
Autodisplay of Human Hyaluronidase Hyal-1 on Escherichia coli and Identification of Plant-Derived Enzyme Inhibitors



Abstract
:1. Introduction
Hyaluronan (HA) and Human Hyaluronidase Hyal-1 (Hyal-1)
2. Results and Discussion
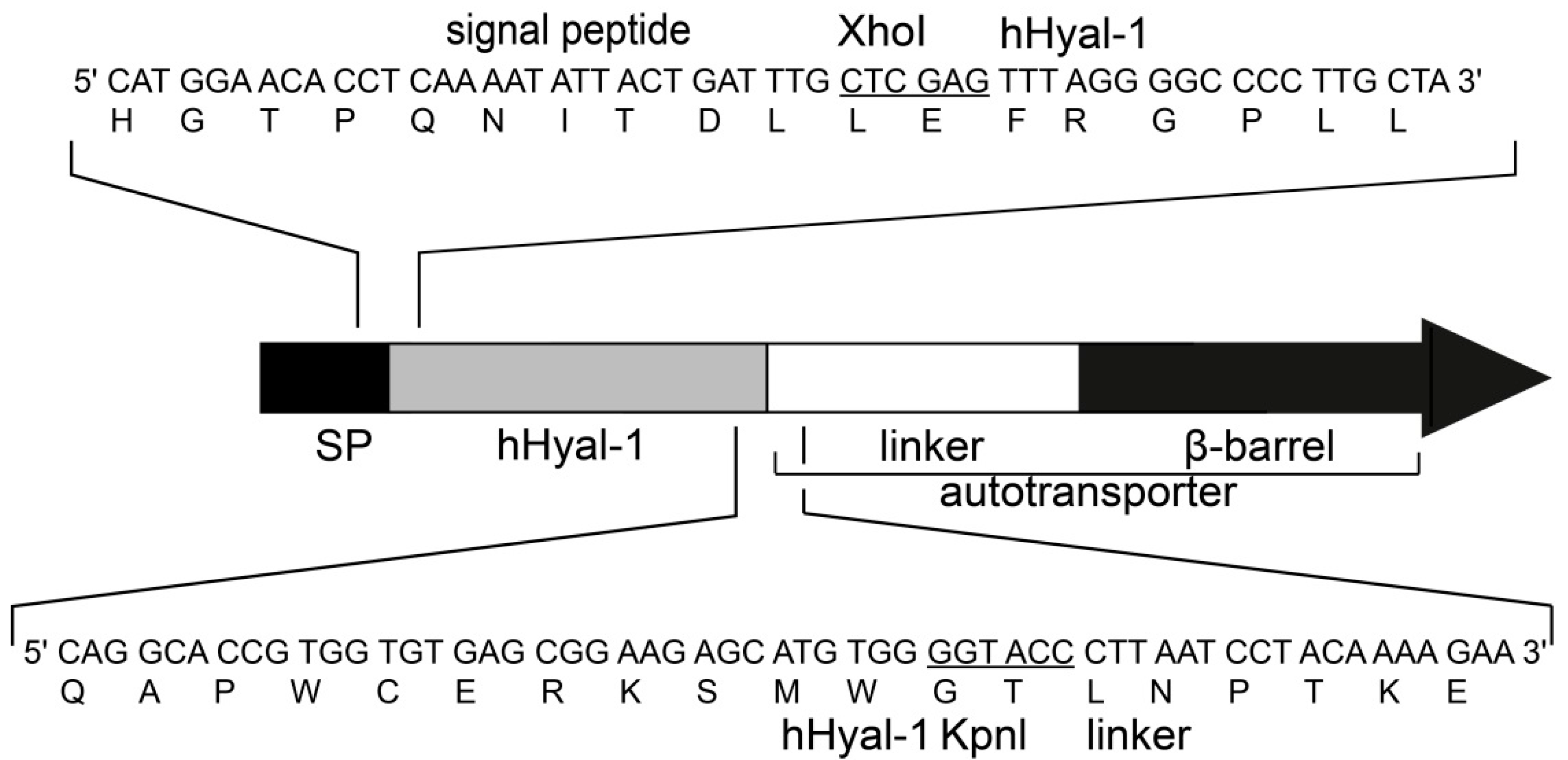
2.1. Artificial Gene Construction for the Surface Display of Hyal-1

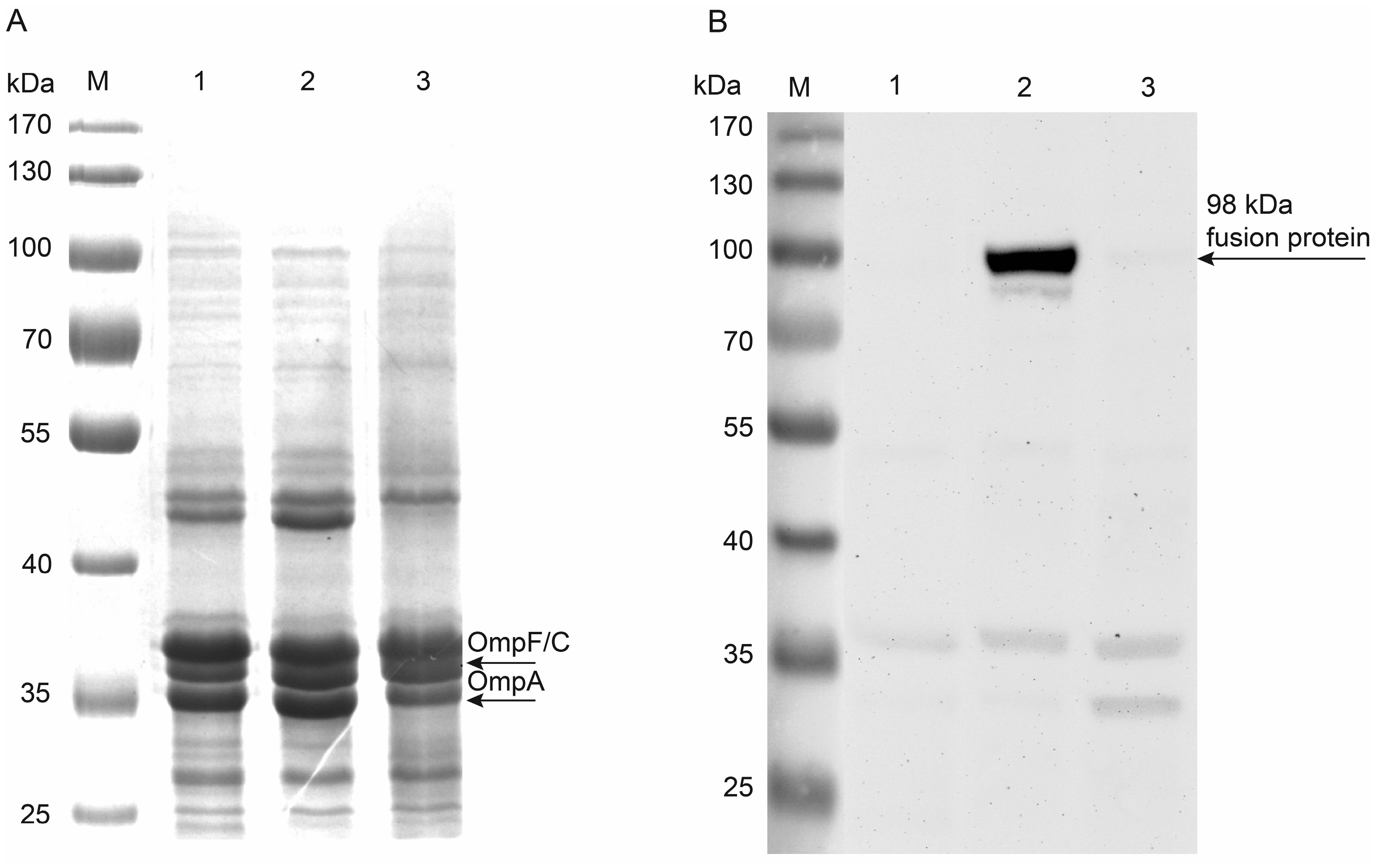
2.2. Surface Display of Hyal-10
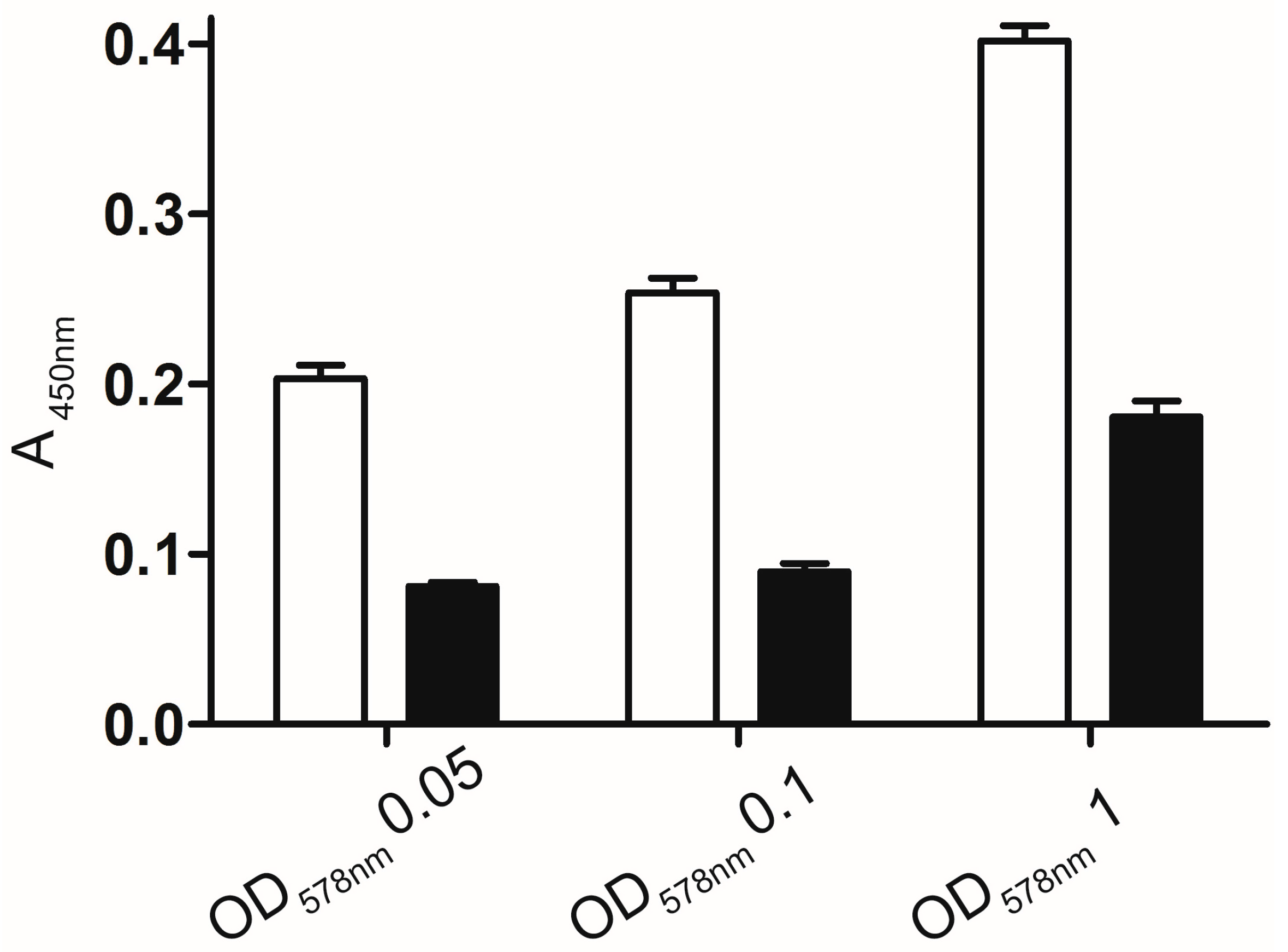
2.2.1. Whole Cell Enzyme Linked Immunosorbent Assay (ELISA)
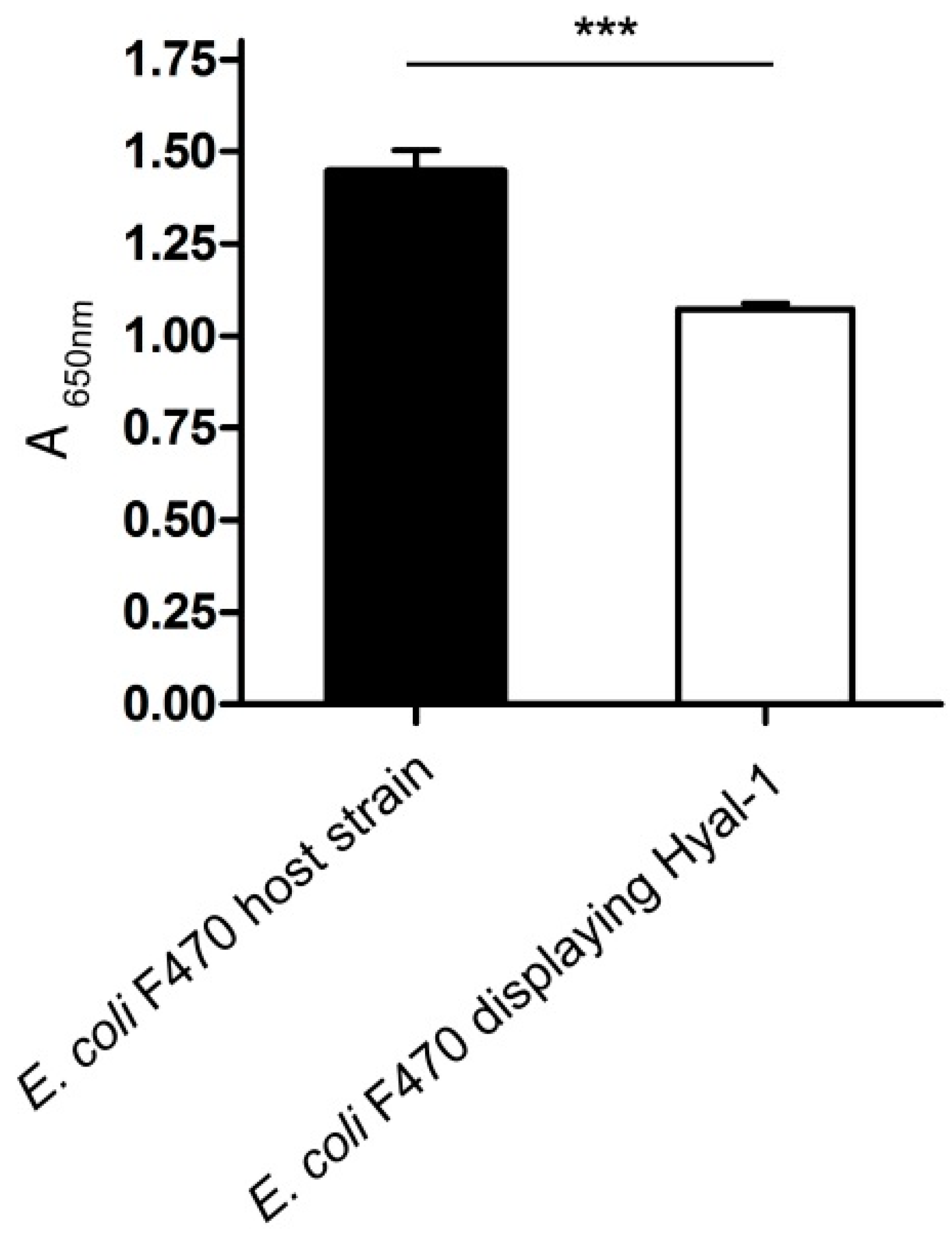
2.2.2. Protease Accessibility Test


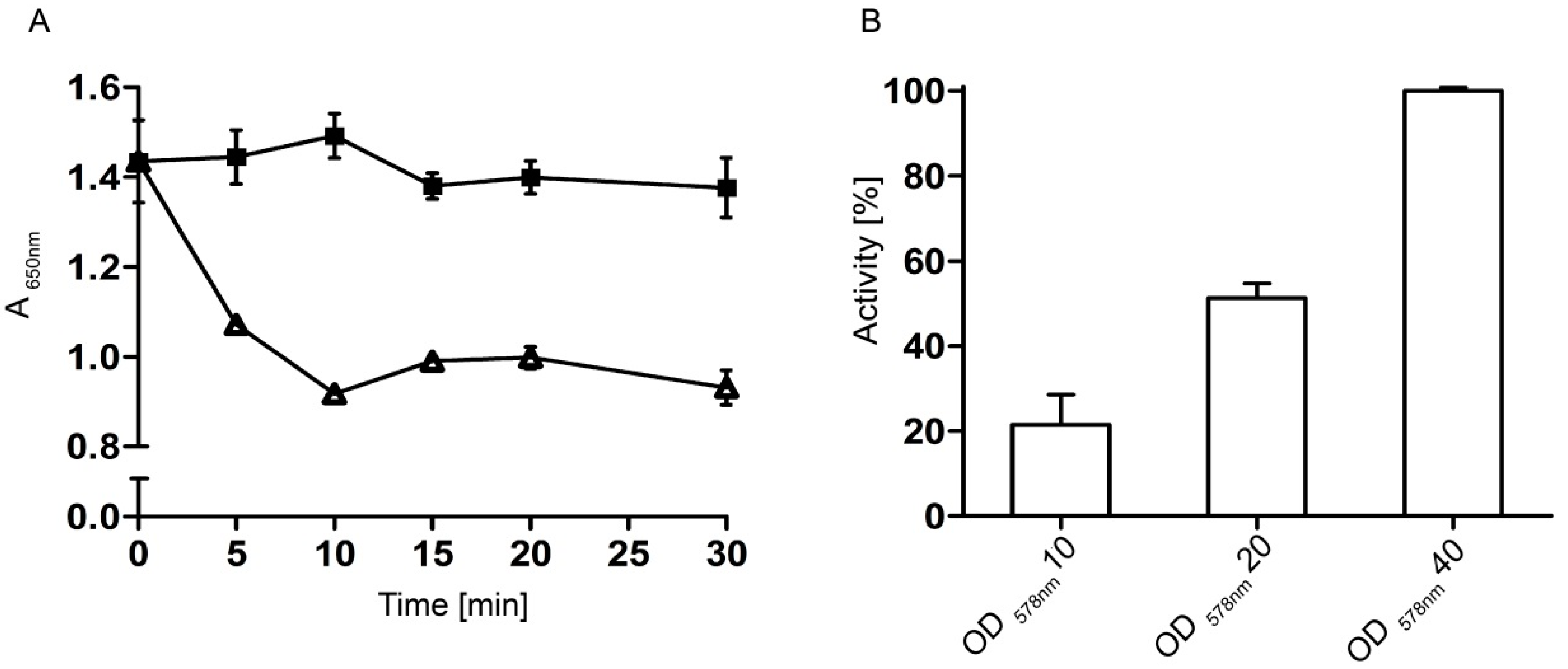
2.3. Hyaluronidase Activity of Whole Cells Displaying Hyal-1
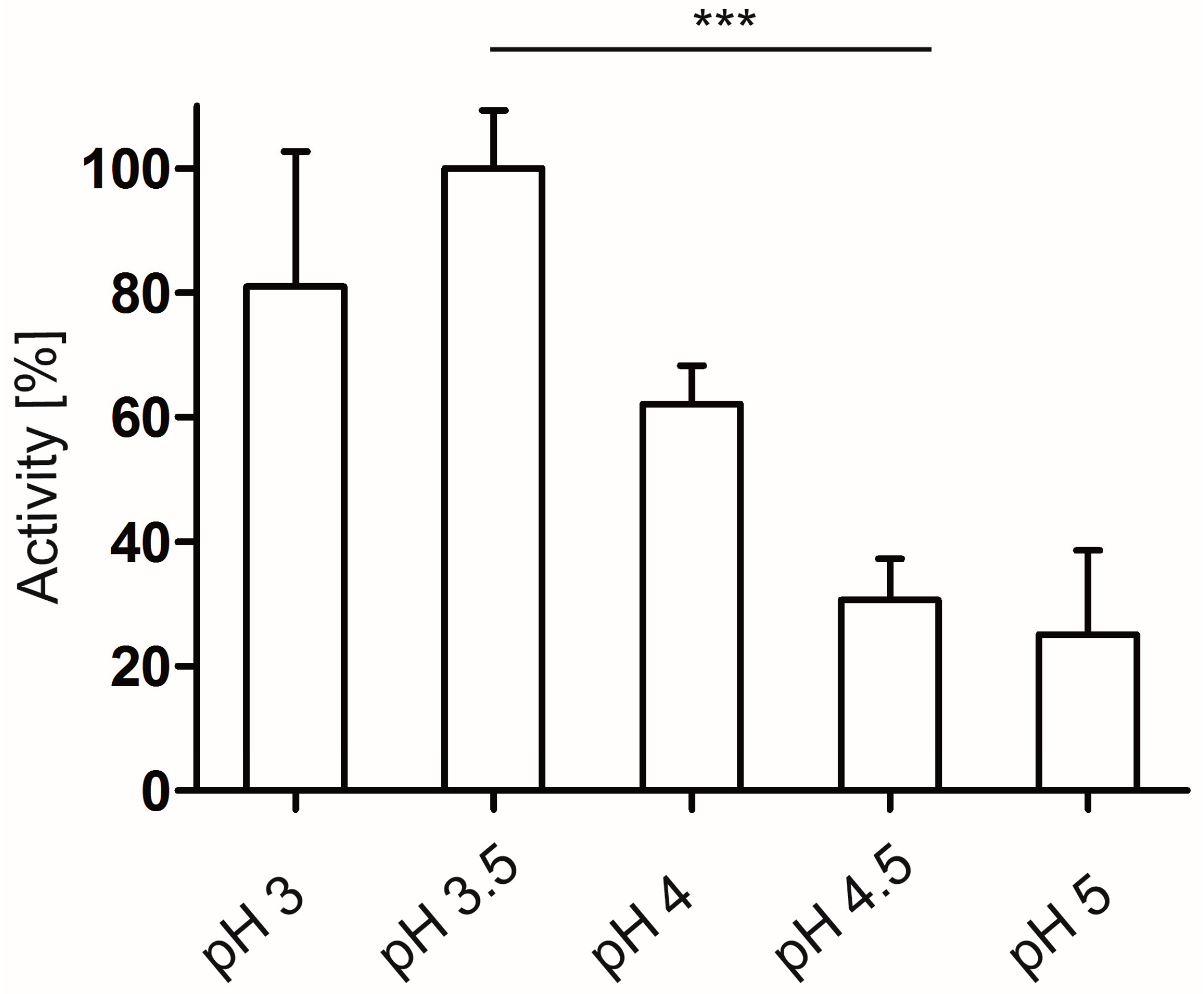
2.3.1. Enzyme Activity Determination and pH Dependency


2.3.2. Optimal Time and Cell Density

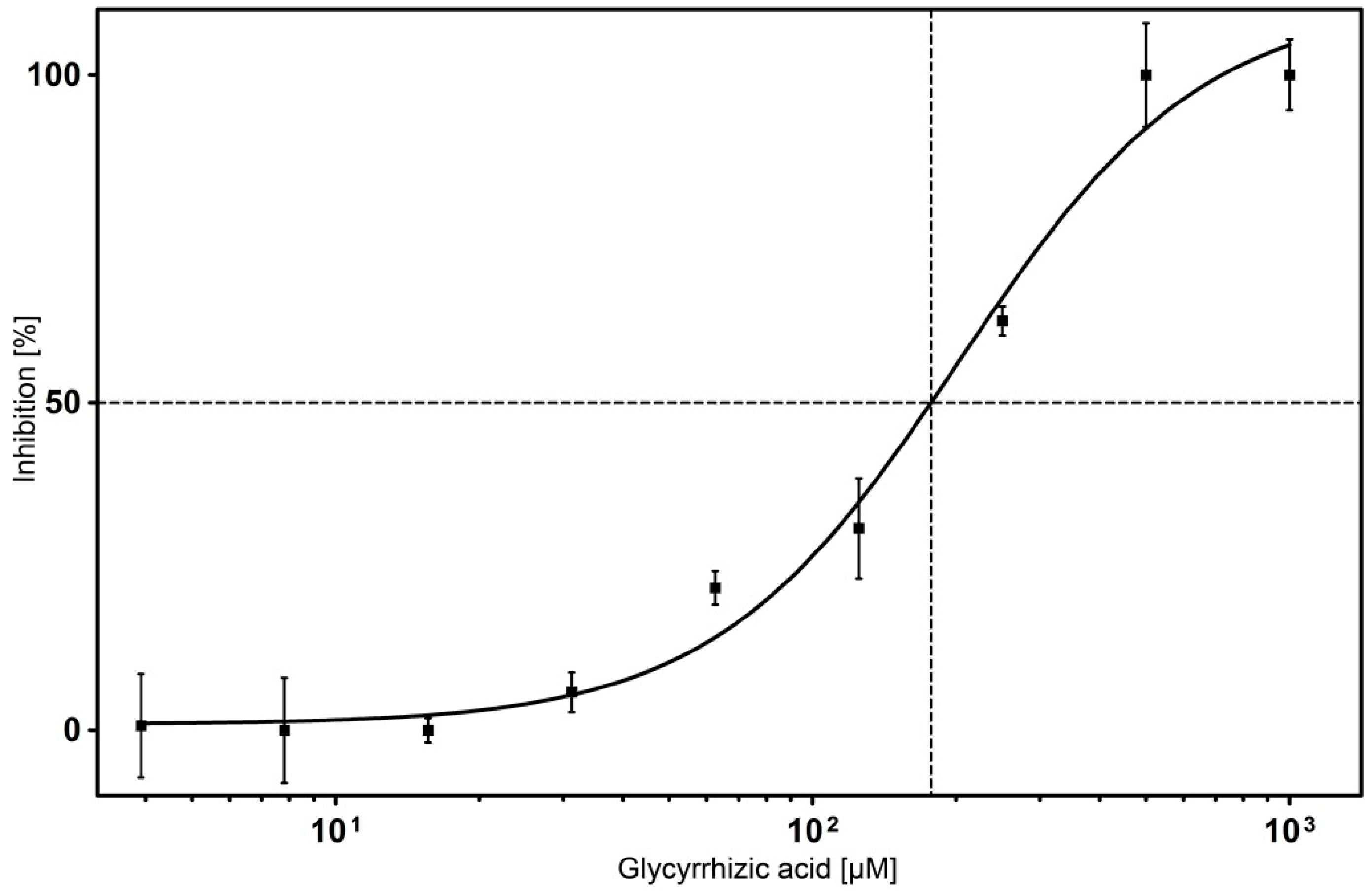
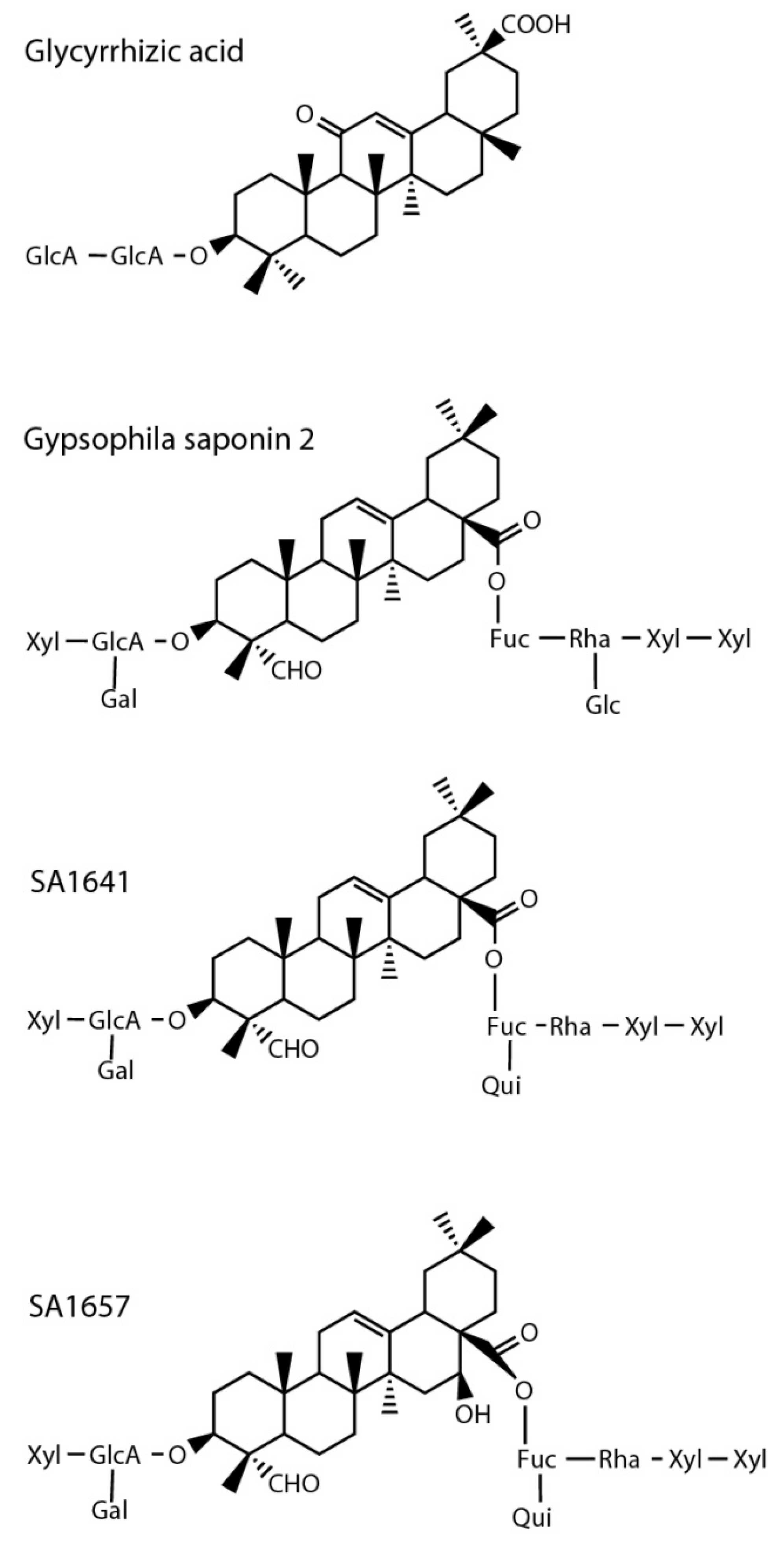
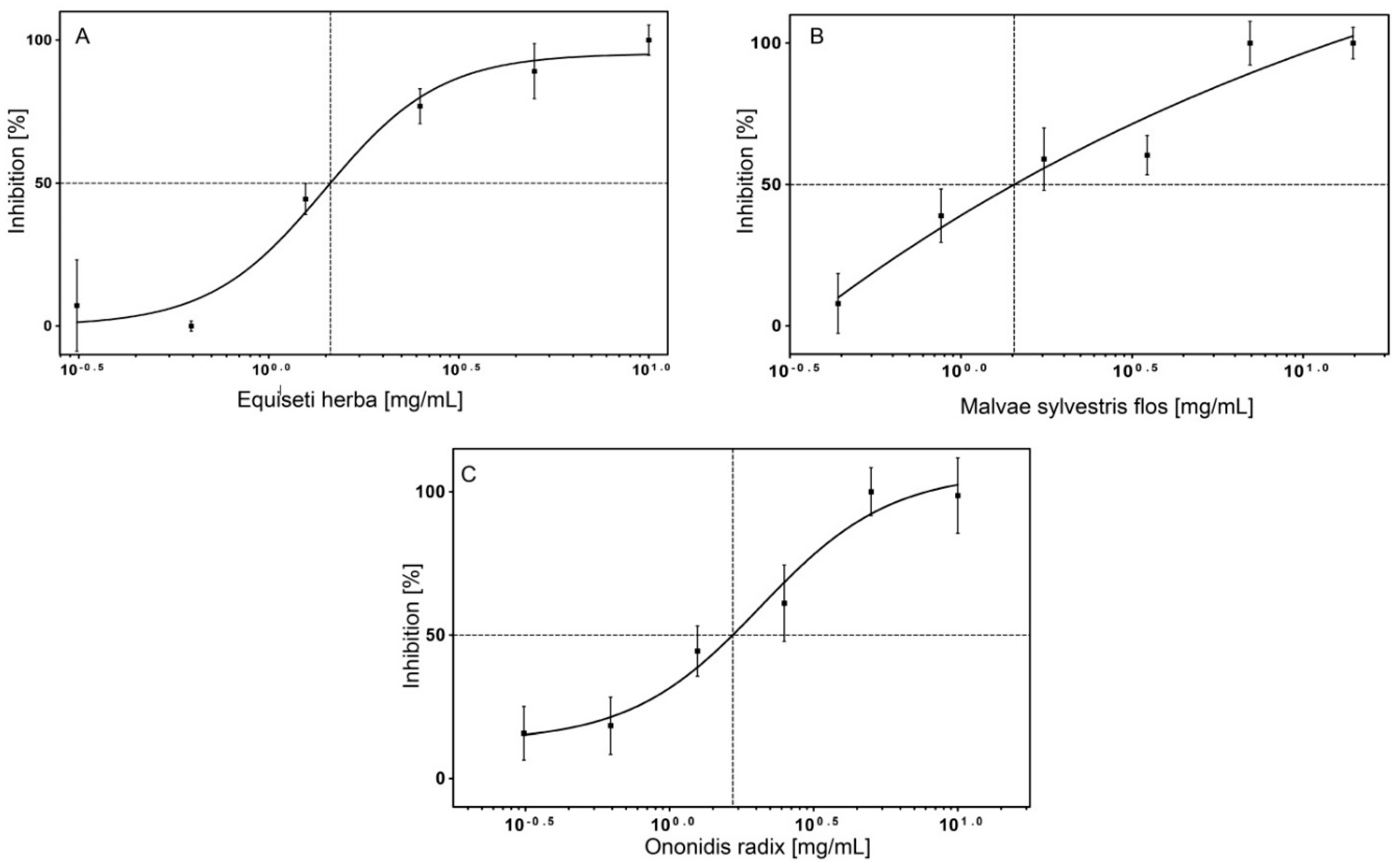
2.4. Inhibitor Testing with Surface Displayed Hyal-1



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | IC50 Value [µM] |
|---|---|
| Gypsophila saponin 2 | 108 |
| SA1657 | 371 |
| SA1641 | 296 |
| Plant Extract | Inhibition % [10 mg/mL] | IC50 Value [mg/mL] |
|---|---|---|
| Hennae folium | 0 | n.d. |
| Equiseti herba | 100 | 1.5 |
| Betulae folium | 61 | n.d. |
| Ononidis radix | 81 | 1.7 |
| Bucco folium | 21 | n.d. |
| Maydis stigma | 47 | n.d. |
| Malvae sylvestris flos | 100 | 1.4 |
| Solidaginis herba | 100 | 4.9 |
| Chebulae fructus | 0 | n.d. |
| Coptis rhizome | 0 | n.d. |
| Cranberry | 10 | n.d. |
| Althaeae radix | 60 | n.d. |
| Hydrastis rhizoma | 7 | n.d. |
| Mahoniae radix | 26 | n.d. |

3. Experimental Section
3.1. Harvesting Whole Cells of E. coli
3.2. “Stains-All” Assay
3.3. Bacterial Strains, Plasmids and Culture Conditions
3.4. Cloning of Artificial Autotransporter Gene Used for Autodisplay
3.5. Whole Cell ELISA
