
Xanthohumol, a Prenylated Chalcone from Hops, Inhibits the Viability and Stemness of Doxorubicin-Resistant MCF-7/ADR Cells


Abstract
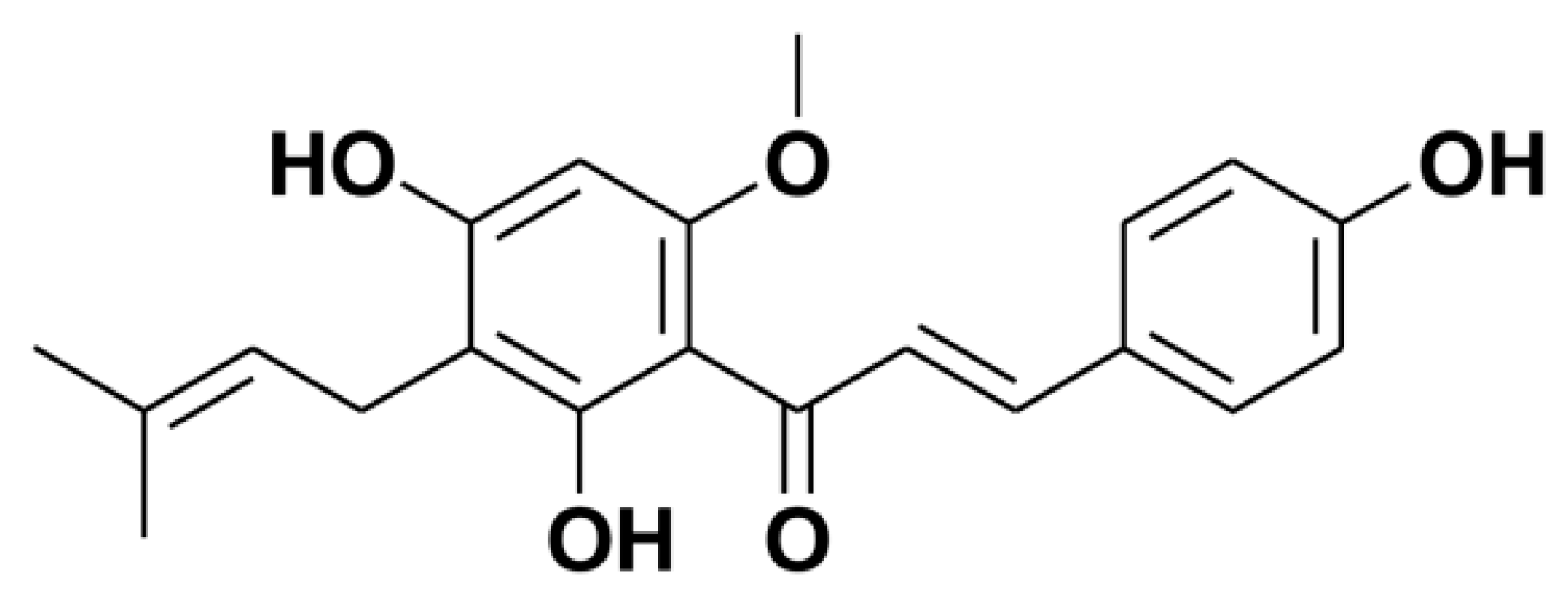
:1. Introduction
2. Results
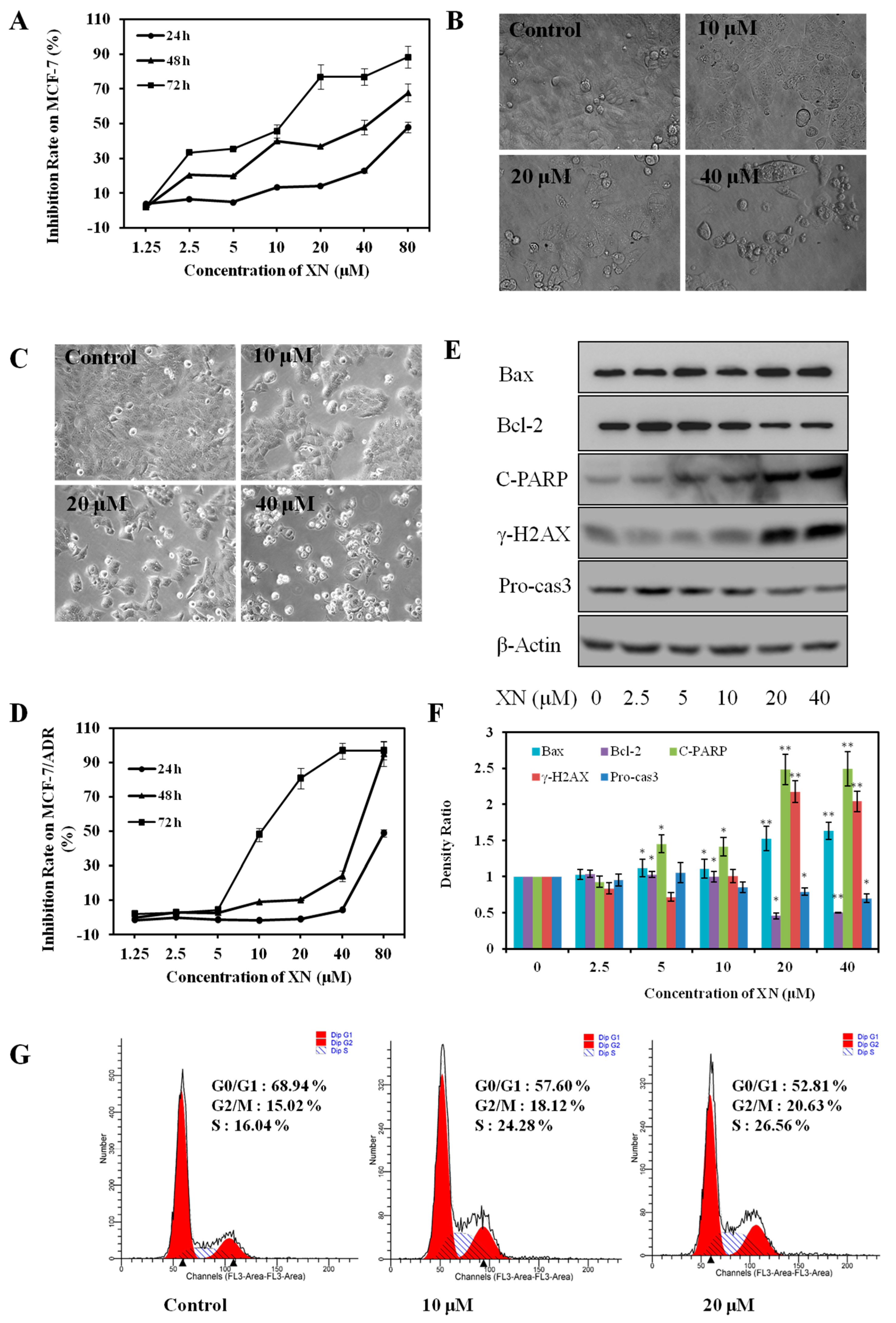
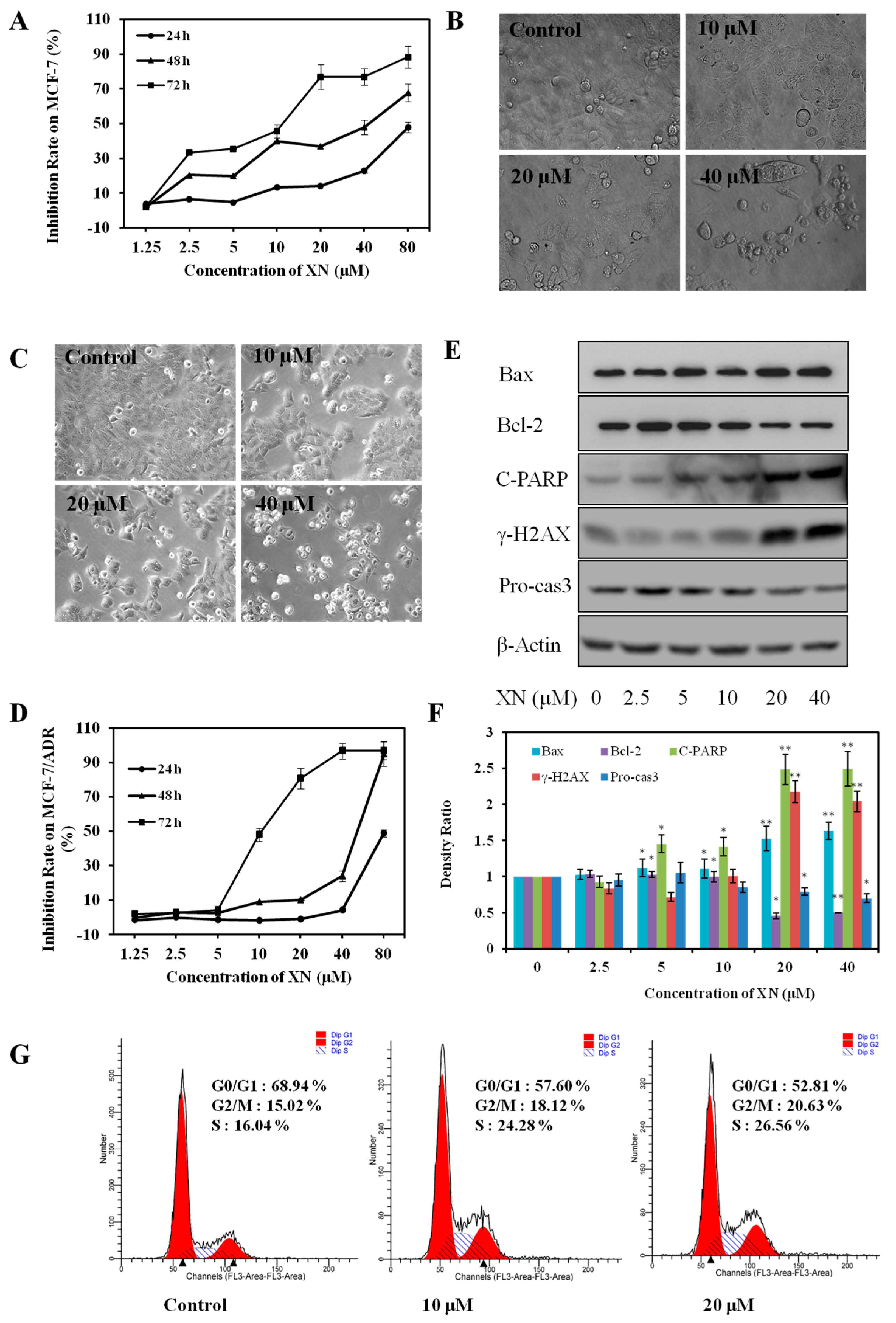
2.1. XN Inhibits Viability, Induces Apoptosis, and Arrests Cell Cycle in MCF-7/ADR Cells
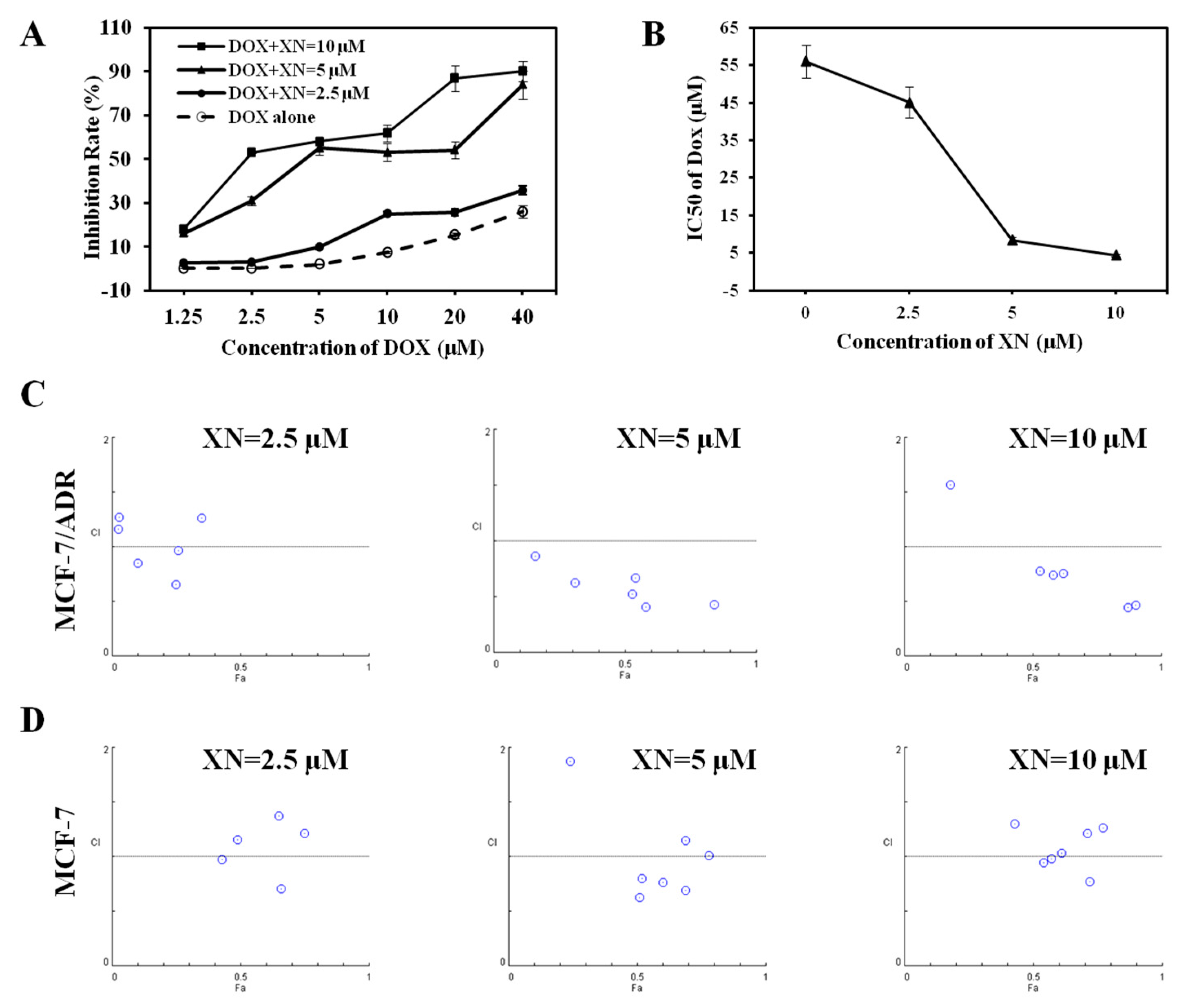
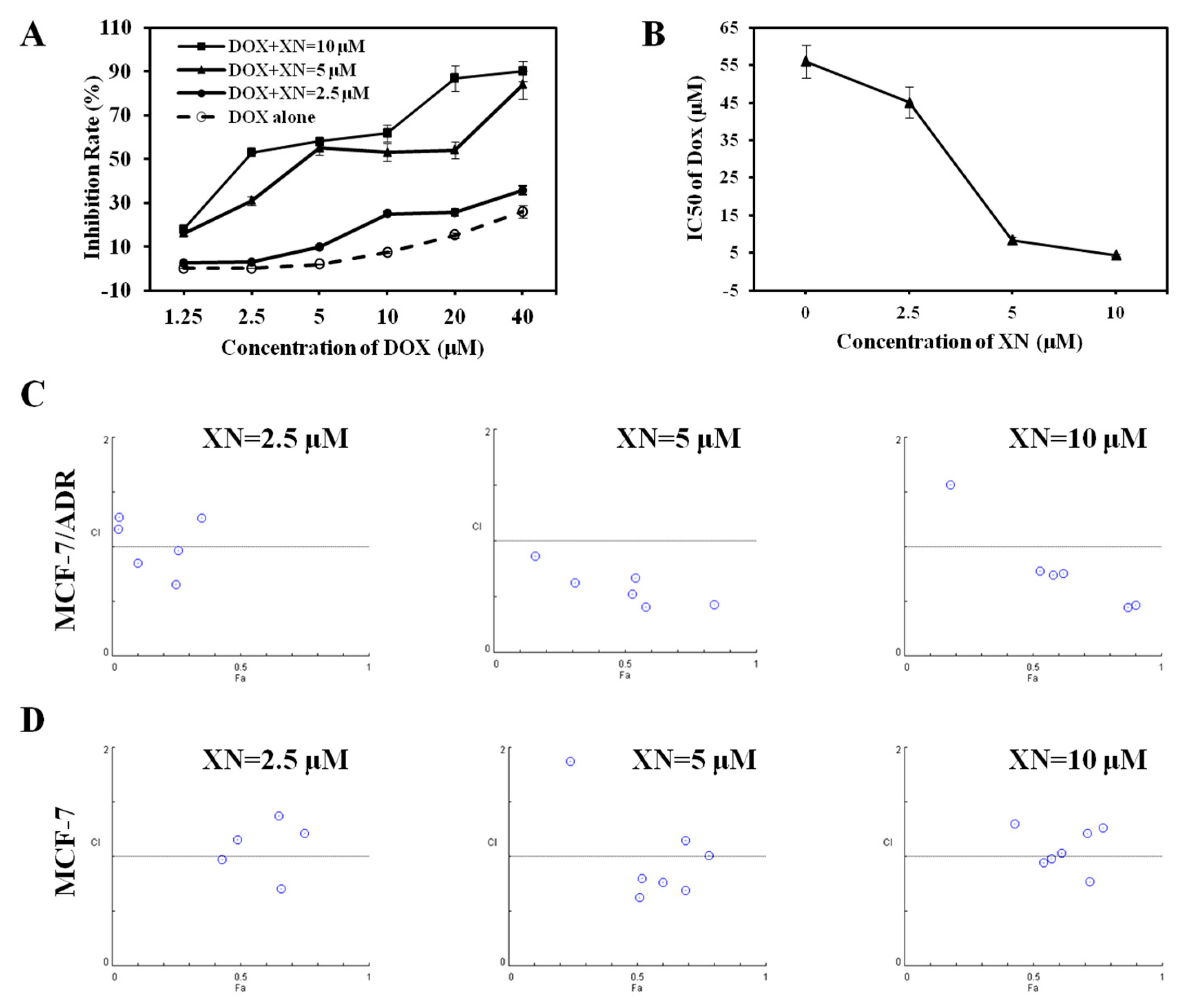
2.2. Synergic Effect of XN in Combination with DOX
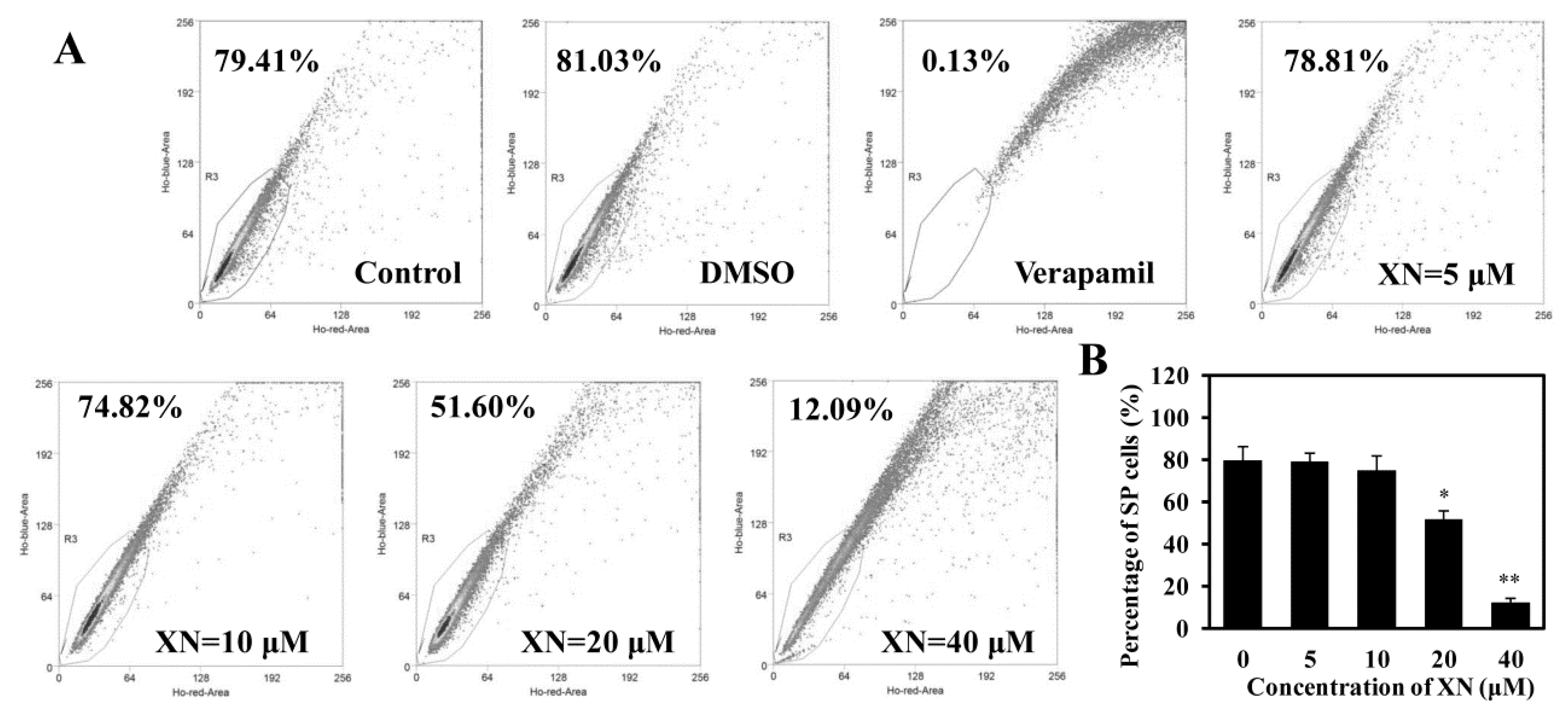
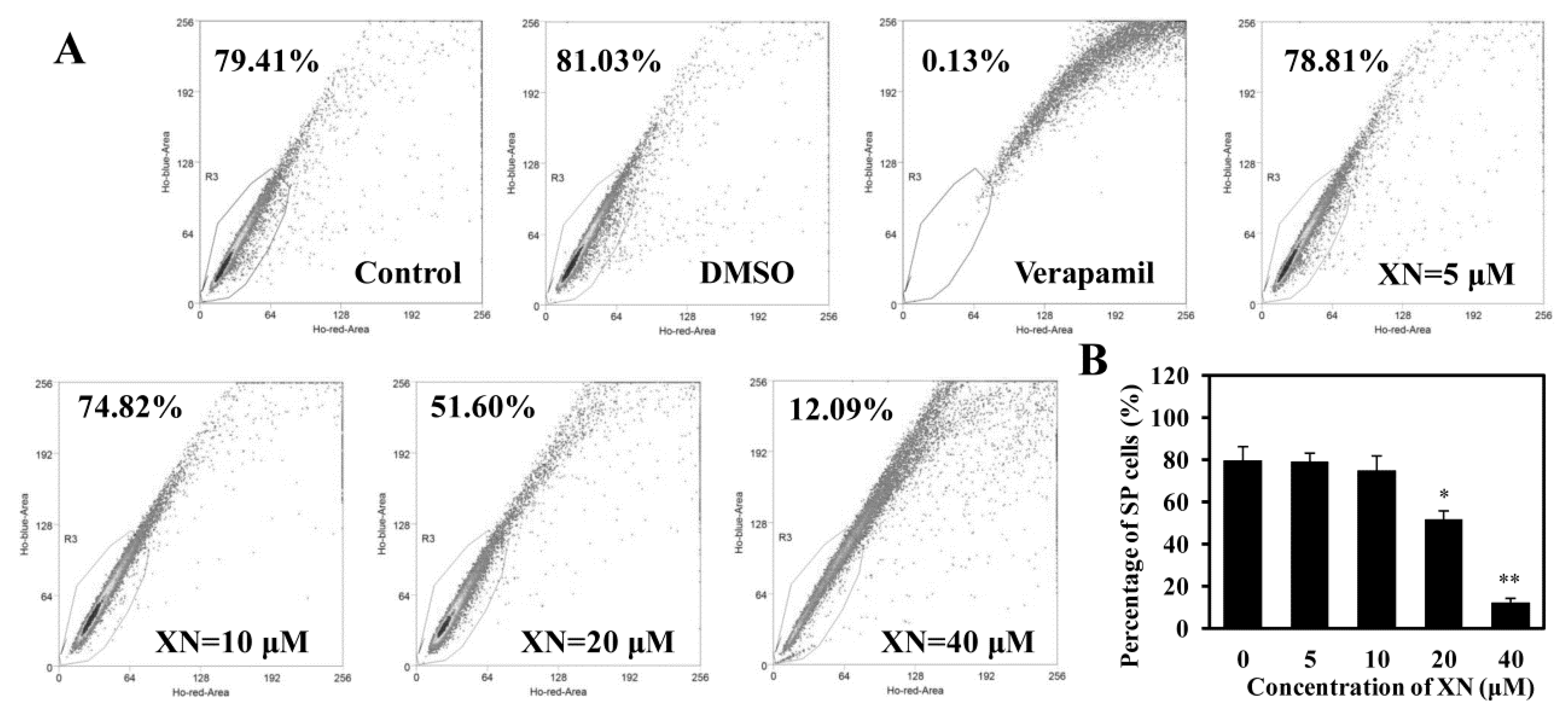
2.3. XN Decreases the SP Cell Fraction in MCF-7/ADR Cells
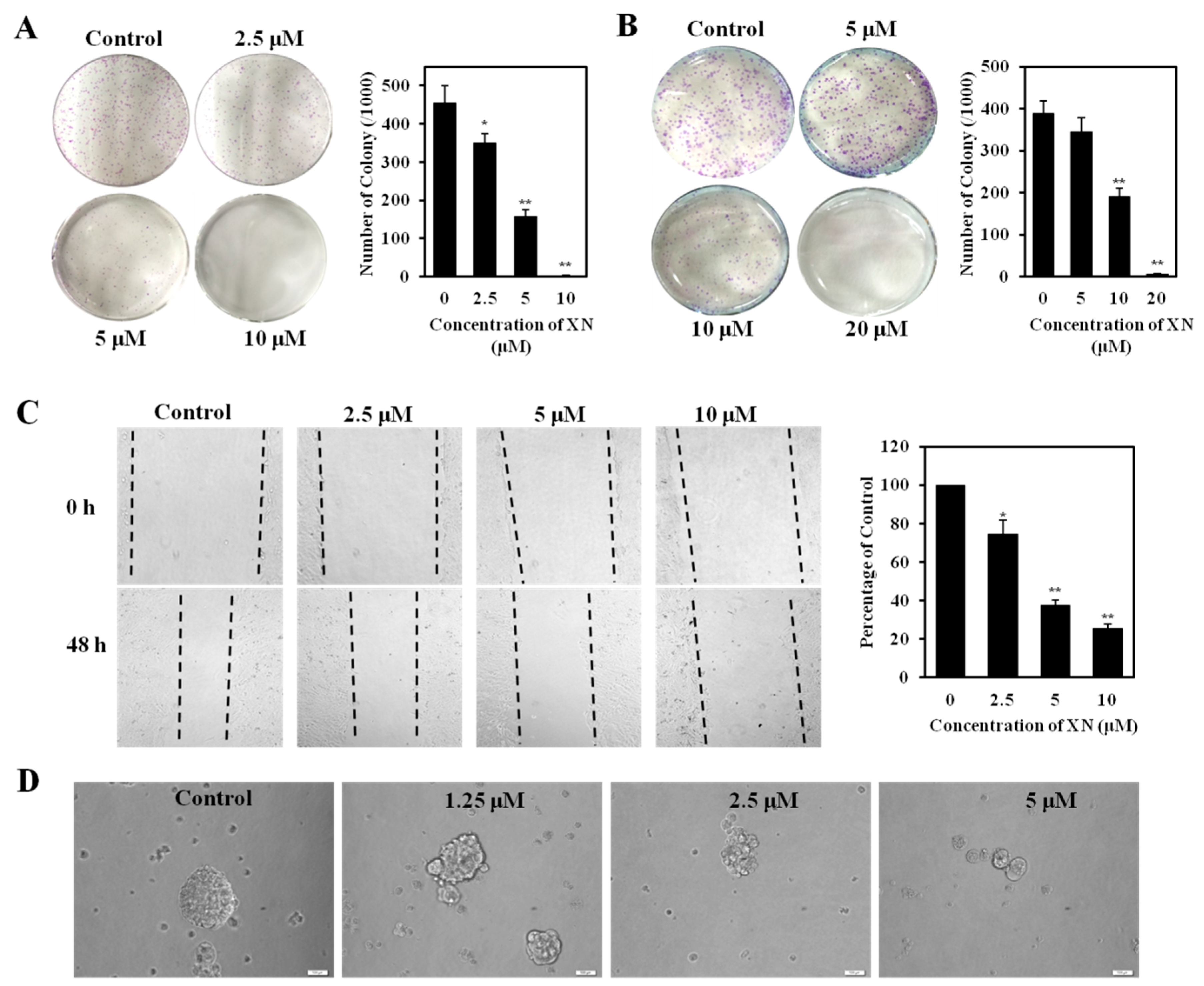
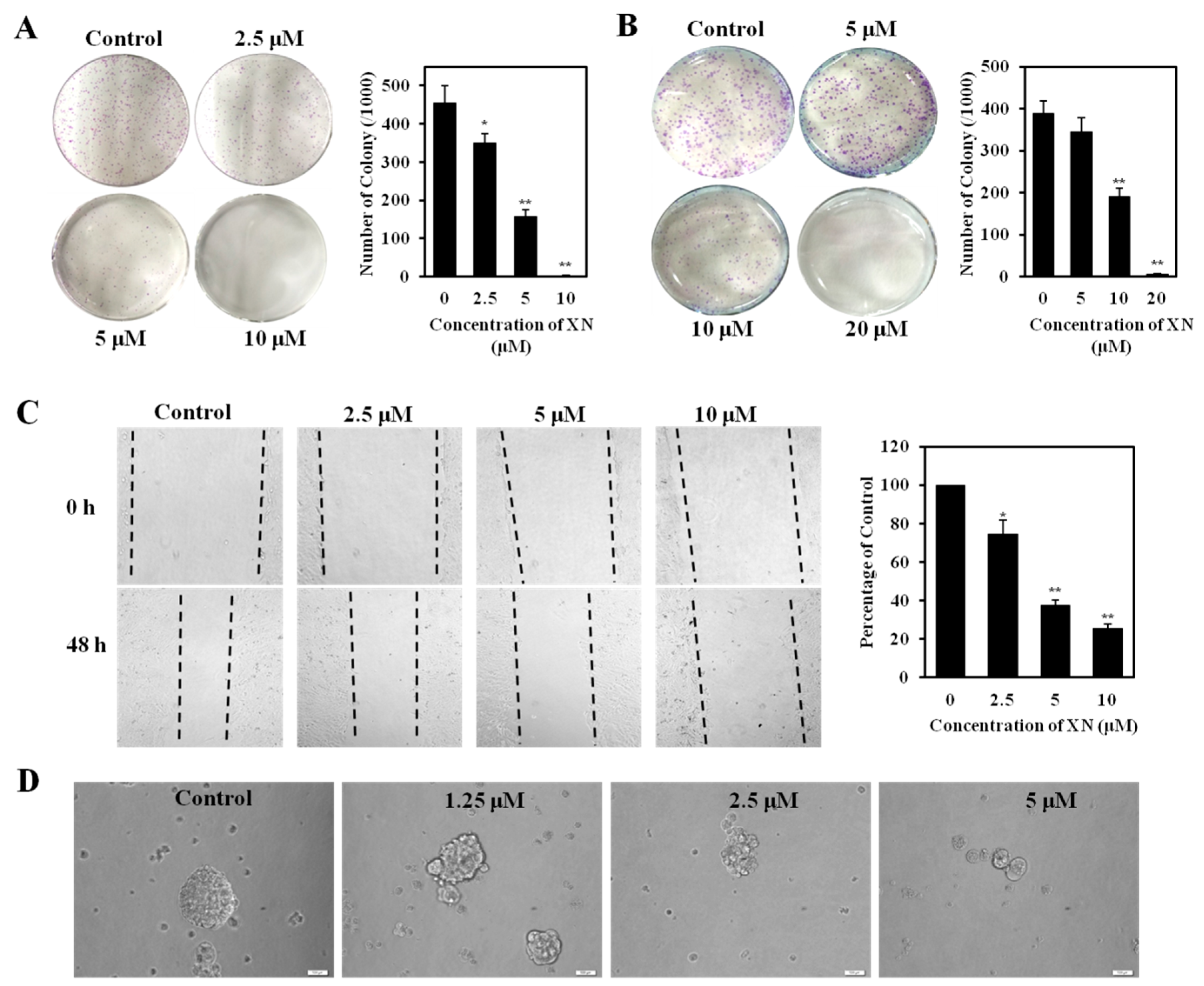
2.4. XN Reduces Clonogenic Survival, Migration, and Sphere Formation of CSC-Like Cells
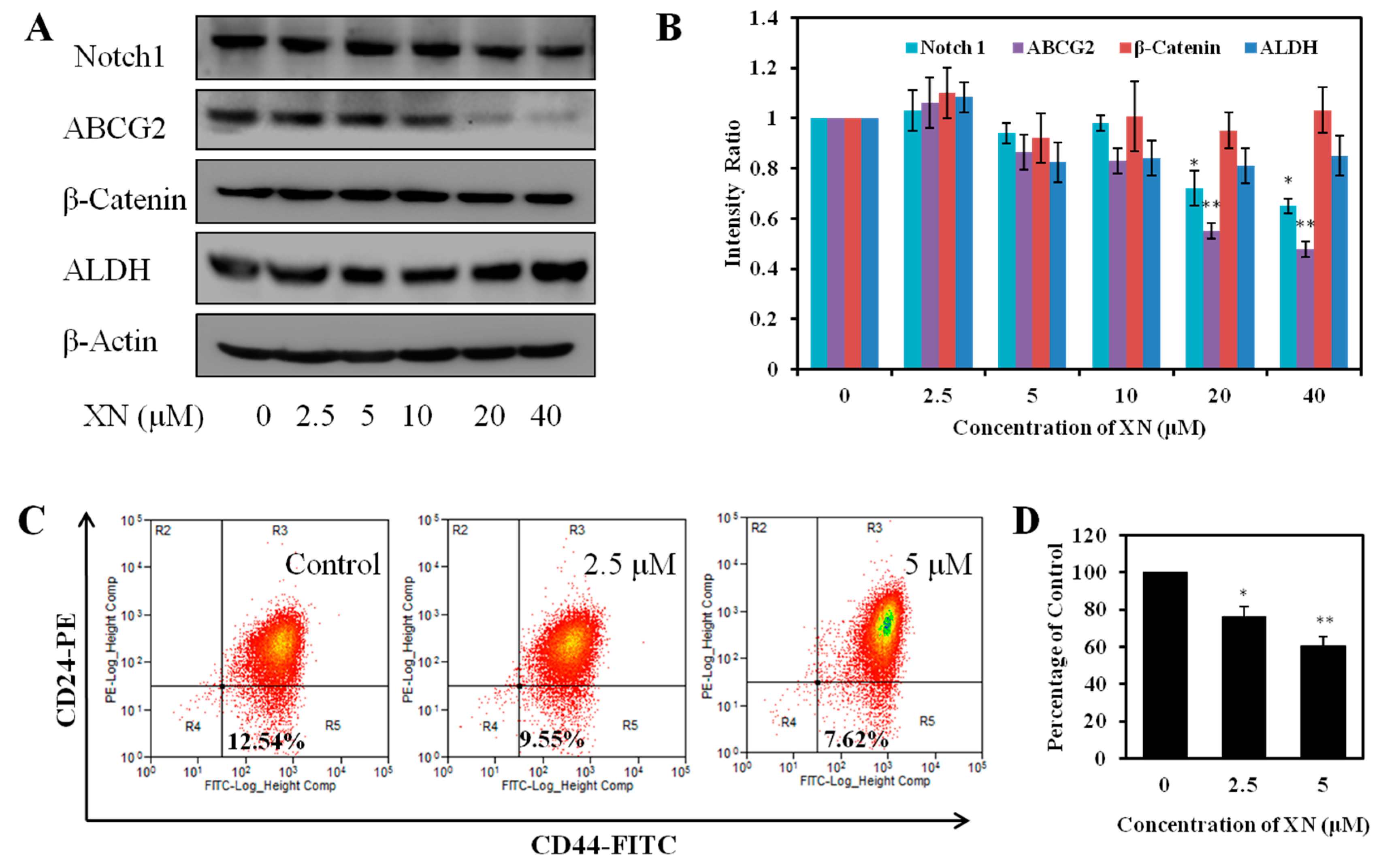
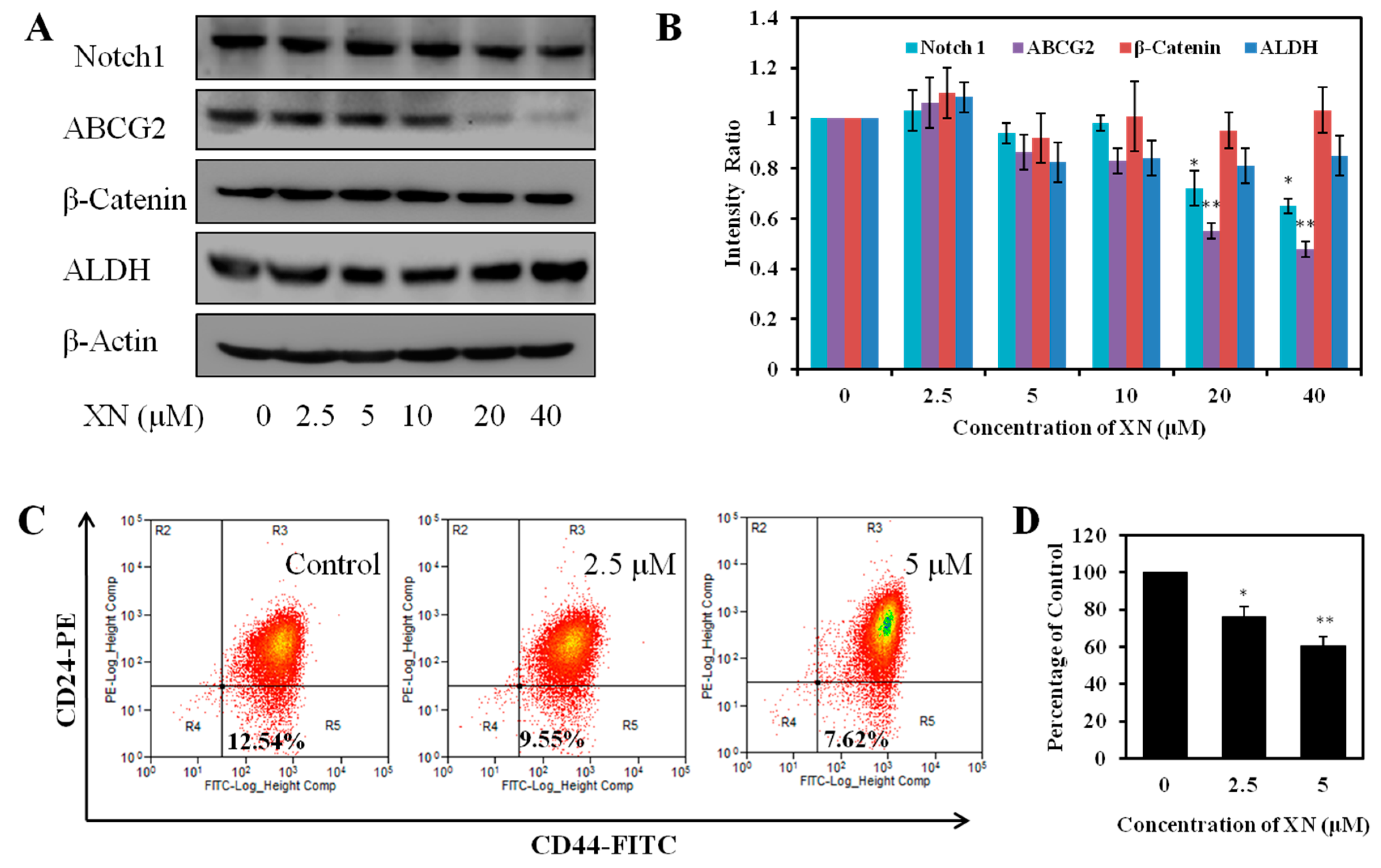
2.5. XN Reduces the Expression of Cancer Stem-Like Cell Markers
3. Discussion
4. Materials and Methods
4.1. Materials and Cells
4.2. Cell Viability Assessment
