GA3 and Other Signal Regulators (MeJA and IAA) Improve Xanthumin Biosynthesis in Different Manners in Xanthium strumarium L.

Abstract
:1. Introduction
2. Results and Discussion
2.1. The Discrepancy between Gland Formation and Xanthumin Production upon Flowering Initiation

{kind=link}
{kind=link}
{kind=link}
| Groups | Mean | SD | df | t | Sig. (p) | |
|---|---|---|---|---|---|---|
| Trichome density (No./mm2) | 80 days | 4254.90 | 90.15 | 2 | ||
| 100 days | 4029.82 | 315.16 | 2 | −1.7325 | >0.05 | |
| Cross-sectional area (µm2) | 80 days | 2305.15 | 140.75 | 2 | ||
| 100 days | 2166.85 | 158.53 | 2 | −13.4710 | >0.05 | |
| Relative xanthumin level | 80 days | 1.00 | 0.13 | 2 | ||
| 100 days | 1.36 | 0.01 | 2 | −9.2520 | <0.01 ** | |
| GA3 concentration (ng/g fresh weight) | 80 days | 0.37 | 0.04 | 2 | ||
| 100 days | 0.69 | 0.08 | 2 | 13.6749 | <0.05 * |
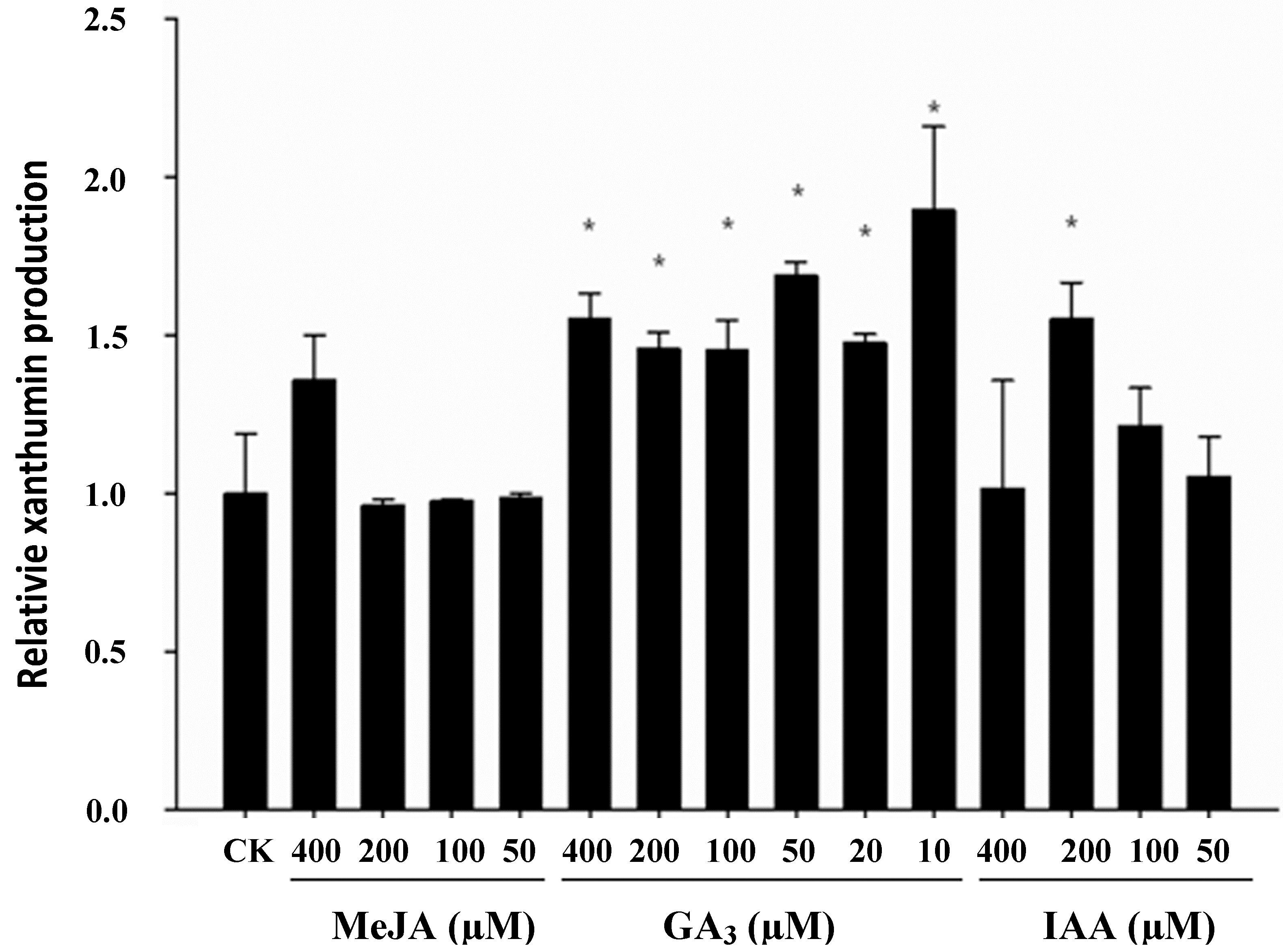
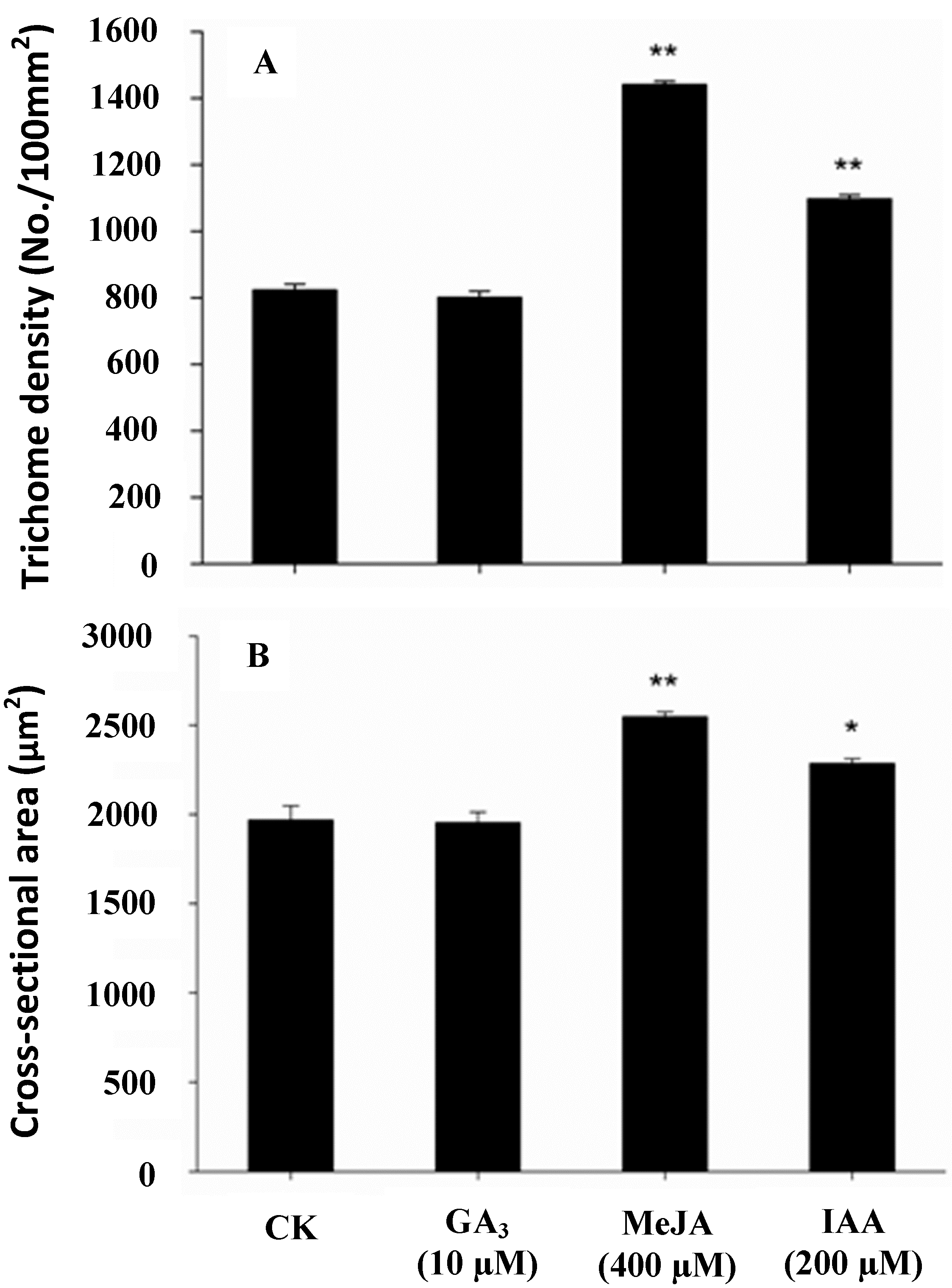
2.2. Effect of Phytohormonal Stimuli on Glandular Trichome Density and Xanthumin Production


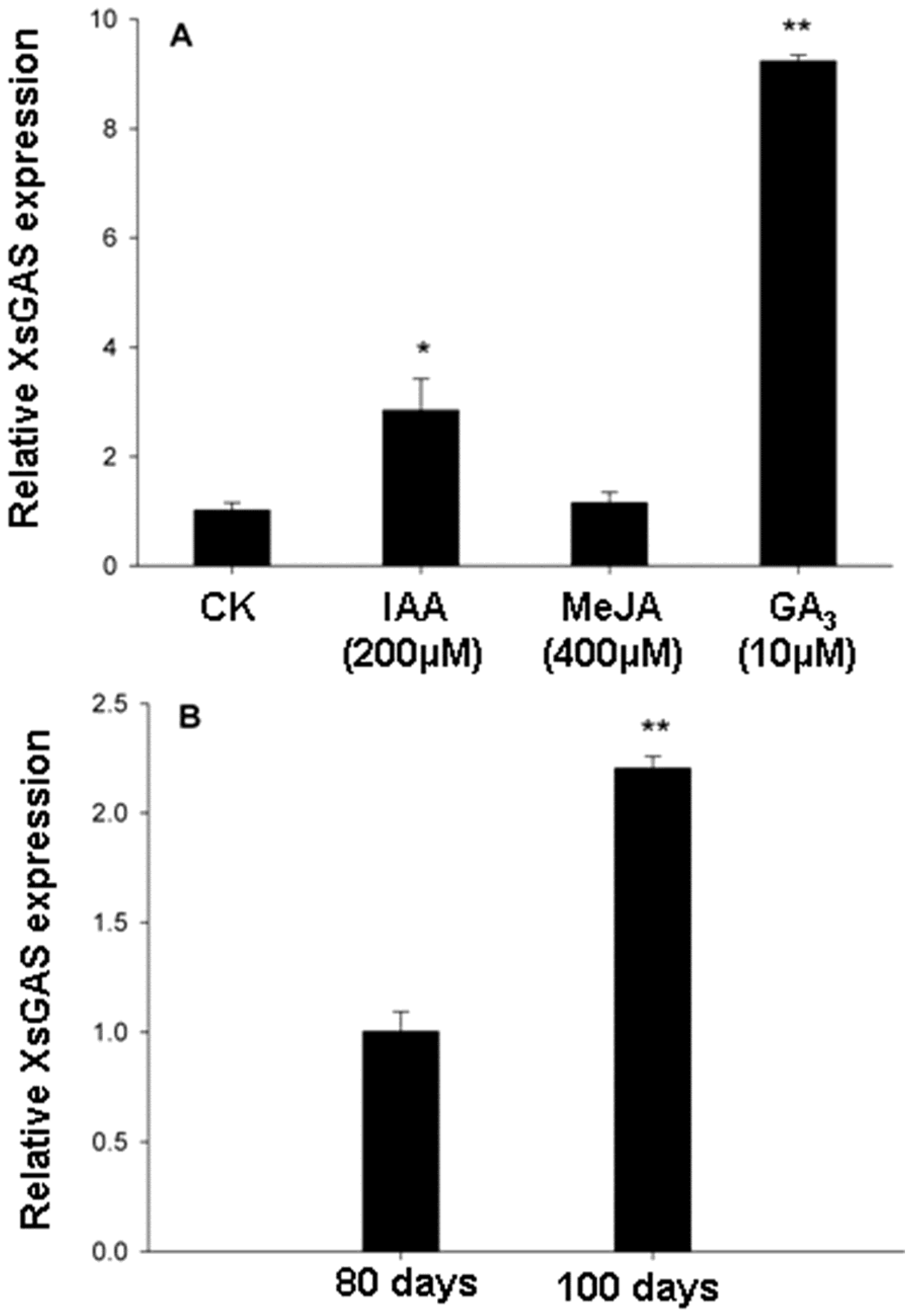
2.3. The Increased Expression of Germacrene A Synthase Gene Induced by GA3 Treatment

3. Experimental Section
3.1. Chemicals and Plant Materials
3.2. Flowering Induction and Phytohormonal Elicitation
3.3. Chemical Extraction and HPLC Analysis
3.4. Glandular Trichome Analysis
3.5. XsGAS Transcript Analysis
4. Conclusions
Supplementary Materials
Supplementary File
