Apoptotic Effects of Chrysin in Human Cancer Cell Lines

Abstract
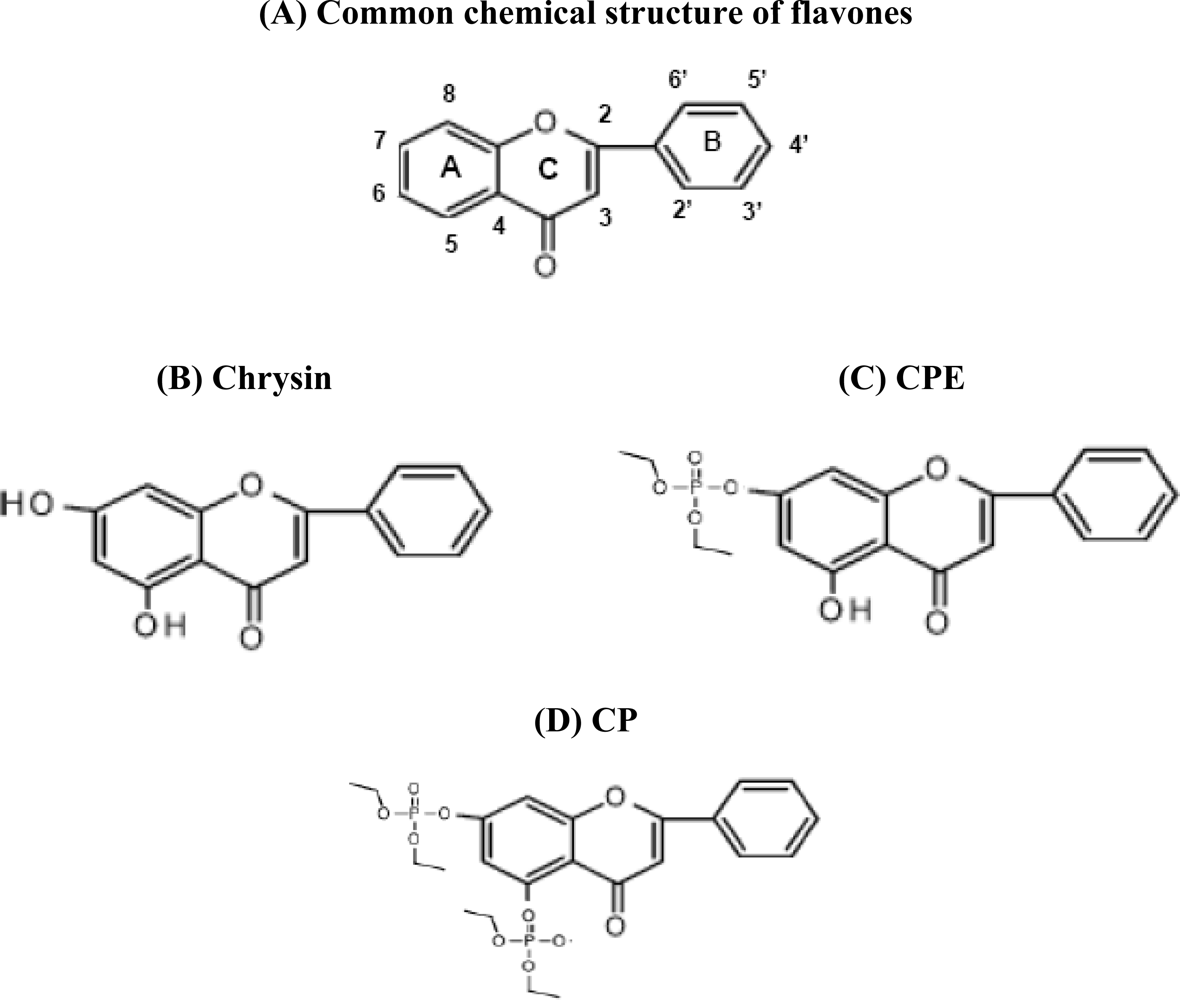
:1. Introduction of Flavonoids
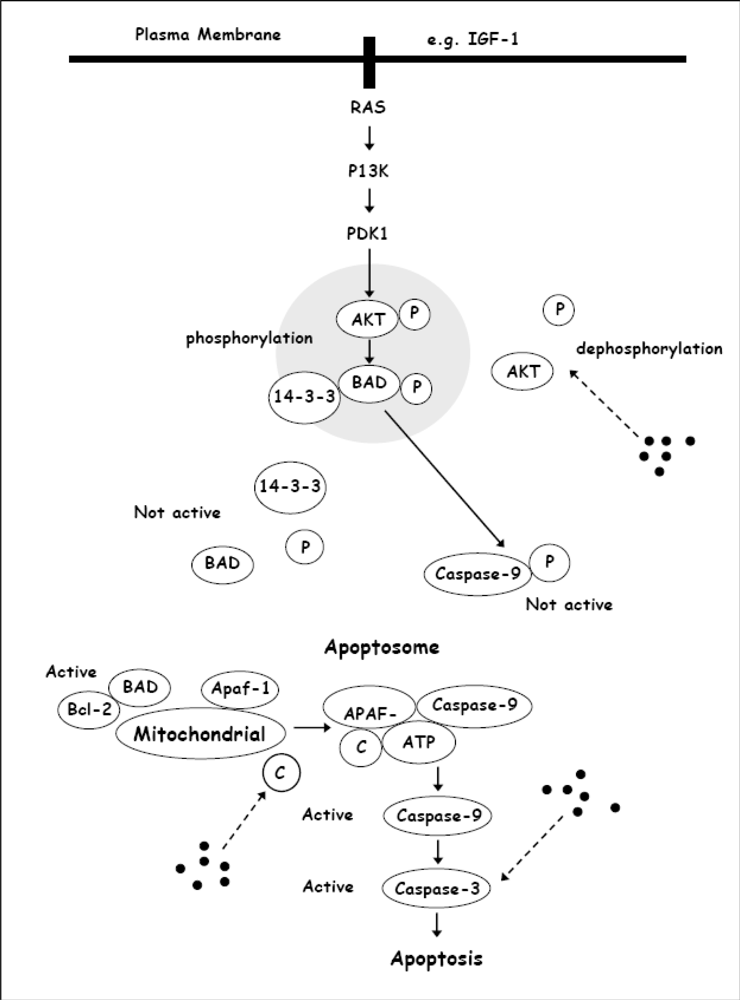
2. Chrysin Inhibits Proliferation and Induces Apoptosis in HeLa Cells
3. Chrysin Induces Apoptosis in Leukemia Cells
4. Cytotoxicity of Chrysin in Esophageal Squamous Carcinoma
5. Chrysin in Malignant Glioma, Breast Carcinoma, Prostate and Other Human Cancers
6. Conclusions
Acknowledgments
References and Notes
- Robards, K; Antolovich, M. Analytical chemistry of fruit bioflavonoids: A review. Analyst 1997, 122, 11–34. [Google Scholar]
- Pietta, PG. Flavonoids as anti-oxidants. J. Nat. Prod 2000, 63, 1035–1042. [Google Scholar]
- Nijveldt, RJ; van Nood, E; van Hoorn, DE; Boelens, PG; van Norren, K; van Leeuwen, PA. Flavonoids: A review of probable mechanisms of action and potential applications. Am. J. Clin. Nutr 2001, 74, 418–425. [Google Scholar]
- Kale, A; Gawande, S; Kotwal, S. Cancer phytotherapeutics: role for flavonoids at the cellular level. Phytother. Res 2008, 22, 567–577. [Google Scholar]
- Awad, R; Arnason, JT; Trudeau, V; Bergeron, C; Budzinski, JW; Foster, BC; Merali, Z. Phytochemical and biological analysis of skullcap (Scutellaria lateriflora L.): A medicinal plant with anxiolytic properties. Phytomedicine 2003, 10, 640–649. [Google Scholar]
- Zheng, X; Meng, WD; Xu, YY; Cao, JG; Qing, FL. Synthesis and anti-cancer effect of chrysin derivatives. Bioorg. Med. Chem. Lett 2003, 13, 881–884. [Google Scholar]
- Ernst, E. Herbal remedies for anxiety - a systematic review of controlled clinical trials. Phytomedicine 2006, 13, 205–208. [Google Scholar]
- Huang, WH; Lee, AR; Yang, CH. Anti-oxidative and anti-inflammatory activities of polyhydroxyflavonoids of Scutellaria baicalensis GEORGI. Biosci. Biotechnol. Biochem 2006, 70, 2371–2380. [Google Scholar]
- Scheck, AC; Perry, K; Hank, NC; Clark, WD. Anti-cancer activity of extracts derived from the mature roots of Scutellaria baicalensis on human malignant brain tumor cells. BMC Complement Altern. Med 2006, 6, 27–35. [Google Scholar]
- Cole, IB; Cao, J; Alan, AR; Saxena, PK; Murch, SJ. Comparisons of Scutellaria baicalensis, Scutellaria lateriflora and Scutellaria racemosa: Genome size, anti-oxidant potential and phytochemistry. Planta Med 2008, 74, 474–481. [Google Scholar]
- Parajuli, P; Joshee, N; Rimando, AM; Mittal, S; Yadav, AK. In vitro anti-tumor mechanisms of various Scutellaria extracts and constituent flavonoids. Planta Med 2009, 75, 41–48. [Google Scholar]