3.6. Membrane Protein Isolation
3.7. Western Blot
4. Conclusions
Author Contributions
Conflicts of Interest
References
- Heldin, P. Importance of hyaluronan biosynthesis and degradation in cell differentiation and tumor formation. Braz. J. Med. Biol. Res. 2003, 36, 967–973. [Google Scholar] [CrossRef] [PubMed]
- Bourguignon, L.Y.; Singleton, P.A.; Diedrich, F.; Stern, R.; Gilad, E. CD44 interaction with Na+-H+ exchanger (NHE1) creates acidic microenvironments leading to hyaluronidase-2 and cathepsin B activation and breast tumor cell invasion. J. Biol. Chem. 2004, 279, 26991–27007. [Google Scholar] [CrossRef] [PubMed]
- Sattar, A.; Rooney, P.; Kumar, S.; Pye, D.; West, D.C.; Scott, I.; Ledger, P. Application of angiogenic oligosaccharides of hyaluronan increases blood vessel numbers in rat skin. J. Investig. Dermatol. 1994, 103, 576–579. [Google Scholar] [CrossRef] [PubMed]
- Slevin, M.; Kumar, S.; Gaffney, J. Angiogenic oligosaccharides of hyaluronan induce multiple signaling pathways affecting vascular endothelial cell mitogenic and wound healing responses. J. Biol. Chem. 2002, 277, 41046–41059. [Google Scholar] [CrossRef] [PubMed]
- Stern, R.; Jedrzejas, M.J. Hyaluronidases: Their genomics, structures, and mechanisms of action. Chem. Rev. 2006, 106, 818–839. [Google Scholar] [CrossRef] [PubMed]
- Noble, P.W.; McKee, C.M.; Cowman, M.; Shin, H.S. Hyaluronan fragments activate an NF-kappa B/I-kappa B alpha autoregulatory loop in murine macrophages. J. Exp. Med. 1996, 183, 2373–2378. [Google Scholar] [CrossRef] [PubMed]
- Jedrzejas, M.J.; Stern, R. Structures of vertebrate hyaluronidases and their unique enzymatic mechanism of hydrolysis. Proteins Struct. Funct. Bioinform. 2005, 61, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Csoka, A.B.; Frost, G.I.; Stern, R. The six Hyaluronidase-like genes in the human und mouse genomes. Matrix Biol. 2001, 20, 499–508. [Google Scholar] [CrossRef]
