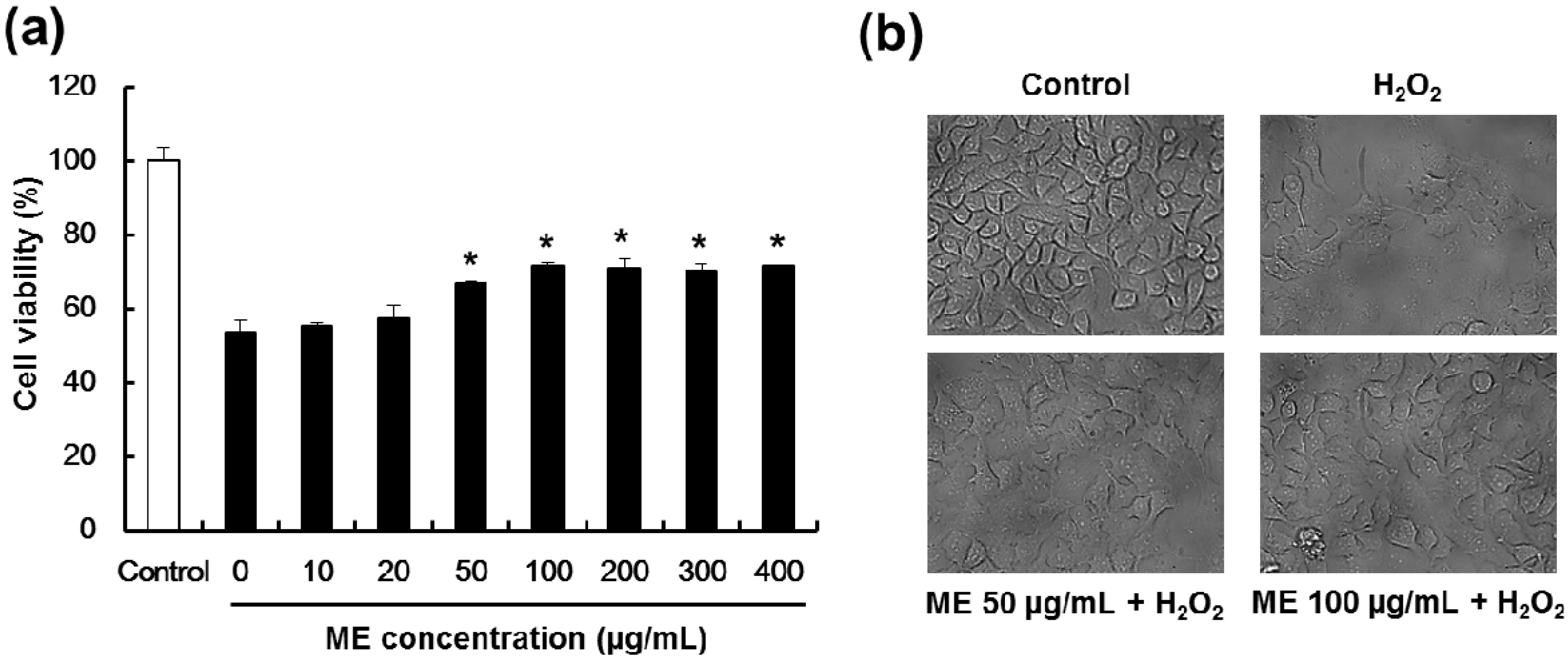
2.3. Mulberry Extract Confers Protection Pancreatic β-Cells Against Hydrogen Peroxide-Induced Toxicity

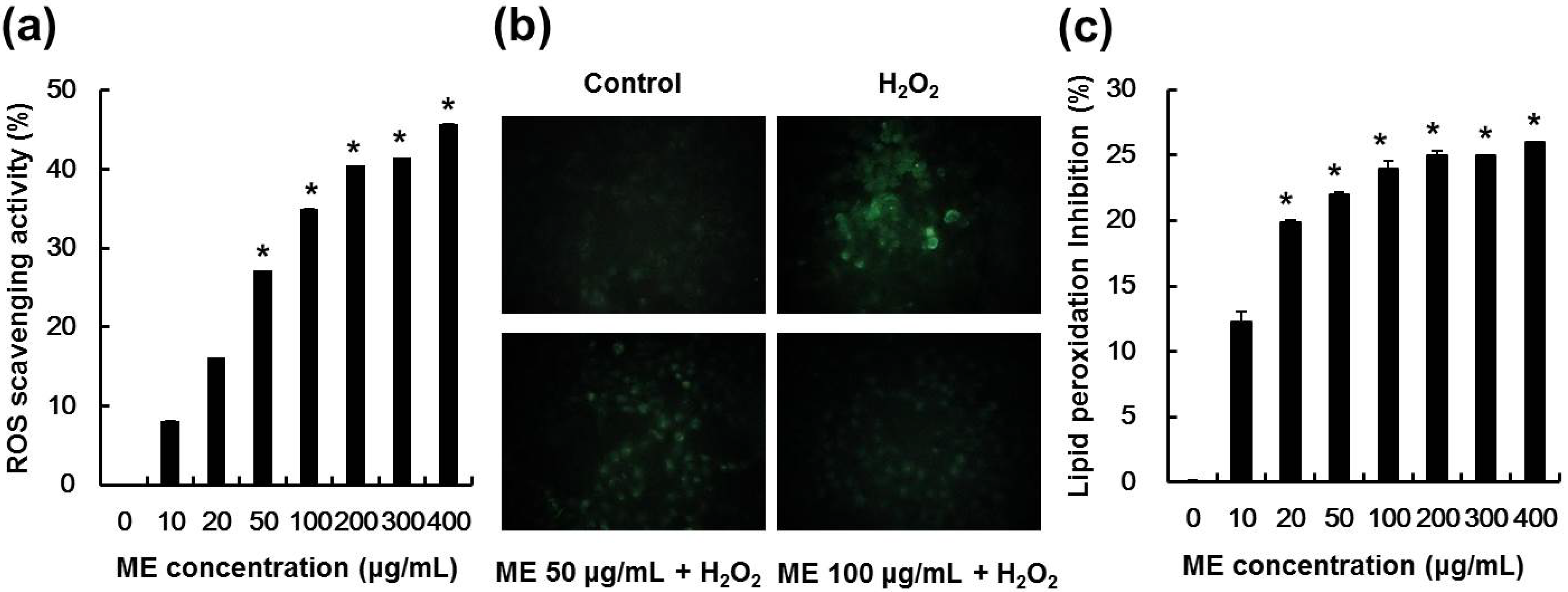
2.4. ME Inhibited Hydrogen Peroxide-Induced ROS Generation and Lipid Peroxidation

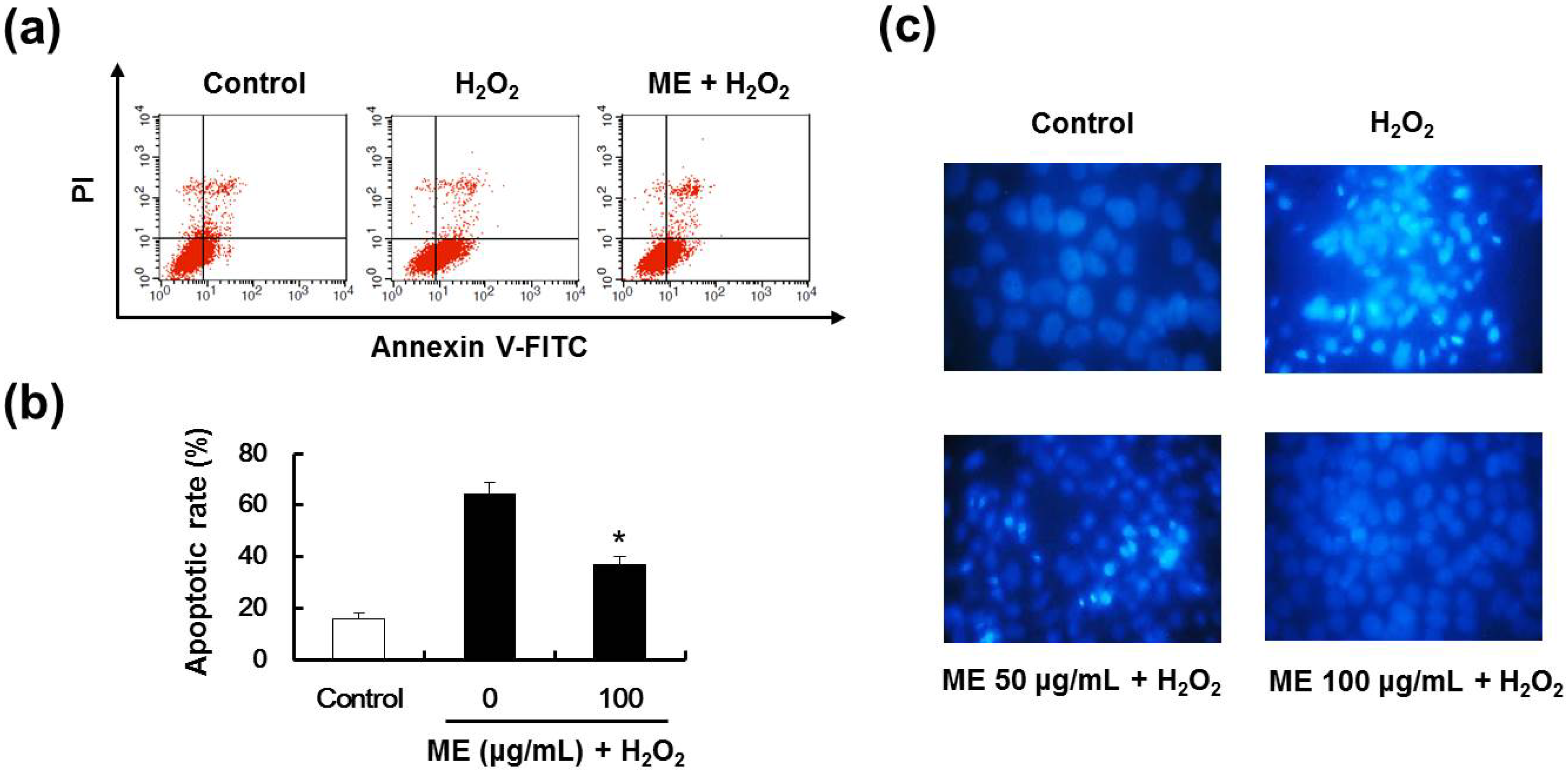
2.5. ME Inhibited Hydrogen Peroxide-Induced Apoptotic Cell Death

3. Experimental
3.1. Materials
3.2. Cell Culture
3.3. DPPH Radical Scavenging Activity
3.4. Cytotoxicity of ME
3.5. Protective Effect of ME and Image Analysis
3.6. Intracellular ROS Scavenging Activity and Image Analysis
3.7. Lipid Peroxidation Inhibitory Activity
3.8. Flow Cytometric Analysis
3.9. Measurement of DNA Condensation and/or Fragmentation
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Maritim, A.C.; Sanders, R.A.; Watkins, J.B., 3rd. Diabetes, oxidative stress, and antioxidants: A review. J. Biochem. Mol. Toxicol. 2003, 17, 24–38. [Google Scholar] [CrossRef]
- Bach, J.F. Insulin-dependent diabetes mellitus as an autoimmune disease. Endocr. Rev. 1994, 15, 516–542. [Google Scholar] [CrossRef]
- Giugliano, D.; Ceriello, A.; Paolisso, G. Oxidative stress and diabetic vascular complications. Diabetes Care 1996, 19, 257–267. [Google Scholar]
- Leal, E.C.; Aveleira, C.A.; Castilho, A.F.; Serra, A.M.; Baptista, F.I.; Hosoya, K.; Forrester, J.V.; Ambrosio, A.F. High glucose and oxidative/nitrosative stress conditions induce apoptosis in retinal endothelial cells by a caspase independent pathway. Exp. Eye Res. 2009, 88, 983–991. [Google Scholar] [CrossRef]
- Lizard, G.; Fournel, S.; Genestier, L.; Dhedin, N.; Chaput, C.; Flacher, M.; Mutin, M.; Panaye, G.; Revillard, J.P. Kinetics of plasma membrane and mitochondrial alterations in cells undergoing apoptosis. Cytometry 1995, 21, 275–283. [Google Scholar]
