Temporal and Spatial Expression of Muc1 During Implantation in Sows

Abstract
:1. Introduction
2. Results and Discussion
2.1. Endometrial Surface Morphological Changes on Day 13 of Pregnancy
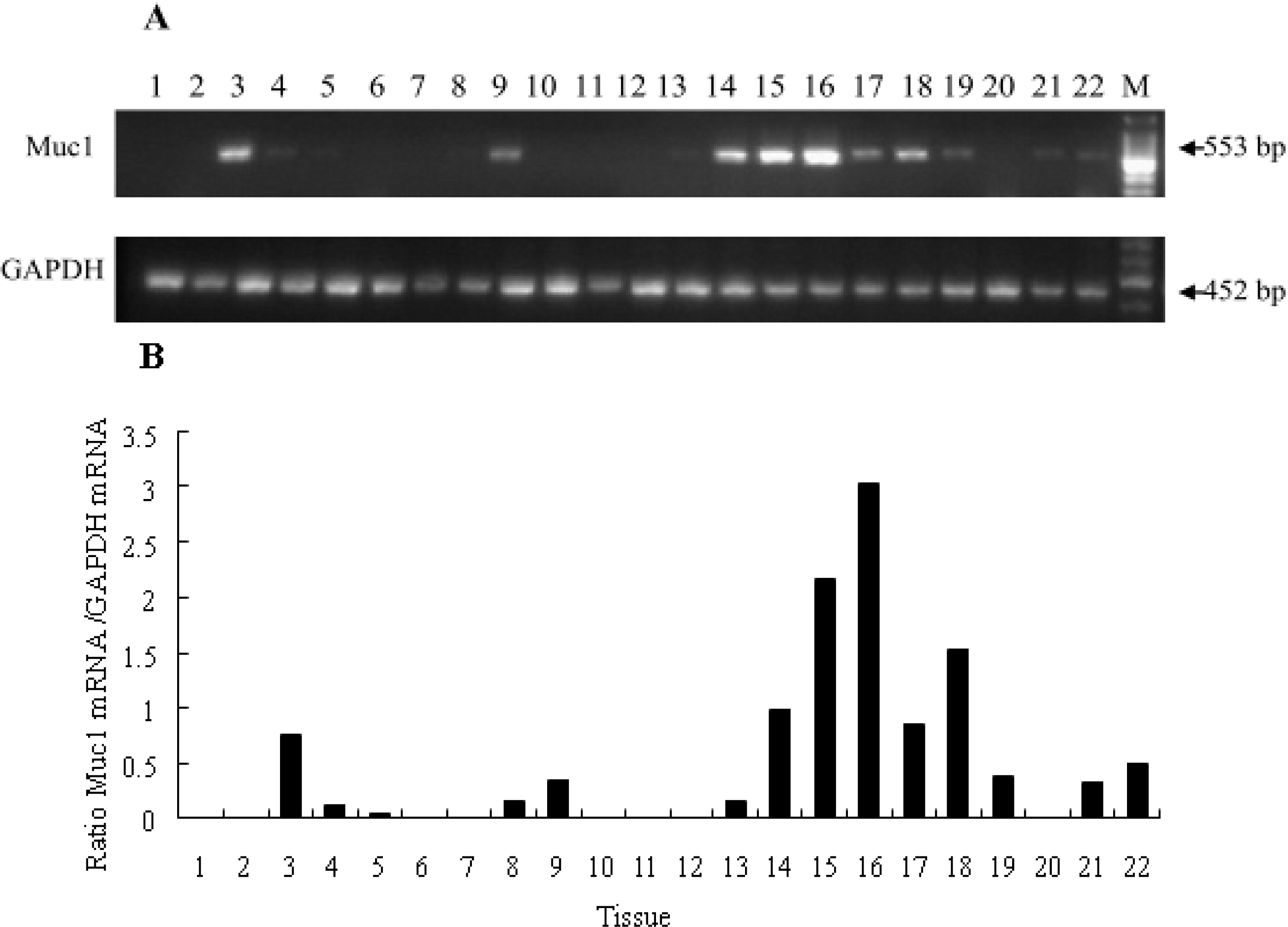
2.2. Tissue Distribution of Muc-1 mRNA in a Sow
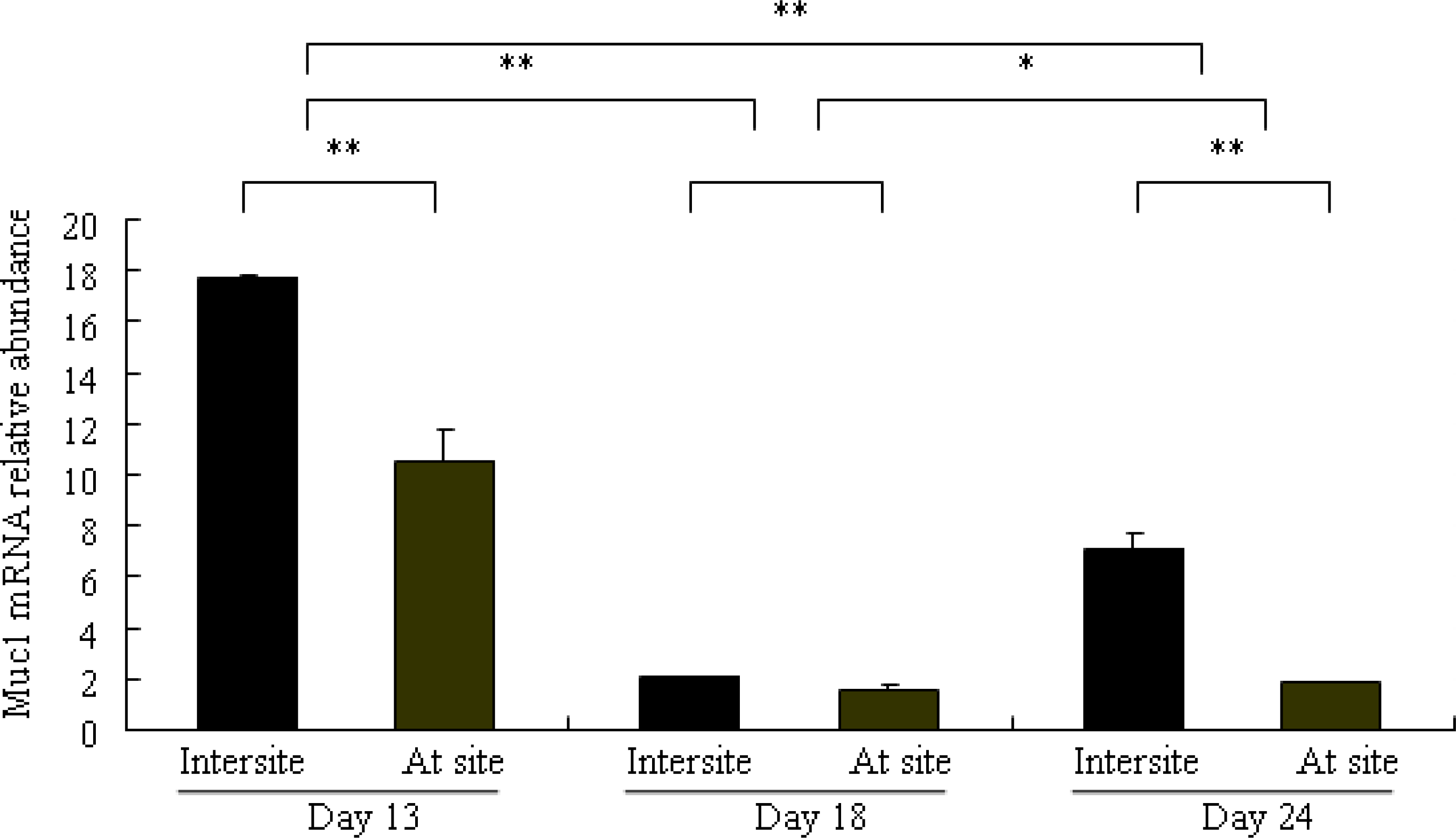
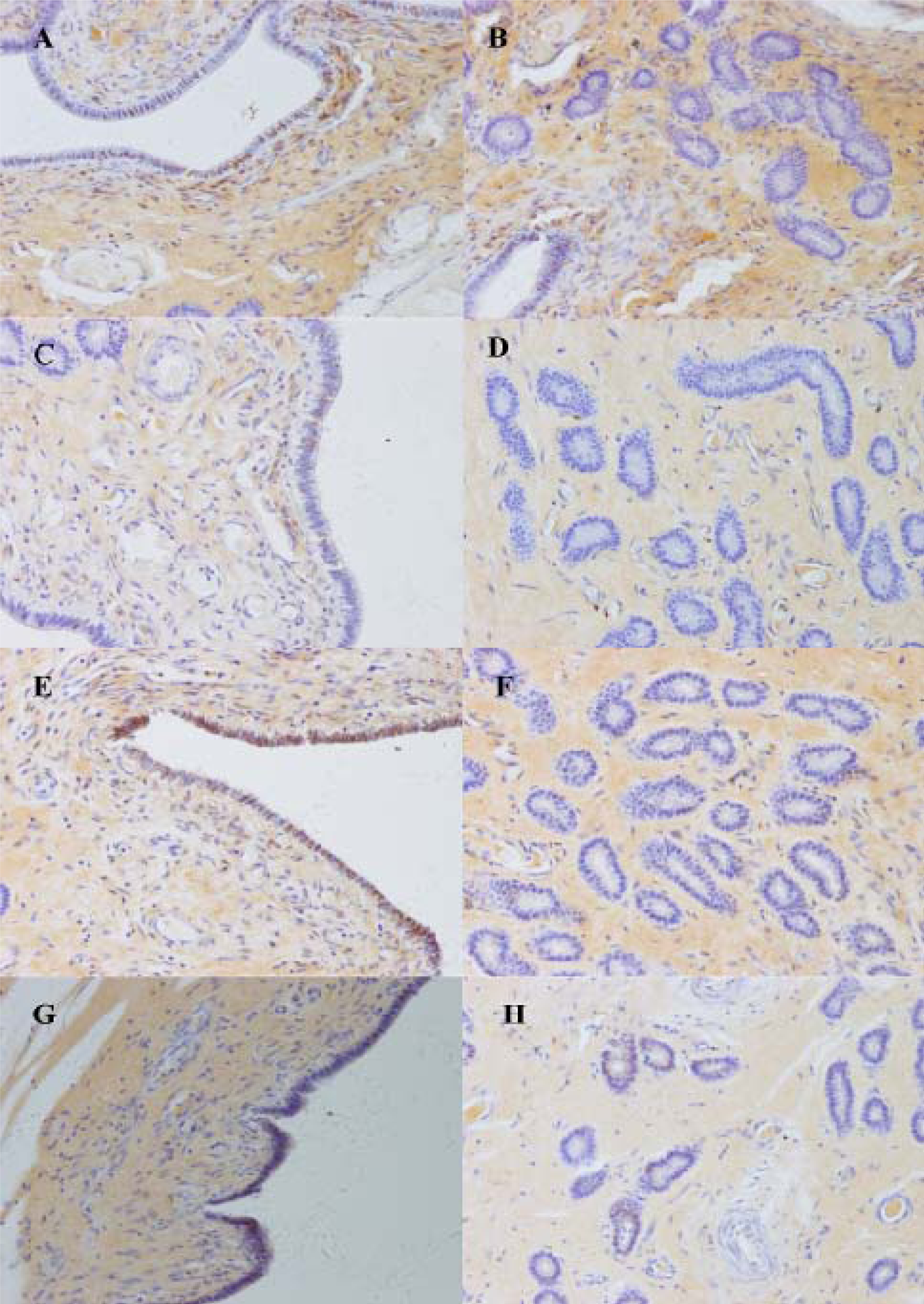
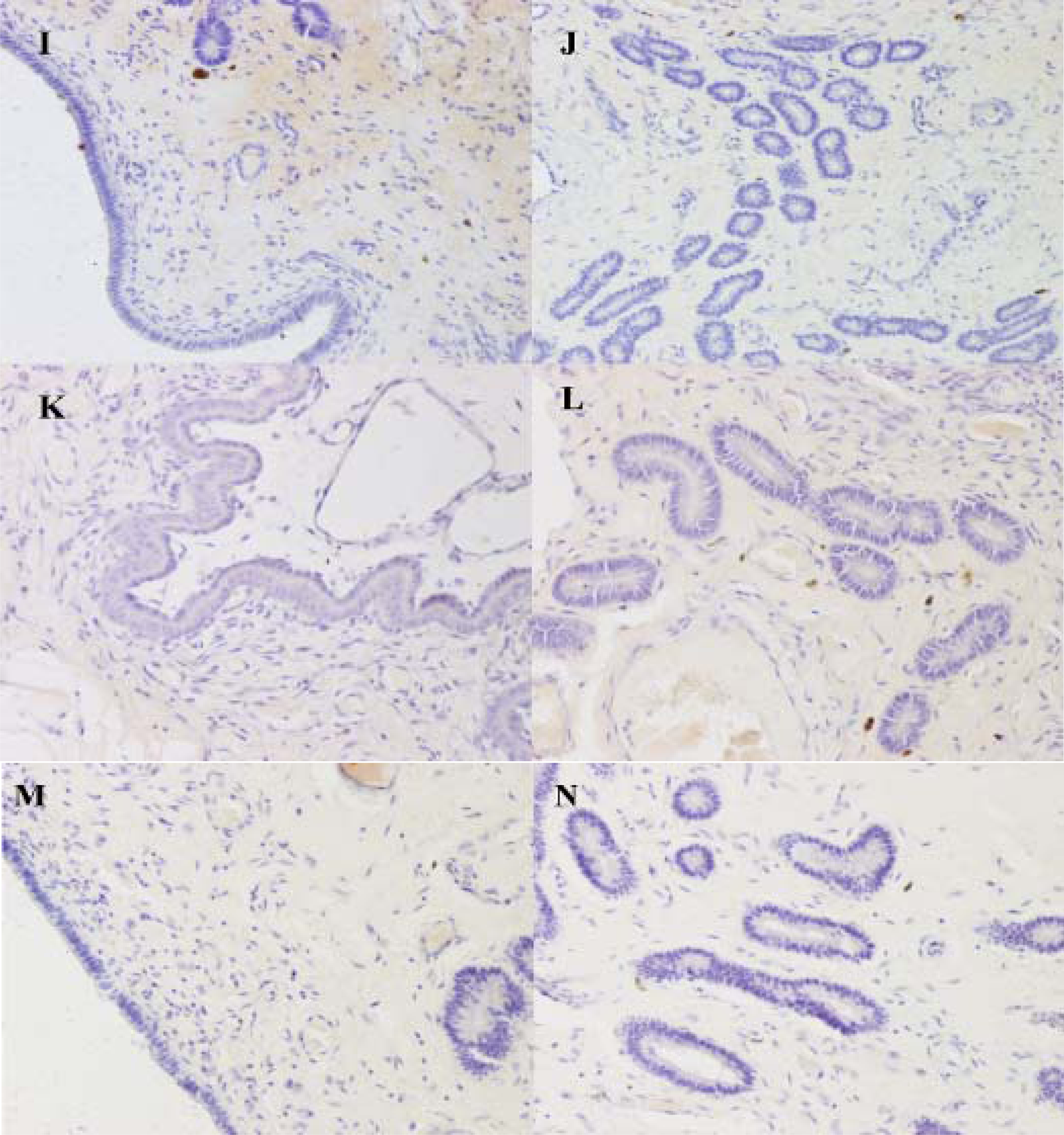
2.3. Differential Expression of Muc1 in Porcine Endometrium
3. Experimental Section
3.1. Animals and Tissue Collection
3.1.1. Animals
3.1.2. Tissue Collection
3.2. Scanning Electron Microscopy (SEM)
3.3. RNA Extraction and Reverse Transcription
3.4. Primer Design
3.5. Semi-Quantitative Polymerase Chain Reaction (SQ-PCR)
3.6. Real-Time Polymerase Chain Reaction (RT-PCR)
3.7. Immunohistochemistry
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
References and Notes
- Kyriazakis, I; Whittemore, CT. Whittemore’s Science and Practice of Pig Production, 3rd ed; Blackwell Publishing: Oxford, UK, 2006; pp. 105–147. [Google Scholar]
- Dantzer, V. Electron microscopy of the initial stages of placentation in the pig. Anat. Embryol 1985, 172, 281–293. [Google Scholar]
- Psychoyos, A. Uterine receptivity for nidation. Ann. N.Y. Acad. Sci 1986, 476, 36–42. [Google Scholar]
- Carson, DD; Bagchi, I; Dey, SK; Enders, AC; Fazleabas, AT; Lessey, BA; Yoshinaga, K. Embryo implantation. Dev. Biol 2000, 223, 217–237. [Google Scholar]