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zhang, X.Q.; Ye, W.C.; Jiang, R.W.; Yin, Z.Q.; Zhao, S.X.; Mak, T.C.; Yao, X.S. Two new eremophilanolides from Xanthium sibiricum. Nat. Prod. Res. 2006, 20, 1265–1270. [Google Scholar] [CrossRef]
- Kim, H.S.; Lee, T.S.; Yeo, S.W.; Seong, L.S.; Yu, T.S. Isolation and characterization of antitumor agents from Xanthium strumarium L. Korean J. Biotechnol. Bioeng. 2003, 18, 324–328. [Google Scholar]
- Ramirez-Erosa, I.; Huang, Y.; Hickie, R.A.; Sutherland, R.G.; Barl, B. Xanthatin and xanthinosin from the burs of Xanthium strumarium L. as potential anticancer agent. Can. J. Physiol. Pharmacol. 2007, 85, 1160–1172. [Google Scholar] [CrossRef]
- Kim, Y.S.; Kim, J.S.; Park, S.H.; Choi, S.U.; Lee, C.O.; Kim, S.K.; Kim, Y.K.; Kim, S.H.; Ryu, S.Y. Two cytotoxic sesquiterpene lactones from the leaves of Xanthium strumarium and their in vitro inhibitory activity on farnesyltransferase. Plant Med. 2003, 69, 375–377. [Google Scholar] [CrossRef]
- Chen, F.F.; Hao, F.H.; Li, C.F.; Gou, J.B.; Lu, D.Y.; Gong, F.J.; Tang, H.R.; Zhang, Y.S. Identifying three ecological chemotypes of Xanthium strumarium L. glandular trichomes using a combined NMR and LC-MS method. PLoS One 2013, 8, e76621. [Google Scholar]
- Yokotani-Tomita, K.; Kato, J.; Kosemura, S.; Yamamura, S.; Kushima, M.; Kakuta, H.; Hasegawa, K. Light-induced auxin-inhibiting substance from sunflower seedlings. Phytochemistry 1997, 46, 503–506. [Google Scholar] [CrossRef]