- Lee, J.H.; Lee, J.S.; Kim, Y.R.; Jung, W.C.; Lee, K.E.; Lee, S.Y.; Hong, E.K. Hispidin isolated from Phellinus linteus protects against hydrogen peroxide-induced oxidative stress in pancreatic MIN6N beta-cells. J. Med. Food 2011, 14, 1431–1438. [Google Scholar] [CrossRef]
- Diplock, A.T.; Charleux, J.L.; Crozier-Willi, G.; Kok, F.J.; Rice-Evans, C.; Roberfroid, M.; Stahl, W.; Vina-Ribes, J. Functional food science and defence against reactive oxidative species. Br. J. Nutr. 1998, 80, S77–S112. [Google Scholar] [CrossRef]
- Reliene, R.; Schiestl, R.H. Antioxidant N-acetyl cysteine reduces incidence and multiplicity of lymphoma in Atm deficient mice. DNA Repair (Amst) 2006, 5, 852–859. [Google Scholar] [CrossRef]
- Sagrista, M.L.; Garcia, A.E.; Africa de Madariaga, M.; Mora, M. Antioxidant and pro-oxidant effect of the thiolic compounds N-acetyl-L-cysteine and glutathione against free radical-induced lipid peroxidation. Free Radic. Res. 2002, 36, 329–340. [Google Scholar]
- Tapsell, L.C.; Hemphill, I.; Cobiac, L.; Patch, C.S.; Sullivan, D.R.; Fenech, M.; Roodenrys, S.; Keogh, J.B.; Clifton, P.M.; Williams, P.G.; et al. Health benefits of herbs and spices: The past, the present, the future. Med. J. Aust. 2006, 185, S4–S24. [Google Scholar]
- Shri, J. Ginger: It’s role in xenobiotic metabolism. ICMR Bull. 2003, 33, 57–63. [Google Scholar]
- Kennedy, D.O.; Wightman, E.L. Herbal extracts and phytochemicals: Plant secondary metabolites and the enhancement of human brain function. Adv. Nutr. 2011, 2, 32–50. [Google Scholar]
- Ma, Y.P. Clinical observation of mulberry as a medicine of pharyngitis treatment. Xinjiang J. Tradit. Chin. Med. 2002, 20, 83–84. [Google Scholar]