- Sharkey, AM; Smith, SK. The endometrium as a cause of implantation failure. Best Pract. Res. Clin. Obstet. Gynaecol 2003, 17, 289–307. [Google Scholar]
- Psychoyos, A; Nikas, G. Uterine pinopodes as markers of uterine receptivity. Assist. Reprod. Rev 1994, 4, 26–32. [Google Scholar]
- Acosta, AA; Elberger, L; Borghi, M; Calamera, JC; Chemes, H; Doncel, GF; Kliman, H; Lema, B; Lustig, L; Papier, S. Endometrial dating and determination of the window of implantation in healthy fertile women. Fertil. Steril 2000, 73, 788–798. [Google Scholar]
- Aplin, JD. MUC-1 glycosylation in endometrium: possible roles of the apical glycocalyx at implantation. Hum. Reprod 1999, 14(Suppl 2), 17–25. [Google Scholar]
- Giudice, LC. Potential biochemical markers of uterine receptivity. Hum. Reprod 1999, 14(Suppl 2), 3–16. [Google Scholar]
- Horne, AW; White, JO; Lalani, EN. The endometrium and embryo implantation. A receptive endometrium depends on more than hormonal influences. BRIT. MED. J 2000, 321, 1301–1302. [Google Scholar]
- Carson, DD; DeSouza, MM; Kardon, R; Zhou, X; Lagow, E; Julian, J. Mucin expression and function in the female reproductive tract. Hum. Reprod. Update 1998, 4, 459–464. [Google Scholar]
- Carson, DD; DeSouza, MM; Regisford, EG. Mucin and proteoglycan functions in embryo implantation. Bioessays 1998, 20, 577–583. [Google Scholar]
- Gendler, SJ; Spicer, AP. Epithelial mucin genes. Annu. Rev. Physiol 1995, 57, 607–634. [Google Scholar]
- Hanisch, FG; Muller, S. MUC1: The polymorphic appearance of a human mucin. Glycobiology 2000, 10, 439–449. [Google Scholar]