4.3. Drug Combination and Calculation of Synergism
4.4. Colony Formation Assay
4.5. Cell Migration Assay
4.6. Side Population (SP) Cells Analysis
4.7. Western Blotting Assay
4.8. Mammosphere Formation Assay
4.9. CD44 and CD24 Staining
4.10. Mammosphere Formation Assay
4.11. Data Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Tang, Y.; Wang, Y.; Kiani, M.F.; Wang, B. Classification, treatment strategy, and associated drug resistance in breast cancer. Clin. Breast Cancer 2016, 16, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Sotiropoulou, P.A.; Christodoulou, M.S.; Silvani, A.; Herold-Mende, C.; Passarella, D. Chemical approaches to targeting drug resistance in cancer stem cells. Drug Discov. Today 2014, 19, 1547–1562. [Google Scholar] [CrossRef] [PubMed]
- Mannello, F. Understanding breast cancer stem cell heterogeneity: Time to move on to a new research paradigm. BMC Med. 2013, 11, 169. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.C.; Kannappan, R.; Reuter, S.; Kim, J.H.; Aggarwal, B.B. Chemosensitization of tumors by resveratrol. Ann. N. Y. Acad. Sci. 2011, 1215, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.T.; Welch, K.D.; Panter, K.E.; Gardner, D.R.; Garrossian, M.; Chang, C.W.T. Cyclopamine: From cyclops lambs to cancer treatment. J. Agric. Food Chem. 2014, 62, 7355–7362. [Google Scholar] [CrossRef] [PubMed]
- Block, K.I.; Gyllenhaal, C.; Lowe, L.; Amedei, A.; Amin, A.R.M.R.; Amin, A.; Aquilano, K.; Arbiser, J.; Arreola, A.; Arzumanyan, A.; et al. Designing a broad-spectrum integrative approach for cancer prevention and treatment. Semin. Cancer Biol. 2015, 35, S276–S304. [Google Scholar] [CrossRef] [PubMed]
- Stevens, J.F.; Taylor, A.W.; Clawson, J.E.; Deinzer, M.L. Fate of xanthohumol and related prenylflavonoids from hops to beer. J. Agric. Food Chem. 1999, 47, 2421–2428. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Hansen, P.E.; Wang, G.; Qiu, L.; Dong, J.; Yin, H.; Qian, Z.; Yang, M.; Miao, J. Pharmacological profile of xanthohumol, a prenylated flavonoid from hops (Humulus lupulus). Molecules 2015, 20, 754–779. [Google Scholar] [CrossRef] [PubMed]