- Hwang, K.H.; Kim, Y.K. Promoting effect and recovery activity from physical stress of the fruit of Morus alba. Biofactors 2004, 21, 267–271. [Google Scholar]
- Kaewkaen, P.; Tong-Un, T.; Wattanathorn, J.; Muchimapura, S.; Kaewrueng, W.; Wongcharoenwanakit, S. Mulberry fruit extract protects against memory impairment and hippocampal damage in animal model of vascular dementia. Evid.-Based Compl. Alt. Med. 2012, 2012, 1–9. [Google Scholar]
- Kwon, H.J.; Chung, J.Y.; Kim, J.Y.; Kwon, O. Comparison of 1-deoxynojirimycin and aqueous mulberry leaf extract with emphasis on postprandial hypoglycemic effects: In vivo and in vitro studies. J. Agric. Food Chem. 2011, 59, 3014–3019. [Google Scholar]
- Jaruchotikamol, A.; Pannangpetch, P. Cytoprotective activity of mulberry leaf extract against oxidative stress-induced cellular injury in rats. Pak. J. Pharm. Sci. 2013, 26, 163–168. [Google Scholar]
- Kim, J.Y.; Kwon, H.J.; Jung, J.Y.; Kwon, H.Y.; Baek, J.G.; Kim, Y.S.; Kwon, O. Comparison of absorption of 1-deoxynojirimycin from mulberry water extract in rats. J. Agric. Food Chem. 2010, 58, 6666–6671. [Google Scholar] [CrossRef]
- Miyahara, C.; Miyazawa, M.; Satoh, S.; Sakai, A.; Mizusaki, S. Inhibitory effects of mulberry leaf extract on postprandial hyperglycemia in normal rats. J. Nutr. Sci. Vitaminol. (Tokyo) 2004, 50, 161–164. [Google Scholar] [CrossRef]
- Fang, F.; Luo, M.L.; Su, N.; Wu, X.R. Effect of mulberry leaves extracts on glucose uptake of insulin-resistant HepG2 cells and the mechanism. Yao Xue Xue Bao 2012, 47, 1452–1456. [Google Scholar]
- Wang, Y.; Xiang, L.; Wang, C.; Tang, C.; He, X. Antidiabetic and antioxidant effects and phytochemicals of mulberry fruit (Morus alba L.) polyphenol enhanced extract. PLoS One 2013, 8, e71144. [Google Scholar]
