
Therapeutic Effect of Cistanoside A on Bone Metabolism of Ovariectomized Mice

Abstract
:
1. Introduction
2. Results and Discussion
2.1. Results
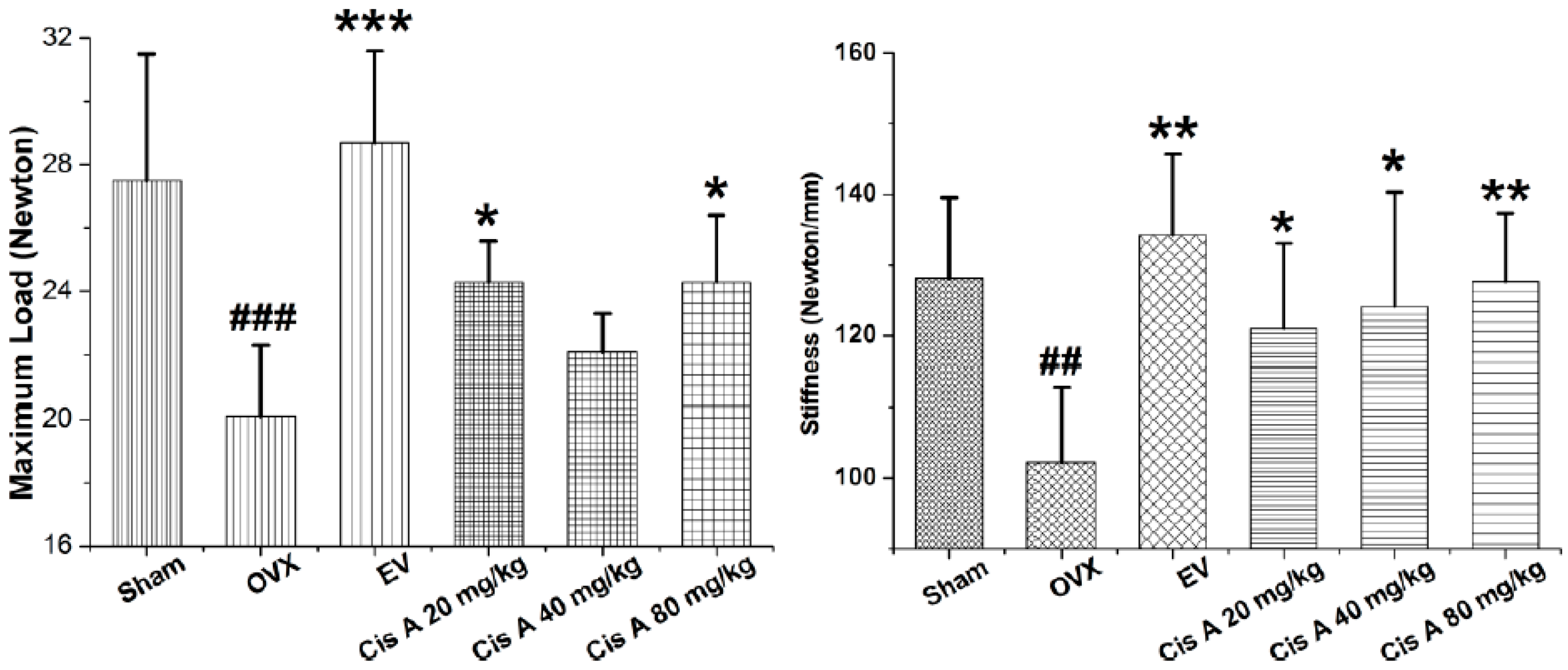
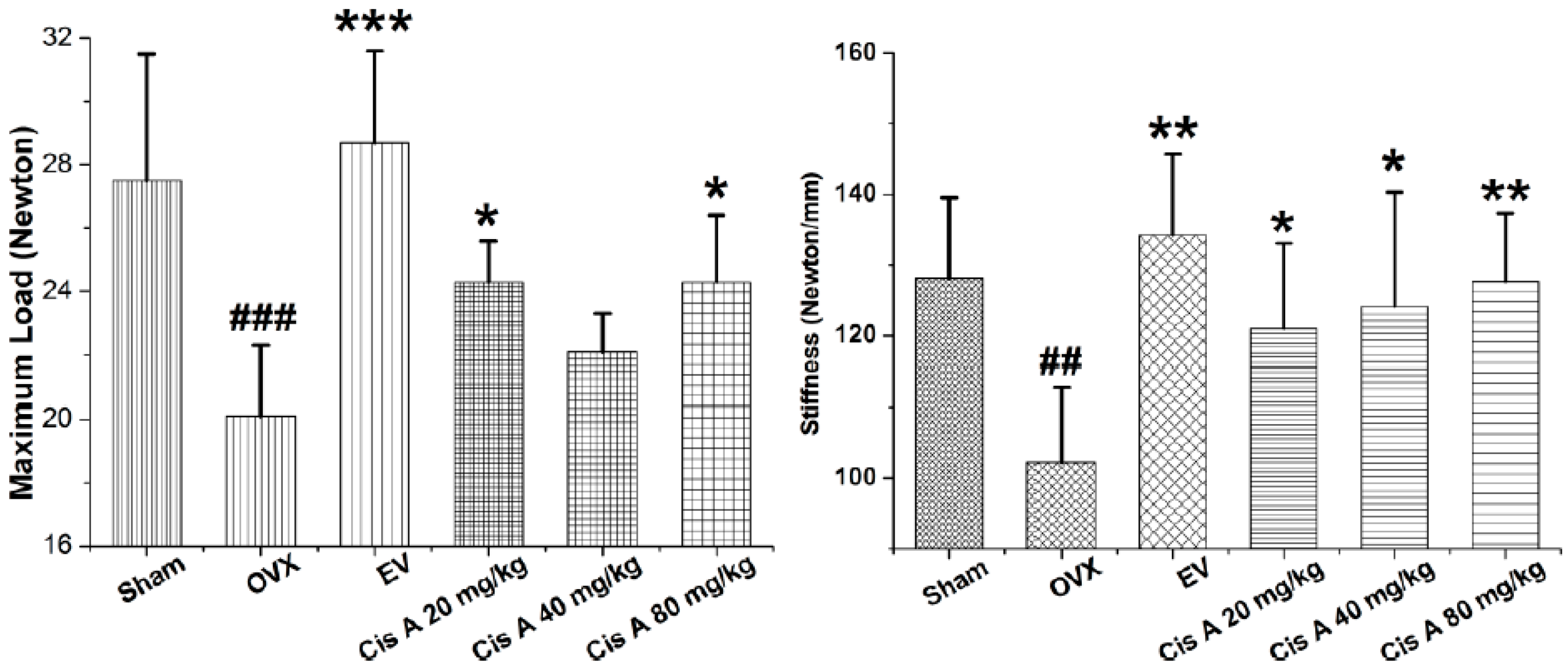
2.1.1. Effects of Cis A on Bone Three-Point Bending Testing
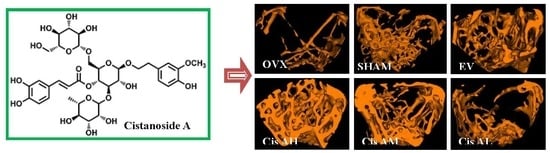
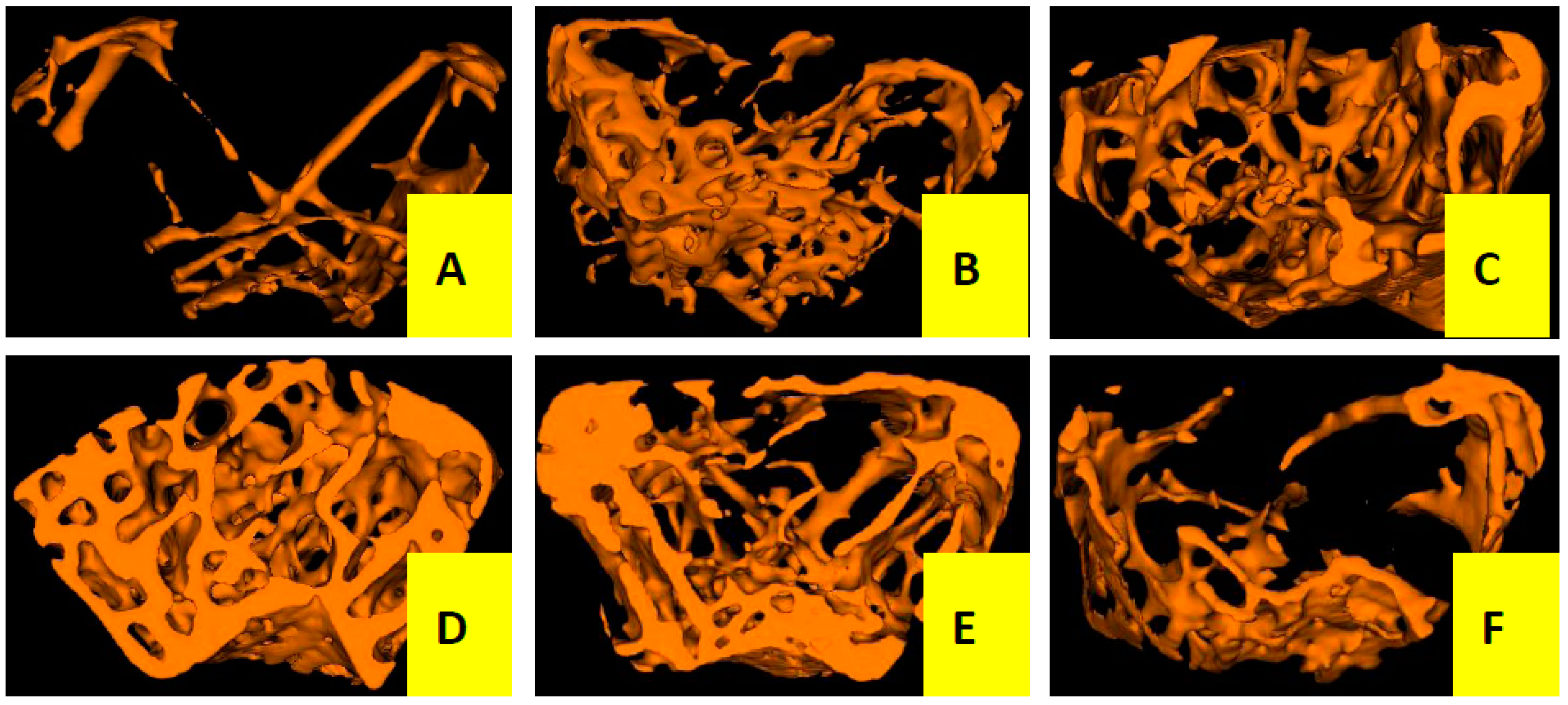
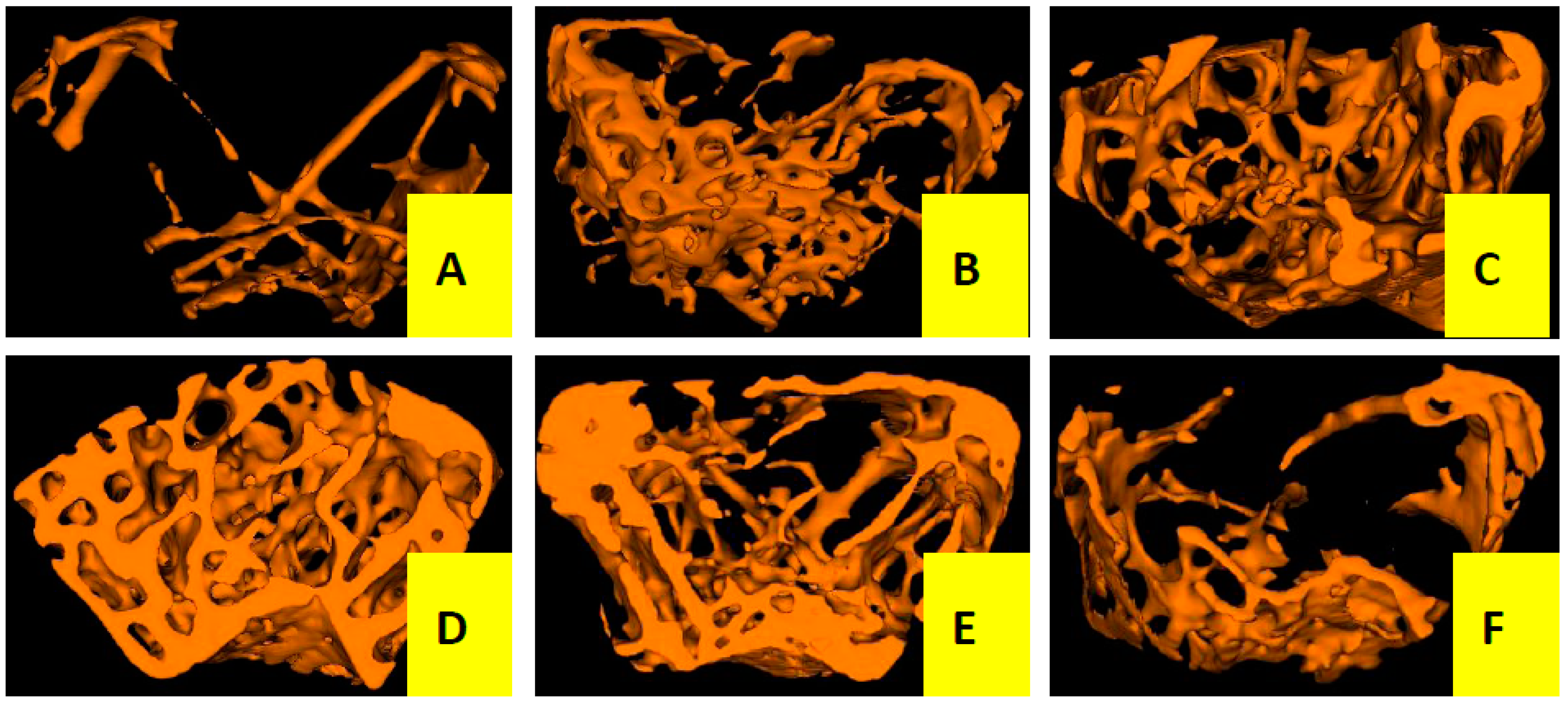
2.1.2. Effects of Cis A on Bone Microarchitecture
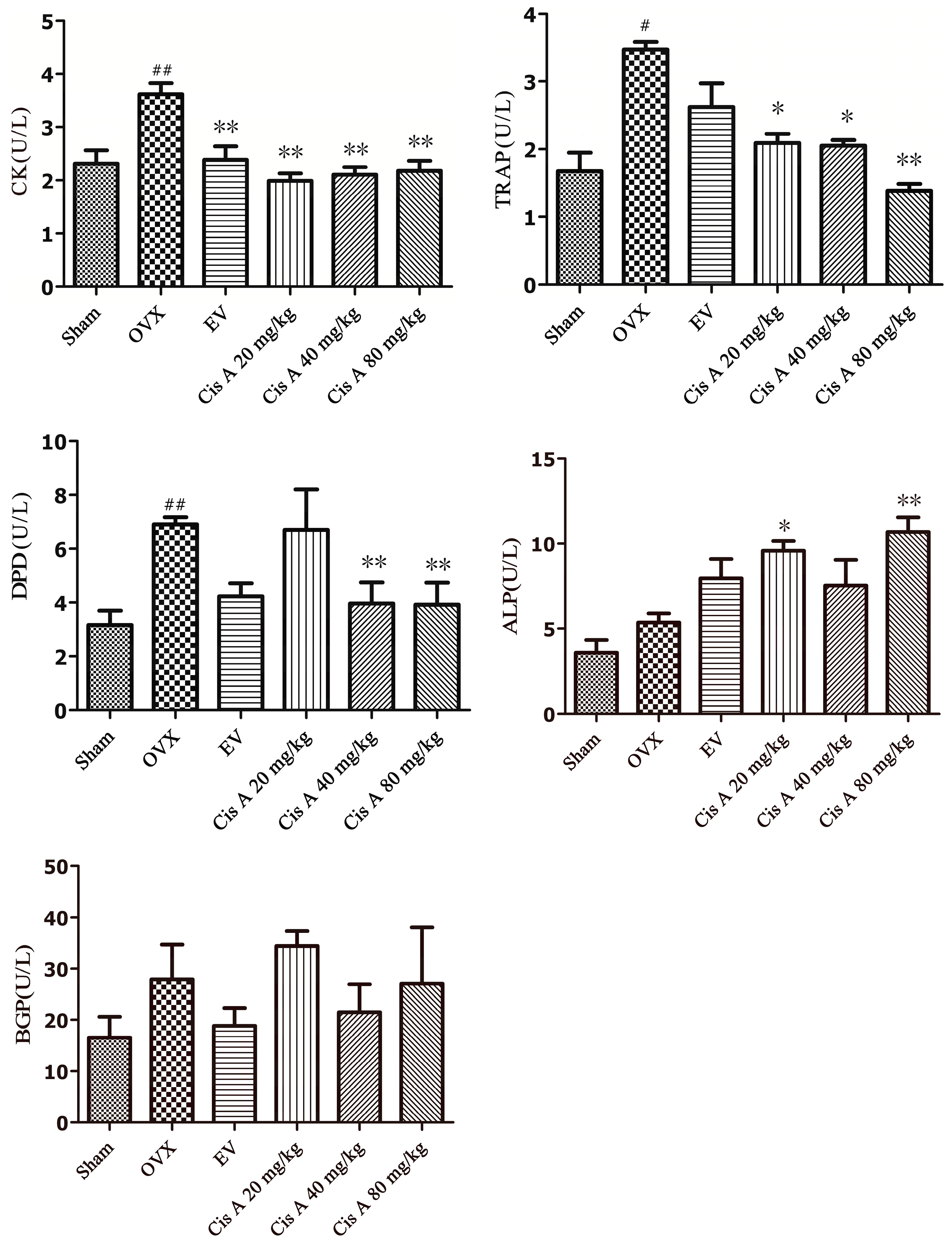
2.1.3. Effects of Cis A on Both the Bone Formation and Resorption Markers
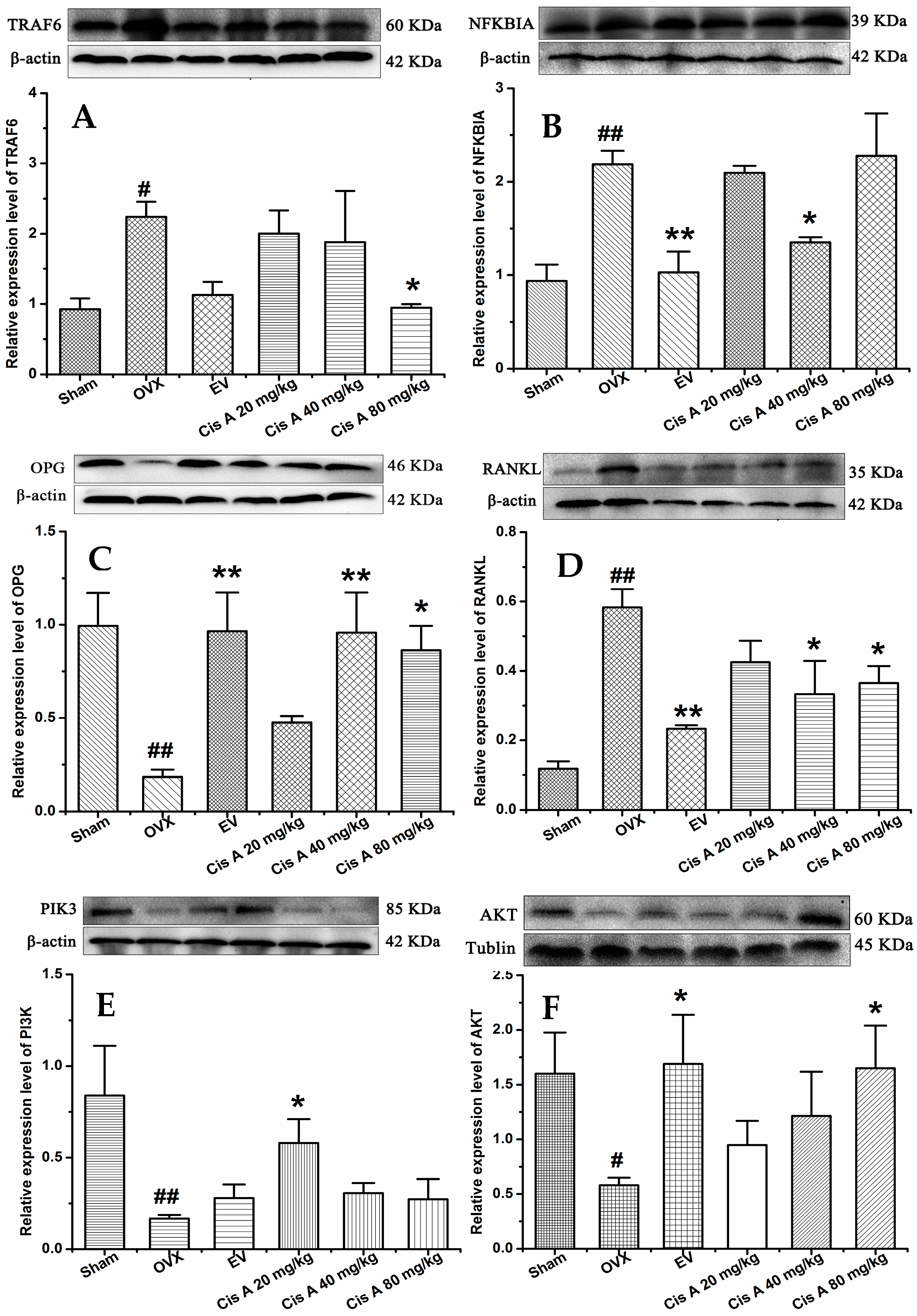
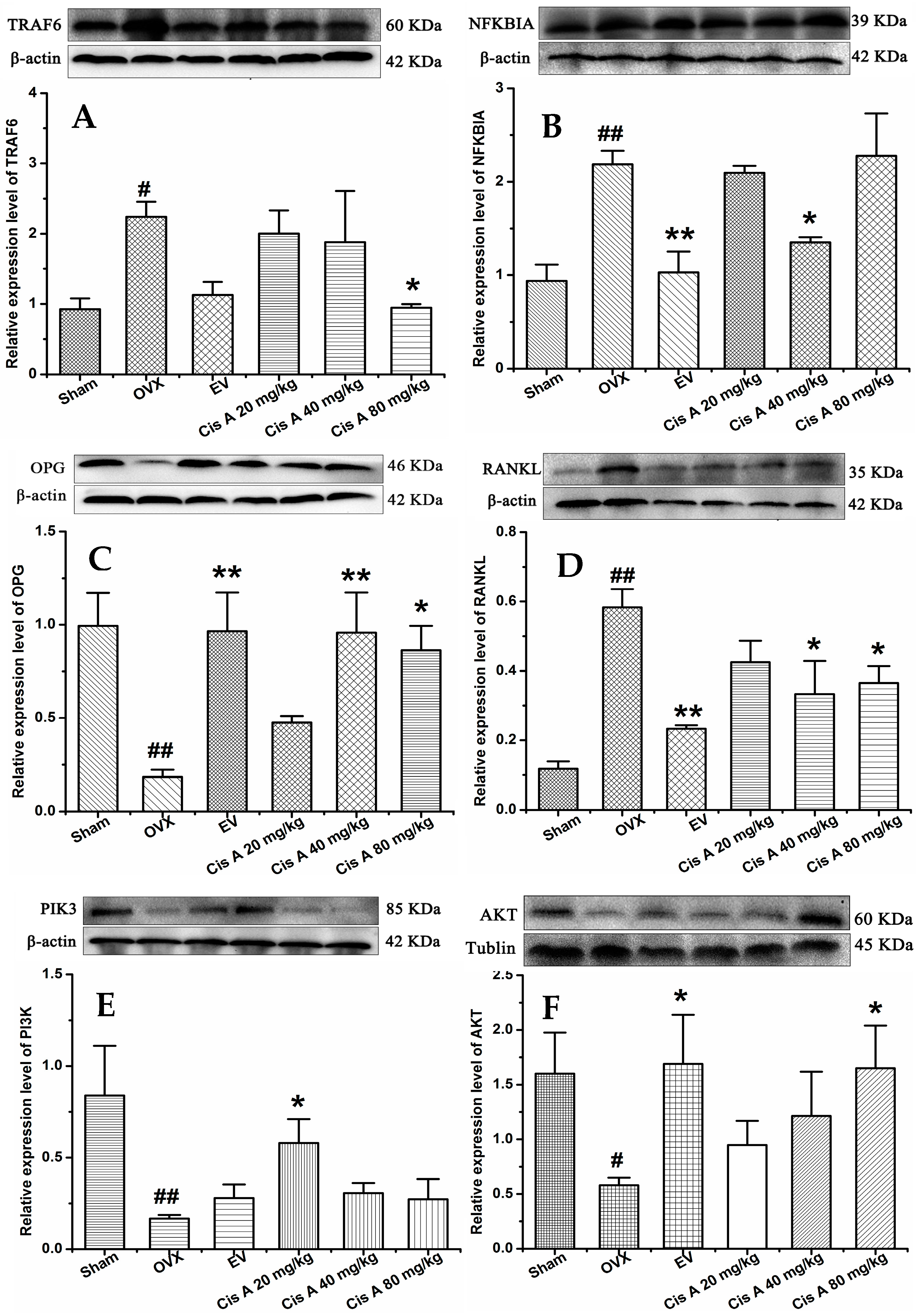
2.1.4. Effects of Cis A on Protein Expression Levels of TRAF6, NF-κB PI3K, Akt, OPG and RANKL
2.2. Discussion
3. Materials and Methods
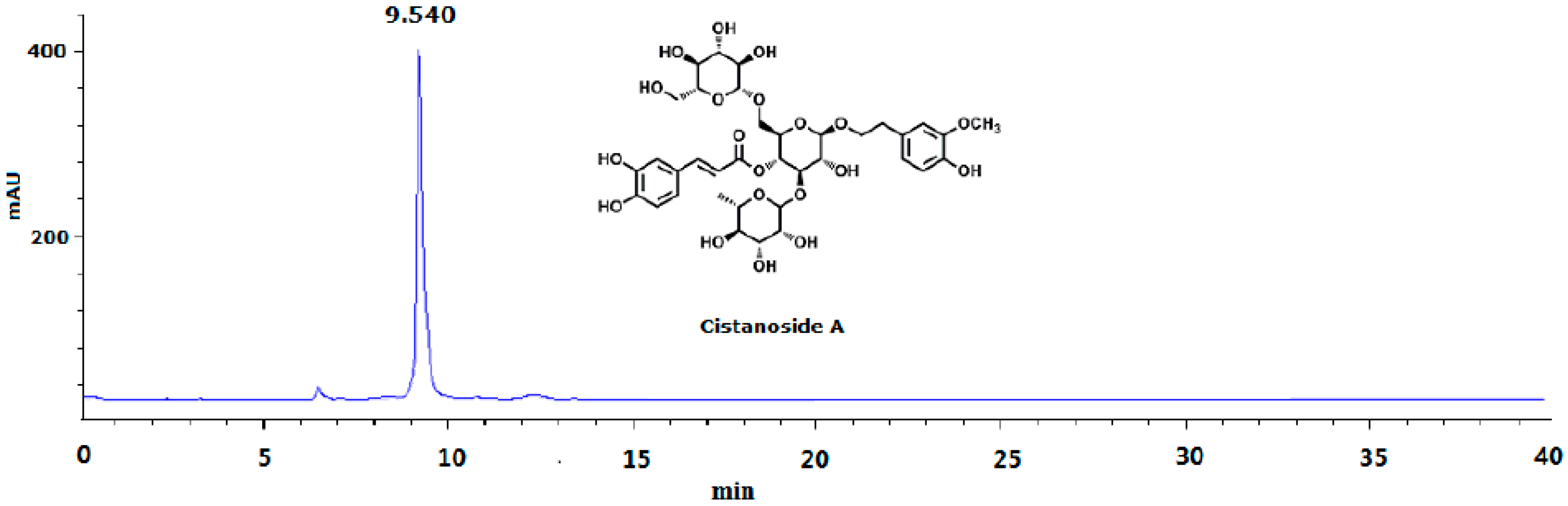
3.1. Plant Materials, Preparation and Determination
3.2. Chemicals and Solvents
3.3. Animal Experiments
3.4. Three-Point Bending Testing
3.5. Micro-Computed Tomography Analysis
3.6. Biochemical Parameters’ Analysis
3.7. Western Blot Analysis
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lane, N.E. Epidemiology, etiology, and diagnosis of osteoporosis. Am. J. Obstet. Gynecol 2006, 194 (Suppl. S2), S3–S11. [Google Scholar] [CrossRef] [PubMed]