- Lagow, E; DeSouza, MM; Carson, DD. Mammalian reproductive tract mucins. Hum. Reprod. Update 1999, 5, 280–292. [Google Scholar]
- Simon, C; Valbuena, D. Embryonic implantation. Ann. Endocrinol. (Paris) 1999, 60, 134–136. [Google Scholar]
- Thathiah, A; Carson, DD. Mucins and blastocyst attachment. Rev. Endocr. Metab. Disord 2002, 3, 87–96. [Google Scholar]
- Braga, VM; Gendler, SJ. Modulation of Muc-1 mucin expression in the mouse uterus during the estrus cycle, early pregnancy and placentation. J. Cell. Sci 1993, 105 Pt 2, 397–405. [Google Scholar]
- Chervenak, JL; Illsley, NP. Episialin acts as an antiadhesive factor in an in vitro model of human endometrial-blastocyst attachment. Biol. Reprod 2000, 63, 294–300. [Google Scholar]
- DeSouza, MM; Surveyor, GA; Price, RE; Julian, J; Kardon, R; Zhou, X; Gendler, S; Hilkens, J; Carson, DD. MUC1/episialin: A critical barrier in the female reproductive tract. J. Reprod. Immunol 1999, 45, 127–158. [Google Scholar]
- Surveyor, GA; Gendler, SJ; Pemberton, L; Das, SK; Chakraborty, I; Julian, J; Pimental, RA; Wegner, CC; Dey, SK; Carson, DD. Expression and steroid hormonal control of Muc-1 in the mouse uterus. Endocrinology 1995, 136, 3639–3647. [Google Scholar]
- Martel, D; Malet, C; Gautray, J; Psychoyos, A. The Endometrium, Hormonal Impacts; Plenum Press: New York, NY, USA, 1981; p. 15. [Google Scholar]