- Raupp, F.M.; Spring, O. New Sesquiterpene Lactones from Sunflower Root Exudate as Germination Stimulants for Orobanche cumana. J. Agric. Food Chem. 2013, 61, 10481–10487. [Google Scholar] [CrossRef]
- Yang, C.; Yuan, C.; Jia, Z. Xanthanolides, germacranolides, and other constituents from Carpesium longifolium. J. Nat. Prod. 2003, 66, 1554–1557. [Google Scholar] [CrossRef]
- De Kraker, J.W.; Franssen, M.C.; de Groot, A.; Konig, W.A.; Bouwmeester, H.J. (+)-Germacrene A biosynthesis. The committed step in the biosynthesis of bitter sesquiterpene lactones in chicory. Plant Physiol. 1998, 117, 1381–1392. [Google Scholar] [CrossRef]
- Nguyen, D.T.; Gopfert, J.C.; Ikezawa, N.; Macnevin, G.; Kathiresan, M.; Conrad, J.; Spring, O.; Ro, D.K. Biochemical conservation and evolution of germacrene A oxidase in asteraceae. J. Biol. Chem. 2010, 285, 16588–16598. [Google Scholar] [CrossRef]
- Bertea, C.M.; Voster, A.; Verstappen, F.W.; Maffei, M.; Beekwilder, J.; Bouwmeester, H.J. Isoprenoid biosynthesis in Artemisia annua: Cloning and heterologous expression of a germacrene A synthase from a glandular trichome cDNA library. Arch. Biochem. Biophys. 2006, 448, 3–12. [Google Scholar] [CrossRef]
- Majdi, M.; Liu, Q.; Karimzadeh, G.; Malboobi, M.A.; Beekwilder, J.; Cankar, K.; Vos, R.; Todorovic, S.; Simonovic, A.; Bouwmeester, H. Biosynthesis and localization of parthenolide in glandular trichomes of feverfew (Tanacetum parthenium L. Schulz Bip). Phytochemistry 2011, 72, 1739–1750. [Google Scholar] [CrossRef]