- Kanis, J.A.; McCloskey, E.V.; Harvey, N.C.; Johansson, H.; Leslie, W.D. Intervention Thresholds and the Diagnosis of Osteoporosis. J. Bone Miner. Res. 2015, 30, 1747–1753. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Li, J.; Jia, X. The antiosteoporotic activity of central-icaritin (CIT) on bone metabolism of ovariectomized rats. Molecules 2014, 19, 18690–18704. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zeng, L.; Xie, J.; Yue, Z.; Deng, H.; Ma, X.; Zheng, C.; Wu, X.; Luo, J.; Liu, M. Inhibition of Osteoclastogenesis and Bone Resorption in vitro andin vivo by a prenylflavonoid xanthohumol from hops. Sci. Rep. 2015, 5, 1–14. [Google Scholar]
- Barzel, U.S. Estrogens in the prevention and treatment of postmenopausal osteoporosis: A review. Am. J. Med. 1988, 85, 847–850. [Google Scholar] [CrossRef]
- Zhu, Z.; Xue, L.M.; Han, T.; Jiao, L.; Qin, L.P.; Li, Y.S.; Zheng, H.C.; Zhang, Q.Y. Antiosteoporotic effects and proteomic characterization of the target and mechanism of an Er-Xian Decoction on osteoblastic UMR-106 and osteoclasts induced from RAW264.7. Molecules 2010, 15, 4695–4710. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.B.; Zheng, C.J.; Qin, L.P.; Sun, L.N.; Han, T.; Jiao, L.; Zhang, Q.Y.; Wu, J.Z. Antiosteoporotic activity of anthraquinones from Morinda officinalis on osteoblasts and osteoclasts. Molecules 2009, 14, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Bonewald, L.F. The amazing osteocyte. J. Bone Miner. Res. 2011, 26, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Banin Hirata, B.K.; Oda, J.M.; Losi Guembarovski, R.; Ariza, C.B.; de Oliveira, C.E.; Watanabe, M.A. Molecular markers for breast cancer: prediction on tumor behavior. Dis. Markers 2014, 2014, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Alipieva, K.; Korkina, L.; Orhan, I.E.; Georgiev, M.I. Verbascoside—A review of its occurrence, (bio)synthesis and pharmacological significance. Biotechnol. Adv. 2014, 32, 1065–1076. [Google Scholar] [CrossRef] [PubMed]
- Pharmacopoeia, Editorial Committee of Chinese Pharmacopoeia. Pharmacopoeia of the People’s Republic of China; China Medical Science and Technology Press: Beijing, China, 2015. [Google Scholar]
- Zhang, H.; Xing, W.W.; Li, Y.S.; Zhu, Z.; Wu, J.Z.; Zhang, Q.Y.; Zhang, W.; Qin, L.P. Effects of a traditional Chinese herbal preparation on osteoblasts and osteoclasts. Maturitas 2008, 61, 334–339. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.-Q.; Zheng, C.