- Brayman, M; Thathiah, A; Carson, DD. MUC1: A multifunctional cell surface component of reproductive tissue epithelia. Reprod. Biol. Endocrinol 2004, 2, 4. [Google Scholar]
- Pemberton, L; Taylor-Papadimitriou, J; Gendler, SJ. Antibodies to the cytoplasmic domain of the MUC1 mucin show conservation throughout mammals. Biochem. Biophys. Res. Commun 1992, 185, 167–175. [Google Scholar]
- Hoffman, LH; Olson, GE; Carson, DD; Chilton, BS. Progesterone and implanting blastocysts regulate Muc1 expression in rabbit uterine epithelium. Endocrinology 1998, 139, 266–271. [Google Scholar]
- Bowen, JA; Bazer, FW; Burghardt, RC. Spatial and temporal analyses of integrin and Muc-1 expression in porcine uterine epithelium and trophectoderm in vivo. Biol. Reprod 1996, 55, 1098–1106. [Google Scholar]
- Meseguer, M; Aplin, JD; Caballero-Campo, P; O’Connor, JE; Martin, JC; Remohi, J; Pellicer, A; Simon, C. Human endometrial mucin MUC1 is up-regulated by progesterone and down-regulated in vitro by the human blastocyst. Biol. Reprod 2001, 64, 590–601. [Google Scholar]
- Burghardt, RC; Johnson, GA; Jaeger, LA; Ka, H; Garlow, JE; Spencer, TE; Bazer, FW. Integrins and extracellular matrix proteins at the maternal-fetal interface in domestic animals. Cells Tissues Organs 2002, 172, 202–217. [Google Scholar]
- Lin, HC; Wang, X; Liu, GF; Fu, JL; Wang, AG. Expression of alphaV and beta3 integrin subunits during implantation in pig. Mol. Reprod. Dev 2007, 74, 1379–1385. [Google Scholar]