- Duthie, G; Crozier, A. Plant-derived phenolic anti-oxidants. Curr. Opin. Clin. Nutr. Metab. Care 2000, 3, 447–451. [Google Scholar]
- Birt, DF; Hendrich, S; Wang, W. Dietary agents in cancer prevention: Flavonoids and isoflavonoids. Pharmacol. Ther 2001, 90, 157–177. [Google Scholar]
- Kim, HK; Cheon, BS; Kim, YH; Kim, SY; Kim, HP. Effects of naturally occurring flavonoids on nitric oxide production in the macrophage cell line RAW 264.7 and their structure-activity relationships. Biochem. Pharmacol 1999, 58, 759–765. [Google Scholar]
- Raso, GM; Meli, R; Di Carlo, G; Pacilio, M; Di Carlo, R. Inhibition of inducible nitric oxide synthase and cyclooxygenase-2 expression by flavonoids in macrophage J774A.1. Life Sci 2001, 68, 921–931. [Google Scholar]
- Lindenmeyer, F; Li, H; Menashi, S; Soria, C; Lu, H. Apigenin acts on the tumor cell invasion process and regulates protease production. Nutr. Cancer 2001, 39, 139–147. [Google Scholar]
- Kuo, ML; Yang, NC. Reversion of v-H-ras-transformed NIH 3T3 cells by apigenin through inhibiting mitogen activated protein kinase and its downstream oncogenes. Biochem. Biophys. Res. Commun 1995, 212, 767–775. [Google Scholar]
- Sanderson, JT; Hordijk, J; Denison, MS; Springsteel, MF; Nantz, MH; van den Berg, M. Induction and inhibition of aromatase (CYP19) activity by natural and synthetic flavonoid compounds in H295R human adrenocortical carcinoma cells. Toxicol. Sci 2004, 82, 70–79. [Google Scholar]
- Weng, MS; Ho, YS; Lin, JK. Chrysin induces G1 phase cell cycle arrest in C6 glioma cells through inducing p21Waf1/Cip1 expression: Involvement of p38 mitogen-activated protein kinase. Biochem. Pharmacol 2005, 69, 1815–1827. [Google Scholar]
- Critchfield, JW; Butera, ST; Folks, TM. Inhibition of HIV activation in latently infected cells by flavonoid compounds. AIDS Res. Hum. Retroviruses 1996, 12, 39–46. [Google Scholar]
- Cho, H; Yun, CW; Park, WK; Kong, JY; Kim, KS; Park, Y; Lee, S; Kim, BK. Modulation of the activity of pro-inflammatory enzymes, COX-2 and iNOS, by chrysin derivatives. Pharmacol. Res 2004, 49, 37–43. [Google Scholar]
- Woodman, OL; Chan, E. Vascular and anti-oxidant actions of flavonols and flavones. Clin. Exp. Pharmacol. Physiol 2004, 31, 786–790. [Google Scholar]
- Gewies, A. Introduction to apoptosis. ApoReview 2003, 1–26. [Google Scholar]
- Zhang, T; Chen, X; Qu, L; Wu, J; Cui, R; Zhao, Y. Chrysin and its phosphate ester inhibit cell proliferation and induce apoptosis in Hela cells. Bioorg. Med. Chem 2004, 12, 6097–6105. [Google Scholar]
- von Brandenstein, MG; Ngum Abety, A; Depping, R; Roth, T; Koehler, M; Dienes, HP; Fries, JW. A p38–p65 transcription complex induced by endothelin-1 mediates signal transduction in cancer cells. Biochim. Biophys. Acta 2008, 1783, 1613–1622. [Google Scholar]