-J.; Zhang, Y.; Hu, C.-L.; Bing, L.; Fu, X.-Y.; Han, L.-Y.; Xu, L.-S.; Rahman, K.; Qin, L.-P. Antiosteoporotic flavonoids from Podocarpium podocarpum. Phytochem. Lett. 2013, 6, 118–122. [Google Scholar] [CrossRef]
- Ye, Q.; Ma, X.Q.; Hu, C.L.; Lin, B.; Xu, L.S.; Zheng, C.J.; Qin, L.P. Antiosteoporotic activity and constituents of Podocarpium podocarpum. Phytomedicine 2015, 22, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Qin, G.; Wen, P.; Wang, Y.; Fu, W.; He, L.; Yao, S.; Zhao, P. Safety assessment of powdered Cistanche deserticola Y.C.Ma by a 90-day feeding test in Sprague-Dawley rats. Drug Chem. Toxicol. 2016, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.X.; Chen, G.M.; Zhao, K.T.; Chen, R.; Lin, C.F. Study on toxicity of Cistanche Deserticola. Chin. J. Health Lab. Technol. 2014, 24, 1098–1100. [Google Scholar]
- Zhang, L.; Yue, X.; Zhang, L.; Zhao, J.; Chen, Y.; Cao, Z.; Liu, Y. Anti-osteoporosis effect of Cistanche deserticola Ma extract in ovariectomized rats. Trop. J. Pharm. Res. 2016, 15, 1929–1933. [Google Scholar] [CrossRef]