- Strathmann, J.; Gerhauser, C. Natural Compounds as Inducers of Cell Death; Springer: Dordrecht, The Netherlands, 2012; Volume 1, pp. 69–93. [Google Scholar]
- Miranda, C.L.; Stevens, J.F.; Helmrich, A.; Henderson, M.C.; Rodriguez, R.J.; Yang, Y.H.; Deinzer, M.L.; Barnes, D.W.; Buhler, D.R. Antiproliferative and cytotoxic effects of prenylated flavonoids from hops (Humulus lupulus) in human cancer cell lines. Food Chem. Toxicol. 1999, 37, 271–285. [Google Scholar] [CrossRef]
- Kang, Y.; Park, M.A.; Heo, S.W.; Park, S.Y.; Kang, K.W.; Park, P.H.; Kim, J.A. The radio-sensitizing effect of xanthohumol is mediated by STAT3 and EGFR suppression in doxorubicin-resistant MCF-7 human breast cancer cells. BBA-Gen. Subj. 2013, 1830, 2638–2648. [Google Scholar] [CrossRef] [PubMed]
- Szliszka, E.; Jaworska, D.; Ksek, M.; Czuba, Z.P.; Krol, W. Targeting death receptor TRAIL-R2 by chalcones for TRAIL-induced apoptosis in cancer cells. Int. J. Mol. Sci. 2012, 13, 15343–15359. [Google Scholar] [CrossRef] [PubMed]