- Ha, U.S.; Bae, W.J.; Kim, S.J.; Yoon, B.I.; Jang, H.; Hong, S.H.; Lee, J.Y.; Hwang, S.Y.; Kim, S.W. Protective effect of cyanidin-3-O-β-d-glucopyranoside fraction from mulberry fruit pigment against oxidative damage in streptozotocin-induced diabetic rat bladder. Neurourol. Urodynam. 2013, 32, 493–499. [Google Scholar]
- Valacchi, G.; Belmonte, G.; Miracco, C.; Eo, H.; Lim, Y. Effect of combined mulberry leaf and fruit extract on liver and skin cholesterol transporters in high fat diet-induced obese mice. Nutr. Res. Pract. 2014, 8, 20–26. [Google Scholar]
- Lee, J.S.; Synytsya, A.; Kim, H.B.; Choi, D.J.; Lee, S.; Lee, J.; Kim, W.J.; Jang, S.; Park, Y.I. Purification, characterization and immunomodulating activity of a pectic polysaccharide isolated from Korean mulberry fruit Oddi (Morus alba L.). Int. Immunopharmacol. 2013, 17, 858–866. [Google Scholar] [CrossRef]
- Rolo, A.P.; Palmeira, C.M. Diabetes and mitochondrial function: Role of hyperglycemia and oxidative stress. Toxicol. Appl. Pharm. 2006, 212, 167–178. [Google Scholar] [CrossRef]
- Zhong, J.; Rao, X.; Xu, J.F.; Yang, P.; Wang, C.Y. The role of endoplasmic reticulum stress in autoimmune-mediated beta-cell destruction in type 1 diabetes. Exp. Diabetes Res. 2012, 2012, 1–12. [Google Scholar]
- Dalton, T.P.; Shertzer, H.G.; Puga, A. Regulation of gene expression by reactive oxygen. Annu. Rev. Pharmacol. 1999, 39, 67–101. [Google Scholar] [CrossRef]
- Evans, J.L.; Goldfine, I.D.; Maddux, B.A.; Grodsky, G.M. Are oxidative stress-activated signaling pathways mediators of insulin resistance and beta-cell dysfunction? Diabetes 2003, 52, 1–8. [Google Scholar]
- Darzynkiewicz, Z.; Juan, G.; Li, X.; Gorczyca, W.; Murakami, T.; Traganos, F. Cytometry in cell necrobiology: Analysis of apoptosis and accidental cell death (necrosis). Cytometry 1997, 27, 1–20. [Google Scholar] [CrossRef]