- Li, T.M.; Huang, H.C.; Su, C.M.; Ho, T.Y.; Wu, C.M.; Chen, W.C.; Fong, Y.C.; Tang, C.H. Cistanche deserticola extract increases bone formation in osteoblasts. J. Pharm. Pharmacol. 2012, 64, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Yang, X.; Yang, Y.; Guo, C.; Zhang, C.; Yang, Z.; Li, P. Antiosteoporotic activity of echinacoside in ovariectomized rats. Phytomedicine 2013, 20, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Yang, Y.; Zhu, P.; Chen, W.; Qi, D.; Shi, X.; Zhang, C.; Yang, Z.; Li, P. Echinacoside promotes bone regeneration by increasing OPG/RANKL ratio in MC3T3-E1 cells. Fitoterapia 2012, 83, 1443–1450. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Q.; Kadota, S.; Tani, T.; Namba, T. Antioxidative effects of phenylethanoids from Cistanche deserticola. Biol. Pharm. Bull. 1996, 19, 1580–1585. [Google Scholar] [CrossRef] [PubMed]
- Nan, Z.D.; Zeng, K.W.; Shi, S.P.; Zhao, M.B.; Jiang, Y.; Tu, P.F. Phenylethanoid glycosides with anti-inflammatory activities from the stems of Cistanche deserticola cultured in Tarim desert. Fitoterapia 2013, 89, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Q.; Tezuka, Y.; Kaneko, T.; Li, H.; Tran, L.Q.; Hase, K.; Namba, T.; Kadota, S. Inhibition of nitric oxide by phenylethanoids in activated macrophages. Eur. J. Pharmacol. 2000, 400, 137–144. [Google Scholar] [CrossRef]