- Chao, K.L.; Muthukumar, L.; Herzberg, O. Structure of human hyaluronidase-1, a hyaluronan hydrolyzing enzyme involved in tumor growth and angiogenesis. Biochemistry 2007, 46, 6911–6920. [Google Scholar] [CrossRef] [PubMed]
- Lokeshwar, V.B.; Rubinowicz, D.; Schroeder, G.L.; Forgacs, E.; Minna, J.D.; Block, N.L.; Nadji, M.; Lokeshwar, B.L. Stromal and epithelial expression of tumor markers hyaluronic acid and HYAL1 hyaluronidase in prostate cancer. J. Biol. Chem. 2001, 276, 11922–11932. [Google Scholar] [CrossRef] [PubMed]
- Lokeshwar, V.B.; Obek, C.; Pham, H.T.; Wei, D.; Young, M.J.; Duncan, R.C.; Soloway, M.S.; Block, N.L. Urinary hyaluronic acid and hyaluronidase: Markers for bladder cancer detection and evaluation of grade. J. Urol. 2000, 163, 348–356. [Google Scholar] [CrossRef]
- Tan, J.X.; Wang, X.Y.; Li, H.Y.; Su, X.L.; Wang, L.A.; Ran, L.A.; Zheng, K.; Ren, G.S. HYAL1 overexpression is correlated with the malignant behavior of human breast cancer. Int. J. Cancer 2011, 128, 1303–1315. [Google Scholar] [CrossRef] [PubMed]
- Mio, K.; Stern, R. Inhibitors of the hyaluronidases. Matrix Biol. 2002, 21, 31–37. [Google Scholar] [CrossRef]
- Jentsch, H.; Pomowski, R.; Kundt, G.; Gocke, R. Treatment of gingivitis with hyaluronan. J. Clin. Periodontol. 2003, 30, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Isoyama, T.; Thwaites, D.; Selzer, M.G.; Carey, R.I.; Barbucci, R.; Lokeshwar, V.B. Differential selectivity of hyaluronidase inhibitors toward acidic and basic hyaluronidases. Glycobiology 2006, 16, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Hofinger, E.S.A.; Spickenreither, M.; Oschmann, J.; Bernhardt, G.; Rudolph, R.; Buschauer, A. Recombinant human hyaluronidase Hyal-1: Insect cells versus Escherichia coli as expression system and identification of low molecular weight inhibitors. Glycobiology 2007, 17, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Jose, J.; Meyer, T.F. The autodisplay story, from discovery to biotechnical and biomedical applications. Microbiol. Mol. Biol. Rev. 2007, 71, 600–619. [Google Scholar] [CrossRef] [PubMed]