- Blois, M.S. Free Radicals in Biological Systems. Science 1960, 132, 306–307. [Google Scholar]
- Jang, J.S.; Lee, J.S.; Lee, J.H.; Kwon, D.S.; Lee, K.E.; Lee, S.Y.; Hong, E.K. Hispidin produced from Phellinus linteus protects pancreatic beta-cells from damage by hydrogen peroxide. Arch. Pharm. Res. 2010, 33, 853–861. [Google Scholar]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Kang, K.A.; Kim, J.S.; Zhang, R.; Piao, M.J.; Maeng, Y.H.; Kang, M.Y.; Lee, I.K.; Kim, B.J.; Hyun, J.W. KIOM-4 protects against oxidative stress-induced mitochondrial damage in pancreatic beta-cells via its antioxidant effects. Evid. Based Complement. Alternat. Med. 2011, 2011, 1–11. [Google Scholar]
- Gschwind, M.; Huber, G. Apoptotic cell death induced by beta-amyloid 1-42 peptide is cell type dependent. J. Neurochem. 1995, 65, 292–300. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.S.; Kim, Y.R.; Park, J.M.; Ha, S.-J.; Kim, Y.E.; Baek, N.I.; Hong, E.K. Mulberry Fruit Extract Protects Pancreatic β-Cells against Hydrogen Peroxide-Induced Apoptosis via Antioxidative Activity. Molecules 2014, 19, 8904-8915. https://doi.org/10.3390/molecules19078904
Lee JS, Kim YR, Park JM, Ha S-J, Kim YE, Baek NI, Hong EK. Mulberry Fruit Extract Protects Pancreatic β-Cells against Hydrogen Peroxide-Induced Apoptosis via Antioxidative Activity. Molecules. 2014; 19(7):8904-8915. https://doi.org/10.3390/molecules19078904
Chicago/Turabian StyleLee, Jong Seok, Young Rae Kim, Jun Myoung Park, Suk-Jin Ha, Young Eon Kim, Nam In Baek, and Eock Kee Hong. 2014. "Mulberry Fruit Extract Protects Pancreatic β-Cells against Hydrogen Peroxide-Induced Apoptosis via Antioxidative Activity" Molecules 19, no. 7: 8904-8915. https://doi.org/10.3390/molecules19078904