- Luo, H.; Cao, R.; Wang, L.; Zhu, L. Protective effect of Cistanchis A on ethanol-induced damage in primary cultured mouse hepatocytes. Biomed. Pharmacother. 2016, 83, 1071–1079. [Google Scholar] [CrossRef] [PubMed]
- Nian, H.; Ma, M.H.; Nian, S.S.; Xu, L.L. Antiosteoporotic activity of icariin in ovariectomized rats. Phytomedicine 2009, 16, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Swaminathan, R. Biochemical markers of bone turnover. Clin. Chim. Acta 2001, 313, 95–105. [Google Scholar] [CrossRef]
- Lim, D.W.; Kim, J.G.; Lee, Y.; Cha, S.H.; Kim, Y.T. Preventive effects of Eleutherococcus senticosus bark extract in OVX-induced osteoporosis in rats. Molecules 2013, 18, 7998–8008. [Google Scholar] [CrossRef] [PubMed]
- Bord, S.; Ireland, D.C.; Beavan, S.R.; Compston, J.E. The effects of estrogen on osteoprotegerin, RANKL, and estrogen receptor expression in human osteoblasts. Bone 2003, 32, 136–141. [Google Scholar] [CrossRef]
- Takayanagi, H. Osteoimmunology: shared mechanisms and crosstalk between the immune and bone systems. Nat. Rev. Immunol. 2007, 7, 292–304. [Google Scholar] [CrossRef] [PubMed]
- Franzoso, G.; Carlson, L.; Xing, L.; Poljak, L.; Shores, E.W.; Brown, K.D.; Leonardi, A.; Tran, T.; Boyce, B.F.; Siebenlist, U. Requirement for NF-kappaB in osteoclast and B-cell development. Gene Dev. 1997, 11, 3482–3496. [Google Scholar] [CrossRef] [PubMed]