- Matsuo, K.; Ohtsuki, K.; Yoshikawa, T.; Shishido, K.; Yokotani-Tomita, K.; Shindo, M. Total synthesis of xanthanolides. Tetrahedron 2010, 66, 8407–8419. [Google Scholar] [CrossRef]
- Hong, G.J.; Xue, X.Y.; Mao, Y.B.; Wang, L.J.; Chen, X.Y. Arabidopsis thaliana MYC2 interacts with DELLA proteins in regulating sesquiterpene synthase gene expression. Plant Cell 2012, 24, 2635–2648. [Google Scholar] [CrossRef]
- Mannan, A.; Ahmed, I.; Arshad, W.; Hussain, I.; Mirza, B. Effects of vegetative and flowering stages on the biosynthesis of artemisinin in Artemisia species. Arch. Pharm. Res. 2011, 34, 1657–1661. [Google Scholar] [CrossRef]
- Richards, D.E.; King, K.E.; Ait-ali, T.; Harberd, N.P. How gibberellin regulates plant growth and development: A molecular genetic analysis of gibberellin signaling. Annu. Rev. Plant Biol. 2001, 52, 67–88. [Google Scholar] [CrossRef]
- Zhou, Z.J.; An, L.J.; Sun, L.L.; Zhu, S.J.; Xi, W.Y.; Broun, P.; Yu, H.; Gan, Y.B. Zinc Finger Protein5 Is Required for the Control of Trichome Initiation by Acting Upstream of Zinc Finger Protein8 in Arabidopsis thaliana. Plant Physiol. 2011, 157, 673–682. [Google Scholar] [CrossRef]
- Rowe, H.C.; Ro, D.K.; Rieseberg, L.H. Response of sunflower (Helianthus annuus L.) leaf surface defenses to exogenous methyl jasmonate. PLoS One 2012, 7, e37191. [Google Scholar] [CrossRef]
- Aerts, R.J.; Alarco, A.M.; de Luca, V. IAAs induce tryptophan decarboxylase activity in radicles of catharanthus seedlings. Plant Physiol. 1992, 100, 1014–1019. [Google Scholar] [CrossRef]