- Johnson, GA; Burghardt, RC; Joyce, MM; Spencer, TE; Bazer, FW; Pfarrer, C; Gray, CA. Osteopontin expression in uterine stroma indicates a decidualization-like differentiation during ovine pregnancy. Biol. Reprod 2003, 68, 1951–1958. [Google Scholar]
- Cunha, GR; Bigsby, RM; Cooke, PS; Sugimura, Y. Stromal-epithelial interactions in adult organs. Cell Differ 1985, 17, 137–148. [Google Scholar]
- Cunha, GR; Chung, LW; Shannon, JM; Taguchi, O; Fujii, H. Hormone-induced morphogenesis and growth: role of mesenchymal-epithelial interactions. Recent Prog. Horm. Res 1983, 39, 559–598. [Google Scholar]
- Guillomot, M; Flechon, JE; Wintenberger-Torres, S. Conceptus attachment in the ewe: An ultrastructural study. Placenta 1981, 2, 169–182. [Google Scholar]
- Lord, E; Murphy, BD; Desmarais, JA; Ledoux, S; Beaudry, D; Palin, MF. Modulation of peroxisome proliferator-activated receptor delta and gamma transcripts in swine endometrial tissue during early gestation. Reproduction 2006, 131, 929–942. [Google Scholar]
- Abd-Elnaeim, MM; Saber, A; Hassan, A; Abou-Elmagd, A; Klisch, K; Jones, CJ; Leiser, R. Development of the areola in the early placenta of the one-humped camel (Camelus dromedarius): A light, scanning and transmission electron microscopical study. Anat. Histol. Embryol 2003, 32, 326–334. [Google Scholar]
- Livak, KJ; Schmittgen, TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar]
- Schmittgen, TD; Livak, KJ. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc 2008, 3, 1101–1108. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day 13 | Day 18 | Day 24 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| LE | GE | S | LE | GE | S | LE | GE | S | |