- Gratz, A.; Bollacke, A.; Stephan, S.; Nienberg, C.; le Borgne, M.; Gotz, C.; Jose, J. Functional display of heterotetrameric human protein kinase CK2 on Escherichia coli: A novel tool for drug discovery. Microb. Cell Factories 2015, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jose, J.; von Schwichow, S. Autodisplay of active sorbitol dehydrogenase (SDH) yields a whole cell biocatalyst for the synthesis of rare sugars. ChemBioChem 2004, 5, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Detzel, C.; Maas, R.; Jose, J. Autodisplay of nitrilase from Alcaligenes faecalis in E. coli yields a whole cell biocatalyst for the synthesis of enantiomerically pure (R)-mandelic acid. ChemCatChem 2011, 3, 719–725. [Google Scholar] [CrossRef]
- Detzel, C.; Maas, R.; Tubeleviciute, A.; Jose, J. Autodisplay of nitrilase from Klebsiella pneumoniae and whole-cell degradation of oxynil herbicides and related compounds. Appl. Microbiol. Biotechnol. 2013, 97, 4887–4896. [Google Scholar] [CrossRef] [PubMed]
- Lattemann, C.T.; Maurer, J.; Gerland, E.; Meyer, T.F. Autodisplay: Functional display of active beta-lactamase on the surface of Escherichia coli by the AIDA-I autotransporter. J. Bacteriol. 2000, 182, 3726–3733. [Google Scholar] [CrossRef] [PubMed]
- Kaessler, A.; Olgen, S.; Jose, J. Autodisplay of catalytically active human hyaluronidase hPH-20 and testing of enzyme inhibitors. Eur. J. Pharm. Sci. 2011, 42, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, S.D.; Hannemann, F.; Teese, M.G.; Bernhardt, R.; Jose, J. Autodisplay of functional CYP106A2 in Escherichia coli. J. Biotechnol. 