- Tan, E.M.; Li, L.; Indran, I.R.; Chew, N.; Yong, E.L. TRAF6 mediates suppression of osteoclastogenesis and prevention of ovariectomy-induced bone loss by a novel prenylflavonoid. J. Bone Miner. Res. 2016. [Google Scholar] [CrossRef] [PubMed]
- Jiao, L.; Cao, D.P.; Qin, L.P.; Han, T.; Zhang, Q.Y.; Zhu, Z.; Yan, F. Antiosteoporotic activity of phenolic compounds from Curculigo orchioides. Phytomedicine 2009, 16, 874–881. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds Cistanoside A and acteoside are available from the authors.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | BMD (mg/cc) | BMC (mg) | TMC (mg) | TMD (mg/cc) | BVF | Tb.Sp (mm) | Tb.N (1/mm) | Tb.Th (mm) |
|---|---|---|---|---|---|---|---|---|
| Sham | 719 ± 79 | 2.07 ± 0.25 | 1.10 ± 0.35 | 1780 ± 64 | 0.22 ± 0.07 | 0.26 ± 0.11 | 3.34 ± 0.91 | 0.06 ± 0.01 |
| OVX | 391 ± 74 # | 0.70 ± 0.43 ## | 0.17 ± 0.19 # | 1717 ± 208 | 0.04 ± 0.03 # | 1.29 ± 0.44 ### | 0.81 ± 0.27 ### | 0.04 ± 0.01 |
| Cis A 80 mg/kg | 915 ± 298 ** | 2.64 ± 0.88 *** | 1.75 ± 0.82 ** | 1773 ± 125 | 0.34 ± 0.13 *** | 0.15 ± 0.05 *** | 4.42 ± 0.62 *** | 0.07 ± 0.02 * |