| intersite | ++ | ± | ++ | +++ | + | + | − | − | − |
| At site | + | − | + | ++ | ++ | ± | − | − | − |
| Primer Name | Primer Sequences (5′–3′) | Annealing Temperature (°C) | Product Size (bp) | Genbank Accession no./references |
|---|---|---|---|---|
| SQ-PCR | ||||
| Muc1-1 | Forward:CACCACCAGCTACTACAAGG | 62 | 553 | AY243508 |
| Reverse:TGCCAGGTTCGAGTAAGAG | ||||
| GAPDH-1 | Forward:ACCACAGTCCATGCCATCAC | 60 | 452 | AF017079/BDBiosciences |
| Reverse:TCCACCACCCTGTTGCTGTA | ||||
| RT-PCR | ||||
| Muc1-2 | Forward:GTGCCGACGAAAGAACTG | 60 | 187 | AY243508 |
| Reverse:TGCCAGGTTCGAGTAAGAG | ||||
| GAPDH-2 | Forward:GTCCACTGGTGTCTTCACGA | 60 | 154 | AF141959/[29] |
| Reverse:GCTGACGATCTTGAGGGAGT |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ren, Q.; Guan, S.; Fu, J.; Wang, A. Temporal and Spatial Expression of Muc1 During Implantation in Sows. Int. J. Mol. Sci. 2010, 11, 2322-2335. https://doi.org/10.3390/ijms11062322
Ren Q, Guan S, Fu J, Wang A. Temporal and Spatial Expression of Muc1 During Implantation in Sows. International Journal of Molecular Sciences. 2010; 11(6):2322-2335. https://doi.org/10.3390/ijms11062322
Chicago/Turabian StyleRen, Qian, Shu Guan, Jinluan Fu, and Aiguo Wang. 2010. "Temporal and Spatial Expression of Muc1 During Implantation in Sows" International Journal of Molecular Sciences 11, no. 6: 2322-2335. https://doi.org/10.3390/ijms11062322