2012, 161, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, S.D.; Jose, J. Expression of active human P450 3A4 on the cell surface of Escherichia coli by Autodisplay. J. Biotechnol. 2012, 161, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Schultheiss, E.; Paar, C.; Schwab, H.; Jose, J. Functional esterase surface display by the autotransporter pathway in Escherichia coli. J. Mol. Catal. B Enzym. 2002, 18, 89–97. [Google Scholar] [CrossRef]
- Tozakidis, I.E.; Sichwart, S.; Teese, M.G.; Jose, J. Autotransporter mediated esterase display on Zymomonas mobilis and Zymobacter palmae. J. Biotechnol. 2014, 191, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Schultheiss, E.; Weiss, S.; Winterer, E.; Maas, R.; Heinzle, E.; Jose, J. Esterase autodisplay: Enzyme engineering and whole-cell activity determination in microplates with pH sensors. Appl. Environ. Microbiol. 2008, 74, 4782–4791. [Google Scholar] [CrossRef] [PubMed]
- Kranen, E.; Detzel, C.; Weber, T.; Jose, J. Autodisplay for the co-expression of lipase and foldase on the surface of E. coli: Washing with designer bugs. Microb. Cell Factories 2014, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jose, J. Autodisplay: Efficient bacterial surface display of recombinant proteins. Appl. Microbiol. Biotechnol. 2006, 69, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.; Jung, M.Y.; Park, J.H.; Jung, G.C.; Hong, Y.S.; Yeom, C.H.; Lee, S. Production of human hyaluronidase in a plant-derived protein expression system: Plant-based transient production of active human hyaluronidase. Protein Expr. Purif. 2010, 74, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Maurer, J.; Jose, J.; Meyer, T.F. Autodisplay: One-component system for efficient surface display and release of soluble recombinant proteins from Escherichia coli. J. Bacteriol. 1997, 179, 794–804. [Google Scholar] [PubMed]