- Monasterio, A; Urdaci, MC; Pinchuk, IV; Lopez-Moratalla, N; Martinez-Irujo, JJ. Flavonoids induce apoptosis in human leukemia U937 cells through caspase- and caspase-calpain-dependent pathways. Nutr. Cancer 2004, 50, 90–100. [Google Scholar]
- Woo, KJ; Jeong, YJ; Park, JW; Kwon, TK. Chrysin-induced apoptosis is mediated through caspase activation and Akt inactivation in U937 leukemia cells. Biochem. Biophys. Res. Commun 2004, 325, 1215–1222. [Google Scholar]
- Woo, KJ; Yoo, YH; Park, JW; Kwon, TK. Bcl-2 attenuates anti-cancer agents-induced apoptosis by sustained activation of Akt/protein kinase B in U937 cells. Apoptosis 2005, 10, 1333–1343. [Google Scholar]
- Lee, SJ; Yoon, JH; Song, KS. Chrysin inhibited stem cell factor (SCF)/c-Kit complex-induced cell proliferation in human myeloid leukemia cells. Biochem. Pharmacol 2007, 74, 215–225. [Google Scholar]
- Ramos, AM; Aller, P. Quercetin decreases intracellular GSH content and potentiates the apoptotic action of the anti-leukemic drug arsenic trioxide in human leukemia cell lines. Biochem. Pharmacol 2008, 75, 1912–1923. [Google Scholar]
- Zhang, Q; Zhao, XH; Wang, ZJ. Flavones and flavonols exert cytotoxic effects on a human oesophageal adenocarcinoma cell line (OE33) by causing G2/M arrest and inducing apoptosis. Food Chem. Toxicol 2008, 46, 2042–2053. [Google Scholar]
- Zhang, Q; Zhao, XH; Wang, ZJ. Cytotoxicity of flavones and flavonols to a human esophageal squamous cell carcinoma cell line (KYSE-510) by induction of G2/M arrest and apoptosis. Toxicol. In Vitro 2009, 23, 797–807. [Google Scholar]
- Wang, HW; Lin, CP; Chiu, JH; Chow, KC; Kuo, KT; Lin, CS; Wang, LS. Reversal of inflammation-associated dihydrodiol dehydrogenases (AKR1C1 and AKR1C2) overexpression and drug resistance in nonsmall cell lung cancer cells by wogonin and chrysin. Int. J. Cancer 2007, 120, 2019–2027. [Google Scholar]
- Wang, W; VanAlstyne, PC; Irons, KA; Chen, S; Stewart, JW; Birt, DF. Individual and interactive effects of apigenin analogs on G2/M cell-cycle arrest in human colon carcinoma cell lines. Nutr. Cancer 2004, 48, 106–114. [Google Scholar]
- Li, X; Huang, Q; Ong, CN; Yang, XF; Shen, HM. Chrysin sensitizes tumor necrosis factor-alpha-induced apoptosis in human tumor cells via suppression of nuclear factor–κB. Cancer Lett 2010, 293, 109–116. [Google Scholar]
- Roberts, R. Apoptosis in Toxicology; Taylor & Francis: London, UK, 2000. [Google Scholar]