| Cis A 40 mg/kg | 732 ± 32 * | 2.00 ± 0.13 * | 1.08 ± 0.06 * | 1791 ± 80 | 0.22 ± 0.01 * | 0.24 ± 0.01 *** | 3.38 ± 0.22 *** | 0.06 ± 0.01 |
| Cis A 20 mg/kg | 689 ± 71 | 1.98 ± 0.20 * | 1.00 ± 0.13 | 1708 ± 35 | 0.20 ± 0.03 * | 0.27 ± 0.04 *** | 3.02 ± 0.29 ** | 0.07 ± 0.01 |
| EV | 1057 ± 335 *** | 3.03 ± 0.95 *** | 2.08 ± 0.90 *** | 2090 ± 299 * | 0.34 ± 0.15 *** | 0.22 ± 0.13 *** | 3.55 ± 1.38 *** | 0.10 ± 0.03 ** |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, X.; Zhang, Z.; Wang, W.; Yao, H.; Ma, X. Therapeutic Effect of Cistanoside A on Bone Metabolism of Ovariectomized Mice. Molecules 2017, 22, 197. https://doi.org/10.3390/molecules22020197
Xu X, Zhang Z, Wang W, Yao H, Ma X. Therapeutic Effect of Cistanoside A on Bone Metabolism of Ovariectomized Mice. Molecules. 2017; 22(2):197. https://doi.org/10.3390/molecules22020197
Chicago/Turabian StyleXu, Xiaoxue, Zhuanzhuan Zhang, Wenping Wang, Huiqin Yao, and Xueqin Ma. 2017. "Therapeutic Effect of Cistanoside A on Bone Metabolism of Ovariectomized Mice" Molecules 22, no. 2: 197. https://doi.org/10.3390/molecules22020197
APA StyleXu, X., Zhang, Z., Wang, W., Yao, H., & Ma, X. (2017). Therapeutic Effect of Cistanoside A on Bone Metabolism of Ovariectomized Mice. Molecules, 22(2), 197. https://doi.org/10.3390/molecules22020197