- Schmidt, G.; Jann, B.; Jann, K. Immunochemistry of R lipopolysaccharides of Escherichia coli—Studies on R-mutants with an incomplete core, derived from E. coli O8:K27. Eur. J. Biochem. 1970, 16, 382–392. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.C.; Park, J.H.; Park, S.H.; Lebeault, J.M.; Pan, J.G. Expression of carboxymethylcellulase on the surface of Escherichia coli using Pseudomonas syringae ice nucleation protein. Enzym. Microb. Technol. 1998, 22, 348–354. [Google Scholar] [CrossRef]
- Park, S.H.; Liu, G.P.; Topping, T.B.; Cover, W.H.; Randall, L.L. Modulation of folding pathways of exported proteins by the leader sequence. Science 1988, 239, 1033–1035. [Google Scholar] [CrossRef] [PubMed]
- Frost, G.I.; Csoka, A.B.; Wong, T.; Stern, R. Purification, cloning, and expression of human plasma hyaluronidase. Biochem. Biophys. Res. Commun. 1997, 236, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Afify, A.M.; Stern, M.; Guntenhoner, M.; Stern, R. Purification and characterization of human serum hyaluronidase. Arch. Biochem. Biophys. 1993, 305, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Bottger, S.; Melzig, M.F. The influence of saponins on cell membrane cholesterol. Bioorg. Med. Chem. 2013, 21, 7118–7124. [Google Scholar] [CrossRef] [PubMed]
- Bottger, S.; Westhof, E.; Siems, K.; Melzig, M.F. Structure-activity relationships of saponins enhancing the cytotoxicity of ribosome-inactivating proteins type I (RIP-I). Toxicon 2013, 73, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Weng, A.; Jenett-Siems, K.; Schmieder, P.; Bachran, D.; Bachran, C.; Gorick, C.; Thakur, M.; Fuchs, H.; Melzig, M.F. A convenient method for saponin isolation in tumour therapy. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2010, 878, 713–718. [Google Scholar] [CrossRef] [PubMed]
- Kuppusamy, U.R.; Das, N.P. Inhibitory effects of flavonoids on several venom hyaluronidases. Experientia 1991, 47, 1196–1200. [Google Scholar] [CrossRef] [PubMed]