- Kelekar, A; Chang, BS; Harlan, JE; Fesik, SW; Thompson, CB. Bad is a BH3 domain-containing protein that forms an inactivating dimer with Bcl-XL. Mol. Cell Biol 1997, 17, 7040–7046. [Google Scholar]
- Pelengaris, S; Khan, M; Evan, G. c-MYC: More than just a matter of life and death. Nat. Rev. Cancer 2002, 2, 764–776. [Google Scholar]
- Debatin, KM. Apoptosis pathways in cancer and cancer therapy. Cancer Immunol. Immunother 2004, 53, 153–159. [Google Scholar]
- Yoshida, K; Hirose, Y; Tanaka, T; Yamada, Y; Kuno, T; Kohno, H; Katayama, M; Qiao, Z; Sakata, K; Sugie, S; Shibata, T; Mori, H. Inhibitory effects of troglitazone, a peroxisome proliferator-activated receptor gamma ligand, in rat tongue carcinogenesis initiated with 4-nitroquinoline 1-oxide. Cancer Sci 2003, 94, 365–371. [Google Scholar]
- Cogulu, O; Biray, C; Gunduz, C; Karaca, E; Aksoylar, S; Sorkun, K; Salih, B; Ozkinay, F. Effects of Manisa propolis on telomerase activity in leukemia cells obtained from the bone marrow of leukemia patients. Int. J. Food Sci. Nutr 2009, 60, 601–605. [Google Scholar]
- Josipovic, P; Orsolic, N. Cytotoxicity of polyphenolic/flavonoid compounds in a leukaemia cell culture. Arh. Hig. Rada. Toksikol 2008, 59, 299–308. [Google Scholar]
- Ozmen, A; Madlener, S; Bauer, S; Krasteva, S; Vonach, C; Giessrigl, B; Gridling, M; Viola, K; Stark, N; Saiko, P; Michel, B; Fritzer-Szekeres, M; Szekeres, T; Askin-Celik, T; Krenn, L; Krupitza, G. In vitro anti-leukemic activity of the ethno-pharmacological plant Scutellaria orientalis ssp. carica endemic to western Turkey. Phytomedicine 2010, 17, 55–62. [Google Scholar]
- Chang, H; Mi, MT; Gu, YY; Yuan, JL; Ling, WH; Lin, H. Effects of flavonoids with different structures on proliferation of leukemia cell line HL-60. Chin. J. Cancer 2007, 26, 1309–1314. [Google Scholar]
- Woo, KJ; Jeong, YJ; Inoue, H; Park, JW; Kwon, TK. Chrysin suppresses lipopolysaccharide-induced cyclooxygenase-2 expression through the inhibition of nuclear factor for IL-6 (NF-IL6) DNA-binding activity. FEBS Lett 2005, 579, 705–711. [Google Scholar]
- Li, HZ; Cao, JG; Deng, YA; Xu, JH; Xie, WY. Induction of apoptosis of human ovarian cancer CoC1 cells by 5-allyl-7-gen-difluoromethylenechrysin through activation of peroxisome-proliferator activated receptor-gamma. Chin. Med. Assoc. J 2007, 87, 2914–2918. [Google Scholar]
- Ai, XH; Zheng, X; Tang, XQ; Sun, L; Zhang, YQ; Qin, Y; Liu, HQ; Xia, H; Cao, JG. Induction of apoptosis of human gastric carcinoma SGC-7901 cell line by 5, 7-dihydroxy-8-nitrochrysin in vitro. World J. Gastroenterol 2007, 13, 3824–3828. [Google Scholar]



{kind=link}
{kind=link}