- Szliszka, E.; Czuba, Z.; Mazur, B.; Sedek, L.; Paradysz, A.; Krol, W. Chalcones enhance TRAIL-induced apoptosis in prostate cancer cells. Int. J. Mol. Sci. 2009, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.W.; Cooney, J.; Jensen, D.; Li, Y.; Paxton, J.W.; Birch, N.P.; Scheepens, A. Hop-derived prenylflavonoids are substrates and inhibitors of the efflux transporter breast cancer resistance protein (BCRP/ABCG2). Mol. Nutr. Food Res. 2014, 14, 2099–2110. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, Y.; Zhong, J. Side population cells and drug resistance in breast cancer. Mol. Med. Rep. 2015, 30, 4297–4302. [Google Scholar] [CrossRef] [PubMed]
- Balbuena, J.; Pachon, G.; Lopez-Torrents, G.; Aran, J.M.; Castresana, J.S.; Petriz, J. ABCG2 is required to control the sonic hedgehog pathway in side population cells with stem-like properties. Cytom. Part A 2011, 79A, 672–683. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, K.; Xie, Y.; Baer, M.R.; Ross, D.D. Role of breast cancer resistance protein (BCRP/ABCG2) in cancer drug resistance. Biochem. Pharmacol. 2012, 83, 1084–1103. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, J.I.; Haber, M.; Henderson, M.J.; Norris, M.D. ABC transporters in cancer: More than just drug efflux pumps. Nat. Rev. Cancer 2010, 10, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Lee, I.-S.; Moon, A. 2-Hydroxychalcone and xanthohumol inhibit invasion of triple negative breast cancer cells. Chem. Biol. Interact. 2013, 203, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Kunnimalaiyaan, S.; Sokolowski, K.M.; Balamurugan, M.; Gamblin, T.C.; Kunnimalaiyaan, M. Xanthohumol inhibits notch signaling and induces apoptosis in hepatocellular carcinoma. PLoS ONE 2015, 10, e0127464. [Google Scholar] [CrossRef] [PubMed]
- Kunnimalaiyaan, S.; Trevino, J.; Tsai, S.; Gamblin, T.C.; Kunnimalaiyaan, M. Xanthohumol-mediated suppression of Notch1 signaling is associated with antitumor activity in human pancreatic cancer cells. Mol. Cancer Ther. 2015, 14, 1395–1403. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.C.; Talalay, P. Quantitative analysis of dose-effect relationships: The combined effects of multiple drugs or enzyme inhibitors. Adv. Enzym. Regul. 1984, 22, 27–55. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compound xanthohumol are available from the authors.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| XN (μM) | DOX (μM) | Effect | CI |
|---|---|---|---|
| 2.5 | 1.25 | 0.026 | 1.16 |
| 2.5 | 2.5 | 0.03 | 1.27 |
| 2.5 | 5 | 0.1 | 0.85 |
| 2.5 | 10 | 0.25 | 0.66 |
| 2.5 | 20 | 0.26 | 0.97 |
| 2.5 | 40 | 0.35 | 1.26 |
| 5 | 1.25 | 0.16 | 0.87 |
| 5 | 2.5 | 0.31 | 0.62 |
| 5 | 5 | 0.58 | 0.41 |
| 5 | 10 | 0.53 | 0.52 |
| 5 | 20 | 0.54 | 0.67 |
| 5 | 40 | 0.84 | 0.42 |
| 10 | 1.25 | 0.18 | 1.56 |
| 10 | 2.5 | 0.53 | 0.77 |
| 10 | 5 | 0.58 | 0.74 |
| 10 | 10 | 0.62 | 0.75 |
| 10 | 20 | 0.87 | 0.44 |
| 10 | 40 | 0.90 | 0.47 |
| XN (μM) | DOX (μM) | Effect | CI |
|---|---|---|---|
| 2.5 | 0.15 | 0.19 | 1.89 |
| 2.5 | 0.3 | 0.22 | 2.33 |
| 2.5 | 0.625 | 0.43 | 0.97 |
| 2.5 | 1.25 | 0.49 | 1.16 |
| 2.5 | 2.5 | 0.66 | 0.71 |
| 2.5 | 5 | 0.65 | 1.38 |
| 2.5 | 10 | 0.75 | 1.21 |
| 5 | 0.15 | 0.24 | 1.87 |
| 5 | 0.3 | 0.51 | 0.63 |
| 5 | 0.625 | 0.52 | 0.80 |
| 5 | 1.25 | 0.6 | 0.76 |
| 5 | 2.5 | 0.69 | 0.69 |