- Hertel, W.; Peschel, G.; Ozegowski, J.H.; Muller, P.J. Inhibitory effects of triterpenes and flavonoids on the enzymatic activity of hyaluronic acid-splitting enzymes. Arch. Pharm. 2006, 339, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Furuya, T.; Yamagata, S.; Shimoyama, Y.; Fujihara, M.; Morishima, N.; Ohtsuki, K. Biochemical characterization of glycyrrhizin as an effective inhibitor for hyaluronidases from bovine testis. Biol. Pharm. Bull. 1997, 20, 973–977. [Google Scholar] [CrossRef] [PubMed]
- Rafsanjany, N.; Lechtenberg, M.; Petereit, F.; Hensel, A. Antiadhesion as a functional concept for protection against uropathogenic Escherichia coli: In vitro studies with traditionally used plants with antiadhesive activity against uropathognic Escherichia coli. J. Ethnopharmacol. 2013, 145, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Hagerman, A.E.; Rice, M.E.; Ritchard, N.T. Mechanisms of protein precipitation for two tannins, pentagalloyl glucose and epicatechin(16) (4→8) catechin (procyanidin). J. Agric. Food Chem. 1998, 46, 2590–2595. [Google Scholar] [CrossRef]
- Schmidt, G.; Jann, B.; Jann, K. Immunochemistry of R lipopolysaccharides of Escherichia coli. Different core regions in the lipopolysaccharides of O group 8. Eur. J. Biochem. 1969, 10, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Petermann, K.; Vordenbaumen, S.; Pyun, J.C.; Braukmann, A.; Bleck, E.; Schneider, M.; Jose, J. Autodisplay of 60-kDa Ro/SS-A antigen and development of a surface display enzyme-linked immunosorbent assay for systemic lupus erythematosus patient sera screening. Anal. Biochem. 2010, 407, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Park, M.; Yoo, G.; Bong, J.H.; Jose, J.; Kang, M.J.; Pyun, J.C. Isolation and characterization of the outer membrane of Escherichia coli with autodisplayed Z-domains. Biochim. Biophys. Acta 2015, 1848, 842–847. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Hyaluronic acid sodium salt from Streptococcus equi, proteinase K and PMSF were obtained from Sigma-Aldrich Co (Munich, Germany). Stabilized 1,4-dioxane, DNaseI were purchased from AppliChem (Darmstadt, Germany). Stains-all was obtained from Acros Organics (part of Fisher Scientific GmbH, Nidderau, Germany). All restriction enzymes, ligase and the corresponding buffers were from NEB (country road, USA). Taq-DNA-polymerase was a mastermix from Eppendorf (Hamburg, Germany). The polyclonal murine antiserum raised against Hyal-1 (epitope aa 60–154, unconjugated) was ordered from Abnova (Berlin, Germany) and the peroxidase conjugated rabbit anti‑mouse IgG was ordered from antibodies-online (Aachen, Germany). The HRP substrate, Western Blot Luminol Reagent (ImmunoCruz™), was from Santa Cruz Biotechnology (Heidelberg, Germany). Glycyrrhizic acid, lysozyme and aprotinin were purchased from Roth (Karlsruhe, Germany).
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orlando, Z.; Lengers, I.; Melzig, M.F.; Buschauer, A.; Hensel, A.; Jose, J. Autodisplay of Human Hyaluronidase Hyal-1 on Escherichia coli and Identification of Plant-Derived Enzyme Inhibitors. Molecules 2015, 20, 15449-15468. https://doi.org/10.3390/molecules200915449
Orlando Z, Lengers I, Melzig MF, Buschauer A, Hensel A, Jose J. Autodisplay of Human Hyaluronidase Hyal-1 on Escherichia coli and Identification of Plant-Derived Enzyme Inhibitors. Molecules. 2015; 20(9):15449-15468. https://doi.org/10.3390/molecules200915449
Chicago/Turabian StyleOrlando, Zoya, Isabelle Lengers, Matthias F. Melzig, Armin Buschauer, Andreas Hensel, and Joachim Jose. 2015. "Autodisplay of Human Hyaluronidase Hyal-1 on Escherichia coli and Identification of Plant-Derived Enzyme Inhibitors" Molecules 20, no. 9: 15449-15468. https://doi.org/10.3390/molecules200915449