| Cancer type | Reference | Effect and molecular mechanism |
|---|---|---|
| Cervical cancer | [24] | Chrysin (IC50 = 14.2 μM) inhibited proliferation and induced apoptosis in HeLa cells, though the effects were not as potent as those of its synthetic derivative compounds. |
| [25] | Chrysin (30 μM) potentially induced p38 and NFkappaB/p65 activation in HeLa cells. | |
| Leukemia | [26] | Chrysin (IC50 = 16 μM) showed to be the most potent flavonoid to reduce cell viability and induce apoptotic DNA fragmentation in U937 cells. |
| [27,28] | Chrysin induced apoptosis in Bcl-2 overexpressing U937 leukemia cells, was associated with activation of caspase-3 and PLC-γ1 degradation. The induction of apoptosis was accompanied by down-regulation of XIAP and inactivation of Akt. | |
| [29] | Chrysin had the ability to abolish SCF/c-Kit signaling by inhibiting the PI3K pathway in MO7e, myeloid leukemia cells. | |
| [30] | Chrysin, alone or in combination with other compounds, decreased Akt phosphorylation and potentially caused mitochondrial dysfunction in THP-1 and HL-60 leukemia cells. | |
| Esophageal squamous carcinoma | [31,32] | Chrysin (IC50 = 63 μM) induced cytotoxicity in KYSE-510 cells in dose- and time-dependent manners. |
| Malignant glioma, breast carcinoma, prostate cancer | [11] | Chrysin (100 μM) showed dose-dependent inhibition of U87-MG, MDA-MB-231, U-251 and PC3 proliferation, and displayed apoptotic activity in U87-MG cells. However, the study did not report details about the apoptotic activity of chrysin in U-251, MDA-MB-231 and PC3 cells. |
| NSCLC | [33] | Chrysin and wogonin showed to have potential as adjuvant therapy for drug-resistant NSCLC, especially in patients with AKR1C1/1C2 overexpression where IL-6-induced AKR1C1/1C2 overexpression and drug resistance could be inhibited by these flavonoids in H23 cells. |
| Colon cancer | [34] | Chrysin caused the SW480 cells in cell-cycle arrest at the G2/M phase in a dose-dependent manner. |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Khoo, B.Y.; Chua, S.L.; Balaram, P. Apoptotic Effects of Chrysin in Human Cancer Cell Lines. Int. J. Mol. Sci. 2010, 11, 2188-2199. https://doi.org/10.3390/ijms11052188
Khoo BY, Chua SL, Balaram P. Apoptotic Effects of Chrysin in Human Cancer Cell Lines. International Journal of Molecular Sciences. 2010; 11(5):2188-2199. https://doi.org/10.3390/ijms11052188
Chicago/Turabian StyleKhoo, Boon Yin, Siang Ling Chua, and Prabha Balaram. 2010. "Apoptotic Effects of Chrysin in Human Cancer Cell Lines" International Journal of Molecular Sciences 11, no. 5: 2188-2199. https://doi.org/10.3390/ijms11052188