| 5 | 5 | 0.69 | 1.15 |
| 5 | 10 | 0.78 | 1.01 |
| 10 | 0.15 | 0.43 | 1.30 |
| 10 | 0.3 | 0.54 | 0.94 |
| 10 | 0.625 | 0.57 | 0.98 |
| 10 | 1.25 | 0.61 | 1.03 |
| 10 | 2.5 | 0.72 | 0.77 |
| 10 | 5 | 0.71 | 1.21 |
| 10 | 10 | 0.77 | 1.26 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, M.; Yin, H.; Qian, X.; Dong, J.; Qian, Z.; Miao, J. Xanthohumol, a Prenylated Chalcone from Hops, Inhibits the Viability and Stemness of Doxorubicin-Resistant MCF-7/ADR Cells. Molecules 2017, 22, 36. https://doi.org/10.3390/molecules22010036
Liu M, Yin H, Qian X, Dong J, Qian Z, Miao J. Xanthohumol, a Prenylated Chalcone from Hops, Inhibits the Viability and Stemness of Doxorubicin-Resistant MCF-7/ADR Cells. Molecules. 2017; 22(1):36. https://doi.org/10.3390/molecules22010036
Chicago/Turabian StyleLiu, Ming, Hua Yin, Xiaokun Qian, Jianjun Dong, Zhonghua Qian, and Jinlai Miao. 2017. "Xanthohumol, a Prenylated Chalcone from Hops, Inhibits the Viability and Stemness of Doxorubicin-Resistant MCF-7/ADR Cells" Molecules 22, no. 1: 36. https://doi.org/10.3390/molecules22010036