- Maes, L.; van Nieuwerburgh, F.C.; Zhang, Y.; Reed, D.W.; Pollier, J.; vande Casteele, S.R.; Inze, D.; Covello, P.S.; Deforce, D.L.; Goossens, A. Dissection of the phytohormonal regulation of trichome formation and biosynthesis of the antimalarial compound artemisinin in Artemisia annua plants. New Phytol. 2011, 189, 176–189. [Google Scholar]
- Bovy, A.; de Vos, R.; Kemper, M.; Schijlen, E.; Almenar Pertejo, M.; Muir, S.; Collins, G.; Robinson, S.; Verhoeyen, M.; Hughes, S.; et al. High-flavonol tomatoes resulting from the heterologous expression of the maize transcription factor genes LC and C1. Plant Cell 2002, 14, 2509–2526. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the xanthumin and other xanthanolides are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, C.; Chen, F.; Zhang, Y. GA3 and Other Signal Regulators (MeJA and IAA) Improve Xanthumin Biosynthesis in Different Manners in Xanthium strumarium L. Molecules 2014, 19, 12898-12908. https://doi.org/10.3390/molecules190912898
Li C, Chen F, Zhang Y. GA3 and Other Signal Regulators (MeJA and IAA) Improve Xanthumin Biosynthesis in Different Manners in Xanthium strumarium L. Molecules. 2014; 19(9):12898-12908. https://doi.org/10.3390/molecules190912898
Chicago/Turabian StyleLi, Changfu, Fangfang Chen, and Yansheng Zhang. 2014. "GA3 and Other Signal Regulators (MeJA and IAA) Improve Xanthumin Biosynthesis in Different Manners in Xanthium strumarium L." Molecules 19, no. 9: 12898-12908. https://doi.org/10.3390/molecules190912898